ARF1 and SAR1 GTPases in Endomembrane Trafficking in Plants

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
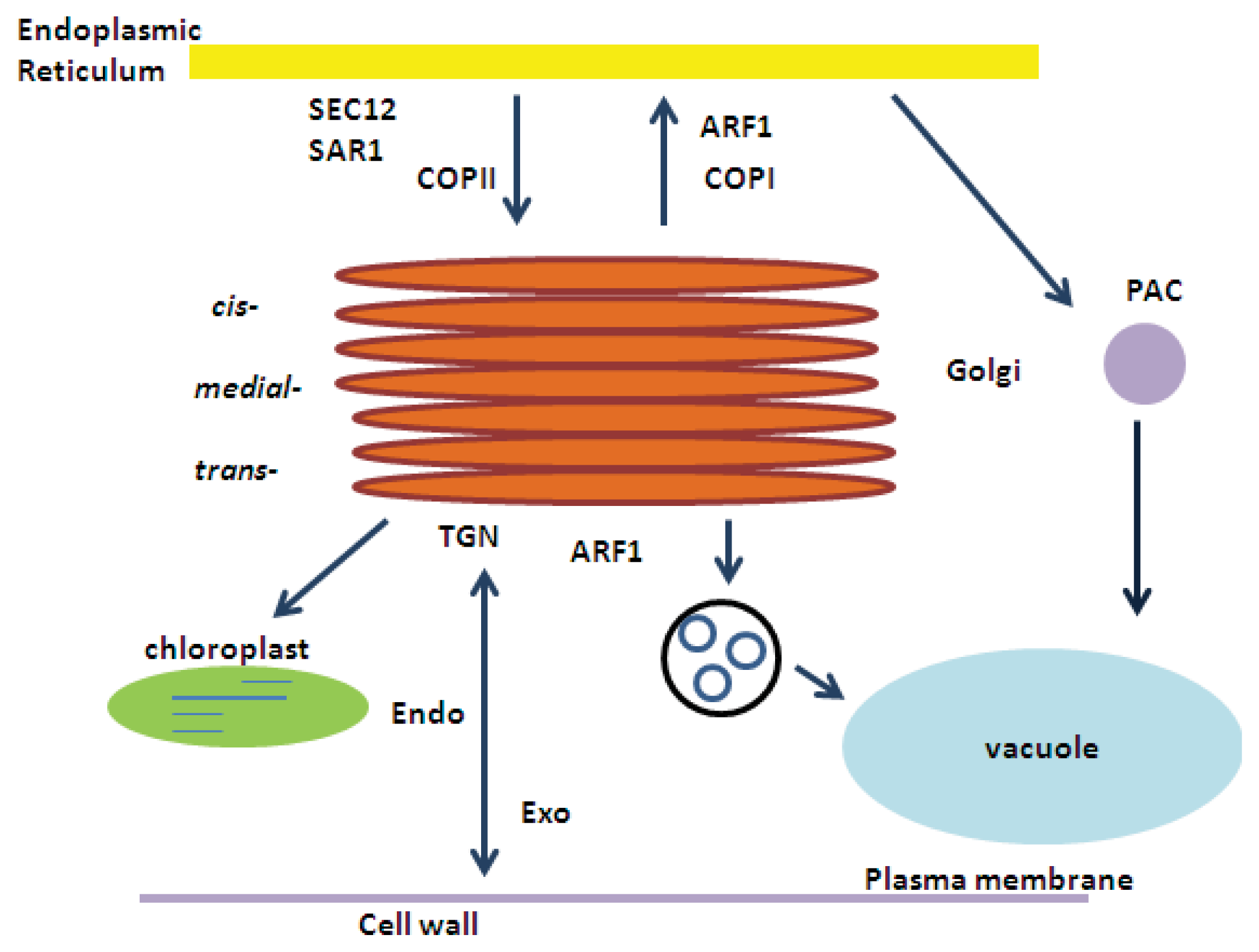
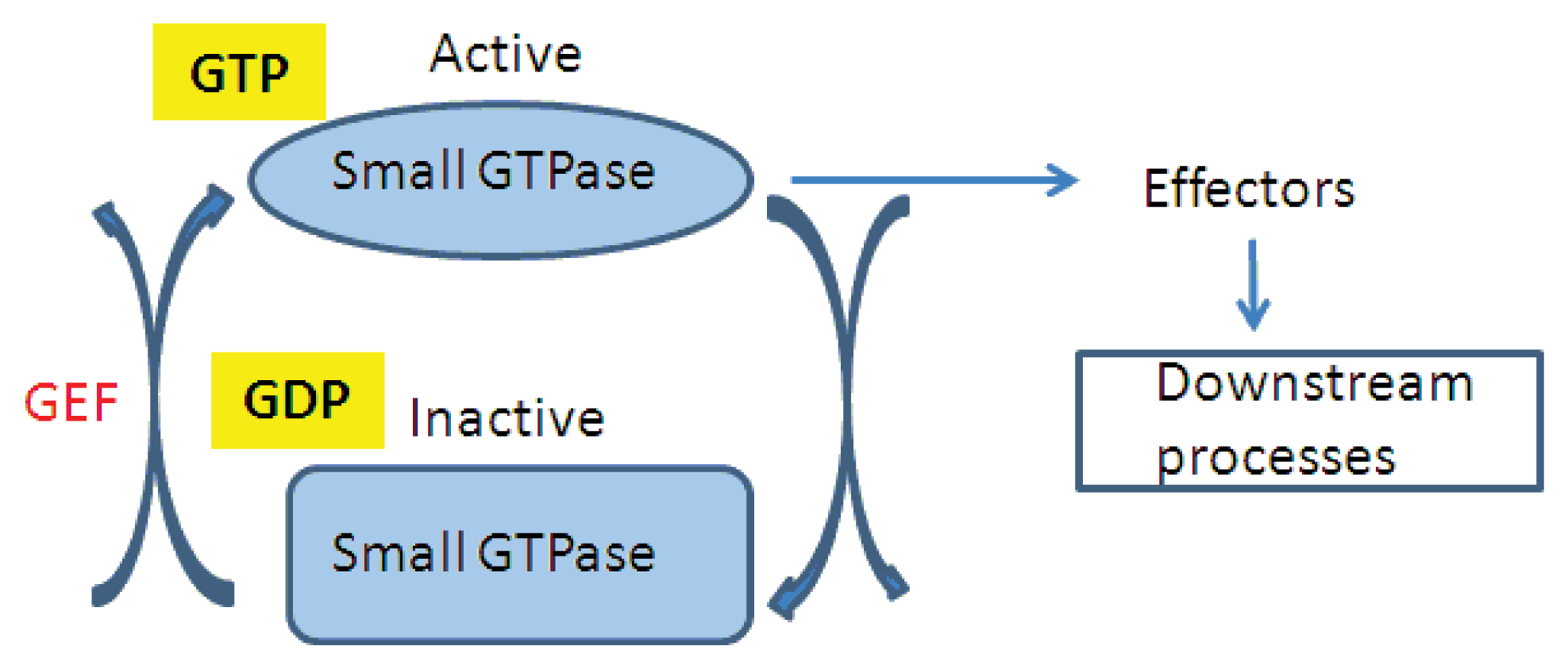
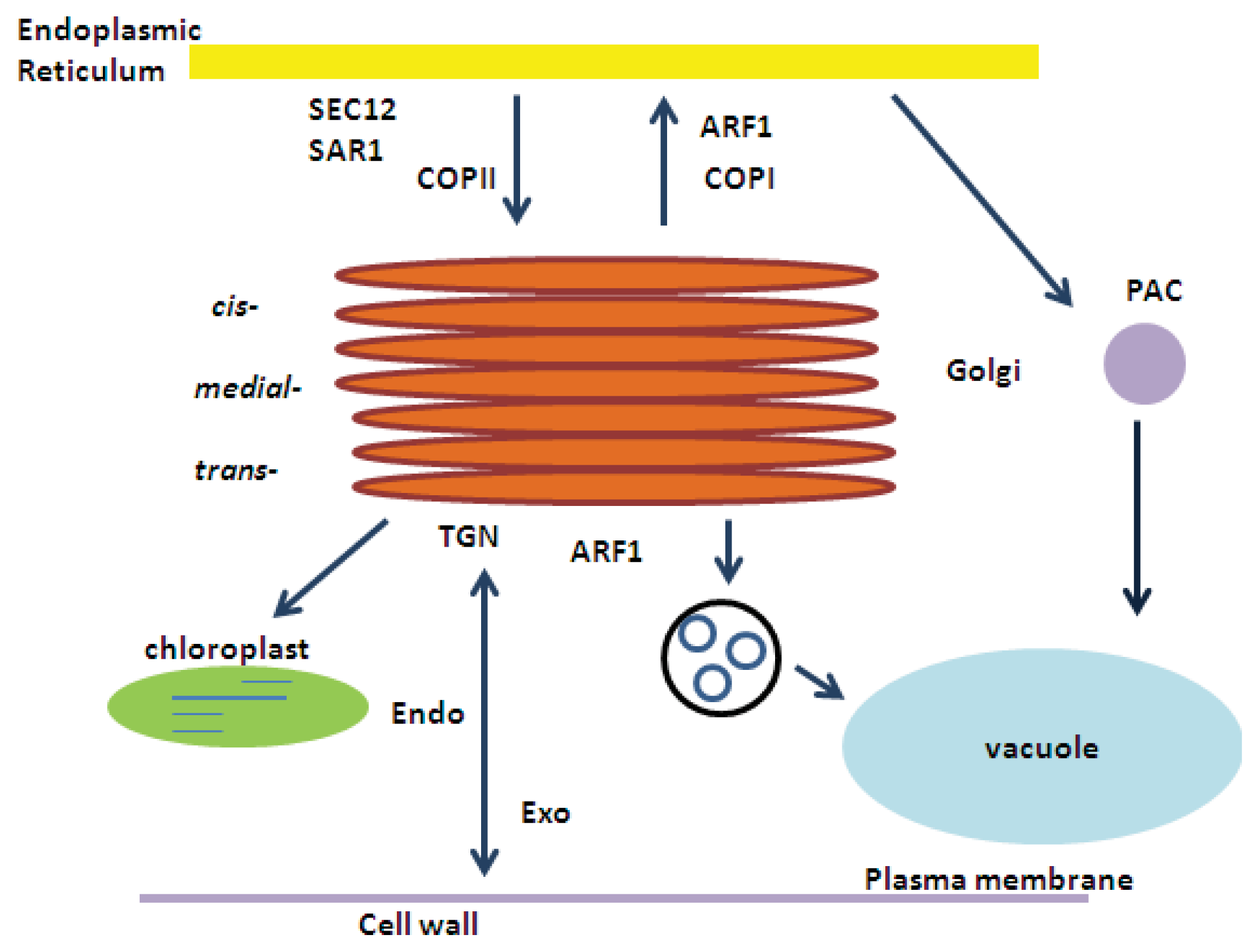
2. ARF1 (ADP-Ribosylation Factor 1) and SAR1 (Secretion-Associated RAS Super Family 1) GTPases
2.1. ARF1
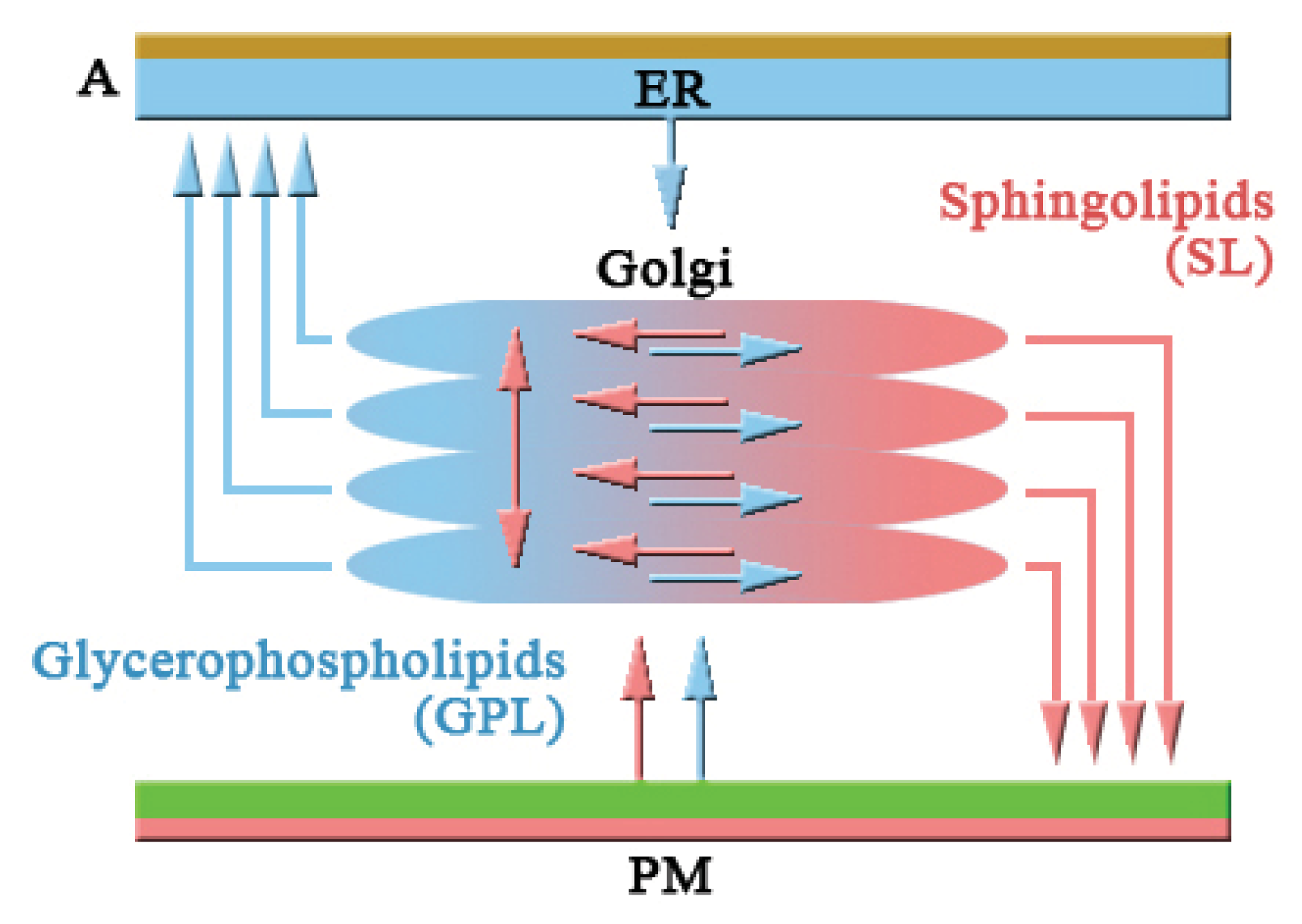
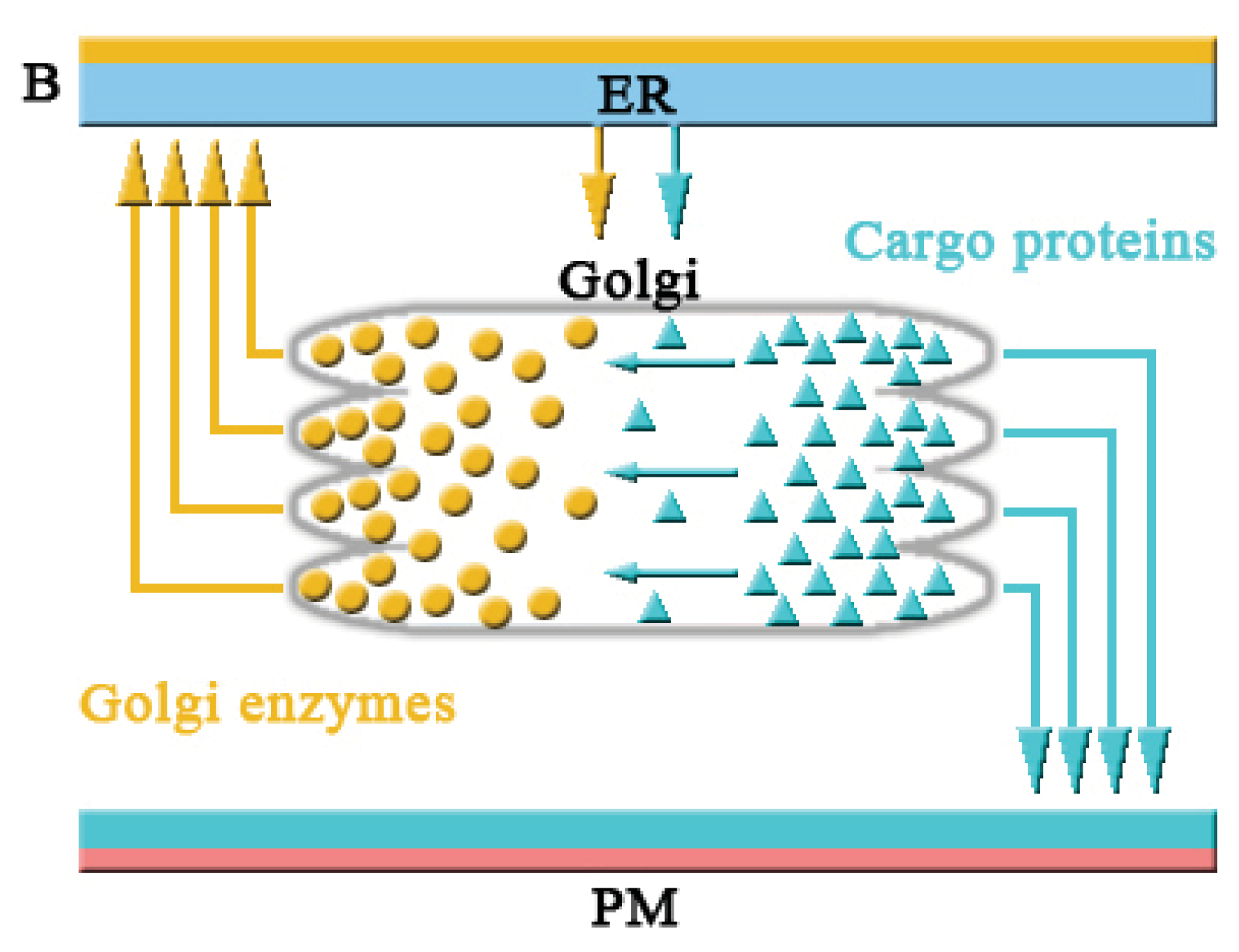
2.1.1. COPI and Retrograde Transport
2.1.2. Intra-Golgi Transport
2.2. SAR1 and COPII
ER-to-Golgi Protein Transport
3. Developmental Regulation of ARF1 and SAR1
4. Light Regulation of ARF1 and SAR1
5. Conclusions and Perspectives
Acknowledgments
Conflicts of Interest
References
- Kahn, R.A.; der, C.J.; Bokoch, G.M. The ras superfamily of GTP-binding proteins: Guidelines on nomenclature. FASEB J 1992, 6, 2512–2513. [Google Scholar]
- Vernoud, V.; Horton, A.C.; Yang, Z.; Nielsen, E. Analysis of the small GTPase gene superfamily of Arabidopsis. Plant Physiol 2003, 131, 1191–1208. [Google Scholar]
- Inoue, H.; Randazzo, P.A. Arf GAPs and their interacting proteins. Traffic 2007, 8, 1465–1475. [Google Scholar]
- Bourne, H.R.; Sanders, D.A.; McCormick, F. The GTPase superfamily: A conserved switch for diverse cell functions. Nature 1990, 348, 125–132. [Google Scholar]
- Jekely, G. Small GTPases and the evolution of the eukaryotic cell. BioEssays 2003, 25, 1129–1138. [Google Scholar]
- Nielsen, E.; Cheung, A.Y.; Ueda, T. The regulatory RAB and ARF GTPases for vesicular trafficking. Plant Physiol 2008, 147, 1516–1526. [Google Scholar]
- Goldberg, J. Structural and functional analysis of the ARF1-ARFGAP complex reveals a role for coatamer in GTP hydrolysis. Cell 1999, 96, 893–902. [Google Scholar]
- D’Souza-Schorey, C.; Chavrier, P. ARF proteins: Roles in membrane traffic and beyond. Nat. Rev. Mol. Cell Biol 2006, 7, 347–358. [Google Scholar]
- Morishige, M.; Hashimoto, S.; Ogawa, E.; Toda, Y.; Kotani, H.; Hirose, M.; Wei, S.; Hashimoto, A.; Yamada, A.; Yano, H.; et al. GEP100 links epidermal growth factor receptor signalling to Arf6 activation to induce breast cancer invasion. Nat. Cell Biol 2008, 10, 85–92. [Google Scholar]
- Muralidharan-Chari, V.; Hoover, H.; Clancy, J.; Schweitzer, J.; Suckow, M.A.; Schroeder, V.; Castellino, F.J.; Schorey, J.S.; D’Souza-Schorey, C. ADP-ribosylation factor 6 regulates tumorigenic and invasive properties in vivo. Cancer Res 2009, 69, 2201–2209. [Google Scholar]
- Tsai, M.M.; Lin, P.Y.; Cheng, W.L.; Tsai, C.Y.; Chi, H.C.; Chen, C.Y.; Tseng, Y.H.; Cheng, Y.F.; Chen, C.D.; Liang, Y.; et al. Overexpression of ADP-ribosylation factor 1 in human gastric carcinoma and its clinicopathological. Cancer Sci 2012, 103, 1136–1144. [Google Scholar]
- Donaldson, J.G.; Jackson, C.L. ARF family G proteins and their regulators: Roles in membrane transport, development and disease. Nat. Rev. Mol. Cell Biol 2011, 12, 362–375. [Google Scholar]
- Sanderfoot, A.; Raikhel, N. The Secretory System of Arabidopsis. In The Arabidopsis Book; American Society of Plant Biologists: Rockville, MD, USA, 2003. [Google Scholar]
- Donohoe, B.S.; Kang, B.H.; Staehelin, L.A. Identification and characterization of COPIa- and COPIb-type vesicle classes associated with plant and algal Golgi. Proc. Natl. Acad. Sci. USA 2007, 104, 163–168. [Google Scholar]
- Ostermann, J.; Orci, L.; Tani, K.; Amherdt, M.; Ravazzola, M.; Elazar, Z.; Rothman, J.E. Stepwise assembly of functionally active transport vesicles. Cell 1993, 75, 1015–1025. [Google Scholar]
- Beck, R.; Prinz, S.; Diestelkotter-Bachert, P.; Rohling, S.; Adolf, F.; Hoehner, K.; Welsch, S.; Ronchi, P.; Brügger, B.; Briggs, J.A.; et al. Coatomer and dimeric ADP ribosylation factor 1 promote distinct steps in membrane scission. J. Cell Biol 2011, 194, 765–777. [Google Scholar]
- Orci, L.; Amherdt, M.; Ravazzola, M.; Perrelet, A.; Rothman, J.E. Exclusion of Golgi residents from transport vesicles budding from Golgi cisternae in intact cells. J. Cell Biol 2000, 150, 1263–1269. [Google Scholar]
- Martínez-Menárguez, J.A.; Prekeris, R.; Oorschot, V.M.J.; Scheller, R.; Slot, J.W.; Geuze, H.J.; Klumperman, J. Peri-Golgi vesicles contain retrograde but not anterograde proteins consistent with the cisternal progression model of intra-Golgi transport. J. Cell Biol 2001, 155, 1213–1224. [Google Scholar]
- Whitney, J.A.; Gomez, M.; Sheff, D.; Kreis, T.E.; Mellman, I. Cytoplasmic coat proteins involved in endosome function. Cell 1995, 83, 703–713. [Google Scholar]
- Gu, F.; Aniento, F.; Parton, R.G.; Gruenberg, J. Functional dissection of COP-I subunits in the biogenesis of multivesicular endosomes. J. Cell Biol. 1997, 139, 1183–1195. [Google Scholar]
- Aniento, F.; Helms, B.; Memon, A. How to make a vesicle: Coat protein-membrane interactions. The Golgi apparatus and the Plant Secretory Pathway. Annu. Plant Rev 2003, 9, 36–62. [Google Scholar]
- Jackson, M.R.; Nilsson, T.; Peterson, P.A. Identification of a consensus motif for retention of transmembrane proteins in the endoplasmic reticulum. EMBO J. 1990, 9, 3153–3162. [Google Scholar]
- Hardt, B.; Bause, E. Lysine can be replaced by histidine but not by arginine as the ER retrieval motif for type I membrane proteins. Biochem. Biophys. Res. Commun 2002, 291, 751–757. [Google Scholar]
- Gaynor, E.C.; Heesen, S.; Graham, T.R.; Aebi, M.; Emr, S.D. Signal-mediated retrieval of a membrane protein from the Golgi to the ER in yeast. J. Cell Biol 1994, 127, 653–665. [Google Scholar]
- Sohn, K.; Orci, L.; Ravazzola, M.; Amherdt, M.; Brunner, M.; Kahn, R.A.; Rothman, J.E. A major transmembrane protein of Golgi-derived COPIcoated vesicles involved in coatomer binding. J. Cell Biol 1996, 135, 1239–1248. [Google Scholar]
- Fiedler, K.; Veit, M.; Stamnes, M.A.; Rothman, J.E. Bimodal interaction of coatomer with the p24 family of putative cargo receptors. Science 1996, 273, 1396–1399. [Google Scholar]
- Goldberg, J. Decoding of sorting signals by coatomer through a GTPase switch in the COPI coat complex. Cell 2000, 100, 671–679. [Google Scholar]
- Nickel, W.; Brugger, B.; Wieland, F. Vesicular transport: The core machinery of COPI recruitment and budding. J. Cell Sci 2002, 115, 3235–3240. [Google Scholar]
- Langhans, M.; Marcote, M.J.; Pimpl, P.; Virgili-López, G.; Robinson, D.G.; Aniento, F. In vivo trafficking and localization of p24 proteins in plant cells. Traffic 2008, 9, 770–785. [Google Scholar]
- Chardin, P.; Paris, S.; Antonny, B.; Robineau, S.; Béraud-Dufour, S.; Jackson, C.L.; Chabre, M. A human exchange factor for ARF contains Sec7- and pleckstrin-homology domains. Nature 1996, 384, 481–484. [Google Scholar]
- Zhao, L.; Helms, J.B.; Brugger, B.; Harter, C.; Martoglio, B.; Graf, R.; Brunner, J.; Wieland, F.T. Direct and GTP-dependent interaction of ADP ribosylation factor 1 with coatomer subunit beta. Proc. Natl. Acad. Sci. USA 1997, 94, 4418–4423. [Google Scholar]
- Donaldson, J.G.; Cassel, D.; Kahn, R.A.; Klaussner, R.D. ADP-ribosylation factor, a small GTP-binding protein, is required for binding of the coatomer protein 3-COP to Golgi membranes. Proc. Natl. Acad. Sci. USA 1992, 89, 6408–6412. [Google Scholar]
- Antonny, B.; Beraud-Dufour, S.; Chardin, P.; Chabre, M. N-terminal hydrophobic residues of the G-protein ADP-ribosylation factor-1 insert into membrane phospholipids upon GDP to GTP exchange. Biochemistry 1997, 36, 4675–4684. [Google Scholar]
- Itoh, T.; de Camilli, P. Membrane trafficking: Dual-key strategy. Nature 2004, 429, 141–143. [Google Scholar]
- Majoul, I.; Straub, M.; Hell, S.W.; Duden, R.; Soling, H.D. KDEL-cargo regulates interactions between proteins involved in COPI vesicle traffic: Measurements in living cells using FRET. Dev. Cell 2001, 1, 139–153. [Google Scholar]
- Contreras, I.; Ortiz-Zapater, E.; Aniento, F. Sorting signals in the cytosolic tail of membrane proteins involved in the interaction with plant ARF1 and coatomer. Plant J 2004, 38, 685–698. [Google Scholar]
- Rojo, M.; Peperkok, R.; Emery, G.; Kellner, R.; Stang, E.; Parton, R.G.; Gruenberg, J. Involvement of the transmembrane protein p23 in biosynthetic protein transport. J. Cell Biol 1997, 139, 1119–1135. [Google Scholar]
- Lavoie, C.; Paiement, J.; Dominguez, M.; Roy, L.; Dahan, S.; Gushue, J.N.; Bergeron, J.J. Roles for alpha (2) p24 and COPI in endoplasmic reticulum cargo exit site formation. J. Cell Biol 1999, 146, 285–300. [Google Scholar]
- Emery, G.; Parton, R.G.; Rojo, M.; Gruenberg, J. The trans-membrane protein p25 forms highly specialized domains that regulate membrane composition and dynamics. J. Cell Sci 2003, 116, 4821–4832. [Google Scholar]
- Contreras, I.; Yang, Y.; Robinson, D.G.; Aniento, F. Plant COPI and COPII coat proteins show a differential affinity for p24 cytosolic tails. Plant Cell Physiol 2004, 45, 1779–1786. [Google Scholar]
- Stearns, T.; Kahn, R.A.; Botstein, D.; Hoyt, M.A. ADP ribosylation factor is an essential protein in Saccharomyces cerevisiae and is encoded by two genes. Mol. Cell. Biol 1990, 10, 6690–6699. [Google Scholar]
- Xu, J.; Scheres, B. Cell polarity: ROPing the ends together. Curr. Opin. Plant Biol 2005, 8, 613–618. [Google Scholar]
- Keskin, B.C.; Yuca, E.; Ertekin, O.; Yüksel, B.; Memon, A.R. Expression characteristics of ARF1 and SAR1 during development and the de-etiolation process. Plant Biol 2012, 14, 24–32. [Google Scholar]
- Gebbie, L.K.; Burn, J.E.; Hocart, C.H.; Williamson, R.E. Genes encoding ADP-ribosylation factors in Arabidopsis thaliana L. Heyn., genome analysis and antisense suppression. J. Exp. Bot 2005, 56, 1079–1091. [Google Scholar]
- Regad, F.; Bardet, C.; Tremousaygue, D.; Moisan, A.; Lescure, B.; Axelos, M. cDNA cloning and expression of an Arabidopsis GTP-binding protein of the ARF family. FEBS Lett 1993, 316, 133–136. [Google Scholar]
- Kobayashi-Uehara, A.; Shimosaka, E.; Handa, H. Cloning and expression analyses of cDNA encoding an ADP-ribosylation factor from wheat: Tissue-specific expression of wheat ARF. Plant Sci 2001, 160, 535–542. [Google Scholar]
- Rothman, J.E.; Wieland, F.T. Protein sorting by transport vesicles. Science 1996, 272, 227–234. [Google Scholar]
- Pepperkok, R.; Whitney, J.A.; Gomez, M.; Kreis, T.E. COPI vesicles accumulating in the presence of a GTP restricted arf1 mutant are depleted of anterograde and retrograde cargo. J. Cell Sci 2000, 113, 135–144. [Google Scholar]
- Lanoix, J.; Ouwendijk, J.; Lin, C.C.; Stark, A.; Love, H.D.; Ostermann, J.; Nilsson, T. GTP hydrolysis by arf-1 mediates sorting & concentration of Golgi resident enzymes into functional COP I vesicles. EMBO J. 1999, 18, 4935–4948. [Google Scholar]
- Glick, B.S.; Elston, T.; Oster, G.A. Cisternal maturation mechanism can explain the asymmetry of the Golgi stack. FEBS Lett 1997, 414, 177–181. [Google Scholar]
- Trucco, A.; Polishchuk, R.S.; Martella, O.; di Pentima, A.; Fusella, A.; di Gian-domenico, D.; San Pietro, E.; Beznoussenko, G.V.; Polishchuk, E.V.; Baldas-Sarre, M.; et al. Secretory traffic triggers the formation of tubular continuities across Golgi sub-compartments. Nat. Cell Biol 2004, 6, 1071–1081. [Google Scholar]
- Matsuura-Tokita, K.; Takeuchi, M.; Ichihara, A.; Mikuriya, K.; Nakano, A. Live imaging of yeast Golgi cisternal maturation. Nature 2006, 441, 1007–1010. [Google Scholar]
- Patterson, G.H.; Hirschberg, K.; Polishchuk, R.S.; Gerlich, D.; Phair, R.D.; Lippincott-Schwartz, J. Transport through the Golgi apparatus by rapid partitioning within a two-phase membrane system. Cell 2008, 133, 1055–1067. [Google Scholar]
- Smith, R.D.; Lupashin, V.V. Role of the conserved oligomeric Golgi (COG) complex in protein glycosylation. Carbohydr. Res 2008, 343, 2024–2031. [Google Scholar]
- Nakano, A.; Muramatsu, M. A novel GTP-binding protein, Sar1p, is involved in transport from the endoplasmic reticulum to the Golgi apparatus. J. Cell Biol 1989, 109, 2677–2691. [Google Scholar]
- Cutrona, M.B.; Beznoussenko, G.V.; Fusella, A.; Martella, O.; Moral, P.; Mironov, A.A. Silencing of mammalian Sar1 isoforms reveals COPII-independent protein sorting and transport. Traffic 2013, 14, 691–708. [Google Scholar]
- Zanetti, G.; Pahuja, K.B.; Studer, S.; Shim, S.; Schekman, R. COPII and the regulation of protein sorting in mammals. Nat. Cell Biol 2011, 14, 20–28. [Google Scholar]
- Villarejo, A.; Burén, S.; Larsson, S.; Déjardin, A.; Monné, M.; Rudhe, C.; Karlsson, J.; Jansson, S.; Lerouge, P.; Rolland, N.; et al. Evidence for a protein transported through the secretory pathway en route to the higher plant chloroplast. Nat. Cell Biol 2005, 7, 1224–1231. [Google Scholar]
- Melville, D.B.; Knapik, E.W. Traffic jams in fish bones. ER-to-Golgi protein transport during zebrafish development. Cell Adhes. Migr 2011, 5, 114–118. [Google Scholar]
- Orci, L.; Stamnes, M.; Ravazzola, M.; Amherdt, M.; Perrelet, A.; Söllner, T.H.; Rothman, J.E. Bidirectional transport by distinct populations of COPI-coated vesicles. Cell 1997, 90, 335–349. [Google Scholar]
- Stephens, D.J.; Pepperkok, R. Illuminating the secretory pathway: When do we need vesicles? J. Cell Sci 2001, 114, 1053–1059. [Google Scholar]
- D’Enfert, C.; Wuestehube, L.J.; Lila, T.; Schekman, R. Sec12p-dependent membrane binding of the small GTP-binding protein Sar1p promotes formation of transport vesicles from the ER. J. Cell Biol 1991, 114, 663–670. [Google Scholar]
- Miller, E.A.; Barlowe, C. Regulation of coat assembly-sorting things out at the ER. Curr. Opin. Cell Biol 2010, 22, 447–453. [Google Scholar]
- Miller, E.A.; Antonny, B.; Hamamoto, S.; Schekman, R. Cargo selection into COPII vesicles is driven by the Sec24p subunit. EMBO J 2002, 21, 6105–6113. [Google Scholar]
- Miller, E.A.; Beilharz, T.H.; Malkus, P.N.; Lee, M.C.S.; Hamamoto, S.; Orci, L.; Schekman, R. Multiple cargo binding sites on the COPII subunit Sec24p ensure capture of diverse membrane proteins into transport vesicles. Cell 2003, 114, 497–509. [Google Scholar]
- Hughes, H.; Budnik, A.; Schmidt, K.; Palmer, K.J.; Mantell, J.; Noakes, C.; Johnson, A.; Carter, D.A.; Verkade, P.; Watson, P.; et al. Organisation of human ER-exit sites: Requirements for the localisation of Sec16 to transitional ER. J. Cell Sci 2009, 122, 2924–2934. [Google Scholar]
- Karhinen, L.; Bastos, R.N.; Jokitalo, E.; Makarow, M. Endoplasmic reticulum exit of a secretory glycoprotein in the absence of Sec24p family proteins in yeast. Traffic 2005, 6, 562–574. [Google Scholar]
- Lee, M.C.S.; Orci, L.; Hamamoto, S.; Futai, E.; Ravazzola, M.; Schekman, R. Sar1p N-terminal helix initiates membrane curvature and completes the fission of a COPII vesicle. Cell 2005, 122, 605–617. [Google Scholar]
- Settles, E.I.; Loftus, A.F.; McKeown, A.N.; Parthasarathy, R. The vesicle trafficking protein Sar1 lowers lipid membrane rigidity. Biophys. J 2010, 99, 1539–1545. [Google Scholar]
- Loftus, A.F.; Hsieh, V.L.; Parthasarathy, R. Modulation of membrane rigidity by the human vesicle trafficking proteins Sar1A and Sar1B. Biochem. Biophys. Res. Commun 2012, 426, 585–589. [Google Scholar]
- Phillipson, B.A.; Pimpl, P.; daSilva, L.L.; Crofts, A.J.; Taylor, J.P.; Movafeghi, A.; Robinson, D.G.; Denecke, J. Secretory bulk flow of soluble proteins is efficient and COPII dependent. Plant Cell 2001, 13, 2005–2020. [Google Scholar]
- DaSilva, L.L.; Snapp, E.L.; Denecke, J.; Lippincott-Schwartz, J.; Hawes, C.; Brandizzi, F. Endoplasmic reticulum export sites and Golgi bodies behave as single mobile secretory units in plant cells. Plant Cell 2004, 16, 1753–1771. [Google Scholar]
- Ward, T.H.; Brandizzi, F. Dynamics of proteins in Golgi membranes: Comparisons between mammalian and plant cells highlighted by photobleaching techniques. Cell. Mol. Life Sci 2004, 61, 172–185. [Google Scholar]
- Kang, B.H.; Staehelin, L.A. ER-to-Golgi transport by COPII vesicles in Arabidopsis involves a ribosome-excluding scaffold that is transferred with the vesicles to the Golgi matrix. Protoplasma 2008, 234, 51–64. [Google Scholar]
- Kuehn, M.J.; Herrmann, J.M.; Schekman, R. COPII-cargo interactions direct protein sorting into ER-derived transport vesicles. Nature 1998, 391, 187–190. [Google Scholar]
- Supek, F.; Madden, D.T.; Hamamoto, S.; Orci, L.; Schekman, R. Sec16p potentiates the action of COPII proteins to bud transport vesicles. J. Cell Biol 2002, 158, 1029–1038. [Google Scholar]
- Gimeno, R.E.; Espenshade, P.; Kaiser, C.A. SED4 encodes a yeast endoplasmic reticulum protein that binds Sec16p and participates in vesicle formation. J. Cell Biol 1995, 131, 325–338. [Google Scholar]
- Saito-Nakano, Y.; Nakano, A. Sed4p functions as a positive regulator of Sar1p probably through inhibition of the GTPase activation by Sec23p. Genes Cells 2000, 5, 1039–1048. [Google Scholar]
- Connerly, P.L.; Esaki, M.; Montegna, E.A.; Strongin, D.E.; Levi, S.; Soderholm, J.; Glick, B.S. Sec16 is a determinant of transitional ER organization. Curr. Biol 2005, 15, 1439–1447. [Google Scholar]
- Watson, P.; Townley, A.K.; Koka, P.; Palmer, K.J.; Stephens, D.J. Sec16 defines endoplasmic reticulum exit sites and is required for secretory cargo export in mammalian cells. Traffic 2006, 7, 1678–1687. [Google Scholar]
- De Matteis, M.A.; Luini, A. Exiting theGolgi complex. Nat. Rev. Mol. Cell Biol 2008, 9, 273–284. [Google Scholar]
- Hanton, S.L.; Chatre, L.; Renna, L.; Matheson, L.A.; Brandizzi, F. De novo formation of plant endoplasmic reticulum export sites is membrane cargo induced and signal mediated. Plant Physiol 2007, 143, 1640–1650. [Google Scholar]
- Bar-Peled, M.; Conceicao, A.; Frigerio, L.; Raikhel, N.V. Expression and regulation of aERD2, a gene encoding the KDEL receptor homolog in plants, and other genes encoding proteins involved in ER-Golgi vesicular trafficking. Plant Cell 1995, 7, 667–676. [Google Scholar]
- Robinson, D.G.; Herranz, M.C.; Bubeck, J.; Pepperkok, R.; Ritzenthaler, C. Membrane dynamics in the early secretory pathway. Crit. Rev. Plant Sci 2007, 26, 199–225. [Google Scholar]
- Hanton, S.L.; Chatre, L.; Matheson, L.A.; Rossi, M.; Held, M.A.; Brandizzi, F. Plant Sar1 isoforms with near-identical protein sequences exhibit different localisations and effects on secretion. Plant Mol. Biol 2008, 67, 283–294. [Google Scholar]
- Jensen, D.; Schekman, R. COPII-mediated vesicle formation at a glance. J. Cell Sci 2011, 124, 1–4. [Google Scholar]
- Fromme, J.C.; Ravazzola, M.; Hamamoto, S.; Al-Balwi, M.; Eyaid, W.; Boyadjiev, S.A.; Cosson, P.; Schekman, R.; Orci, L. The genetic basis of a craniofacial disease provides insight into COPII coat assembly. Dev. Cell 2007, 13, 623–634. [Google Scholar]
- Sarmah, S.; Barallo-Gimeno, A.; Melville, D.B.; Topczewski, J.; Solnica-Krezel, L.; Knapik, E.W. Sec24D-dependent transport of extracellular matrix proteins is required for zebrafish skeletal morphogenesis. PLoS One 2010, 5, e10367. [Google Scholar]
- Barlowe, C. Signals for COPII-dependent export from the ER: What’s the ticket out? Trends Cell Biol 2003, 13, 295–300. [Google Scholar]
- Sieben, C.; Mikosch, M.; Brandizzi, F.; Homann, U. Interaction of the K(+)-channel KAT1 with the coat protein complex II coat component Sec24 depends on a di-acidic endoplasmic reticulum export motif. Plant J. 2008, 56, 997–1006. [Google Scholar]
- Marti, L.; Fornaciari, S.; Renna, L.; Stefano, G.; Brandizzi, F. COPII-mediated traffic in plants. Trends Plant Sci 2010, 15, 522–528. [Google Scholar]
- Jürgens, G. Membrane trafficking in plants. Annu. Rev. Cell Dev. Biol 2004, 20, 481–504. [Google Scholar]
- Mazel, A.; Leshem, Y.; Tiwari, B.S.; Levine, A. Induction of salt and osmotic stress tolerance by overexpression of an intracellular vesicle trafficking protein AtRab7 (AtRabG3e). Plant Physiol 2004, 134, 118–128. [Google Scholar]
- Jung, Y.H.; Agrawal, G.K.; Rakwal, R.; Kim, J.A.; Lee, M.O.; Choi, P.G.; Kim, Y.J.; Kim, M.J.; Shibato, J.; Kim, S.H.; et al. Functional characterization of OsRacB GTPase—A potentially negative regulator of basal disease resistance in rice. Plant Physiol. Biochem 2006, 44, 68–77. [Google Scholar]
- Campanoni, P.; Blatt, M.R. Membrane trafficking and polar root growth in root hairs and pollen tubes. J. Exp. Bot 2007, 58, 65–74. [Google Scholar]
- Galway, M.E.; Heckman, J.W.; Schiefelbein, J.W. Growth and ultrastructure of Arabidopsis root hairs: The rhd3 mutation alters vacuole enlargement and tip growth. Planta 1997, 201, 209–218. [Google Scholar]
- Voigt, B.; Timmers, T.; Samaj, J.; Muller, J.; Baluska, F.; Menzel, D. GFP-FABD2 fusion construct allows in vivo visualization of the dynamic actin cytoskeleton in all cells of Arabidopsis seedlings. Eur. J. Cell Biol 2005, 84, 595–608. [Google Scholar]
- Clouse, S.D. Integration of light and brassinosteroid signals in etiolated seedling growth. Trends Plant Sci 2001, 6, 443–445. [Google Scholar]
- Nagano, Y.; Okada, Y.; Narita, H.; Asaka, Y.; Sasaki, Y. Location of light-repressible, small GTP-binding protein of the YPT/rab family in the growing zone of etiolated pea stems. Proc. Natl. Acad. Sci. USA 1995, 92, 6314–6318. [Google Scholar]
- Inaba, T.; Nagano, Y.; Sakakibara, T.; Sasaki, Y. Identification of a cis-regulatory element involved in phytochrome down-regulated expression of the pea small GTPase gene pra2. Plant Physiol 1999, 120, 491–499. [Google Scholar]
- Inaba, T.; Nagano, Y.; Reid, J.B.; Sasaki, Y. DE1 a 12-base pair cis-regulatory element sufficient to confer dark-inducible and light down-regulated expression to a minimal promoter in pea. J. Biol. Chem 2000, 275, 19723–19727. [Google Scholar]
- Neff, J.C.; Hobbie, S.H.; Vitousek, P.M. Controls over the production and stoichiometry of dissolved organic carbon, nitrogen and phosphorus in tropical soils. Biogeochemistry 2000, 51, 283–302. [Google Scholar]
- Jiao, Y.; Ma, L.; Strickland, E.; Deng, X.W. Conservation and divergence of light-regulated genome expression patterns during seedling development in rice and Arabidopsis. Plant Cell 2005, 17, 3239–3256. [Google Scholar]
- Schekman, R.; Orci, L. Coat proteins and vesicle budding. Science 1996, 271, 1526–1533. [Google Scholar]
- Bar-Peled, M.; Raikhel, N.V. Characterization of AtSec12 and AtSar1, proteins likely involved in endoplasmic reticulum and Golgi transport. Plant Physiol 1997, 114, 315–324. [Google Scholar]
- Aridor, M.; Fish, K.N.; Bannykh, S.; Weissman, J.; Roberts, T.H.; Lippincott-Schwartz, J.; Balch, W.E. The Sar1 GTPase coordinates biosynthetic cargo selection with endoplasmic reticulum export site assembly. J. Cell Biol 2001, 152, 213–229. [Google Scholar]
- Takeuchi, M.; Ueda, T.; Yahara, N.; Nakano, A. Arf1 GTPase plays roles in the protein traffic between the endoplasmic reticulum and the Golgi apparatus in tobacco and Arabidopsis cultured cells. Plant J 2002, 31, 499–515. [Google Scholar]
- Takeuchi, M.; Tada, M.; Saito, C.; Yashiroda, H.; Nakano, A. Isolation of a tobacco cDNA encoding Sar1 GTPase and analysis of its dominant mutations in vesicular traffic using a yeast complementation system. Plant Cell Physiol 1998, 39, 590–599. [Google Scholar]
- Kim, W.Y.; Cheong, N.E.; Je, D.Y.; Kim, M.G.; Lim, C.O.; Bahk, J.D.; Cho, M.J.; Lee, S.Y. The presence of a Sar1 gene family in Brassica campestris that suppresses a yeast vesicular transport mutation Sec12-1. Plant Mol. Biol 1997, 33, 1025–1035. [Google Scholar]
- Andreeva, A.V.; Kutuzov, M.A.; Evans, D.E.; Hawes, C.R. Proteins involved in membrane transport between the ER and the Golgi apparatus: 21 putative plant homologues revealed by dbEST searching. Cell Biol. Int 1998, 22, 145–160. [Google Scholar]
- Yang, Y.D.; Elamawi, R.; Bubeck, J.; Pepperkok, R.; Ritzenthaler, C.; Robinson, D.G. Dynamics of COPII vesicles and the Golgi apparatus in cultured Nicotiana tabacum BY-2 cells provides evidence for transient association of Golgi stacks with endoplasmic reticulum exit sites. Plant Cell 2005, 17, 1513–1531. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cevher-Keskin, B. ARF1 and SAR1 GTPases in Endomembrane Trafficking in Plants. Int. J. Mol. Sci. 2013, 14, 18181-18199. https://doi.org/10.3390/ijms140918181
Cevher-Keskin B. ARF1 and SAR1 GTPases in Endomembrane Trafficking in Plants. International Journal of Molecular Sciences. 2013; 14(9):18181-18199. https://doi.org/10.3390/ijms140918181
Chicago/Turabian StyleCevher-Keskin, Birsen. 2013. "ARF1 and SAR1 GTPases in Endomembrane Trafficking in Plants" International Journal of Molecular Sciences 14, no. 9: 18181-18199. https://doi.org/10.3390/ijms140918181
APA StyleCevher-Keskin, B. (2013). ARF1 and SAR1 GTPases in Endomembrane Trafficking in Plants. International Journal of Molecular Sciences, 14(9), 18181-18199. https://doi.org/10.3390/ijms140918181