TrkB Receptor Signalling: Implications in Neurodegenerative, Psychiatric and Proliferative Disorders


{kind=link}
{kind=link}
Abstract
:1. Introduction
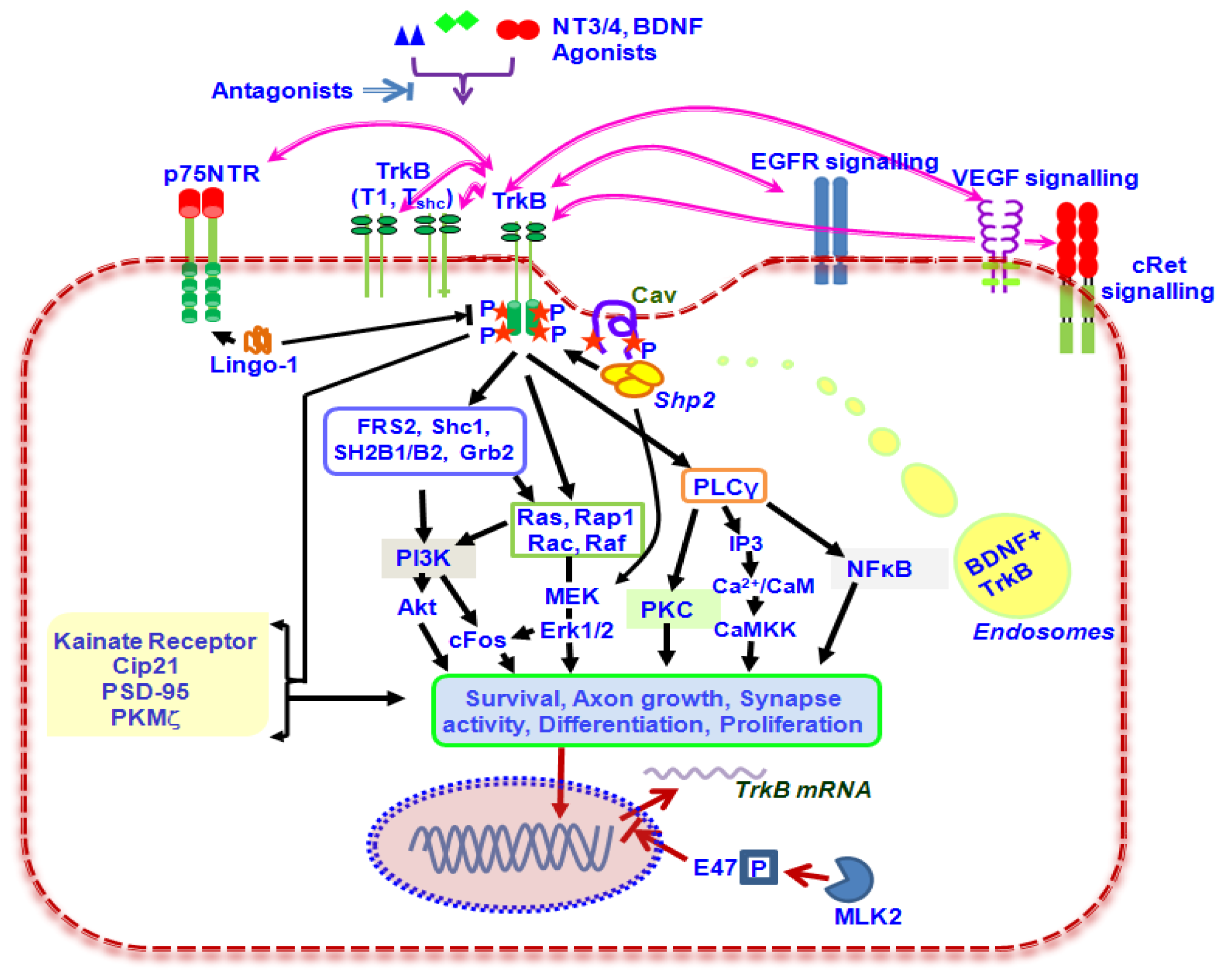
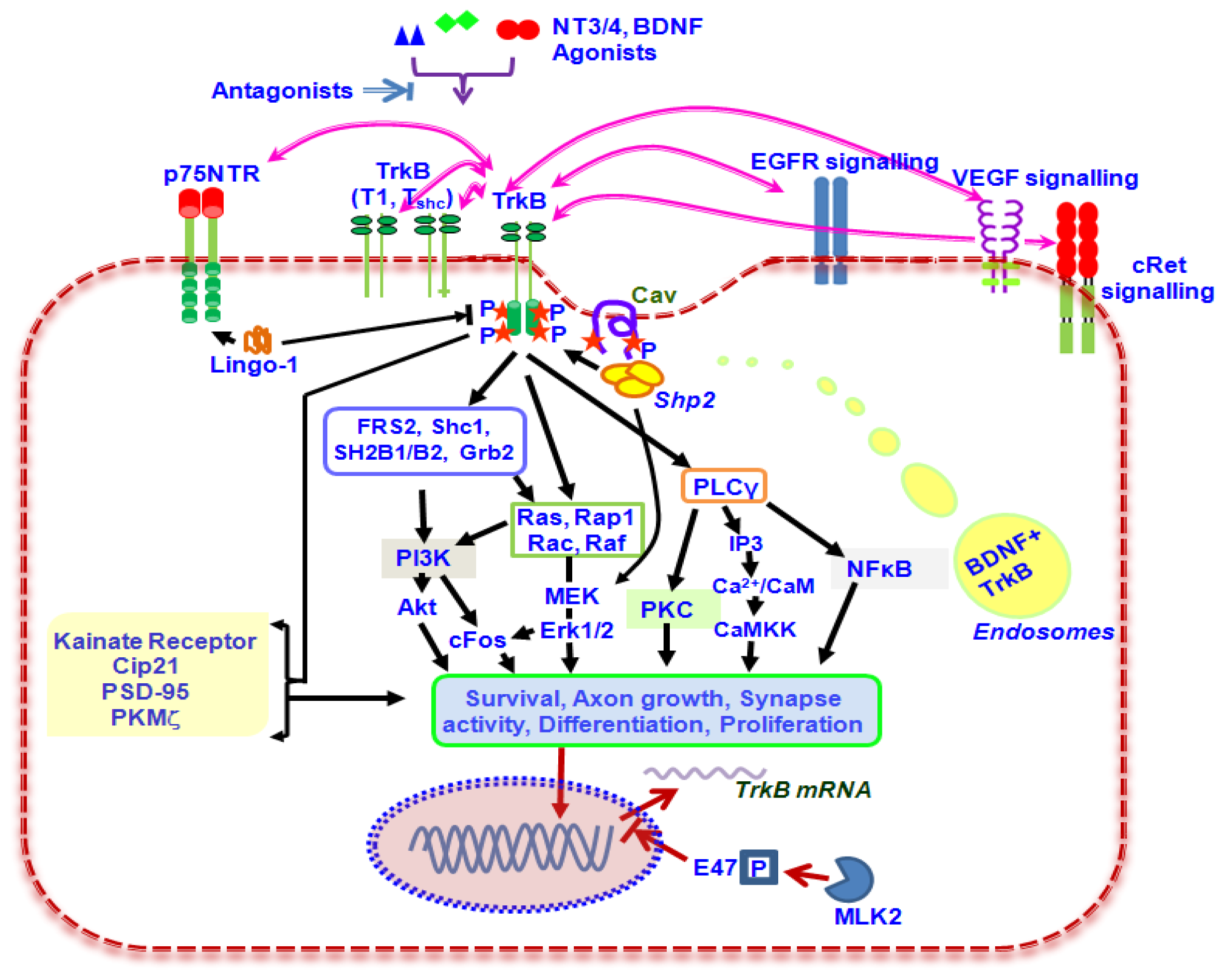
2. Biological Functions of TrkB Signalling
3. Role of TrkB Splice Variants
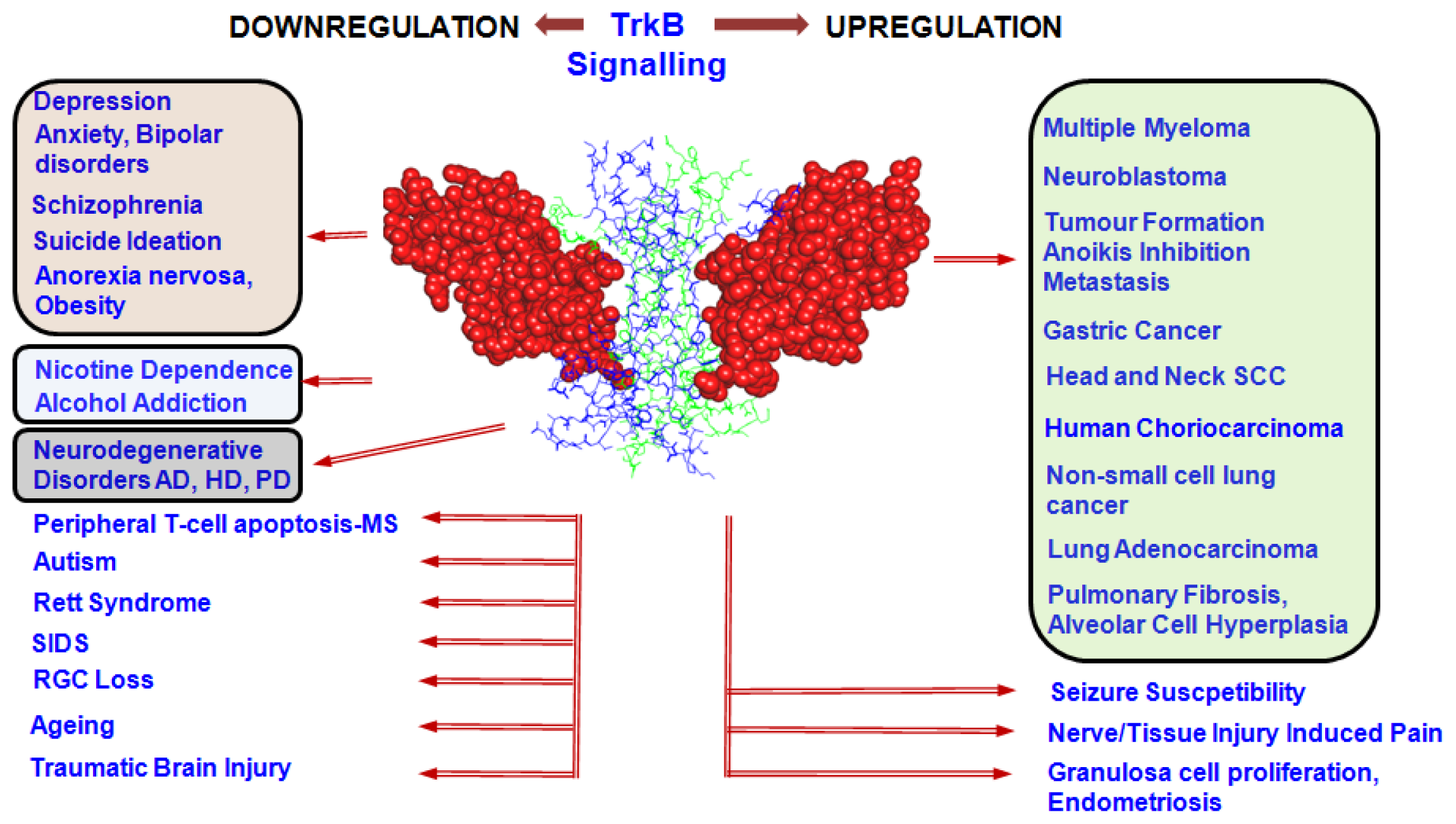
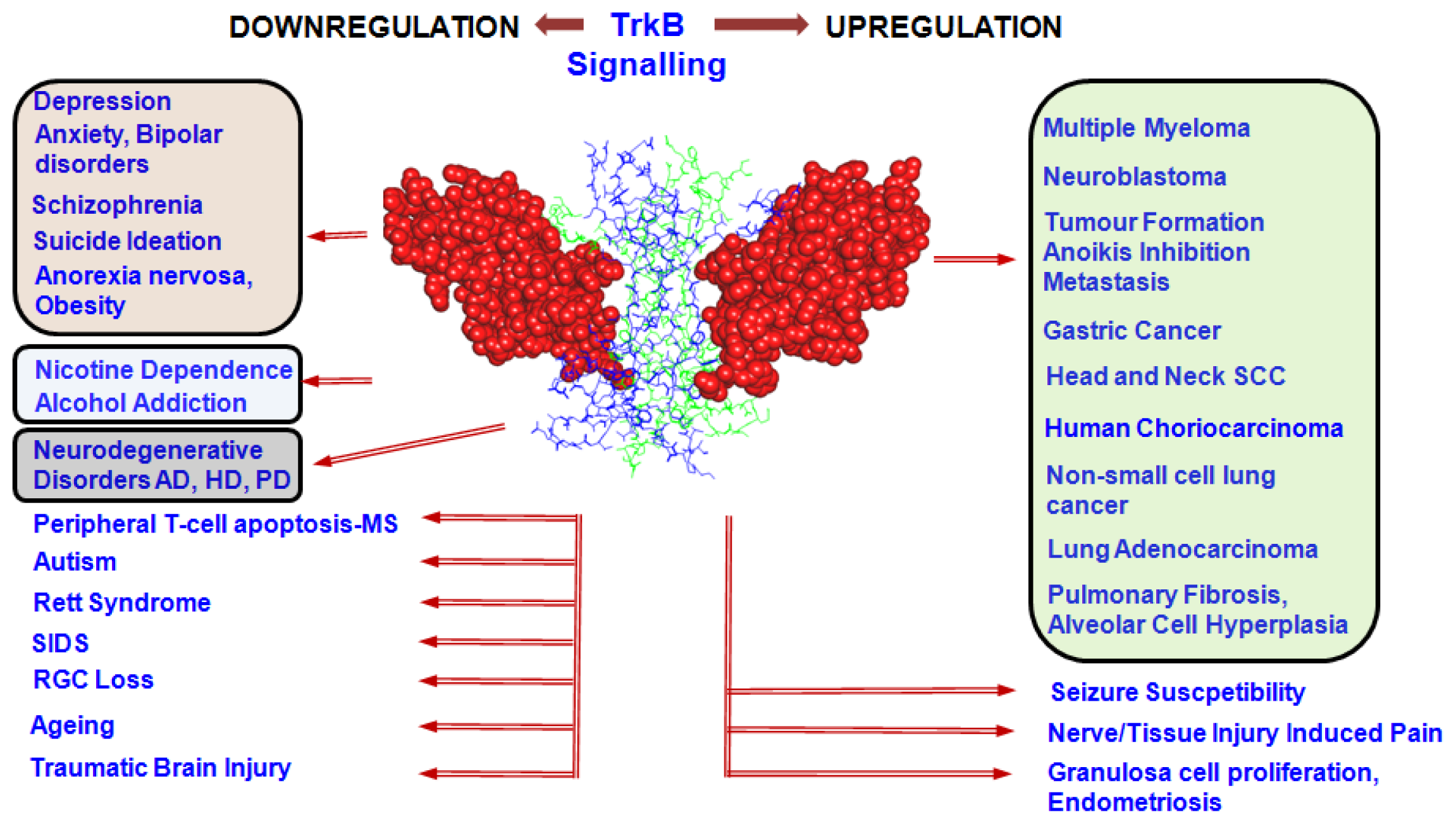
4. Involvement of TrkB Signalling in Disease Processes
4.1. TrkB in Proliferative Disorders
4.2. TrkB in Psychiatric Disorders
4.3. TrkB in Neurodegenerative Disorders
4.4. TrkB in Other Disorders
5. Therapeutic Potential of TrkB Pathway Modulation
6. Conclusions
Acknowledgements
Abbreviations
| Trk | Tropomyosin Related Kinase |
| P75NTR | p75 Neurotrophin Receptor |
| SH2 | Src Homology 2 |
| FRS2 | Fibroblast growth factor receptor substrate 2 |
| Shc | Src homology 2 domain containing |
| SH2B | Src homology 2 domain containing adaptor protein B |
| SH2B2 | SH2B adaptor protein 2 |
| MAPK | Mitogen Activated Protein Kinase |
| VEGF | Vascular Endothelial Growth factor |
| EGFR | Epidermal Growth Factor Receptor |
| PSD-95 | Postsynaptic Density Protein-95 |
| PTPN11 | Tyrosine-protein phosphatase non-receptor type 11 |
| Shp2 | SH2 domain containing Tyrosine Phosphatse 2 |
| Pyk2 | Protein tyrosine Kinase 2 |
| Nogo | Neurite Outgrowth inhibitor |
| NOTCH2 | Neurogenic locus notch homolog protein 2 |
| JNK | c-Jun N-terminal kinase. |
References
- Reichardt, L.F. Neurotrophin-regulated signalling pathways. Philos. TR Soc. B 2006, 361, 1545–1564. [Google Scholar]
- Massa, S.M.; Yang, T.; Xie, Y.; Shi, J.; Bilgen, M.; Joyce, J.N.; Nehama, D.; Rajadas, J.; Longo, F.M. Small molecule BDNF mimetics activate TrkB signaling and prevent neuronal degeneration in rodents. J. Clin. Invest 2010, 120, 1774–1785. [Google Scholar]
- Stoilov, P.; Castren, E.; Stamm, S. Analysis of the human TrkB gene genomic organization reveals novel TrkB isoforms, unusual gene length, and splicing mechanism. Biochem. Biophys. Res. Commun 2002, 290, 1054–1065. [Google Scholar]
- Yeo, G.S.; Connie Hung, C.C.; Rochford, J.; Keogh, J.; Gray, J.; Sivaramakrishnan, S.; O'Rahilly, S.; Farooqi, I.S. A de novo mutation affecting human TrkB associated with severe obesity and developmental delay. Nat. Neurosci 2004, 7, 1187–1189. [Google Scholar]
- Barbacid, M. The trk family of neurotrophin receptors. J. Neurobiol 1994, 25, 1386–1403. [Google Scholar]
- Shelton, D.L.; Sutherland, J.; Gripp, J.; Camerato, T.; Armanini, M.P.; Phillips, H.S.; Carroll, K.; Spencer, S.D.; Levinson, A.D. Human trks: Molecular cloning, tissue distribution, and expression of extracellular domain immunoadhesins. J. Neurosci 1995, 15, 477–491. [Google Scholar]
- Calavia, M.G.; Feito, J.; Lopez-Iglesias, L.; de Carlos, F.; Garcia-Suarez, O.; Perez-Pinera, P.; Cobo, J.; Vega, J.A. The lamellar cells in human Meissner corpuscles express TrkB. Neurosci. Lett 2010, 468, 106–109. [Google Scholar]
- Kaplan, D.R.; Miller, F.D. Neurotrophin signal transduction in the nervous system. Curr. Opin. Neurobiol 2000, 10, 381–391. [Google Scholar]
- Minichiello, L. TrkB signalling pathways in LTP and learning. Nat. Rev. Neurosci 2009, 10, 850–860. [Google Scholar]
- Cowley, S.; Paterson, H.; Kemp, P.; Marshall, C.J. Activation of MAP kinase kinase is necessary and sufficient for PC12 differentiation and for transformation of NIH 3T3 cells. Cell 1994, 77, 841–852. [Google Scholar]
- Meakin, S.O.; MacDonald, J.I.; Gryz, E.A.; Kubu, C.J.; Verdi, J.M. The signaling adapter FRS-2 competes with Shc for binding to the nerve growth factor receptor TrkA. A model for discriminating proliferation and differentiation. J. Biol. Chem 1999, 274, 9861–9870. [Google Scholar]
- Huang, E.J.; Reichardt, L.F. Trk receptors: Roles in neuronal signal transduction. Annu. Rev. Biochem 2003, 72, 609–642. [Google Scholar]
- Miyamoto, Y.; Yamauchi, J.; Tanoue, A.; Wu, C.; Mobley, W.C. TrkB binds and tyrosine-phosphorylates Tiam1, leading to activation of Rac1 and induction of changes in cellular morphology. Proc. Natl. Acad. Sci. USA 2006, 103, 10444–10449. [Google Scholar]
- Geetha, T.; Wooten, M.W. Association of the atypical protein kinase C-interacting protein p62/ZIP with nerve growth factor receptor TrkA regulates receptor trafficking and Erk5 signaling. J. Biol. Chem 2003, 278, 4730–4739. [Google Scholar]
- Arevalo, J.C.; Yano, H.; Teng, K.K.; Chao, M.V. A unique pathway for sustained neurotrophin signaling through an ankyrin-rich membrane-spanning protein. EMBO J 2004, 23, 2358–2368. [Google Scholar]
- Liu, Y.; Encinas, M.; Comella, J.X.; Aldea, M.; Gallego, C. Basic helix-loop-helix proteins bind to TrkB and p21(Cip1) promoters linking differentiation and cell cycle arrest in neuroblastoma cells. Mol. Cell Biol 2004, 24, 2662–2672. [Google Scholar]
- Pedraza, N.; Rafel, M.; Navarro, I.; Encinas, M.; Aldea, M.; Gallego, C. Mixed lineage kinase phosphorylates transcription factor E47 and inhibits TrkB expression to link neuronal death and survival pathways. J. Biol. Chem 2009, 284, 32980–32988. [Google Scholar]
- Soppet, D.; Escandon, E.; Maragos, J.; Middlemas, D.S.; Reid, S.W.; Blair, J.; Burton, L.E.; Stanton, B.R.; Kaplan, D.R.; Hunter, T.; et al. The neurotrophic factors brain-derived neurotrophic factor and neurotrophin-3 are ligands for the trkB tyrosine kinase receptor. Cell 1991, 65, 895–903. [Google Scholar]
- McCarty, J.H.; Feinstein, S.C. Activation loop tyrosines contribute varying roles to TrkB autophosphorylation and signal transduction. Oncogene 1998, 16, 1691–700. [Google Scholar]
- Easton, J.B.; Moody, N.M.; Zhu, X.; Middlemas, D.S. Brain-derived neurotrophic factor induces phosphorylation of fibroblast growth factor receptor substrate 2. J. Biol. Chem 1999, 274, 11321–11327. [Google Scholar]
- Atwal, J.K.; Massie, B.; Miller, F.D.; Kaplan, D.R. The TrkB-Shc site signals neuronal survival and local axon growth via MEK and P13-kinase. Neuron 2000, 27, 265–277. [Google Scholar]
- Bilderback, T.R.; Gazula, V.R.; Lisanti, M.P.; Dobrowsky, R.T. Caveolin interacts with Trk A and p75(NTR) and regulates neurotrophin signaling pathways. J. Biol. Chem 1999, 274, 257–263. [Google Scholar]
- Suzuki, S.; Numakawa, T.; Shimazu, K.; Koshimizu, H.; Hara, T.; Hatanaka, H.; Mei, L.; Lu, B.; Kojima, M. BDNF-induced recruitment of TrkB receptor into neuronal lipid rafts: Roles in synaptic modulation. J. Cell Biol 2004, 167, 1205–1215. [Google Scholar]
- Okada, N.; Wada, K.; Goldsmith, B.A.; Koizumi, S. SHP-2 is involved in neurotrophin signaling. Biochem. Biophys. Res. Commun 1996, 229, 607–611. [Google Scholar]
- Basavarajappa, D.K.; Gupta, V.K.; Dighe, R.; Rajala, A.; Rajala, R.V. Phosphorylated Grb14 is an endogenous inhibitor of retinal protein tyrosine phosphatase 1B, and light-dependent activation of Src phosphorylates Grb14. Mol. Cell Biol 2011, 31, 3975–3987. [Google Scholar]
- Basavarajappa, D.K.; Gupta, V.K.; Rajala, R.V. Protein tyrosine phosphatase 1B: A novel molecular target for retinal degenerative diseases. Adv. Exp. Med. Biol 2012, 723, 829–834. [Google Scholar]
- Rusanescu, G.; Yang, W.; Bai, A.; Neel, B.G.; Feig, L.A. Tyrosine phosphatase SHP-2 is a mediator of activity-dependent neuronal excitotoxicity. EMBO J 2005, 24, 305–314. [Google Scholar]
- Easton, J.B.; Royer, A.R.; Middlemas, D.S. The protein tyrosine phosphatase, Shp2, is required for the complete activation of the RAS/MAPK pathway by brain-derived neurotrophic factor. J. Neurochem 2006, 97, 834–845. [Google Scholar]
- Gupta, V.K.; You, Y.; Klistorner, A.; Graham, S.L. Shp-2 regulates the TrkB receptor activity in the retinal ganglion cells under glaucomatous stress. Biochim. Biophys. Acta 2012, 1822, 1643–1649. [Google Scholar]
- You, Y.; Gupta, V.; Li, J.; Klistorner, A.; Graham, S.L. Optic neuropathies: Characteristic features and mechanisms of retinal ganglion cell loss. Rev. Neurosci. 2013. [Google Scholar] [CrossRef]
- Kumamaru, E.; Numakawa, T.; Adachi, N.; Kunugi, H. Glucocorticoid suppresses BDNF-stimulated MAPK/ERK pathway via inhibiting interaction of Shp2 with TrkB. FEBS Lett 2011, 585, 3224–3228. [Google Scholar]
- Klein, R.; Smeyne, R.J.; Wurst, W.; Long, L.K.; Auerbach, B.A.; Joyner, A.L.; Barbacid, M. Targeted disruption of the trkB neurotrophin receptor gene results in nervous system lesions and neonatal death. Cell 1993, 75, 113–122. [Google Scholar]
- Schimmang, T.; Minichiello, L.; Vazquez, E.; San Jose, I.; Giraldez, F.; Klein, R.; Represa, J. Developing inner ear sensory neurons require TrkB and TrkC receptors for innervation of their peripheral targets. Development 1995, 121, 3381–3391. [Google Scholar]
- Minichiello, L.; Casagranda, F.; Tatche, R.S.; Stucky, C.L.; Postigo, A.; Lewin, G.R.; Davies, A.M.; Klein, R. Point mutation in trkB causes loss of NT4-dependent neurons without major effects on diverse BDNF responses. Neuron 1998, 21, 335–345. [Google Scholar]
- Webster, M.J.; Herman, M.M.; Kleinman, J.E.; Shannon Weickert, C. BDNF and trkB mRNA expression in the hippocampus and temporal cortex during the human lifespan. Gene Exp. Patterns 2006, 6, 941–951. [Google Scholar]
- Sallert, M.; Rantamaki, T.; Vesikansa, A.; Anthoni, H.; Harju, K.; Yli-Kauhaluoma, J.; Taira, T.; Castren, E.; Lauri, S.E. Brain-derived neurotrophic factor controls activity-dependent maturation of CA1 synapses by downregulating tonic activation of presynaptic kainate receptors. J. Neurosci 2009, 29, 11294–11303. [Google Scholar]
- Wang, X.; Ratnam, J.; Zou, B.; England, P.M.; Basbaum, A.I. TrkB signaling is required for both the induction and maintenance of tissue and nerve injury-induced persistent pain. J. Neurosci 2009, 29, 5508–5515. [Google Scholar]
- Han, L.; Zhang, Z.; Qin, W.; Sun, W. Neurotrophic receptor TrkB: Is it a predictor of poor prognosis for carcinoma patients? Med. Hypotheses 2007, 68, 407–409. [Google Scholar]
- Edsjo, A.; Lavenius, E.; Nilsson, H.; Hoehner, J.C.; Simonsson, P.; Culp, L.A.; Martinsson, T.; Larsson, C.; Pahlman, S. Expression of trkB in human neuroblastoma in relation to MYCN expression and retinoic acid treatment. Lab. Invest 2003, 83, 813–823. [Google Scholar]
- Nakamura, K.; Martin, K.C.; Jackson, J.K.; Beppu, K.; Woo, C.W.; Thiele, C.J. Brain-derived neurotrophic factor activation of TrkB induces vascular endothelial growth factor expression via hypoxia-inducible factor-1alpha in neuroblastoma cells. Cancer Res 2006, 66, 4249–4255. [Google Scholar]
- Esposito, C.L.; D’Alessio, A.; de Franciscis, V.; Cerchia, L. A cross-talk between TrkB and ret tyrosine kinases receptors mediates neuroblastoma cells differentiation. PLoS One 2008, 3, e1643. [Google Scholar]
- Qiu, L.; Zhou, C.; Sun, Y.; Di, W.; Scheffler, E.; Healey, S.; Kouttab, N.; Chu, W.; Wan, Y. Crosstalk between EGFR and TrkB enhances ovarian cancer cell migration and proliferation. Int. J. Oncol 2006, 29, 1003–1011. [Google Scholar]
- Siu, M.K.; Wong, O.G.; Cheung, A.N. TrkB as a therapeutic target for ovarian cancer. Expert Opin. Ther. Targets 2009, 13, 1169–1178. [Google Scholar]
- Hayashi, A.; Nakatani, K.; Nishioka, J.; Sakamoto, Y.; Jinda, S.; Wada, H.; Nobori, T. Neurotrophic receptor tyrosine kinase B induces c-fos-associated cell survival. Int. J. Mol. Med 2009, 24, 807–811. [Google Scholar]
- Au, C.W.; Siu, M.K.; Liao, X.; Wong, E.S.; Ngan, H.Y.; Tam, K.F.; Chan, D.C.; Chan, Q.K.; Cheung, A.N. Tyrosine kinase B receptor and BDNF expression in ovarian cancers—Effect on cell migration, angiogenesis and clinical outcome. Cancer Lett 2009, 281, 151–161. [Google Scholar]
- Grishanin, R.N.; Yang, H.; Liu, X.; Donohue-Rolfe, K.; Nune, G.C.; Zang, K.; Xu, B.; Duncan, J.L.; Lavail, M.M.; Copenhagen, D.R.; et al. Retinal TrkB receptors regulate neural development in the inner, but not outer, retina. Mol. Cell Neurosci 2008, 38, 431–443. [Google Scholar]
- Cheng, L.; Sapieha, P.; Kittlerova, P.; Hauswirth, W.W.; di Polo, A. TrkB gene transfer protects retinal ganglion cells from axotomy-induced death in vivo. J. Neurosci 2002, 22, 3977–3986. [Google Scholar]
- Von Bartheld, C.S.; Williams, R.; Lefcort, F.; Clary, D.O.; Reichardt, L.F.; Bothwell, M. Retrograde transport of neurotrophins from the eye to the brain in chick embryos: Roles of the p75NTR and trkB receptors. J. Neurosci 1996, 16, 2995–3008. [Google Scholar]
- Turner, B.A.; Sparrow, J.; Cai, B.; Monroe, J.; Mikawa, T.; Hempstead, B.L. TrkB/BDNF signaling regulates photoreceptor progenitor cell fate decisions. Dev. Biol 2006, 299, 455–465. [Google Scholar]
- Di Polo, A.; Cheng, L.; Bray, G.M.; Aguayo, A.J. Colocalization of TrkB and brain-derived neurotrophic factor proteins in green-red-sensitive cone outer segments. Invest. Ophth. Vis. Sci 2000, 41, 4014–4021. [Google Scholar]
- Trifunovic, D.; Sahaboglu, A.; Kaur, J.; Mencl, S.; Zrenner, E.; Ueffing, M.; Arango-Gonzalez, B.; Paquet-Durand, F. Neuroprotective strategies for the treatment of inherited photoreceptor degeneration. Curr. Mol. Med 2012, 12, 598–612. [Google Scholar]
- Harada, C.; Guo, X.; Namekata, K.; Kimura, A.; Nakamura, K.; Tanaka, K.; Parada, L.F.; Harada, T. Glia- and neuron-specific functions of TrkB signalling during retinal degeneration and regeneration. Nat. Commun 2011, 2, 189. [Google Scholar]
- Liu, Z.Z.; Zhu, L.Q.; Eide, F.F. Critical role of TrkB and brain-derived neurotrophic factor in the differentiation and survival of retinal pigment epithelium. J. Neurosci 1997, 17, 8749–8755. [Google Scholar]
- Saito, T.; Abe, T.; Wakusawa, R.; Sato, H.; Asai, H.; Tokita-Ishikawa, Y.; Nishida, K. TrkB-T1 receptors on Muller cells play critical role in brain-derived neurotrophic factor-mediated photoreceptor protection against phototoxicity. Curr. Eye Res 2009, 34, 580–588. [Google Scholar]
- Yoshii, A.; Murata, Y.; Kim, J.; Zhang, C.; Shokat, K.M.; Constantine-Paton, M. TrkB and protein kinase Mzeta regulate synaptic localization of PSD-95 in developing cortex. J. Neurosci 2011, 31, 11894–11904. [Google Scholar]
- Patapoutian, A.; Reichardt, L.F. Trk receptors: Mediators of neurotrophin action. Curr. Opin. Neurobiol 2001, 11, 272–280. [Google Scholar]
- Allen, S.J.; Dawbarn, D.; Eckford, S.D.; Wilcock, G.K.; Ashcroft, M.; Colebrook, S.M.; Feeney, R.; MacGowan, S.H. Cloning of a non-catalytic form of human trkB and distribution of messenger RNA for trkB in human brain. Neuroscience 1994, 60, 825–834. [Google Scholar]
- Haapasalo, A.; Sipola, I.; Larsson, K.; Akerman, K.E.; Stoilov, P.; Stamm, S.; Wong, G.; Castren, E. Regulation of TRKB surface expression by brain-derived neurotrophic factor and truncated TRKB isoforms. J. Biol. Chem 2002, 277, 43160–43167. [Google Scholar]
- Fryer, R.H.; Kaplan, D.R.; Kromer, L.F. Truncated trkB receptors on nonneuronal cells inhibit BDNF-induced neurite outgrowth in vitro. Exp. Neurol 1997, 148, 616–627. [Google Scholar]
- Fenner, B.M. Truncated TrkB: Beyond a dominant negative receptor. Cytokine Growth Factor Rev 2012, 23, 15–24. [Google Scholar]
- Yacoubian, T.A.; Lo, D.C. Truncated and full-length TrkB receptors regulate distinct modes of dendritic growth. Nat. Neurosci 2000, 3, 342–349. [Google Scholar]
- Kafitz, K.W.; Rose, C.R.; Thoenen, H.; Konnerth, A. Neurotrophin-evoked rapid excitation through TrkB receptors. Nature 1999, 401, 918–921. [Google Scholar]
- Blum, R.; Kafitz, K.W.; Konnerth, A. Neurotrophin-evoked depolarization requires the sodium channel Na(V)1.9. Nature 2002, 419, 687–693. [Google Scholar]
- Rose, C.R.; Blum, R.; Pichler, B.; Lepier, A.; Kafitz, K.W.; Konnerth, A. Truncated TrkB-T1 mediates neurotrophin-evoked calcium signalling in glia cells. Nature 2003, 426, 74–78. [Google Scholar]
- Nakagawara, A. Trk receptor tyrosine kinases: A bridge between cancer and neural development. Cancer Lett 2001, 169, 107–114. [Google Scholar]
- Zhang, J.; Zheng, Y.; Wang, Y.; Tong, H. The studies on the correlation for gene expression of tyrosine-kinase receptors and vascular endothelial growth factor in human neuroblastomas. J. Pediatr. Hematol. Oncol 2010, 32, 180–184. [Google Scholar]
- Hecht, M.; Schulte, J.H.; Eggert, A.; Wilting, J.; Schweigerer, L. The neurotrophin receptor TrkB cooperates with c-Met in enhancing neuroblastoma invasiveness. Carcinogenesis 2005, 26, 2105–2115. [Google Scholar]
- Schulte, J.H.; Schramm, A.; Klein-Hitpass, L.; Klenk, M.; Wessels, H.; Hauffa, B.P.; Eils, J.; Eils, R.; Brodeur, G.M.; Schweigerer, L.; et al. Microarray analysis reveals differential gene expression patterns and regulation of single target genes contributing to the opposing phenotype of TrkA- and TrkB-expressing neuroblastomas. Oncogene 2005, 24, 165–177. [Google Scholar]
- Geiger, T.R.; Peeper, D.S. Critical role for TrkB kinase function in anoikis suppression, tumorigenesis, and metastasis. Cancer Res 2007, 67, 6221–6229. [Google Scholar]
- Geiger, T.R.; Peeper, D.S. The neurotrophic receptor TrkB in anoikis resistance and metastasis: A perspective. Cancer Res 2005, 65, 7033–7036. [Google Scholar]
- Greenman, C.; Stephens, P.; Smith, R.; Dalgliesh, G.L.; Hunter, C.; Bignell, G.; Davies, H.; Teague, J.; Butler, A.; Stevens, C.; et al. Patterns of somatic mutation in human cancer genomes. Nature 2007, 446, 153–158. [Google Scholar]
- Pearse, R.N.; Swendeman, S.L.; Li, Y.; Rafii, D.; Hempstead, B.L. A neurotrophin axis in myeloma: TrkB and BDNF promote tumor-cell survival. Blood 2005, 105, 4429–4436. [Google Scholar]
- Tanaka, K.; Mohri, Y.; Nishioka, J.; Kobayashi, M.; Ohi, M.; Miki, C.; Tonouchi, H.; Nobori, T.; Kusunoki, M. Neurotrophic receptor, tropomyosin-related kinase B as an independent prognostic marker in gastric cancer patients. J. Surg. Oncol 2009, 99, 307–310. [Google Scholar]
- Kupferman, M.E.; Jiffar, T.; El-Naggar, A.; Yilmaz, T.; Zhou, G.; Xie, T.; Feng, L.; Wang, J.; Holsinger, F.C.; Yu, D.; et al. TrkB induces EMT and has a key role in invasion of head and neck squamous cell carcinoma. Oncogene 2010, 29, 2047–2059. [Google Scholar]
- Zhang, S.; Guo, D.; Luo, W.; Zhang, Q.; Zhang, Y.; Li, C.; Lu, Y.; Cui, Z.; Qiu, X. TrkB is highly expressed in NSCLC and mediates BDNF-induced the activation of Pyk2 signaling and the invasion of A549 cells. BMC Cancer 2010, 10, 43. [Google Scholar]
- Avcuoglu, S.; Wygrecka, M.; Marsh, L.M.; Gunther, A.; Seeger, W.; Weissmann, N.; Fink, L.; Morty, R.E.; Kwapiszewska, G. Neurotrophic tyrosine kinase receptor B/neurotrophin 4 signaling axis is perturbed in clinical and experimental pulmonary fibrosis. Am. J. Respir. Cell Mol. Biol 2011, 45, 768–780. [Google Scholar]
- Lin, E.; Hong, C.J.; Hwang, J.P.; Liou, Y.J.; Yang, C.H.; Cheng, D.; Tsai, S.J. Gene-gene interactions of the brain-derived neurotrophic-factor and neurotrophic tyrosine kinase receptor 2 genes in geriatric depression. Rejuven. Res 2009, 12, 387–393. [Google Scholar]
- Hung, Y.Y.; Lin, C.J.; Huang, T.L. Higher serum tropomyosin-related kinase B protein level in major depression. Progr. Neuropsychopharmacol. Biol. Psychiatry 2010, 34, 610–612. [Google Scholar]
- Ernst, C.; Wanner, B.; Brezo, J.; Vitaro, F.; Tremblay, R.; Turecki, G. A deletion in tropomyosin-related kinase B and the development of human anxiety. Biol. Psychiatry 2011, 69, 604–607. [Google Scholar]
- Soontornniyomkij, B.; Everall, I.P.; Chana, G.; Tsuang, M.T.; Achim, C.L.; Soontornniyomkij, V. Tyrosine kinase B protein expression is reduced in the cerebellum of patients with bipolar disorder. J. Affect. Disord 2011, 133, 646–654. [Google Scholar]
- Thompson Ray, M.; Weickert, C.S.; Wyatt, E.; Webster, M.J. Decreased BDNF, trkB-TK+ and GAD67 mRNA expression in the hippocampus of individuals with schizophrenia and mood disorders. J. Psychiatry Neurosci 2011, 36, 195–203. [Google Scholar]
- Iritani, S.; Niizato, K.; Nawa, H.; Ikeda, K.; Emson, P.C. Immunohistochemical study of brain-derived neurotrophic factor and its receptor, TrkB, in the hippocampal formation of schizophrenic brains. Progr. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 801–807. [Google Scholar]
- Dwivedi, Y.; Rizavi, H.S.; Conley, R.R.; Roberts, R.C.; Tamminga, C.A.; Pandey, G.N. Altered gene expression of brain-derived neurotrophic factor and receptor tyrosine kinase B in postmortem brain of suicide subjects. Arch. Gen. Psychiatry 2003, 60, 804–815. [Google Scholar]
- Beuten, J.; Ma, J.Z.; Payne, T.J.; Dupont, R.T.; Lou, X.Y.; Crews, K.M.; Elston, R.C.; Li, M.D. Association of specific haplotypes of neurotrophic tyrosine kinase receptor 2 gene (NTRK2) with vulnerability to nicotine dependence in African-Americans and European-Americans. Biol. Psychiatry 2007, 61, 48–55. [Google Scholar]
- Xu, K.; Anderson, T.R.; Neyer, K.M.; Lamparella, N.; Jenkins, G.; Zhou, Z.; Yuan, Q.; Virkkunen, M.; Lipsky, R.H. Nucleotide sequence variation within the human tyrosine kinase B neurotrophin receptor gene: Association with antisocial alcohol dependence. Pharmacogenomics J 2007, 7, 368–379. [Google Scholar]
- Chen, Z.; Simmons, M.S.; Perry, R.T.; Wiener, H.W.; Harrell, L.E.; Go, R.C. Genetic association of neurotrophic tyrosine kinase receptor type 2 (NTRK2) With Alzheimer’s disease. Am. J. Med. Genet. B 2008, 147, 363–369. [Google Scholar]
- Baydyuk, M.; Nguyen, M.T.; Xu, B. Chronic deprivation of TrkB signaling leads to selective late-onset nigrostriatal dopaminergic degeneration. Exp. Neurol 2011, 228, 118–125. [Google Scholar]
- Colombo, E.; Cordiglieri, C.; Melli, G.; Newcombe, J.; Krumbholz, M.; Parada, L.F.; Medico, E.; Hohlfeld, R.; Meinl, E.; Farina, C. Stimulation of the neurotrophin receptor TrkB on astrocytes drives nitric oxide production and neurodegeneration. J. Exp. Med 2012, 209, 521–535. [Google Scholar]
- De Santi, L.; Cantalupo, L.; Tassi, M.; Raspadori, D.; Cioni, C.; Annunziata, P. Higher expression of BDNF receptor gp145trkB is associated with lower apoptosis intensity in T cell lines in multiple sclerosis. J. Neurol. Sci 2009, 277, 65–70. [Google Scholar]
- Correia, C.T.; Coutinho, A.M.; Sequeira, A.F.; Sousa, I.G.; Lourenco Venda, L.; Almeida, J.P.; Abreu, R.L.; Lobo, C.; Miguel, T.S.; Conroy, J.; et al. Increased BDNF levels and NTRK2 gene association suggest a disruption of BDNF/TrkB signaling in autism. Genes Brain Behav 2010, 9, 841–848. [Google Scholar]
- Louhivuori, V.; Vicario, A.; Uutela, M.; Rantamaki, T.; Louhivuori, L.M.; Castren, E.; Tongiorgi, E.; Akerman, K.E.; Castren, M.L. BDNF and TrkB in neuronal differentiation of Fmr1-knockout mouse. Neurobiol. Dis 2011, 41, 469–480. [Google Scholar]
- Abuhatzira, L.; Makedonski, K.; Kaufman, Y.; Razin, A.; Shemer, R. MeCP2 deficiency in the brain decreases BDNF levels by REST/CoREST-mediated repression and increases TRKB production. Epigenetics 2007, 2, 214–222. [Google Scholar]
- Tang, S.; Machaalani, R.; Waters, K.A. Expression of brain-derived neurotrophic factor and TrkB receptor in the sudden infant death syndrome brainstem. Respir. Physiol. Neurobiol 2012, 180, 25–33. [Google Scholar]
- He, X.P.; Kotloski, R.; Nef, S.; Luikart, B.W.; Parada, L.F.; McNamara, J.O. Conditional deletion of TrkB but not BDNF prevents epileptogenesis in the kindling model. Neuron 2004, 43, 31–42. [Google Scholar]
- Marvanova, M.; Lakso, M.; Pirhonen, J.; Nawa, H.; Wong, G.; Castren, E. The neuroprotective agent memantine induces brain-derived neurotrophic factor and trkB receptor expression in rat brain. Mol. Cell Neurosci 2001, 18, 247–258. [Google Scholar]
- Ly, T.; Gupta, N.; Weinreb, R.N.; Kaufman, P.L.; Yucel, Y.H. Dendrite plasticity in the lateral geniculate nucleus in primate glaucoma. Vis. Res 2011, 51, 243–250. [Google Scholar]
- Vrabec, J.P.; Levin, L.A. The neurobiology of cell death in glaucoma. Eye 2007, 21, S11–S14. [Google Scholar]
- Johnson, E.C.; Guo, Y.; Cepurna, W.O.; Morrison, J.C. Neurotrophin roles in retinal ganglion cell survival: Lessons from rat glaucoma models. Exp. Eye Res 2009, 88, 808–815. [Google Scholar]
- Pease, M.E.; McKinnon, S.J.; Quigley, H.A.; Kerrigan-Baumrind, L.A.; Zack, D.J. Obstructed axonal transport of BDNF and its receptor TrkB in experimental glaucoma. Invest. Ophth. Vis. Sci 2000, 41, 764–774. [Google Scholar]
- Fu, Q.L.; Hu, B.; Li, X.; Shao, Z.; Shi, J.B.; Wu, W.; So, K.F.; Mi, S. LINGO-1 negatively regulates TrkB phosphorylation after ocular hypertension. Eur. J. Neurosci 2010, 31, 1091–1097. [Google Scholar]
- Mi, S.; Lee, X.; Shao, Z.; Thill, G.; Ji, B.; Relton, J.; Levesque, M.; Allaire, N.; Perrin, S.; Sands, B.; et al. LINGO-1 is a component of the Nogo-66 receptor/p75 signaling complex. Nat. Neurosci 2004, 7, 221–228. [Google Scholar]
- Ribases, M.; Gratacos, M.; Badia, A.; Jimenez, L.; Solano, R.; Vallejo, J.; Fernandez-Aranda, F.; Estivill, X. Contribution of NTRK2 to the genetic susceptibility to anorexia nervosa, harm avoidance and minimum body mass index. Mol. Psychiatry 2005, 10, 851–860. [Google Scholar]
- Dorfman, M.D.; Kerr, B.; Garcia-Rudaz, C.; Paredes, A.H.; Dissen, G.A.; Ojeda, S.R. Neurotrophins acting via TRKB receptors activate the JAGGED1-NOTCH2 cell-cell communication pathway to facilitate early ovarian development. Endocrinology 2011, 152, 5005–5016. [Google Scholar]
- Harel, S.; Jin, S.; Fisch, B.; Feldberg, D.; Krissi, H.; Felz, C.; Freimann, S.; Tan, S.L.; Ao, A.; Abir, R. Tyrosine kinase B receptor and its activated neurotrophins in ovaries from human fetuses and adults. Mol. Hum. Reprod 2006, 12, 357–365. [Google Scholar]
- Mayeur, S.; Silhol, M.; Moitrot, E.; Barbaux, S.; Breton, C.; Gabory, A.; Vaiman, D.; Dutriez-Casteloot, I.; Fajardy, I.; Vambergue, A.; et al. Placental BDNF/TrkB signaling system is modulated by fetal growth disturbances in rat and human. Placenta 2010, 31, 785–791. [Google Scholar]
- Anger, D.L.; Zhang, B.; Boutross-Tadross, O.; Foster, W.G. Tyrosine receptor kinase B (TrkB) protein expression in the human endometrium. Endocrine 2007, 31, 167–173. [Google Scholar]
- Schindowski, K.; Belarbi, K.; Buee, L. Neurotrophic factors in Alzheimer’s disease: Role of axonal transport. Genes Brain Behav 2008, 7, 43–56. [Google Scholar]
- Zhang, Y.W.; Xu, H. Molecular and cellular mechanisms for Alzheimer’s disease: Understanding APP metabolism. Curr. Mol. Med 2007, 7, 687–96. [Google Scholar]
- Zuccato, C.; Cattaneo, E. Role of brain-derived neurotrophic factor in Huntington’s disease. Progr. Neurobiol 2007, 81, 294–330. [Google Scholar]
- Wright, B.L.; Barker, R.A. Established and emerging therapies for Huntington’s disease. Curr. Mol. Med 2007, 7, 579–587. [Google Scholar]
- Fumagalli, F.; Racagni, G.; Riva, M.A. Shedding light into the role of BDNF in the pharmacotherapy of Parkinson’s disease. Pharmacogenomics J 2006, 6, 95–104. [Google Scholar]
- Ogier, M.; Wang, H.; Hong, E.; Wang, Q.; Greenberg, M.E.; Katz, D.M. Brain-derived neurotrophic factor expression and respiratory function improve after ampakine treatment in a mouse model of Rett syndrome. J. Neurosci 2007, 27, 10912–10917. [Google Scholar]
- Griesbach, G.S.; Sutton, R.L.; Hovda, D.A.; Ying, Z.; Gomez-Pinilla, F. Controlled contusion injury alters molecular systems associated with cognitive performance. J. Neurosci. Res 2009, 87, 795–805. [Google Scholar]
- Mattson, M.P.; Magnus, T. Ageing and neuronal vulnerability. Nat. Rev. Neurosci 2006, 7, 278–294. [Google Scholar]
- Thoenen, H.; Sendtner, M. Neurotrophins: From enthusiastic expectations through sobering experiences to rational therapeutic approaches. Nat. Neurosci 2002, 5, 1046–1050. [Google Scholar]
- Zhang, Y.H.; Chi, X.X.; Nicol, G.D. Brain-derived neurotrophic factor enhances the excitability of rat sensory neurons through activation of the p75 neurotrophin receptor and the sphingomyelin pathway. J. Physiol 2008, 586, 3113–3127. [Google Scholar]
- Poduslo, J.F.; Curran, G.L. Permeability at the blood-brain and blood-nerve barriers of the neurotrophic factors: NGF, CNTF, NT-3, BDNF. Brain Res. Mol. Brain Res 1996, 36, 280–286. [Google Scholar]
- Morse, J.K.; Wiegand, S.J.; Anderson, K.; You, Y.; Cai, N.; Carnahan, J.; Miller, J.; DiStefano, P.S.; Altar, C.A.; Lindsay, R.M.; et al. Brain-derived neurotrophic factor (BDNF) prevents the degeneration of medial septal cholinergic neurons following fimbria transection. J. Neurosci 1993, 13, 4146–4156. [Google Scholar]
- Zuccato, C.; Marullo, M.; Vitali, B.; Tarditi, A.; Mariotti, C.; Valenza, M.; Lahiri, N.; Wild, E.J.; Sassone, J.; Ciammola, A.; et al. Brain-derived neurotrophic factor in patients with Huntington’s disease. PLoS One 2011, 6, e22966. [Google Scholar]
- Fletcher, J.M.; Hughes, R.A. Novel monocyclic and bicyclic loop mimetics of brain-derived neurotrophic factor. J. Pept. Sci 2006, 12, 515–524. [Google Scholar]
- Gupta, V.K.; You, Y.; Li, J.C.; Klistorner, A.; Graham, S.L. Protective effects of 7,8-dihydroxyflavone on retinal ganglion and RGC-5 cells against excitotoxic and oxidative stress. J. Mol. Neurosci 2012, 49, 96–104. [Google Scholar]
- Zeng, Y.; Liu, Y.; Wu, M.; Liu, J.; Hu, Q. Activation of trkb by 7,8-dihydroxyflavone prevents fear memory defects and facilitates amygdalar synaptic plasticity in aging. J. Alzheimers Dis 2012, 31, 765–778. [Google Scholar]
- Mantilla, C.B.; Ermilov, L.G. The novel TrkB receptor agonist 7,8-dihydroxyflavone enhances neuromuscular transmission. Muscle Nerve 2012, 45, 274–276. [Google Scholar]
- Johnson, R.A.; Lam, M.; Punzo, A.M.; Li, H.; Lin, B.R.; Ye, K.; Mitchell, G.S.; Chang, Q. 7,8-dihydroxyflavone exhibits therapeutic efficacy in a mouse model of Rett syndrome. J. Appl. Physiol 2012, 112, 704–710. [Google Scholar]
- Liu, X.; Chan, C.B.; Jang, S.W.; Pradoldej, S.; Huang, J.; He, K.; Phun, L.H.; France, S.; Xiao, G.; Jia, Y.; et al. A synthetic 7,8-dihydroxyflavone derivative promotes neurogenesis and exhibits potent antidepressant effect. J. Med. Chem 2010, 53, 8274–8286. [Google Scholar]
- Devi, L.; Ohno, M. 7,8-dihydroxyflavone, a small-molecule TrkB agonist, reverses memory deficits and BACE1 elevation in a mouse model of Alzheimer’s disease. Neuropsychopharmacology 2012, 37, 434–444. [Google Scholar]
- Fumagalli, F.; Racagni, G.; Riva, M.A. The expanding role of BDNF: A therapeutic target for Alzheimer’s disease? Pharmacogenomics J 2006, 6, 8–15. [Google Scholar]
- Autio, H.; Matlik, K.; Rantamaki, T.; Lindemann, L.; Hoener, M.C.; Chao, M.; Arumae, U.; Castren, E. Acetylcholinesterase inhibitors rapidly activate Trk neurotrophin receptors in the mouse hippocampus. Neuropharmacology 2011, 61, 1291–1296. [Google Scholar]
- Gupta, V.B.; Anitha, S.; Hegde, M.L.; Zecca, L.; Garruto, R.M.; Ravid, R.; Shankar, S.K.; Stein, R.; Shanmugavelu, P.; Jagannatha Rao, K.S. Aluminium in Alzheimer’s disease: Are we still at a crossroad? Cell Mol. Life Sci 2005, 62, 143–158. [Google Scholar]
- Gupta, V.B.; Indi, S.S.; Rao, K.S. Studies on the role of amino acid stereospecificity in amyloid beta aggregation. J. Mol. Neurosci 2008, 34, 35–43. [Google Scholar]
- Arancibia, S.; Silhol, M.; Mouliere, F.; Meffre, J.; Hollinger, I.; Maurice, T.; Tapia-Arancibia, L. Protective effect of BDNF against beta-amyloid induced neurotoxicity in vitro and in vivo in rats. Neurobiol. Dis 2008, 31, 316–326. [Google Scholar]
- Jang, S.W.; Liu, X.; Chan, C.B.; France, S.A.; Sayeed, I.; Tang, W.; Lin, X.; Xiao, G.; Andero, R.; Chang, Q.; et al. Deoxygedunin, a natural product with potent neurotrophic activity in mice. PLoS One 2010, 5, e11528. [Google Scholar]
- Fletcher, J.M.; Morton, C.J.; Zwar, R.A.; Murray, S.S.; O’Leary, P.D.; Hughes, R.A. Design of a conformationally defined and proteolytically stable circular mimetic of brain-derived neurotrophic factor. J. Biol. Chem 2008, 283, 33375–33383. [Google Scholar]
- Khursigara, G.; Bertin, J.; Yano, H.; Moffett, H.; DiStefano, P.S.; Chao, M.V. A prosurvival function for the p75 receptor death domain mediated via the caspase recruitment domain receptor-interacting protein 2. J. Neurosci 2001, 21, 5854–5863. [Google Scholar]
- Bai, Y.; Xu, J.; Brahimi, F.; Zhuo, Y.; Sarunic, M.V.; Saragovi, H.U. An agonistic TrkB mAb causes sustained TrkB activation, delays RGC death, and protects the retinal structure in optic nerve axotomy and in glaucoma. Invest. Ophth. Vis. Sci 2010, 51, 4722–4731. [Google Scholar]
- Soto, I.; Rosenthal, J.J.; Blagburn, J.M.; Blanco, R.E. Fibroblast growth factor 2 applied to the optic nerve after axotomy up-regulates BDNF and TrkB in ganglion cells by activating the ERK and PKA signaling pathways. J. Neurochem 2006, 96, 82–96. [Google Scholar]
- Chen, H.; Weber, A.J. BDNF enhances retinal ganglion cell survival in cats with optic nerve damage. Invest. Ophth. Vis. Sci 2001, 42, 966–974. [Google Scholar]
- Birnbaum, S.; Chen, J.; Kwon, C.H.; Kernie, S.G.; Bassel-Duby, R.; Parada, L.F. TrkB regulates hippocampal neurogenesis and governs sensitivity to antidepressive treatment. Neuron 2008, 59, 399–412. [Google Scholar]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nature 2008, 455, 894–902. [Google Scholar]
- Shen, J.; Ghai, K.; Sompol, P.; Liu, X.; Cao, X.; Iuvone, P.M.; Ye, K. N-acetyl serotonin derivatives as potent neuroprotectants for retinas. Proc. Natl. Acad. Sci. USA 2012, 109, 3540–3545. [Google Scholar]
- Rantamaki, T.; Vesa, L.; Antila, H.; di Lieto, A.; Tammela, P.; Schmitt, A.; Lesch, K.P.; Rios, M.; Castren, E. Antidepressant drugs transactivate TrkB neurotrophin receptors in the adult rodent brain independently of BDNF and monoamine transporter blockade. PLoS One 2011, 6, e20567. [Google Scholar]
- Hashimoto, R.; Takei, N.; Shimazu, K.; Christ, L.; Lu, B.; Chuang, D.M. Lithium induces brain-derived neurotrophic factor and activates TrkB in rodent cortical neurons: An essential step for neuroprotection against glutamate excitotoxicity. Neuropharmacology 2002, 43, 1173–1179. [Google Scholar]
- Furmaga, H.; Carreno, F.R.; Frazer, A. Vagal nerve stimulation rapidly activates brain-derived neurotrophic factor receptor TrkB in rat brain. PLoS One 2012, 7, e34844. [Google Scholar]
- Al-Majed, A.A.; Brushart, T.M.; Gordon, T. Electrical stimulation accelerates and increases expression of BDNF and trkB mRNA in regenerating rat femoral motoneurons. Eur. J. Neurosci 2000, 12, 4381–4390. [Google Scholar]
- Tsao, D.; Thomsen, H.K.; Chou, J.; Stratton, J.; Hagen, M.; Loo, C.; Garcia, C.; Sloane, D.L.; Rosenthal, A.; Lin, J.C. TrkB agonists ameliorate obesity and associated metabolic conditions in mice. Endocrinology 2008, 149, 1038–1048. [Google Scholar]
- Brodeur, G.M.; Minturn, J.E.; Ho, R.; Simpson, A.M.; Iyer, R.; Varela, C.R.; Light, J.E.; Kolla, V.; Evans, A.E. Trk receptor expression and inhibition in neuroblastomas. Clin. Cancer Res 2009, 15, 3244–3250. [Google Scholar]
- Kawamura, N.; Kawamura, K.; Manabe, M.; Tanaka, T. Inhibition of brain-derived neurotrophic factor/tyrosine kinase B signaling suppresses choriocarcinoma cell growth. Endocrinology 2010, 151, 3006–3014. [Google Scholar]
- Ng, Y.K.; Wong, E.Y.; Lau, C.P.; Chan, J.P.; Wong, S.C.; Chan, A.S.; Kwan, M.P.; Tsao, S.W.; Tsang, C.M.; Lai, P.B.; et al. K252a induces anoikis-sensitization with suppression of cellular migration in Epstein-Barr virus (EBV)—Associated nasopharyngeal carcinoma cells. Invest. New Drugs 2012, 30, 48–58. [Google Scholar]
- Yalta, K.; Yilmaz, M.B.; Turgut, O.O.; Tandogan, I. Markers of inflammation and thrombin generation: Additional guides in determining the therapeutic strategy for malign ventricular arrhythmias after an acute myocardial infarction? Int. J. Cardiol 2010, 145, 492–493. [Google Scholar]
- Ho, R.; Minturn, J.E.; Simpson, A.M.; Iyer, R.; Light, J.E.; Evans, A.E.; Brodeur, G.M. The effect of P75 on Trk receptors in neuroblastomas. Cancer Lett 2011, 305, 76–85. [Google Scholar]
- Dubreuil, C.I.; Winton, M.J.; McKerracher, L. Rho activation patterns after spinal cord injury and the role of activated Rho in apoptosis in the central nervous system. J. Cell Biol 2003, 162, 233–243. [Google Scholar]
- Fu, Q.L.; Hu, B.; Wu, W.T.; Pepinsky, R.B.; Mi, S.; So, K.F. Blocking LINGO-1 function promotes retinal ganglion cell survival following ocular hypertension and optic nerve transection. Invest. Ophth. Vis. Sci 2008, 49, 975–985. [Google Scholar]
- Stranahan, A.M.; Arumugam, T.V.; Mattson, M.P. Lowering corticosterone levels reinstates hippocampal brain-derived neurotropic factor and Trkb expression without influencing deficits in hypothalamic brain-derived neurotropic factor expression in leptin receptor-deficient mice. Neuroendocrinology 2011, 93, 58–64. [Google Scholar]
- Zhang, L.; Hu, Y.; Sun, C.Y.; Li, J.; Guo, T.; Huang, J.; Chu, Z.B. Lentiviral shRNA silencing of BDNF inhibits in vivo multiple myeloma growth and angiogenesis via down-regulated stroma-derived VEGF expression in the bone marrow milieu. Cancer Sci 2010, 101, 1117–1124. [Google Scholar]
- Shin, J.; Kim, J.; Yu, Y.; Heo, J. Evaluation of change of neurotrophin receptors (TrkA, TrkB, and TrkC) in the retina after intravitreal injection of anti-vegf antibody. ARVO Meet. Abstr 2010, 51, 721. [Google Scholar]
- Folkman, J. Fundamental concepts of the angiogenic process. Curr. Mol. Med 2003, 3, 643–651. [Google Scholar]
- Sondell, M.; Sundler, F.; Kanje, M. Vascular endothelial growth factor is a neurotrophic factor which stimulates axonal outgrowth through the flk-1 receptor. Eur. J. Neurosci 2000, 12, 4243–4254. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gupta, V.K.; You, Y.; Gupta, V.B.; Klistorner, A.; Graham, S.L. TrkB Receptor Signalling: Implications in Neurodegenerative, Psychiatric and Proliferative Disorders. Int. J. Mol. Sci. 2013, 14, 10122-10142. https://doi.org/10.3390/ijms140510122
Gupta VK, You Y, Gupta VB, Klistorner A, Graham SL. TrkB Receptor Signalling: Implications in Neurodegenerative, Psychiatric and Proliferative Disorders. International Journal of Molecular Sciences. 2013; 14(5):10122-10142. https://doi.org/10.3390/ijms140510122
Chicago/Turabian StyleGupta, Vivek K., Yuyi You, Veer Bala Gupta, Alexander Klistorner, and Stuart L. Graham. 2013. "TrkB Receptor Signalling: Implications in Neurodegenerative, Psychiatric and Proliferative Disorders" International Journal of Molecular Sciences 14, no. 5: 10122-10142. https://doi.org/10.3390/ijms140510122
APA StyleGupta, V. K., You, Y., Gupta, V. B., Klistorner, A., & Graham, S. L. (2013). TrkB Receptor Signalling: Implications in Neurodegenerative, Psychiatric and Proliferative Disorders. International Journal of Molecular Sciences, 14(5), 10122-10142. https://doi.org/10.3390/ijms140510122