Abstract
microRNAs (miRNAs) cause mRNA degradation or translation suppression of their target genes. Previous studies have found direct involvement of miRNAs in cancer initiation and progression. Artificial miRNAs, designed to target single or multiple genes of interest, provide a new therapeutic strategy for cancer. This study investigates the anti-tumor effect of a novel artificial miRNA, miR P-27-5p, on breast cancer. In this study, we reveal that miR P-27-5p downregulates the differential gene expressions associated with the protein modification process and regulation of cell cycle in T-47D cells. Introduction of this novel artificial miRNA, miR P-27-5p, into breast cell lines inhibits cell proliferation and induces the first “gap” phase (G1) cell cycle arrest in cancer cell lines but does not affect normal breast cells. We further show that miR P-27-5p targets the 3′-untranslated mRNA region (3′-UTR) of cyclin-dependent kinase 4 (CDK4) and reduces both the mRNA and protein level of CDK4, which in turn, interferes with phosphorylation of the retinoblastoma protein (RB1). Overall, our data suggest that the effects of miR p-27-5p on cell proliferation and G1 cell cycle arrest are through the downregulation of CDK4 and the suppression of RB1 phosphorylation. This study opens avenues for future therapies targeting breast cancer.
1. Introduction
microRNAs (miRNAs) cause mRNA degradation or translation suppression of their target genes. Previous studies have shown that miRNAs play crucial roles in tumorigenesis by targeting the mRNAs of oncogenes or tumor suppressors [1,2]. Many reports indicate that miRNA expression is altered in tumor tissues, suggesting that these miRNAs could be potential markers for detection and prognosis in human cancers [3,4]. Because of the critical role of miRNAs as regulators of cell fate, analyzing and manipulating miRNAs within cancer cells may provide powerful avenues for diagnosis, prognosis, drug discovery, and therapeutics [5]. miRNA P-27-5p (miR P-27-5p) is a novel miRNA we recently discovered in breast cancer cells [6]. In this study, we investigate why this artificial miRNA can suppress breast cancer cell growth.
Microarray is a powerful high-throughput technique for determining changes in global gene expressions in functional genomics [7,8]. It can measure expression of tens of thousands of discrete sequences in a single array [9]. The exon array is useful for exon-level expression profiling at the whole-genome scale on a single array [10]. The oligonucleotide probes of exon arrays greatly differ from those of conventional 3′ expression arrays in their design, density, and coverage. This technique has been used for novel genes discoveries [11], gene function determination, drug evaluation, pathway dissection, and clinical sample classification [9,12].
The G1/S cell cycle checkpoint controls the passage of eukaryotic cells from G1 into the DNA synthesis phase (S). Two cell cycle kinases, cyclin-dependent kinase 4/6 (CDK4/6)-cyclin D [13–15] and CDK2-cyclin E [16], and the transcription complex that includes retinoblastoma protein (RB1) and E2F are essential in controlling this checkpoint [17]. CDK4, a member of the Ser/Thr protein kinase family, binds cyclin D and subsequently phosphorylates RB1, leading to cell cycle regulation, which is thought to occur through hyperphosphorylation-induced release of the E2F transcription factors from the large pocket [17]. A breakdown in the regulation of this cycle can lead to out-of-control growth and contribute to tumor formation. Recently, many inhibitors specifically targeted to CDK4 have been developed for the treatment of breast cancer [14,15].
In this study, we show that miR P-27-5p inhibits the growth of breast cancer cells and induces cell cycle arrest at the G1 phase. Exon array, Western blot, real-time polymerase chain reaction (PCR), and luciferase report analyses have revealed that miR P-27-5p targets CDK4. Our observations indicate that miR P-27-5p inhibits cancer cell proliferation and triggers G1 cell cycle arrest by targeting CDK4 and suppressing phosphorylation of RB1.
2. Results and Discussion
2.1. miR P-27-5p Downregulates Gene Expressions Associated with Cell Growth, Cell Cycle, and Phosphorylation in T-47D Cells
In addition to translation suppression, miRNAs cause mRNA degradation of their target genes; the changes at the mRNA level can be detected by microarray experiments [18,19]. We used a high-throughput exon array to identify the miR P-27-5p–regulated genes and potential targets. The results of exon array data have been submitted to the GEO database, and the series record is GSE28657.
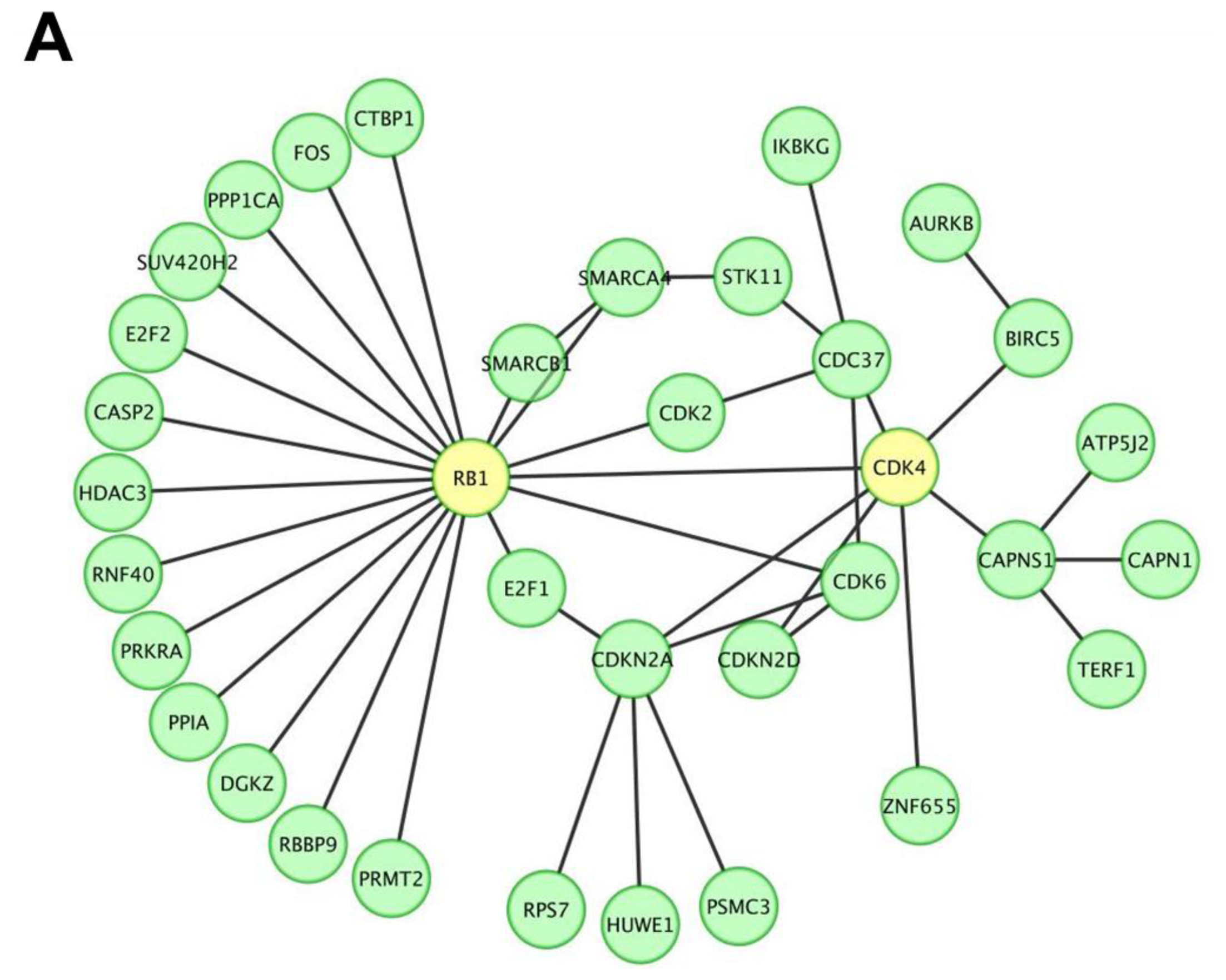
To screen and investigate the gene expression profile and possible biological functions of miR P-27-5p regulation in breast cancer T-47D cells, we used our previously developed approach [20,21]. There were 2590 genes with a significant downregulated change in expression level between control and miR P-27-5p–transfected T-47D cells (Supporting Information Table S.1). These downregulated expressed genes were used to construct the protein-protein interaction (PPI) network. The human protein interaction network (PIN) was downloaded from the Human Protein Reference Database [22], and only the largest connected component was studied. The differentially downregulated expressed proteins in the miR P-27-5p–related network was further predicted by functional enrichment analysis. Among the PPI networks, we found that CDK4 and RB1 were involved in the network and the RB1 interacted with 19 proteins including CDK4 (Figure 1A).
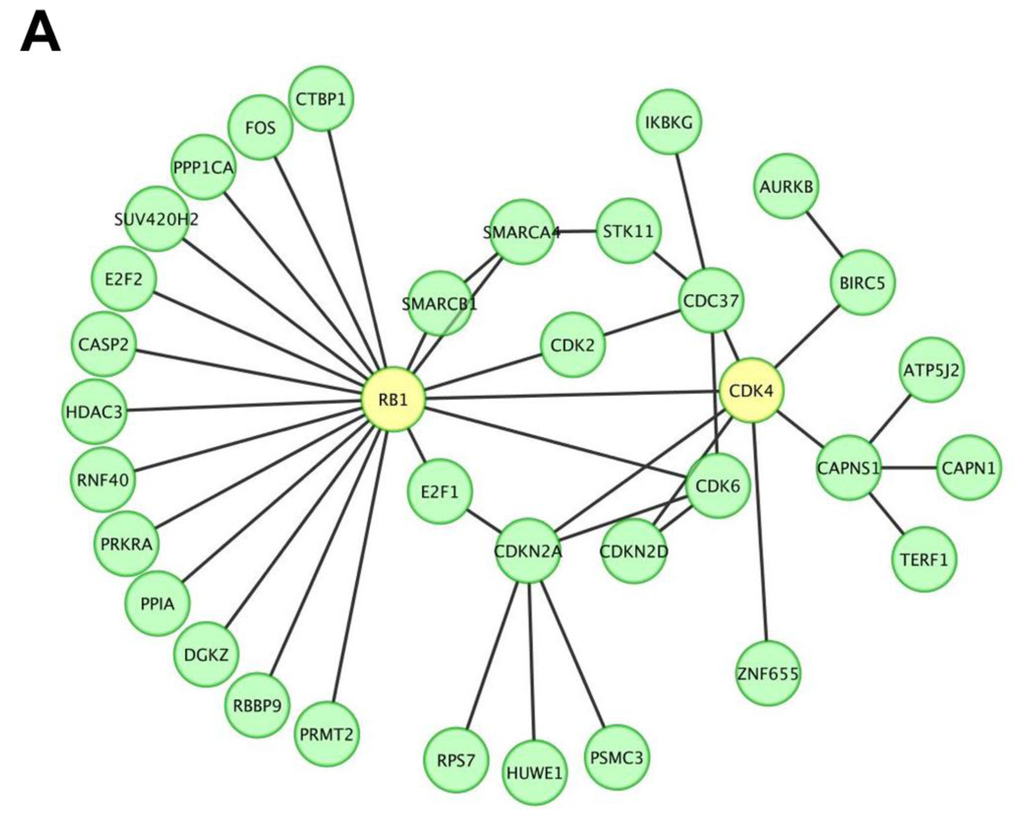
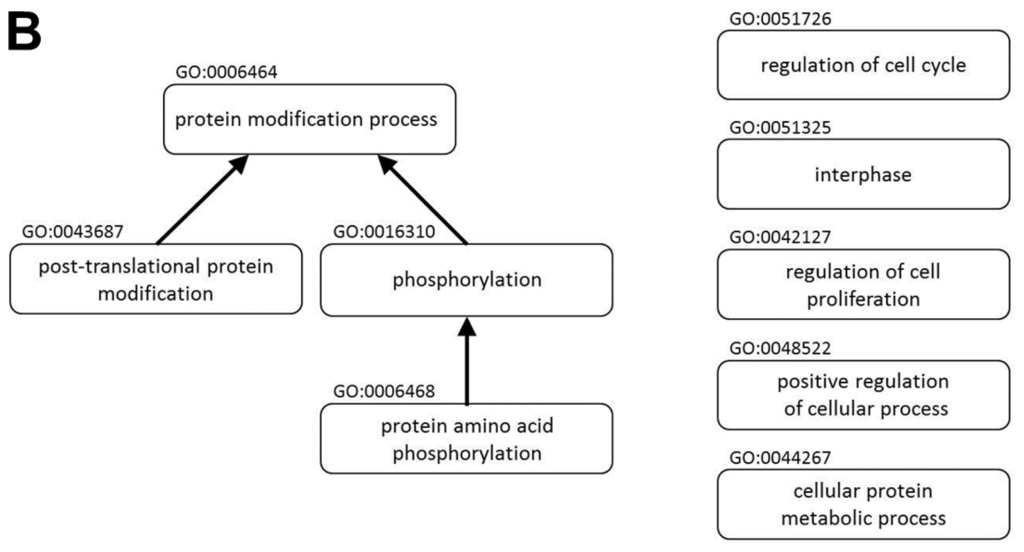
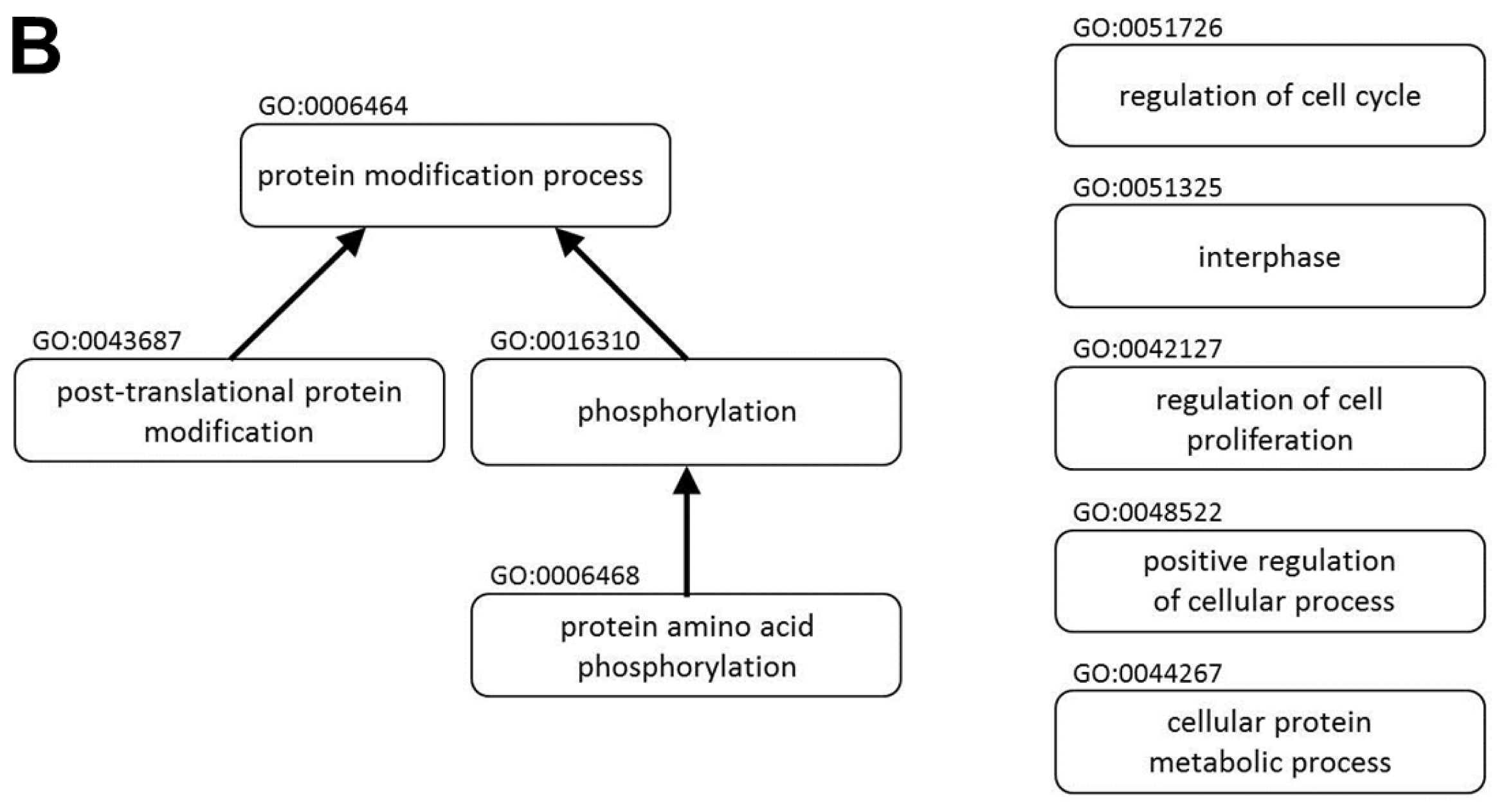
Figure 1.
A protein-protein interaction network and the biological functions regulated by miR P-27-5p in T-47D cells. Gene expression profiles were determined using exon arrays. (A) The significantly differentially expressed proteins in miR P-27-5p–overexpressing tumor cells were used to construct the protein-protein interaction (PPI) network; (B) All proteins in the network were further analyzed for clustering of functional profiles by using BiNGO. It uncovered key functional relationships, particularly cell proliferation and cycle and phosphorylation.
All proteins in the network were further analyzed for clustering of functional profiles using BiNGO (p < 0.001). BiNGO [23,24], a Cytoscape [24] plug-in, was used to determine which Gene Ontology (GO) terms were significantly overrepresented (the hypergeometric test, Benjamini and Hochberg False Discovery Rate correction, p ≤ 0.001) in miR P-27-5p–related networks. Key functional relationships were revealed, including the protein modification process, protein amino acid phosphorylation, phosphorylation, regulation of cell proliferation, posttranslational protein modification, the cellular protein metabolic process, positive regulation of cellular process, interphase, and regulation of cell cycle (Figure 1B and Table 1).

Table 1.
The functions and genes regulated by miR P-27-5p.
2.2. miR P-27-5p Overexpression Inhibits the Growth of Breast Cancer Cells
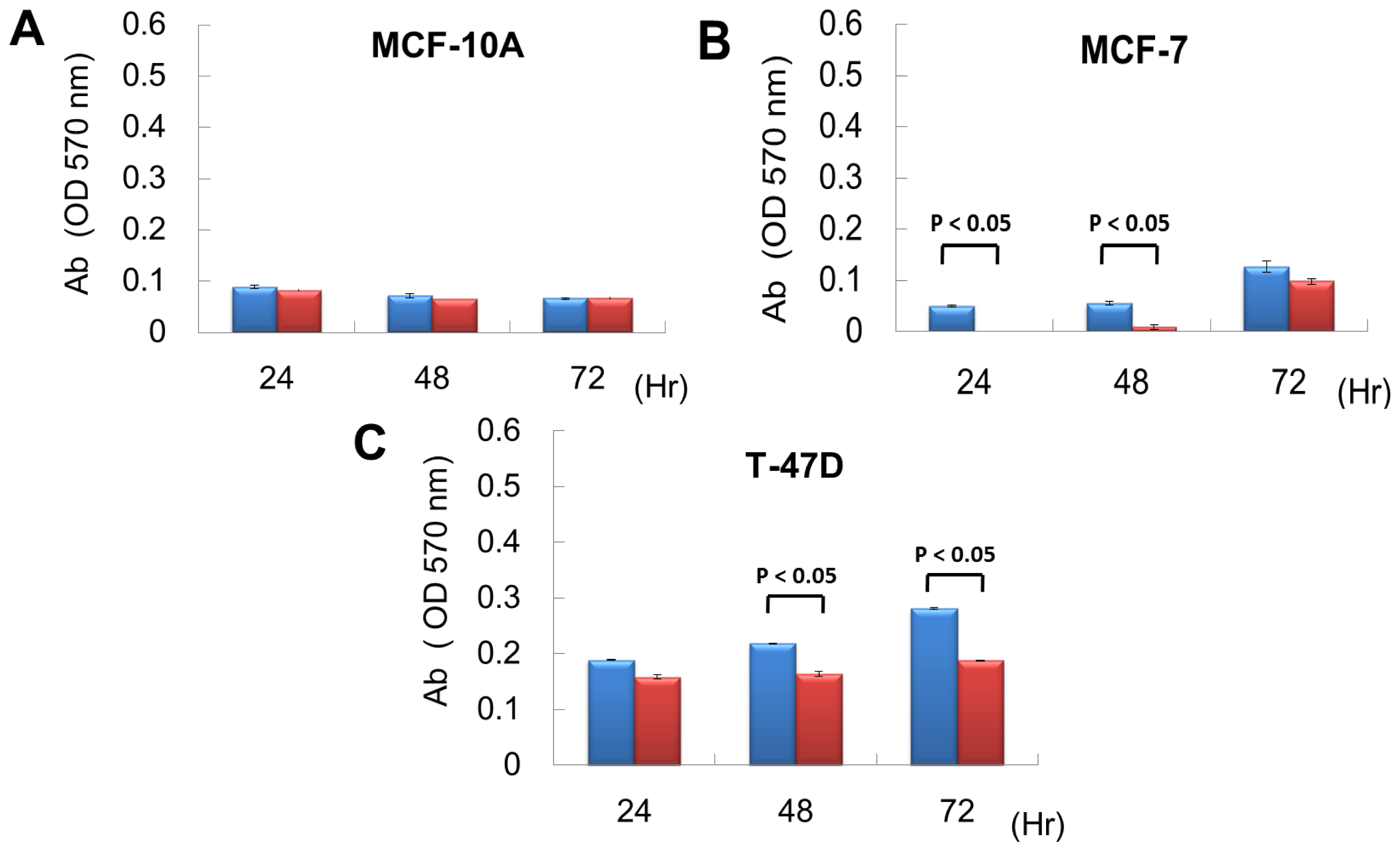
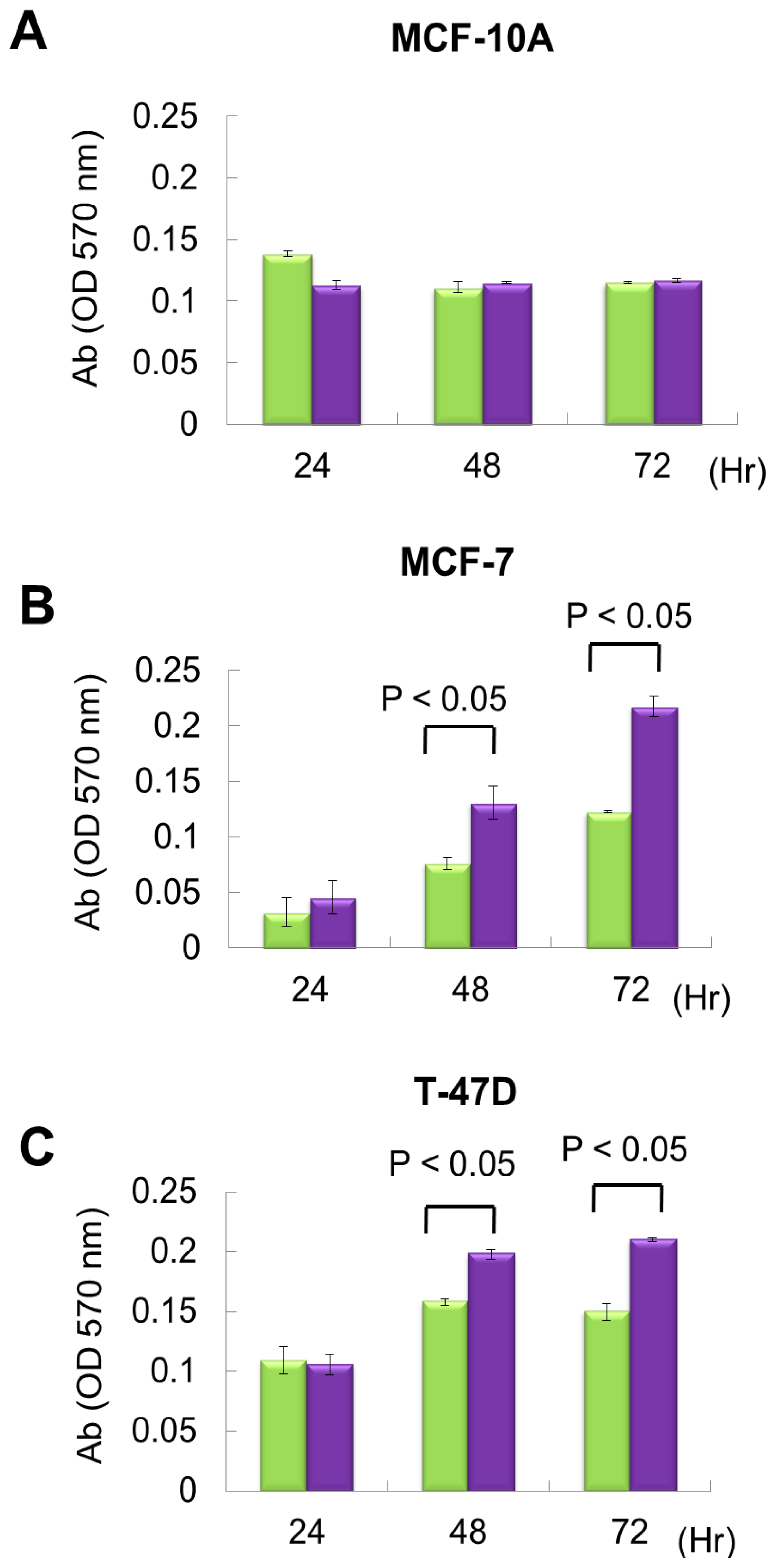
From exon array data, network and functional enrichment analysis, we found that miR P-27-5p overexpression could downregulate the gene expression levels involved in cell proliferation. To evaluate the effect of miR P-27-5p on cell growth, a methylthiazoletetrazolium (MTT) cell proliferation assay was used as described above. MCF-10A, MCF-7 and T-47D were transfected with miR P-27-5p mimics or negative control (NC). MTT assays were performed at 24 h, 48 h, and 72 h. As shown in Figure 2, transfection of miR P-27-5p into cell lines significantly inhibited cell proliferation of breast cancer cells MCF-7 and T-47D, but not the normal cells MCF-10A. Additionally, we found that transfection of antisense miR P-27-5p into cancer cells promoted cell proliferation (Figure 3). These results indicate that miR P-27-5p has an adverse effect on breast cancer cell proliferation.
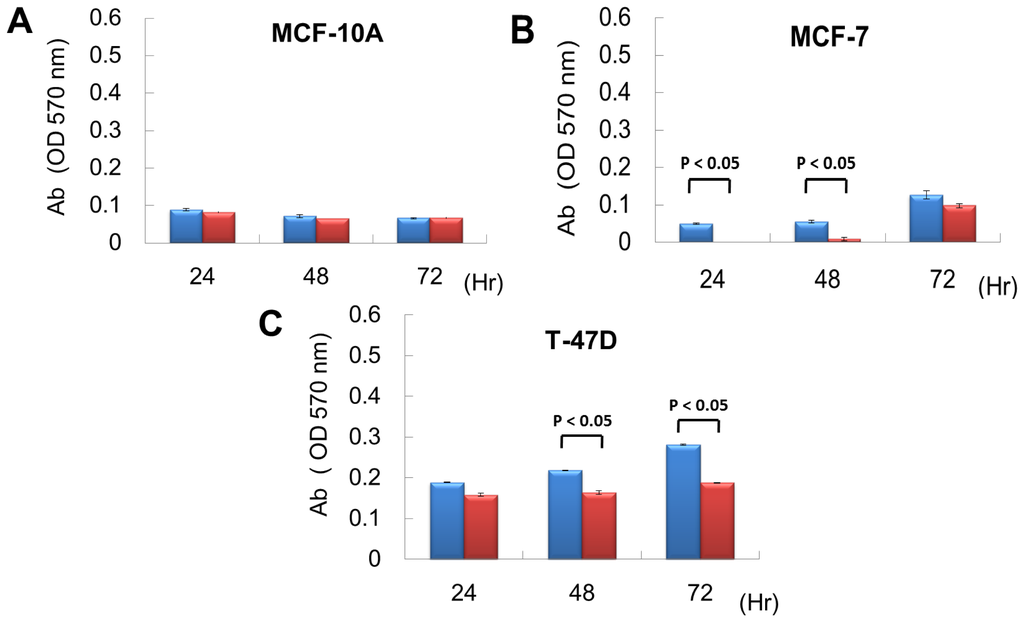
Figure 2.
Inhibitory effect of miR P-27-5p on cell proliferation. The breast normal cells MCF-10A (A) and cancer cells MCF-7 (B) and T-47D (C) were transfected with 60 nM miR P-27-5p (red bar) and NC mimetics (blue bar). Cell viability was determined using an MTT assay. The relative cell proliferation was examined at the indicated time points by MTT. The absorbance of MTT by each sample was recorded at 570 nm after staining. The error bar shows SE for three independent experiments.
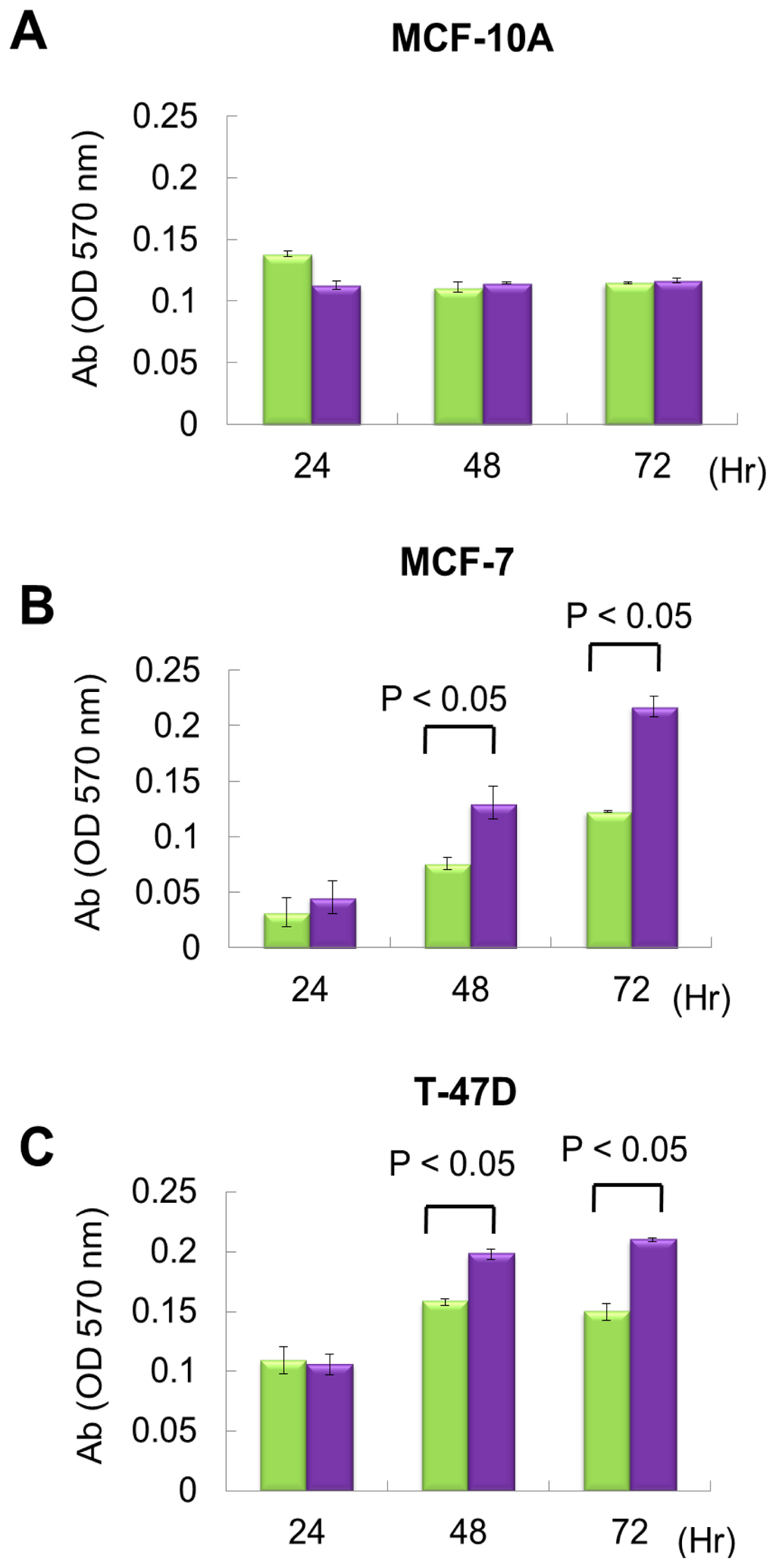
Figure 3.
The effect of downregulation of miR P-27-5p on cell proliferation. The breast normal cells MCF-10A (A) and cancer cells MCF-7 (B) and T-47D (C) were transfected with 60 nM antisense miR P-27-5p (purple bar) and NC (green bar), respectively. Cell viability was determined using an MTT assay. The relative cell proliferation was examined at the indicated time points by methylthiazoletetrazolium (MTT). The absorbance of MTT by each sample was recorded at 570 nm after staining. The error bar shows SE for three independent experiments.
2.3. miR P-27-5p Induces G1 Arrest in Breast Cancer Cells
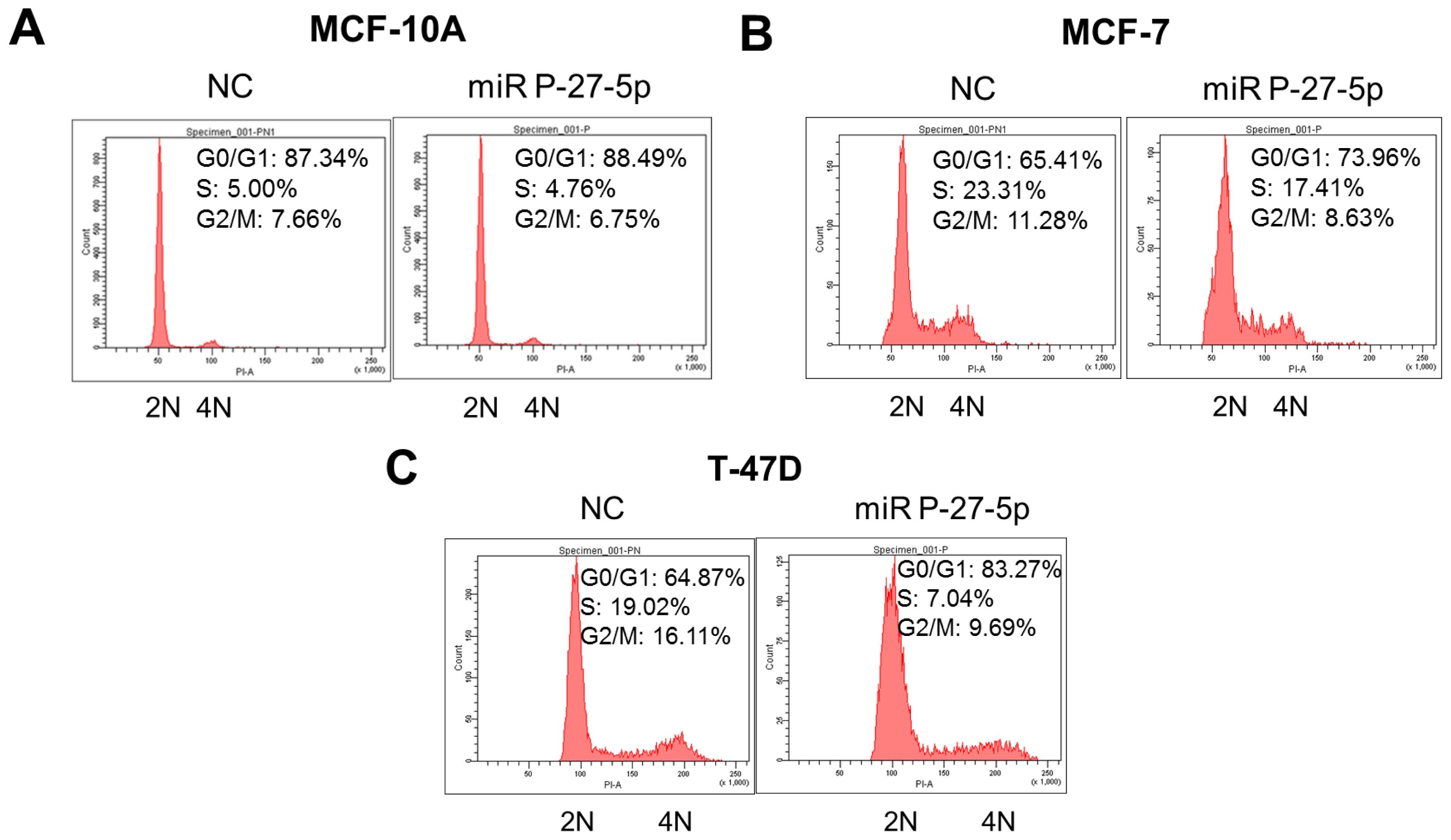
In addition to cell proliferation, miR P-27-5p downregulates the genes involved in the cell cycle. To investigate the effect of miR P-27-5p on cell cycle progression of breast normal/cancer cells, their DNA contents were analyzed by flow cytometry, and the derived data were used to investigate the phase distribution of the cell cycle. Cells with 2 n and 4 n DNA content correspond to the G0/G1 and G2/M phases, respectively. Cells with a DNA content between 2 n and 4 n correspond to the S phase. As shown in Figure 4, after transfection with miR P-27-5p, the DNA percentage of the G0/G1 phase was increased from 65.41% to 73.96% in MCF-7 cells and from 64.87% to 83.27% in T-47D cells, but no significant change in MCF-10A cells was noted. These results show that miR P-27-5p induces G1 cell cycle arrest in breast cancer cells but not in normal cells.
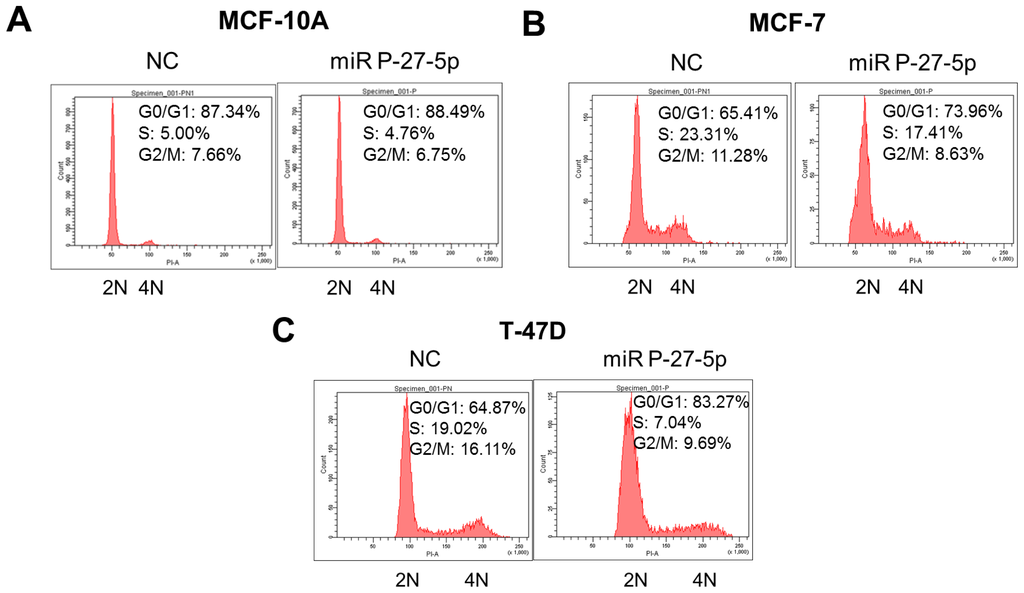
Figure 4.
G0/G1 arrest by miR P-27-5p. The breast normal cells (MCF-10A) (A), and cancer cells MCF-7 (B) and T-47D (C) were transfected with miR P-27-5p and NC mimetics (100 nM) for 48 h. Then, cell cycle distributions of these cells were analyzed by flow cytometry. 2N: cells with diploid DNA content; and 4N: cells with tetraploid DNA content. The experiments were performed in triplicates.
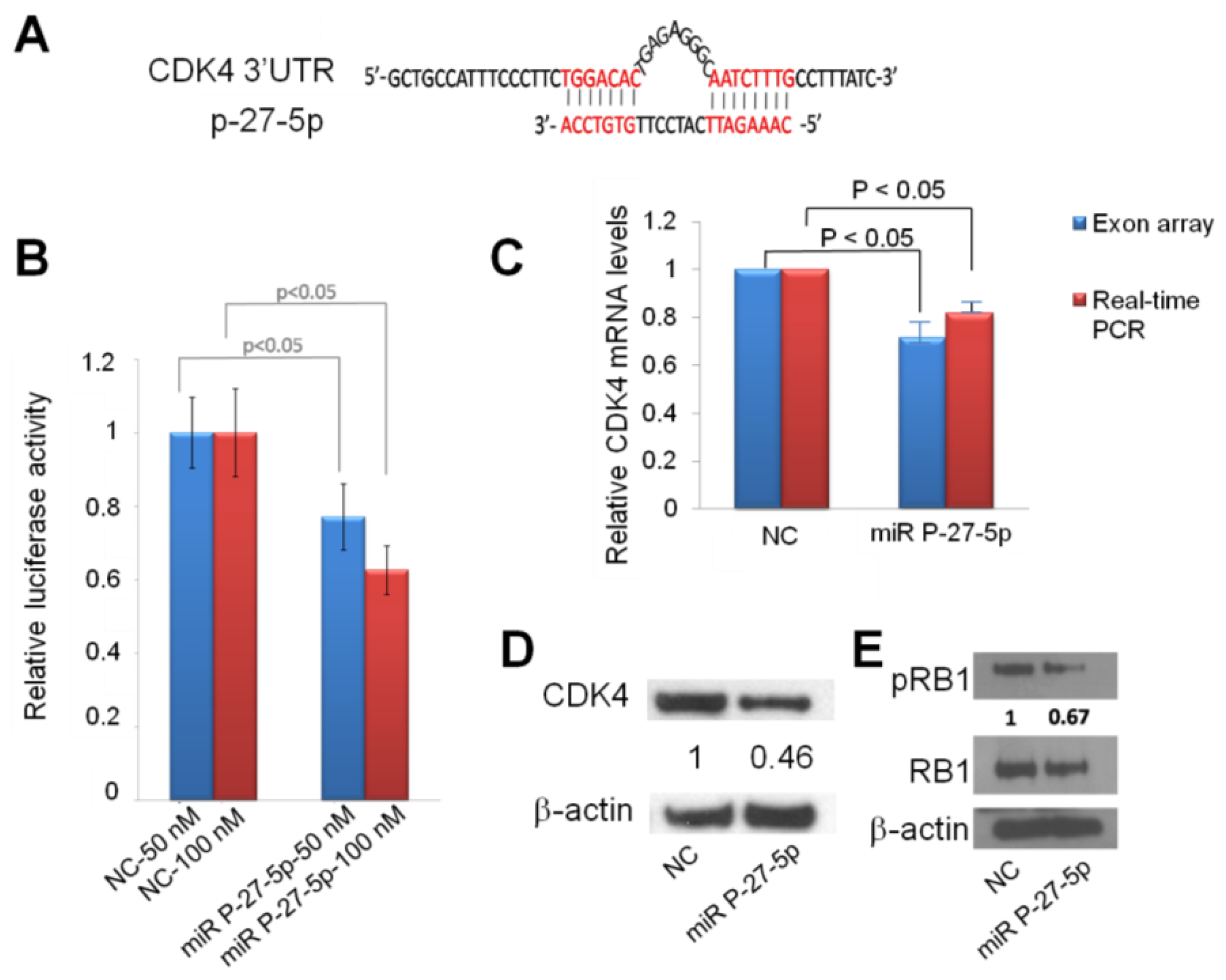
2.4. Down-Regulation of CDK4 by miR P-27-5p at mRNA and Protein Levels via Direct Targeting 3′-UTR
From exon array and network analysis, we found CDK4 is important in miR P-27-5p–regulated function. We used the sequence alignment method to examine whether CDK4 is a potential target of miR P-27-5p. Figure 5A shows that the sequence of miR P-27-5p is complimentary to the 3′-UTR region of CDK4. To investigate whether miR P-27-5p directly recognizes the 3′-UTR of CDK4, we cloned the fragment containing the presumed target site into 3′-UTR of the luciferase gene. Figure 5B shows that transfection with miR P-27-5p resulted in a significant decrease in luciferase activity compared to that in NC-transfected cells. To validate CDK4 as a target of miR P-27-5p, downregulation of CDK4 at the mRNA and protein level was examined by exon array, real-time PCR, and Western blot analyses (Figure 5C,D). CDK4 mRNA and protein in miR P-27-5p–transfected cells were decreased compared to NC-transfected cells. These results suggest that CDK4 is the functional target of miR P-27-5p in T-47D cells and that miR P-27-5p inhibits cell proliferation and causes cell cycle arrest via targeting CDK4.
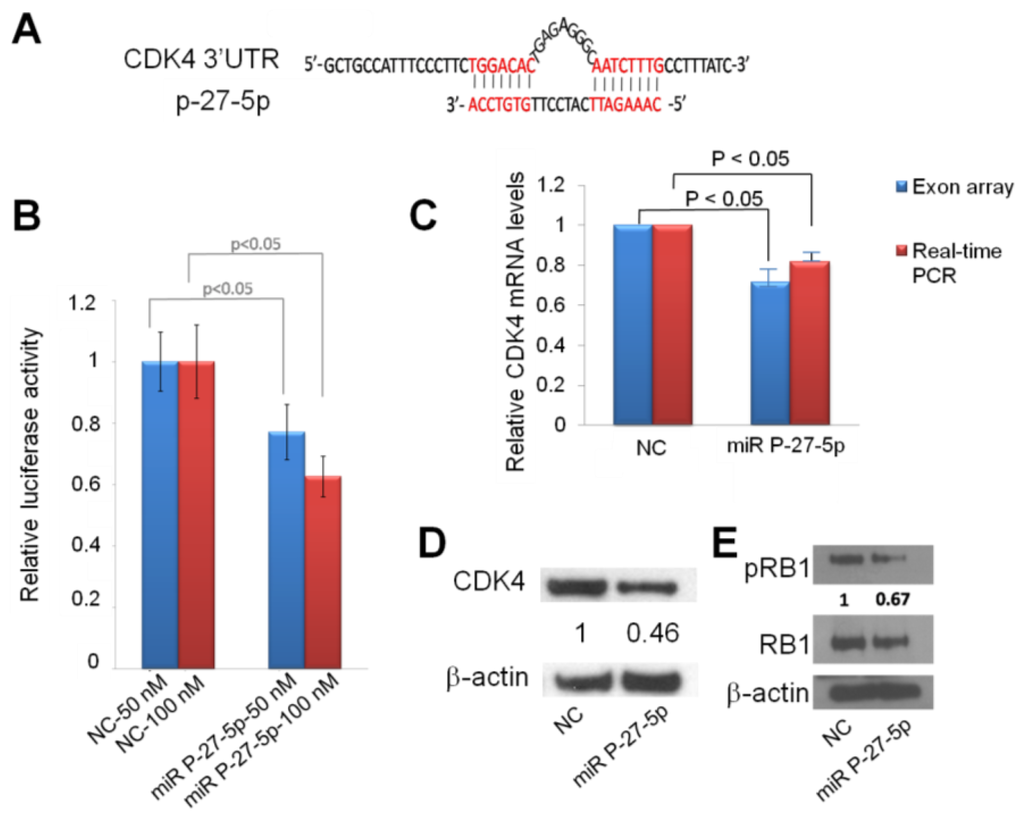
Figure 5.
Down-regulation of CDK4 by miR P-27-5p at the mRNA and protein levels via direct targeting of 3′-UTR. (A) The sequences of CDK4 3′-UTR and miR P-27-5p. (B) The effect of miR 27-5p on the expression of CDK4 by using luciferase assays. Cells were cotransfected with miR P-27-5p duplex or NC with CDK4 3′-UTR. The reporter assays shown in this study were based on data averaged from at least three independent transfections. (C) CDK4 mRNA expression was detected by exon array and real-time PCR at 48 h after transfection with miR P-27-5p or NC mimetics and normalized against that of GADPH. (D) CDK4 protein, (E) Phosphorylated RB1, and RB1 proteins were measured by Western blot at 48 h after transfection with miR P-27-5p or NC mimetics. β-actin was used as the internal control. All the experiments were conducted in triplicates.
2.5. miR P-27-5p Inhibits RB1 Phosphorylation
To investigate the association of the miR P-27-5p–induced G1 arrest with RB1 phosphorylation, Western blot analysis was conducted using an RB1 phosphospecific antibody. The results showed a decrease in RB1 phosphorylation after transfection with miR P-27-5p (Figure 5E). When dephosphorylated, RB1 interacts with E2F transcription factors and prevents transcription of genes required for progression through the cell cycle. By contrast, when phosphorylated by cell cycle-dependent kinases like CDK2 and CDK4, RB1 no longer interacts with E2F and the cell cycle proceeds through the G1/S checkpoint [17]. Loss of cell cycle control is a hallmark of cancer, and aberrations in the cyclin-CDK-RB pathway are common in breast cancer [25]. Therefore, targeted inhibition of CDK4 activity has a role in the treatment of breast cancer [14,26].
2.6. Discussion
Our previous report suggested that miR P-27-5p may be an alternative mature form of miR-802 [6]. Our unpublished data indicate that miR P-27-5p is expressed higher in MCF-10A than MCF-7 and T47D tumor cells. However, previous study reported endogenous miR-802 was not detected in breast cancer patients based on miRNA expression profiles [27]. On the other hand, miR-802 was investigated in mice by using expression and sequencing studies [28] and Kuhn et al. used miRNA expression profiling, miRNA RT-PCR and miRNA in situ hybridization experiments to identify miR-802 was up-regulated in fetal brain and heart specimens from individuals with Down syndrome when comparing with control groups [29]. According to these, miR-802 is expressed in other tissues, but not in breast tissues. Thus, the expression levels of miR P-27-5p and miR-802 in breast tissues are different.
Our previous study showed that miR P-27-5p could target Laminin β3 (LAMB3) [6]. LAMB3 has been detected in various cancers and affects tumor progression. For example, promoter demethylation of LAMB3 causes aberrant LAMB3 expression in gastric cancer and tumor cells stably expressing LAMB3 have elevated migration and adhesiveness [30]. Patients with esophageal carcinoma have worse 5-year survival rates [31]. Additionally, LAMB3 overexpression promotes migration of prostate cancer LNCaP cells and tumor growth in mice [32]. Thus, these results indicate that LAMB3 is involved in tumor progression, such as tumor growth and metastasis, and miR P-27-5p may affects tumor progression through regulating LAMB3 expression.
From our exon array, network analysis, and functional enrichment analysis, we predicted that miR P-27-5p may have a function in cell cycle and cell proliferation. With cell viability assay and cell cycle analysis, we further confirmed that miR P-27-5p induces cancer cell cycle arrest at the G1 phase and inhibits cancer cell proliferation. De Guire et al. find that artificial miRNAs reproduce the effects of E2Fs inhibition in both normal human fibroblasts and prostate cancer cells, where they inhibit cell proliferation and induced cellular senescence [33]. Brown and Naldini suggest utilizing artificial miRNA target sites to exploit or inhibit endogenous miRNA regulation for therapeutic and experimental applications [34]. Idogawa et al. efficiently used a single recombinant adenovirus expressing p53 and p21-targeting artificial miRNAs to induce apoptosis in human cancer cells [35]. Collectively, artificial miR P-27-5p may represent a novel therapeutic option in breast cancer treatment.
Previous studies showed that CDK4 regulates progression through the G1/S phase of the cell cycle by binding to cyclin D1 to phosphorylate RB1 and releasing E2F transcription factors for progression through the cell cycle [36,37]. Decreased cyclin D1 and cyclin D1-CDK4/6 kinase activity reduces the invasion and migration potential of MDA-MB-231 breast cancer cells [37]. In this present study, functional enrichment of miR p-27-5p-regulated network showed that CDK4 is involved in many biological functions such as posttranslational protein modification, protein amino acid phosphorylation, phosphorylation, cellular protein metabolic process, positive regulation of cellular process, protein modification process, regulation of cell cycle, regulation of cell proliferation, and interphase (Table 1). We found that CDK4 and RB1 were hubs within this network and RB1 was one of the interacting proteins of CDK4. RB1 is involved in positive regulation of cellular process, regulation of cell cycle, regulation of cell proliferation, and interphase. These results are consistent with the reports described by Prud’homme et al. [36] and Zhong et al. [37]. Furthermore, we identified their expression levels of CDK4 and phosphorylated RB1 were reduced in response to miR p-27-5p over-expression. Thus, these results indicate that miR p-27-5p may play important roles in cell cycle and proliferation by regulating a small subset of genes within its regulated PIN.
In this study, artificial miR P-27-5p targets the 3′-UTR of CDK4 with mismatches to the central region of its targeting site (Figure 5A). Ye et al. designed artificial miRNAs (AmiRs) with mismatches targeting the 3′-UTR of viral genome. These small AmiRs combined with pRNA-folate conjugates could form a promising system for antiviral drug development [38]. Furthermore, we found that miR P-27-5p suppresses the expressions of CDK4, both in mRNA and protein levels (Figure 5C,D), and inhibits RB1 phosphorylation (Figure 5D), which is one of the CDK4-interacting proteins (Figure 1A). As described previously, CDK4 regulates progression through the G1/S phase of the cell cycle by binding to cyclin D1 to phosphorylate RB1 [36,37]. All of the evidence implies that the miR P-27-5p–induced cell cycle and inhibited cell proliferation may be through targeting CDK4 and its interacting protein, RB1.
3. Experimental Section
3.1. Cell Culture and Transfection
We used one human mammary epithelial (normal) cell line (MCF-10A) and three breast cancer cell lines (MCF-7, T-47D) in this study. All the cell lines were purchased from the Bioresource Collection and Research Center, Hinchu, Taiwan. Human breast MCF-10A cells were cultured in DMEM/F12 supplemented with 5% horse serum and MCF-7 and T-47D cells were cultured in DMEM supplemented with 10% FBS and 100 μg/mL penicillin/streptomycin at 37 °C in a humidified atmosphere of 5% CO2. miR P-27-5p mimics and their corresponding NC were obtained from Ambion (Applied Biosystems/Ambion, Austin, TX, USA). Transfection was carried out using Lipofectamine 2000 (Invitrogen, Inc.) according to the manufacturer’s instructions [6].
3.2. Exon Arrays and Data Analysis
Total cellular RNA was extracted with the use of TRIzol reagent (Invitrogen), and purity was confirmed by spectrophotometry (A260/A280 ratio) and capillary electrophoresis (Agilent 2100 Bioanalyzer, Agilent). RNA processing and hybridization onto Affymetrix Human Exon 1.0 ST arrays were performed according to the manufacturer’s protocol. Microarray (n = 2 per group) analysis was performed by dChip software (dChip 2010.01; Cheng Li Lab of Computational Cancer Genomics: Boston, MA, USA, 2010) [39]. Raw data (CEL files) and annotation data were used for computing the signal value of exons with Quantile normalization. The exon signal data were later transformed to the Affymetrix U133 2.0 PLUS probe set by the export function of dChip software to obtain the gene expression value for further analysis. We have submitted the exon array data to the GEO database, and the series record is GSE28657.
3.3. Protein Interaction Network and Functional Enrichment
Among the significantly differentially expressed genes (p < 0.05), the downregulated genes were chosen due to possible miR p-27-5p targets. The protein interaction networks with these downregulated genes were constructed based on human PIN from the Human Protein Reference Database (version 9; Institute of Bioinformatics & Pandey lab: Bangalore, India; Baltimore, MD, USA, 2010). The possible functions of this network were analyzed by GO functional enrichment, which were performed by BiNGO with a threshold of p < 0.001 and a GO level of more than 5 [20]. Enriched functions with the CDK4 gene were selected for further discussion. The method was described previously [20].
3.4. Cell Viability Assay
Cells (2 × 104 cells/mL) were seeded on 24-well plates. After 24 h, the cells were transfected with miR P-27-5p (60 nM). At 24 h, 48 h, or 72 h, MTT (100 μL) was added to each 1 mL of culture medium for 4 h of incubation at 37 °C and measured at 570 nm by ELISA reader. All experiments were performed in triplicates.
3.5. Cell Cycle Analysis
Cells (4 × 105) were transfected with miR P-27-5p or a control vector at a final concentration of 100 nM using Lipofectamine 2000 (Invitrogen, Inc.) for 48 h. After incubation, floating and adherent cells were harvested. Cells were washed carefully with cold PBS at 4 °C. Cell pellets were resuspended in 100 μL of a binding buffer. Annexin-V FITC (0.2 μg/100 μL) and PI (10 μg/mL) were added to the cells and left to incubate in the dark for 15 min at room temperature. All data were obtained using flow cytometry with a FACSCanto cytometer (Becton Dickinson, San Jose, CA, USA). The flow cytometric analysis was performed using FCS Express (version 4; DeNovo software: Los Angeles, CA, USA, 2012). All experiments were performed in triplicates.
3.6. Construction of the CDK4 3′-UTR Report Plasmids and Luciferase Assay
A CDK4 3′-UTR luciferase reporter was created by inserting full-length human CDK4 3′-UTR into the SacI and HindIII sites in the pMIRREPORT luciferase expression vector (Ambion), as described before [6]. Specific fragments, including the miR P-27-5p targeting site of CDK4 3′-UTR, were generated by the primers: 5′-GGG GAG CTC GTT ACC TCA CCG ACG GTA CCT T-3′ (forward) and 5′-GGG AAG CTT CTG GTA ATA AAG AAA CAA AAC-3′ (reverse). Clones were selected after colony PCR and restriction enzyme digestion. The clones were verified by sequencing (Mission Biotech Co., Ltd.). All the restriction enzymes were purchased from New England Biolabs. Reporter activity was assayed using the Dual-Light system (Applied Biosystems) and was normalized to β-galactosidase activity to control for transfection efficiency variation among different wells according to the manufacturer’s instructions. Luminescent signal was quantified by the Spectramax M5 ELISA reader (Molecular Devices). All experiments were performed in triplicates.
3.7. Quantitative Real-Time PCR
Total RNA was isolated from T-47D cells, and first-strand cDNA synthesis was generated. Extracted first-strand cDNAs were analyzed using a BioRad iCycler iQ Real-Time Detection System with the SYBR Green dye (Molecular Probes, Eugene, OR, USA). SYBR Green yields a strong fluorescent signal on binding double-stranded DNA, enabling the quantification of gene expression by measurement of the intensity of the fluorescent light. The mRNA expressions of CDK4 were normalized to RNA content for each sample by using GADPH gene products as internal controls. The relative expression levels were calculated as the ratio of expression from miR P-27-5p transfected cancer cells to the control. All reactions were run in triplicates. The sequences of the primers specific to CDK4 were: forward primer, 5′-CTCTGCGTCCAGCTGCTCCG-3′; and reverse primer, 5′-ATCAAGGGAGACCCTCACGC-3′.
3.8. Western Blot
The protein (20 μg) was loaded on a 10% SDS-PAGE and subsequently transferred onto PVDF membranes (Amersham Biosciences) at 150 V for 1.5 h. The membranes were blocked in 5% nonfat milk in PBST containing 0.1% Tween 20, incubated with primary antibody overnight at 4 °C, and followed by incubation with a secondary antibody (a goat antirabbit-conjugated IgG, 1:10000 dilution, Upstate). CDK4, phospho-RB1, RB1, and β-actin were purchased from Cell Signaling, Epitomics, Epitomics, and Millipore, respectively. After incubation with secondary antibodies, immunoblots were visualized with the ECL detection kit (Amersham Biosciences) and exposed to x-ray film. The protein bands were quantified using ImageMaster software version 6.0 (Amersham Pharmacia Biotech, Geneva, Switzerland, 2005), and the data were normalized to β-actin.
3.9. Statistical Analysis
Statistical analysis was performed using Microsoft Office Excel 2007 (Microsoft Corporation: Redmond, WA, USA, 2007). Student’s t test and P-values were tested for multiple comparisons by controlling the false discovery rate. Changes were considered significant if the false discovery rate was less than 0.05.
4. Conclusions
Our results reveal that miR P-27-5p targets and negatively regulates CDK4, which in turn suppresses RB1 phosphorylation, thereby preventing the progression of cancer cell cycle and promoting G1 arrest. As a consequence, cancer cell proliferation is inhibited. Thus, on the therapeutic side, artificial miR P-27-5p has the potential for the application in the treatment of breast cancer. miRNA can affect many downstream targets, which in turn form a complicated network. Therefore, there is considerable potential for the miRNA-regulated network to be a novel therapeutic target. The therapeutic strategy could be used for cancer as well as the other diseases such as neurological and cardiovascular disorders. RNA interference and miRNA oligonucleotides have clinical potential. In the future, using the miR P-27-5p-regulated network as a novel therapeutic target, especially responsible for G1 arrest, may help reduce off-target effects and alleviate the risk of side effects.
Supplementary Material
ijms-13-06352-s001.xlsAcknowledgments
This work was supported by the National Science Council of Taiwan (NSC 99-2628-B-001-009-MY3, NSC 99-2621-B-002-005-MY3, NSC 99-2621-B-010-001-MY3, and NSC 99-2621-B-001-003-MY3) and National Taiwan University Cutting-Edge Steering Research Project (10R70602C3). We thank Technology Commons, College of Life Science, National Taiwan University, for providing technical assistance.
References
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar]
- Ventura, A.; Jacks, T. MicroRNAs and cancer: Short RNAs go a long way. Cell 2009, 136, 586–591. [Google Scholar]
- Mattie, M.D.; Benz, C.C.; Bowers, J.; Sensinger, K.; Wong, L.; Scott, G.K.; Fedele, V.; Ginzinger, D.; Getts, R.; Haqq, C. Optimized high-throughput microRNA expression profiling provides novel biomarker assessment of clinical prostate and breast cancer biopsies. Mol. Cancer 2006, 5. [Google Scholar] [CrossRef]
- Yanaihara, N.; Caplen, N.; Bowman, E.; Seike, M.; Kumamoto, K.; Yi, M.; Stephens, R.M.; Okamoto, A.; Yokota, J.; Tanaka, T.; et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell 2006, 9, 189–198. [Google Scholar]
- Lieberman, J. Micromanaging cancer. N. Engl. J. Med 2009, 361, 1500–1501. [Google Scholar]
- Chang, Y.M.; Juan, H.F.; Lee, T.Y.; Chang, Y.Y.; Yeh, Y.M.; Li, W.H.; Shih, A.C. Prediction of human miRNAs using tissue-selective motifs in 3′ UTRs. Proc. Natl. Acad. Sci. USA 2008, 105, 17061–17066. [Google Scholar]
- Yue, H.; Eastman, P.S.; Wang, B.B.; Minor, J.; Doctolero, M.H.; Nuttall, R.L.; Stack, R.; Becker, J.W.; Montgomery, J.R.; Vainer, M.; et al. An evaluation of the performance of cDNA microarrays for detecting changes in global mRNA expression. Nucleic Acids Res 2001, 29. [Google Scholar] [CrossRef]
- Ideker, T.; Thorsson, V.; Ranish, J.A.; Christmas, R.; Buhler, J.; Eng, J.K.; Bumgarner, R.; Goodlett, D.R.; Aebersold, R.; Hood, L. Integrated genomic and proteomic analyses of a systematically perturbed metabolic network. Science 2001, 292, 929–934. [Google Scholar]
- Hughes, T.R.; Marton, M.J.; Jones, A.R.; Roberts, C.J.; Stoughton, R.; Armour, C.D.; Bennett, H.A.; Coffey, E.; Dai, H.; He, Y.D.; et al. Functional discovery via a compendium of expression profiles. Cell 2000, 102, 109–126. [Google Scholar]
- Abdueva, D.; Wing, M.R.; Schaub, B.; Triche, T.J. Experimental comparison and evaluation of the Affymetrix exon and U133Plus2 GeneChip arrays. PLoS One 2007, 2. [Google Scholar] [CrossRef]
- Seo, J.; Kim, M.; Kim, J. Identification of novel genes differentially expressed in PMA-induced HL-60 cells using cDNA microarrays. Mol. Cells 2000, 10, 733–739. [Google Scholar]
- Le Naour, F.; Hohenkirk, L.; Grolleau, A.; Misek, D.E.; Lescure, P.; Geiger, J.D.; Hanash, S.; Beretta, L. Profiling changes in gene expression during differentiation and maturation of monocyte-derived dendritic cells using both oligonucleotide microarrays and proteomics. J. Biol. Chem 2001, 276, 17920–17931. [Google Scholar]
- Dong, Y.; Sui, L.; Sugimoto, K.; Tai, Y.; Tokuda, M. Cyclin D1-CDK4 complex, a possible critical factor for cell proliferation and prognosis in laryngeal squamous cell carcinomas. Int. J. Cancer 2001, 95, 209–215. [Google Scholar]
- Dean, J.L.; Thangavel, C.; McClendon, A.K.; Reed, C.A.; Knudsen, E.S. Therapeutic CDK4/6 inhibition in breast cancer: Key mechanisms of response and failure. Oncogene 2010, 29, 4018–4032. [Google Scholar]
- Sun, Y.; Li, Y.X.; Wu, H.J.; Wu, S.H.; Wang, Y.A.; Luo, D.Z.; Liao, D.J. Effects of an indolocarbazole-derived CDK4 inhibitor on breast cancer cells. J. Cancer 2011, 2, 36–51. [Google Scholar]
- Lin, S.L.; Chang, D.C.; Ying, S.Y.; Leu, D.; Wu, D.T. MicroRNA miR-302 inhibits the tumorigenecity of human pluripotent stem cells by coordinate suppression of the CDK2 and CDK4/6 cell cycle pathways. Cancer Res 2010, 70, 9473–9482. [Google Scholar]
- Burkhart, D.L.; Sage, J. Cellular mechanisms of tumour suppression by the retinoblastoma gene. Nat. Rev. Cancer 2008, 8, 671–682. [Google Scholar]
- Yoon, S.; Choi, Y.C.; Lee, S.; Jeong, Y.; Yoon, J.; Baek, K. Induction of growth arrest by miR-542–3p that targets survivin. FEBS Lett 2010, 584, 4048–4052. [Google Scholar]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsley, P.S.; Johnson, J.M. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005, 433, 769–773. [Google Scholar]
- Tseng, C.W.; Lin, C.C.; Chen, C.N.; Huang, H.C.; Juan, H.F. Integrative network analysis reveals active microRNAs and their functions in gastric cancer. BMC Syst. Biol 2011, 5. [Google Scholar] [CrossRef]
- Tseng, C.W.; Yang, J.C.; Chen, C.N.; Huang, H.C.; Chuang, K.N.; Lin, C.C.; Lai, H.S.; Lee, P.H.; Chang, K.J.; Juan, H.F. Identification of 14-3-3beta in human gastric cancer cells and its potency as a diagnostic and prognostic biomarker. Proteomics 2011, 11, 2423–2439. [Google Scholar]
- Mathivanan, S.; Periaswamy, B.; Gandhi, T.K.; Kandasamy, K.; Suresh, S.; Mohmood, R.; Ramachandra, Y.L.; Pandey, A. An evaluation of human protein-protein interaction data in the public domain. BMC Bioinform 2006, 7. [Google Scholar] [CrossRef]
- Maere, S.; Heymans, K.; Kuiper, M. BiNGO: A Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 2005, 21, 3448–3449. [Google Scholar]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res 2003, 13, 2498–2504. [Google Scholar]
- Sutherland, R.L.; Musgrove, E.A. CDK inhibitors as potential breast cancer therapeutics: New evidence for enhanced efficacy in ER+ disease. Breast Cancer Res 2009, 11. [Google Scholar] [CrossRef]
- Zelivianski, S.; Cooley, A.; Kall, R.; Jeruss, J.S. Cyclin-dependent kinase 4-mediated phosphorylation inhibits Smad3 activity in cyclin D-overexpressing breast cancer cells. Mol. Cancer Res 2010, 8, 1375–1387. [Google Scholar]
- Farazi, T.A.; Horlings, H.M.; ten Hoeve, J.J.; Mihailovic, A.; Halfwerk, H.; Morozov, P.; Brown, M.; Hafner, M.; Reyal, F.; van Kouwenhove, M.; et al. MicroRNA sequence and expression analysis in breast tumors by deep sequencing. Cancer Res 2011, 71, 4443–4453. [Google Scholar]
- Takada, S.; Berezikov, E.; Yamashita, Y.; Lagos-Quintana, M.; Kloosterman, W.P.; Enomoto, M.; Hatanaka, H.; Fujiwara, S.; Watanabe, H.; Soda, M.; et al. Mouse microRNA profiles determined with a new and sensitive cloning method. Nucleic Acids Res 2006, 34. [Google Scholar] [CrossRef]
- Kuhn, D.E.; Nuovo, G.J.; Martin, M.M.; Malana, G.E.; Pleister, A.P.; Jiang, J.; Schmittgen, T.D.; Terry, A.V., Jr; Gardiner, K.; Head, E.; et al. Human chromosome 21-derived miRNAs are overexpressed in down syndrome brains and hearts. Biochem. Biophys. Res. Commun 2008, 370, 473–477. [Google Scholar]
- Kwon, O.H.; Park, J.L.; Kim, M.; Kim, J.H.; Lee, H.C.; Kim, H.J.; Noh, S.M.; Song, K.S.; Yoo, H.S.; Paik, S.G.; et al. Aberrant up-regulation of LAMB3 and LAMC2 by promoter demethylation in gastric cancer. Biochem. Biophys. Res. Commun 2011, 406, 539–545. [Google Scholar]
- Kita, Y.; Mimori, K.; Tanaka, F.; Matsumoto, T.; Haraguchi, N.; Ishikawa, K.; Matsuzaki, S.; Fukuyoshi, Y.; Inoue, H.; Natsugoe, S.; et al. Clinical significance of LAMB3 and COL7A1 mRNA in esophageal squamous cell carcinoma. Eur. J. Surg. Oncol 2009, 35, 52–58. [Google Scholar]
- Calaluce, R.; Bearss, D.J.; Barrera, J.; Zhao, Y.; Han, H.; Beck, S.K.; McDaniel, K.; Nagle, R.B. Laminin-5 beta3A expression in LNCaP human prostate carcinoma cells increases cell migration and tumorigenicity. Neoplasia 2004, 6, 468–479. [Google Scholar]
- De Guire, V.; Caron, M.; Scott, N.; Menard, C.; Gaumont-Leclerc, M.F.; Chartrand, P.; Major, F.; Ferbeyre, G. Designing small multiple-target artificial RNAs. Nucleic Acids Res 2010, 38. [Google Scholar] [CrossRef]
- Brown, B.D.; Naldini, L. Exploiting and antagonizing microRNA regulation for therapeutic and experimental applications. Nat. Rev. Genet 2009, 10, 578–585. [Google Scholar]
- Idogawa, M.; Sasaki, Y.; Suzuki, H.; Mita, H.; Imai, K.; Shinomura, Y.; Tokino, T. A single recombinant adenovirus expressing p53 and p21-targeting artificial microRNAs efficiently induces apoptosis in human cancer cells. Clin. Cancer Res 2009, 15, 3725–3732. [Google Scholar]
- Prud’homme, G.J.; Glinka, Y.; Toulina, A.; Ace, O.; Subramaniam, V.; Jothy, S. Breast cancer stem-like cells are inhibited by a non-toxic aryl hydrocarbon receptor agonist. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Zhong, Z.; Yeow, W.S.; Zou, C.; Wassell, R.; Wang, C.; Pestell, R.G.; Quong, J.N.; Quong, A.A. Cyclin D1/cyclin-dependent kinase 4 interacts with filamin A and affects the migration and invasion potential of breast cancer cells. Cancer Res 2010, 70, 2105–2114. [Google Scholar]
- Ye, X.; Liu, Z.; Hemida, M.G.; Yang, D. Targeted delivery of mutant tolerant anti-coxsackievirus artificial microRNAs using folate conjugated bacteriophage Phi29 pRNA. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- dChip Software: Analysis and visualization of gene expression and SNP microarrays. Available online: http://biosun1.harvard.edu/complab/dchip accessed on 3 January 2012.
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).