Rhinacanthus nasutus Extracts Prevent Glutamate and Amyloid-β Neurotoxicity in HT-22 Mouse Hippocampal Cells: Possible Active Compounds Include Lupeol, Stigmasterol and β-Sitosterol

 ,
, 

Abstract
:1. Introduction
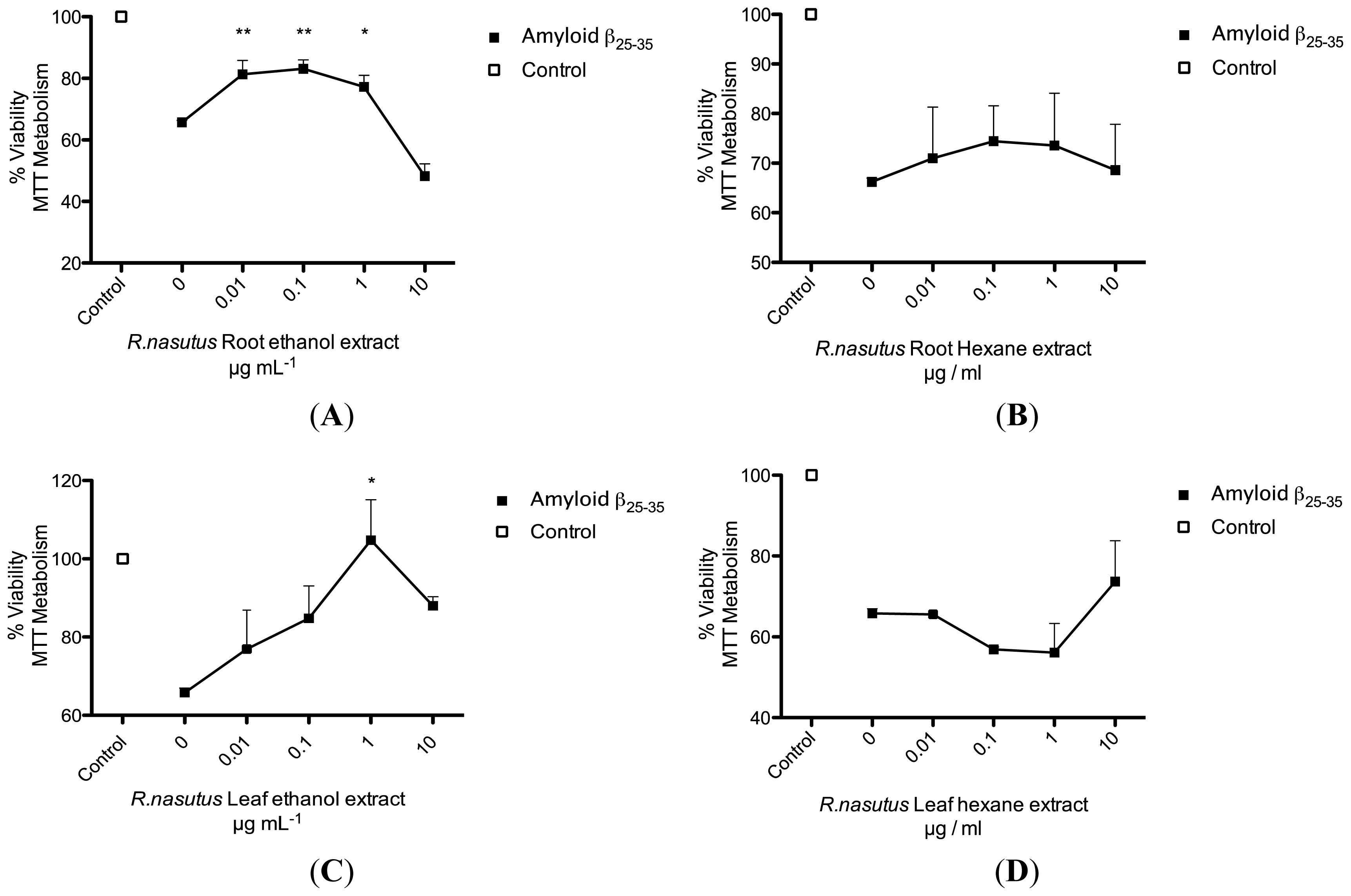
2. Results and Discussion
2.1. Rhinacanthus Nasutus and Its Antioxidant Properties
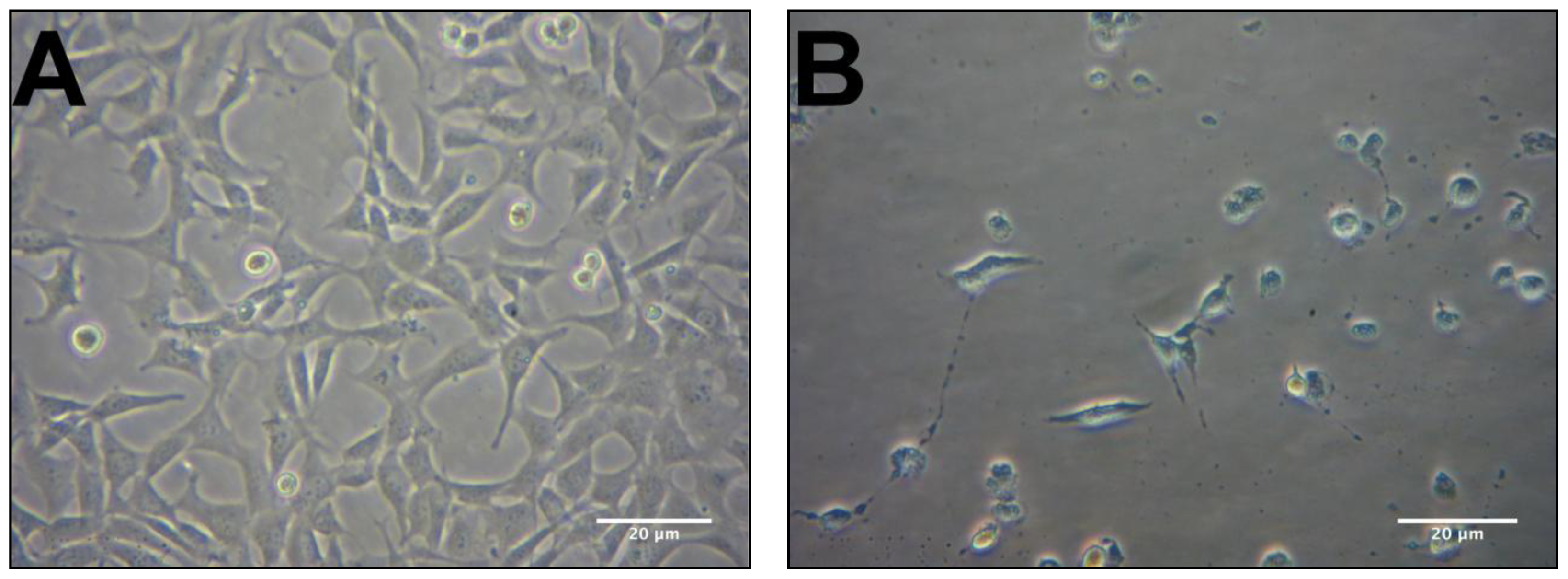
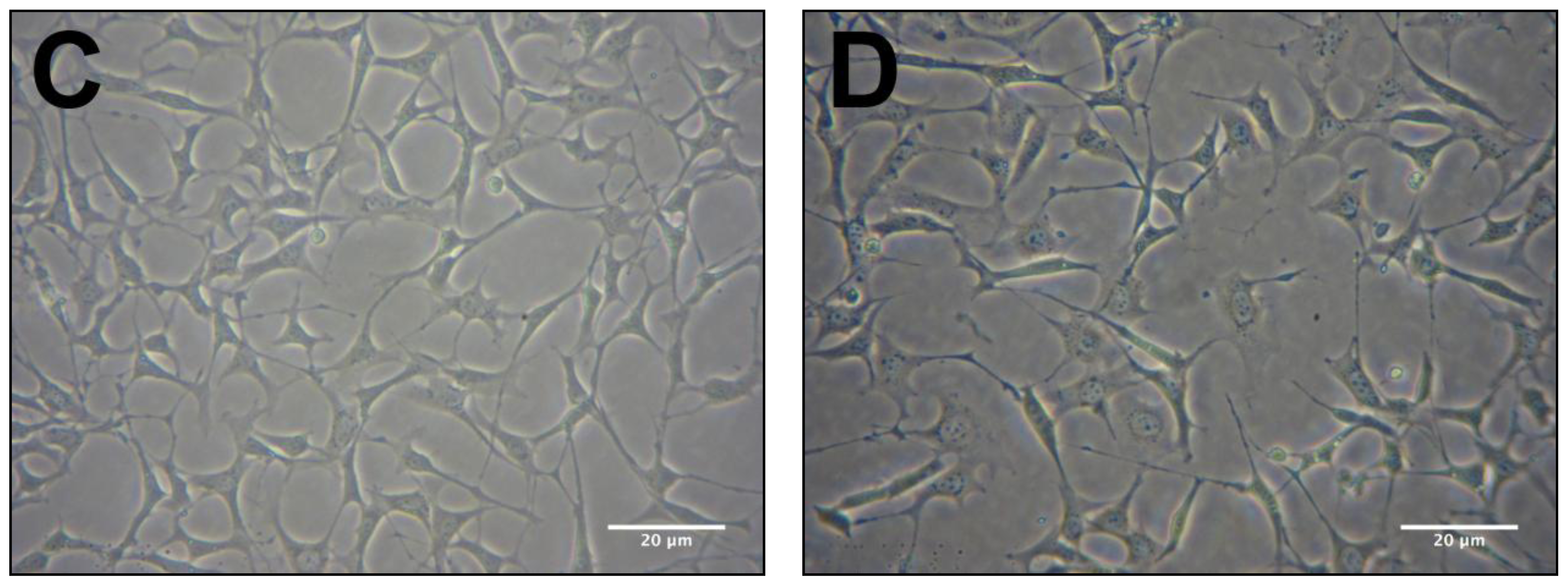
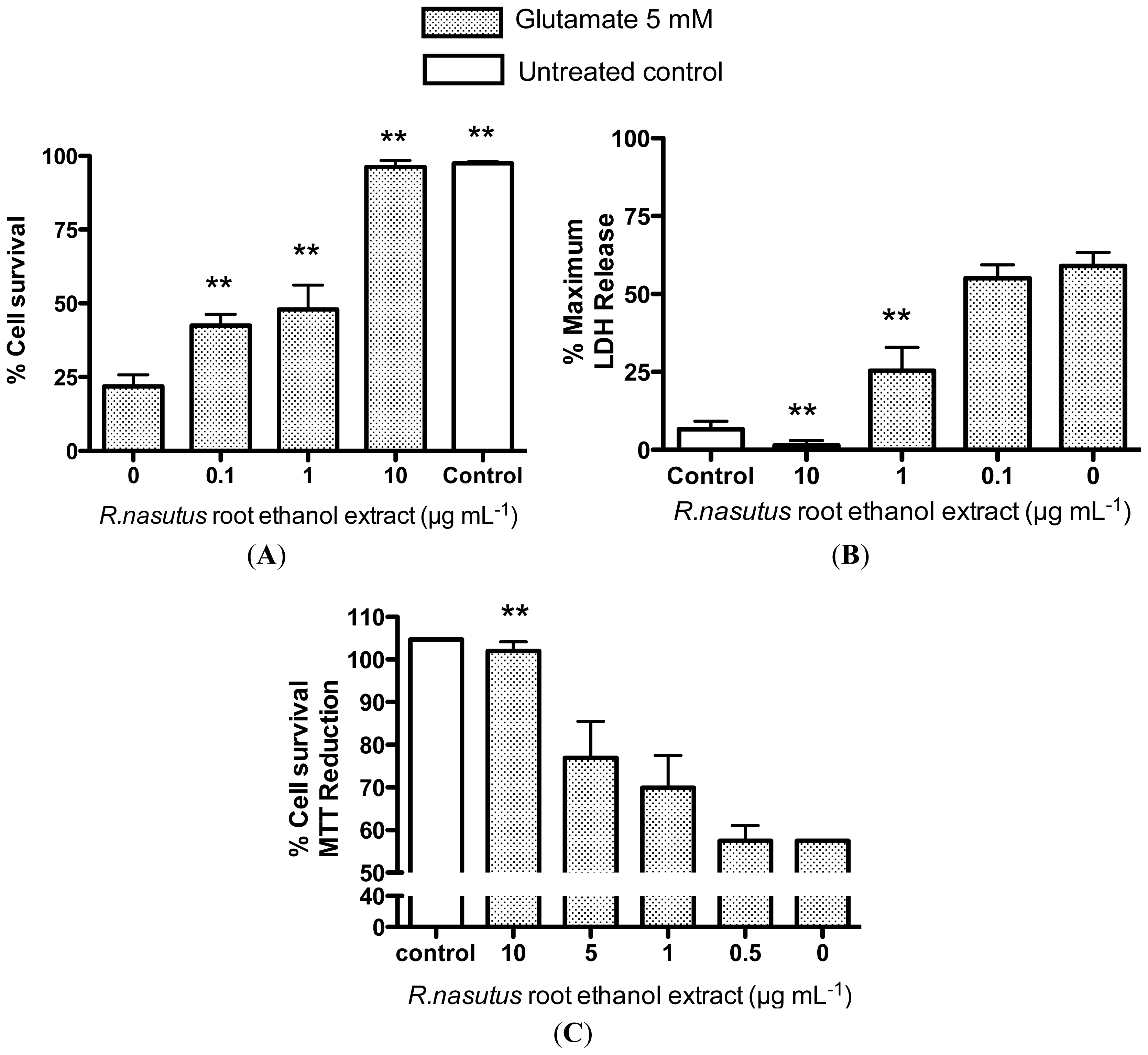
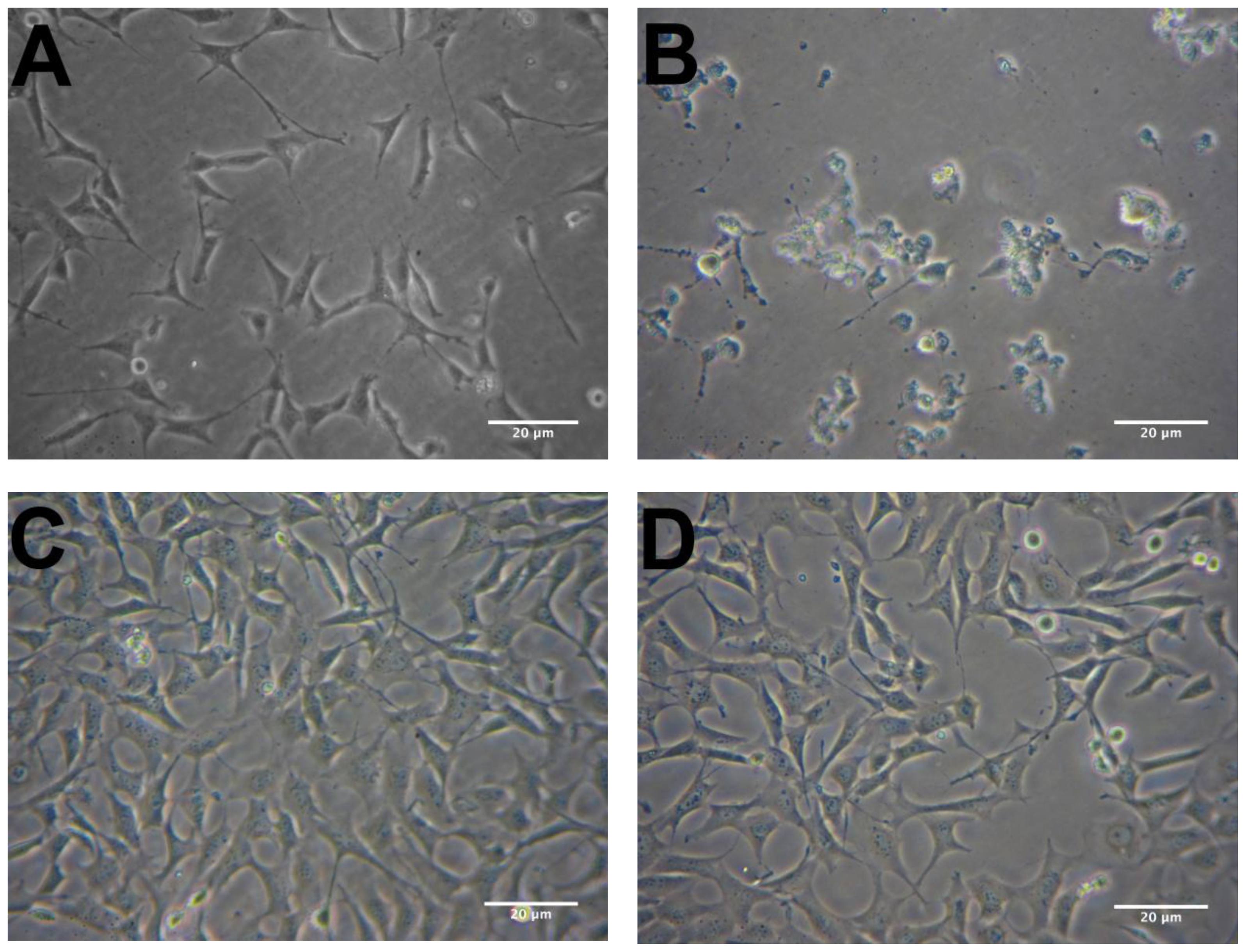
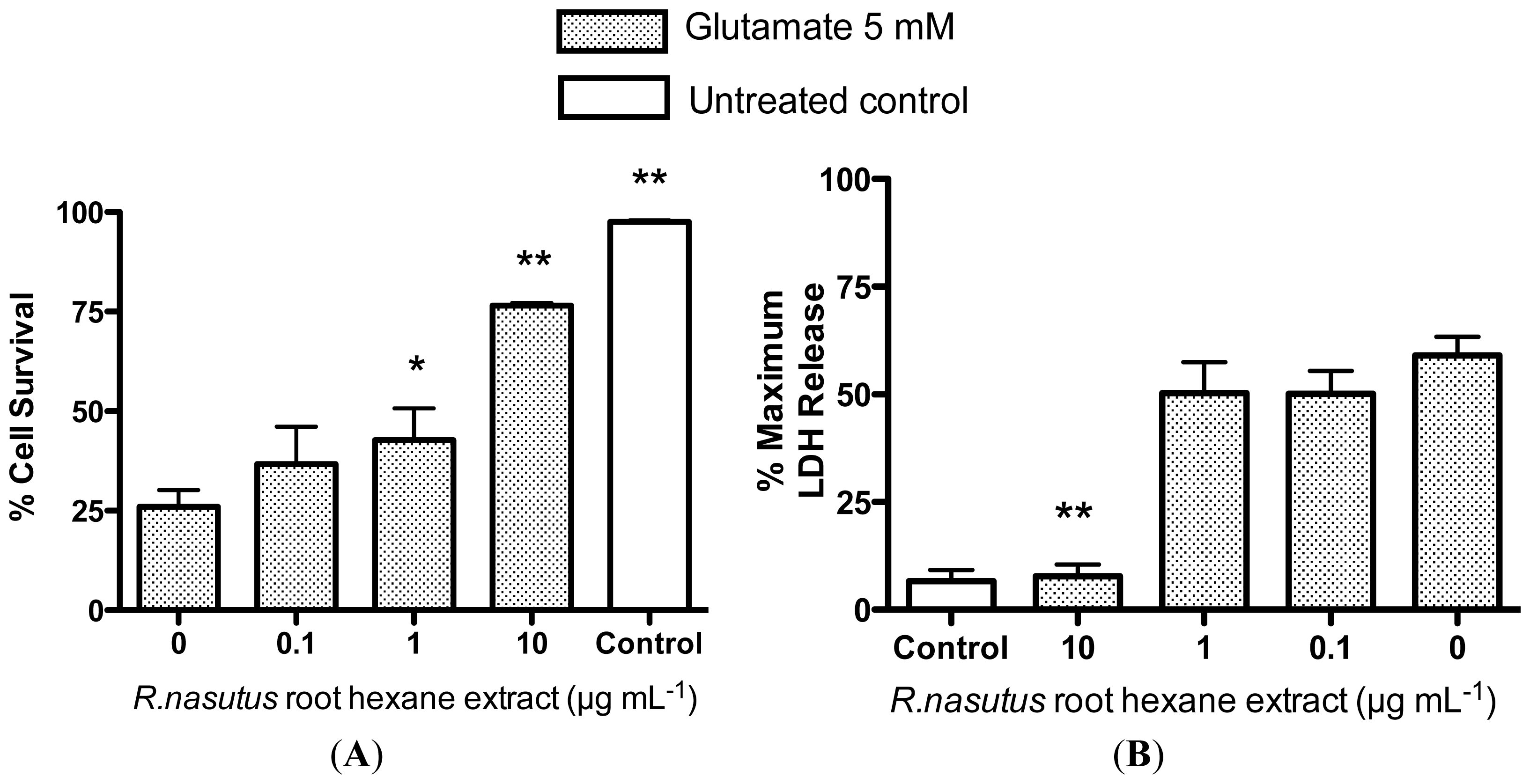
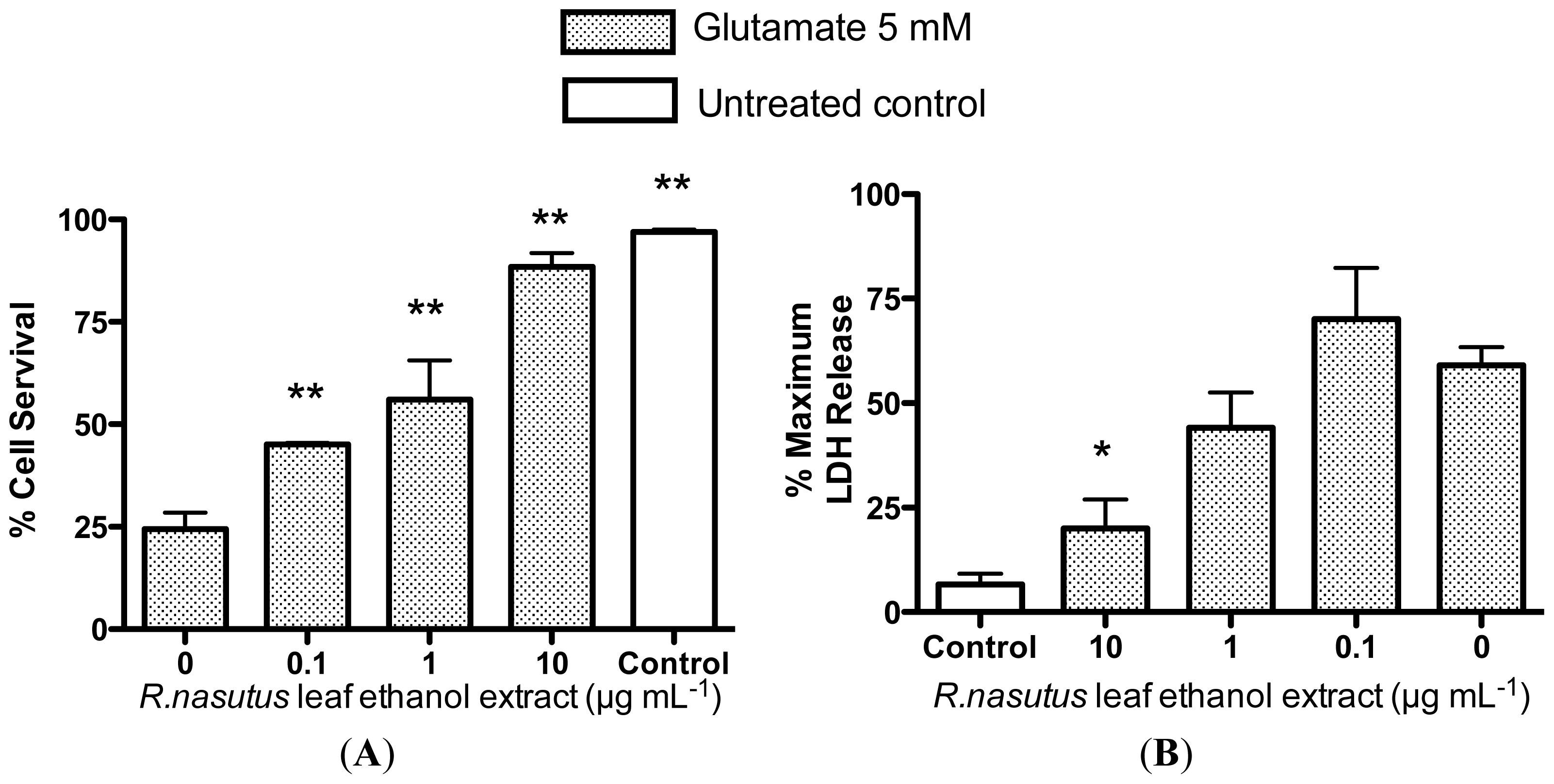
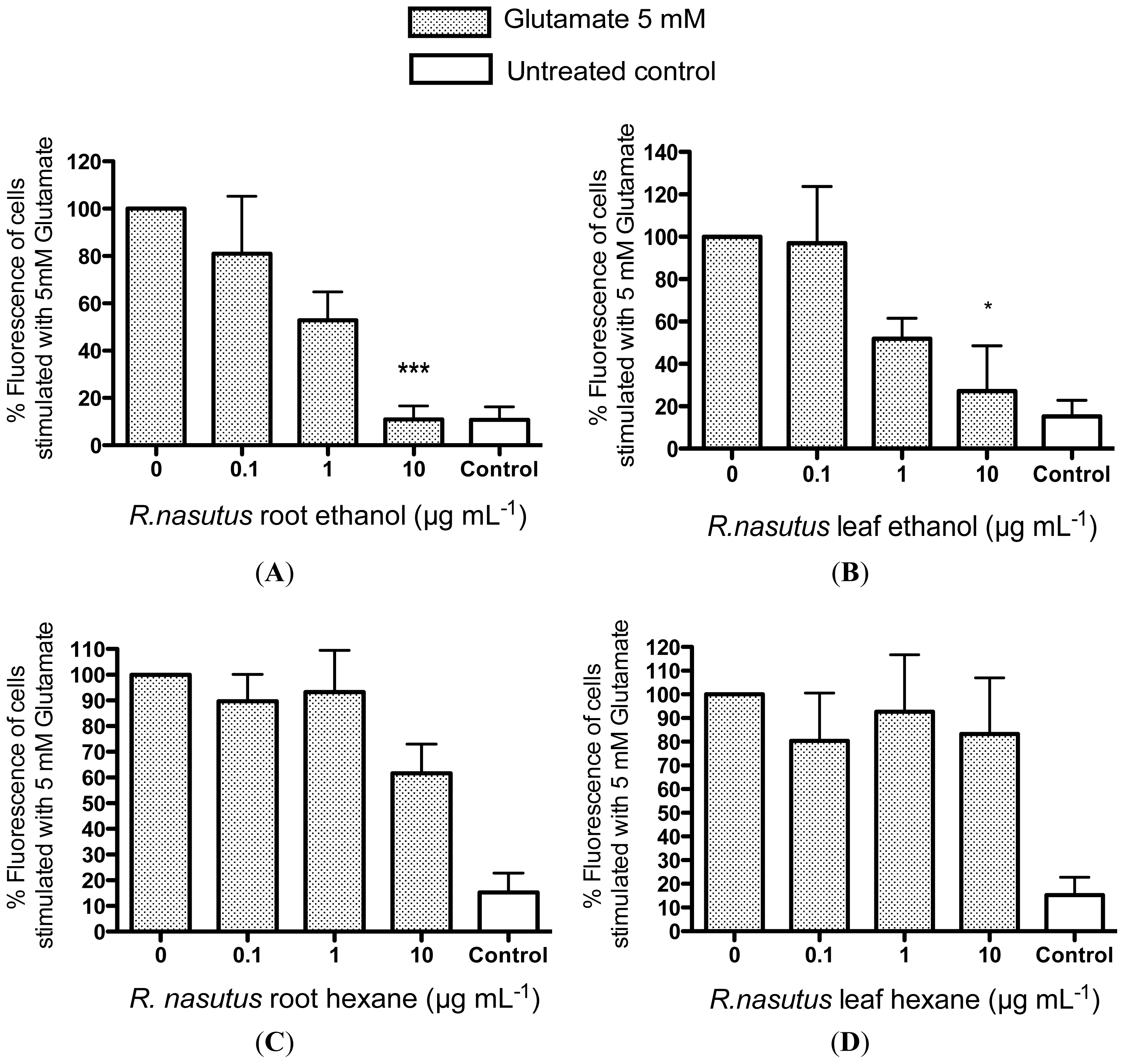
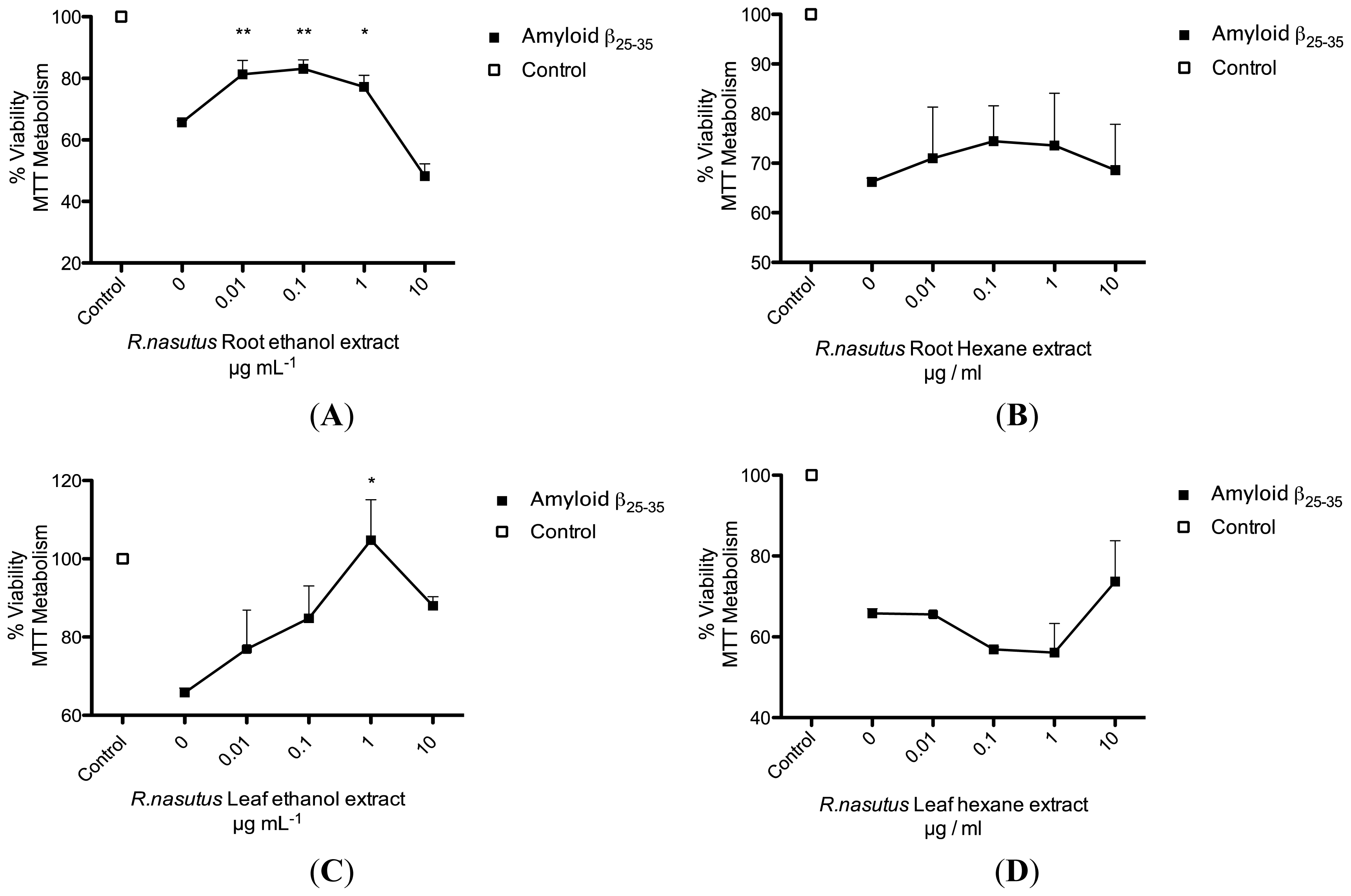
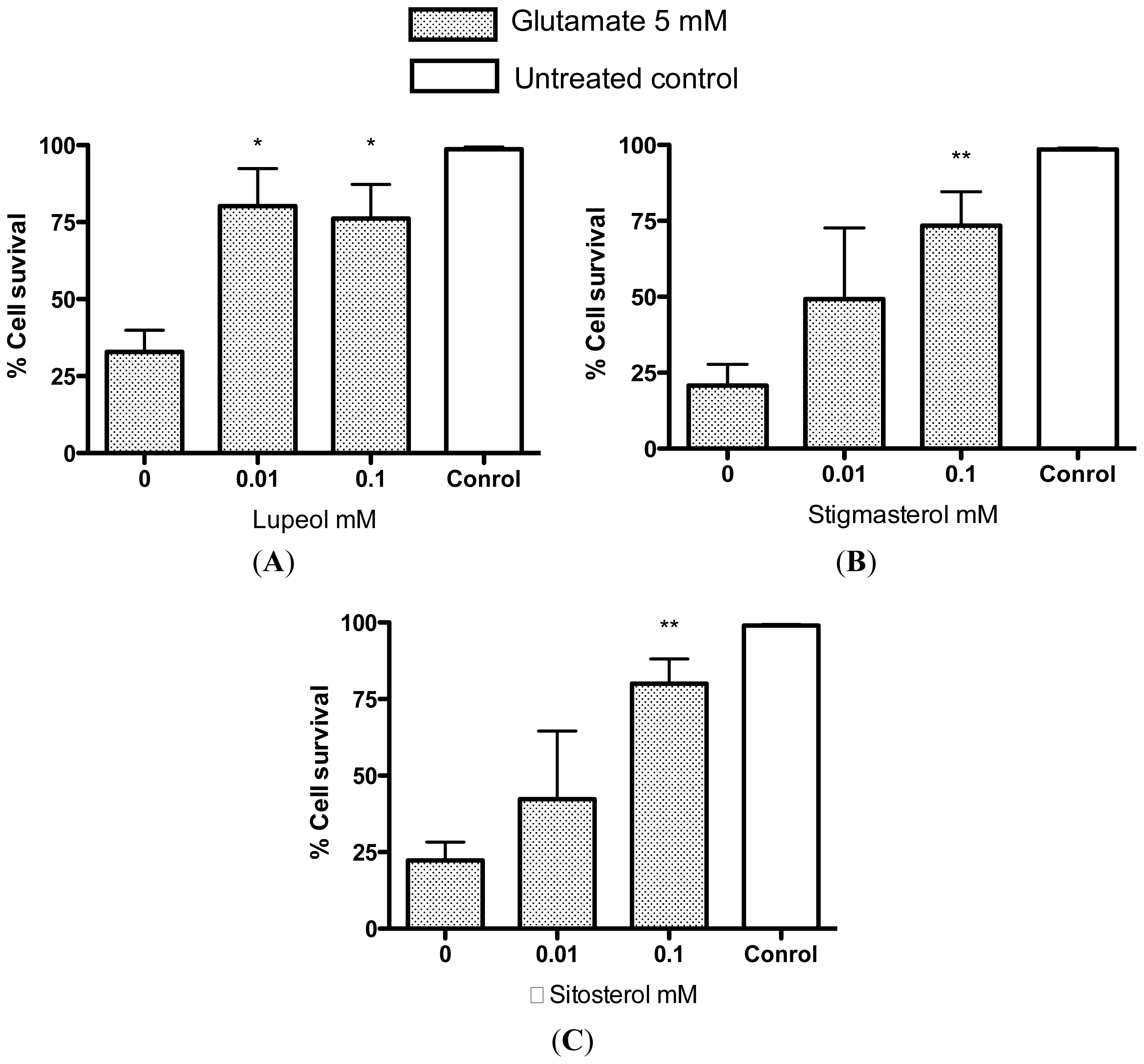
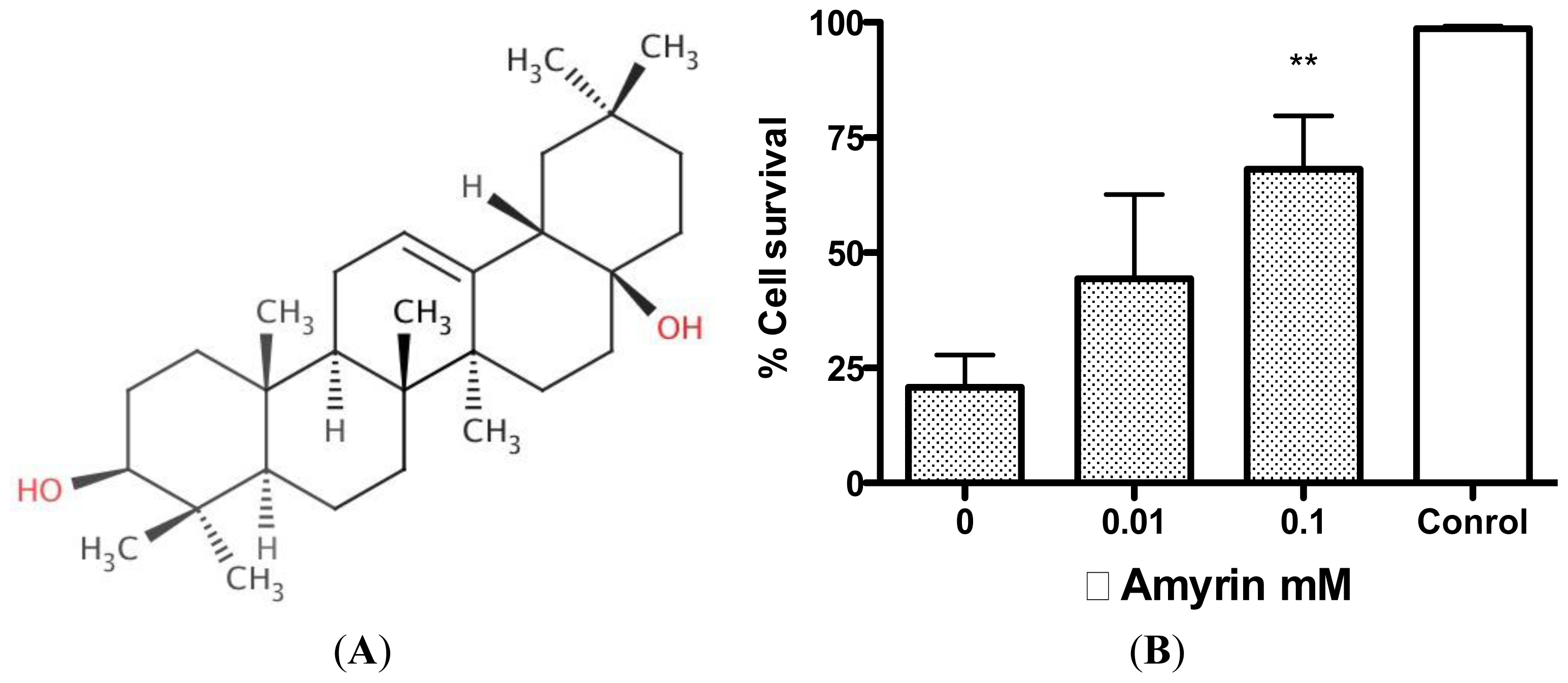
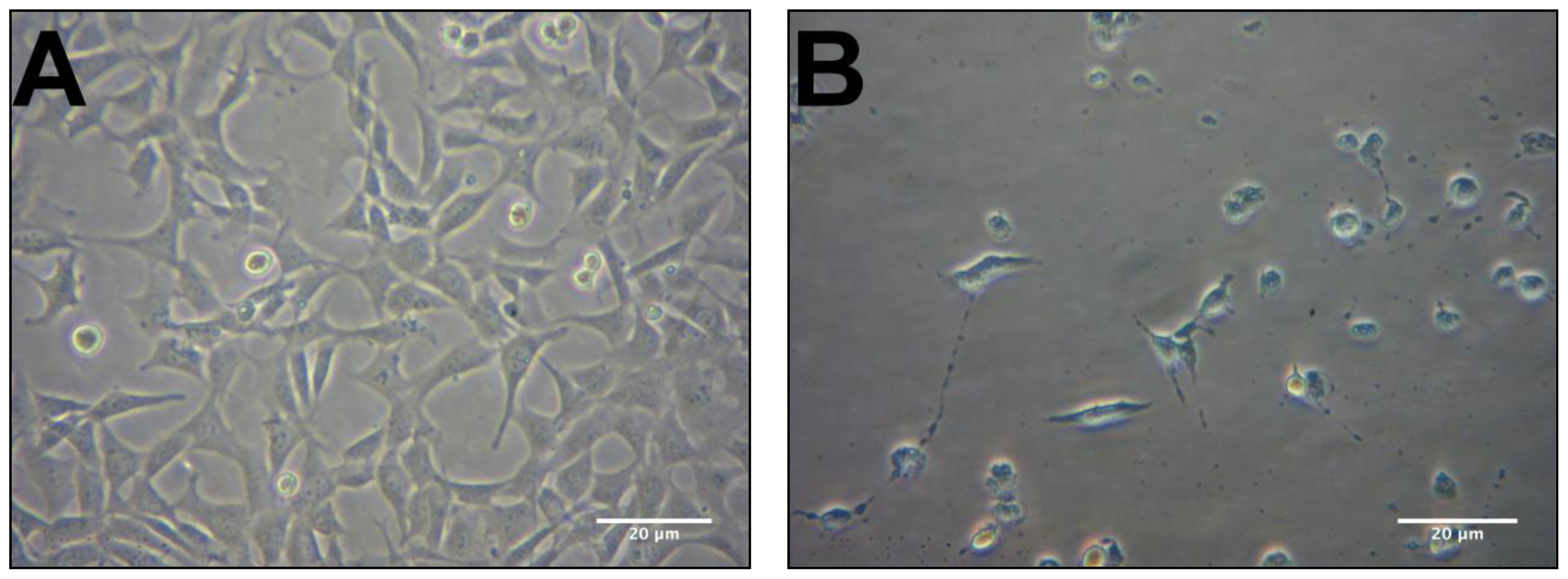
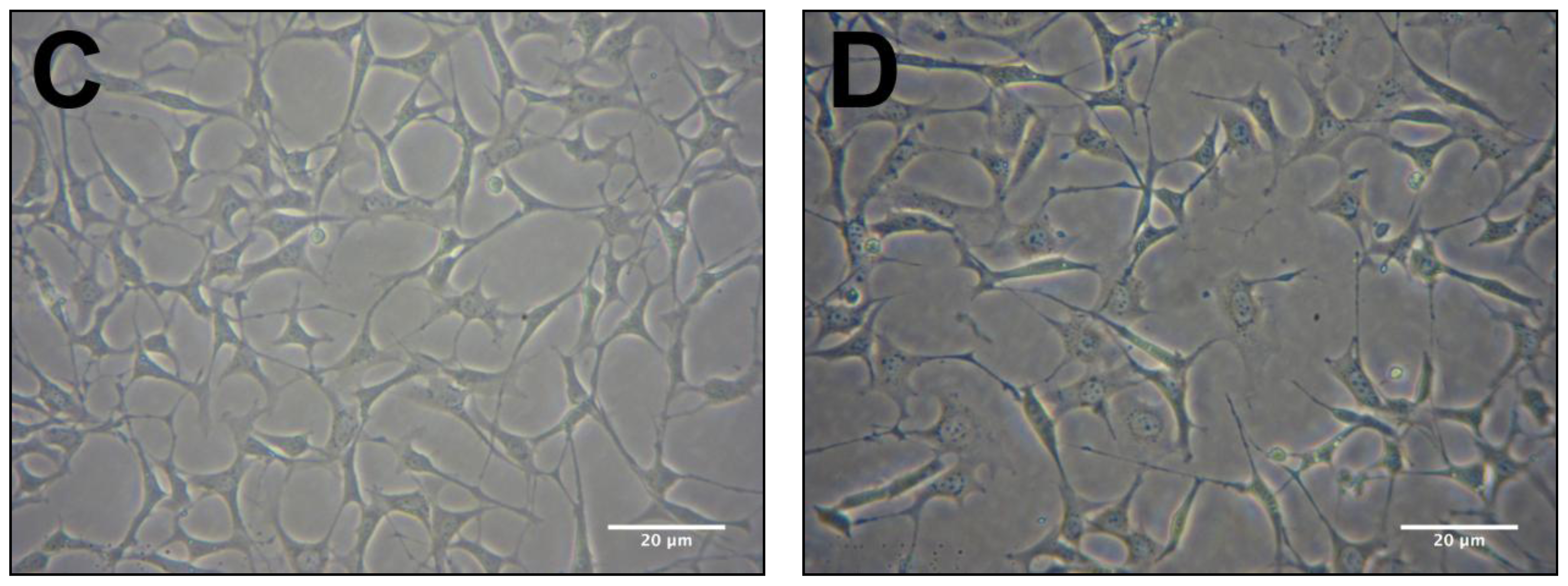
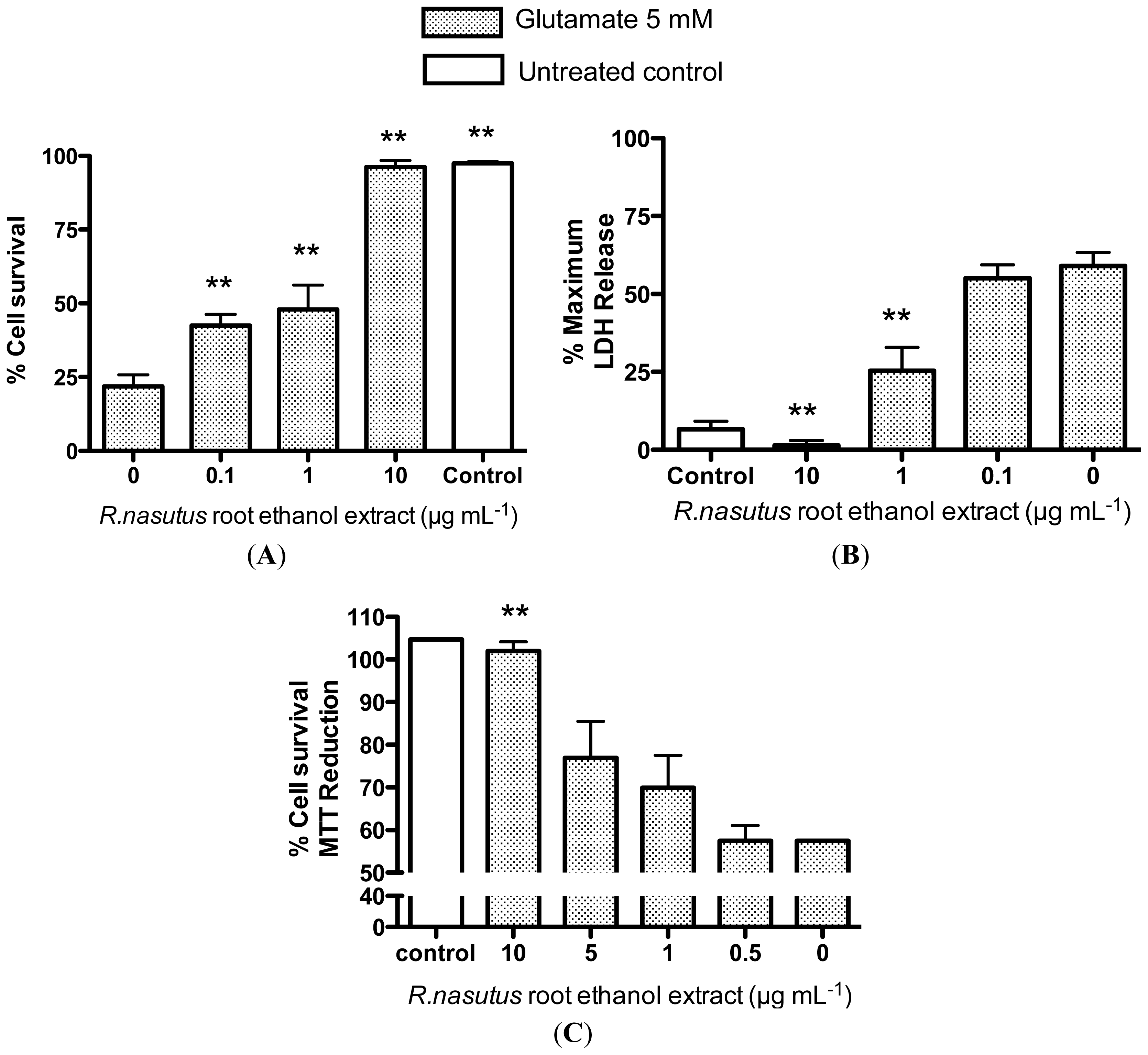
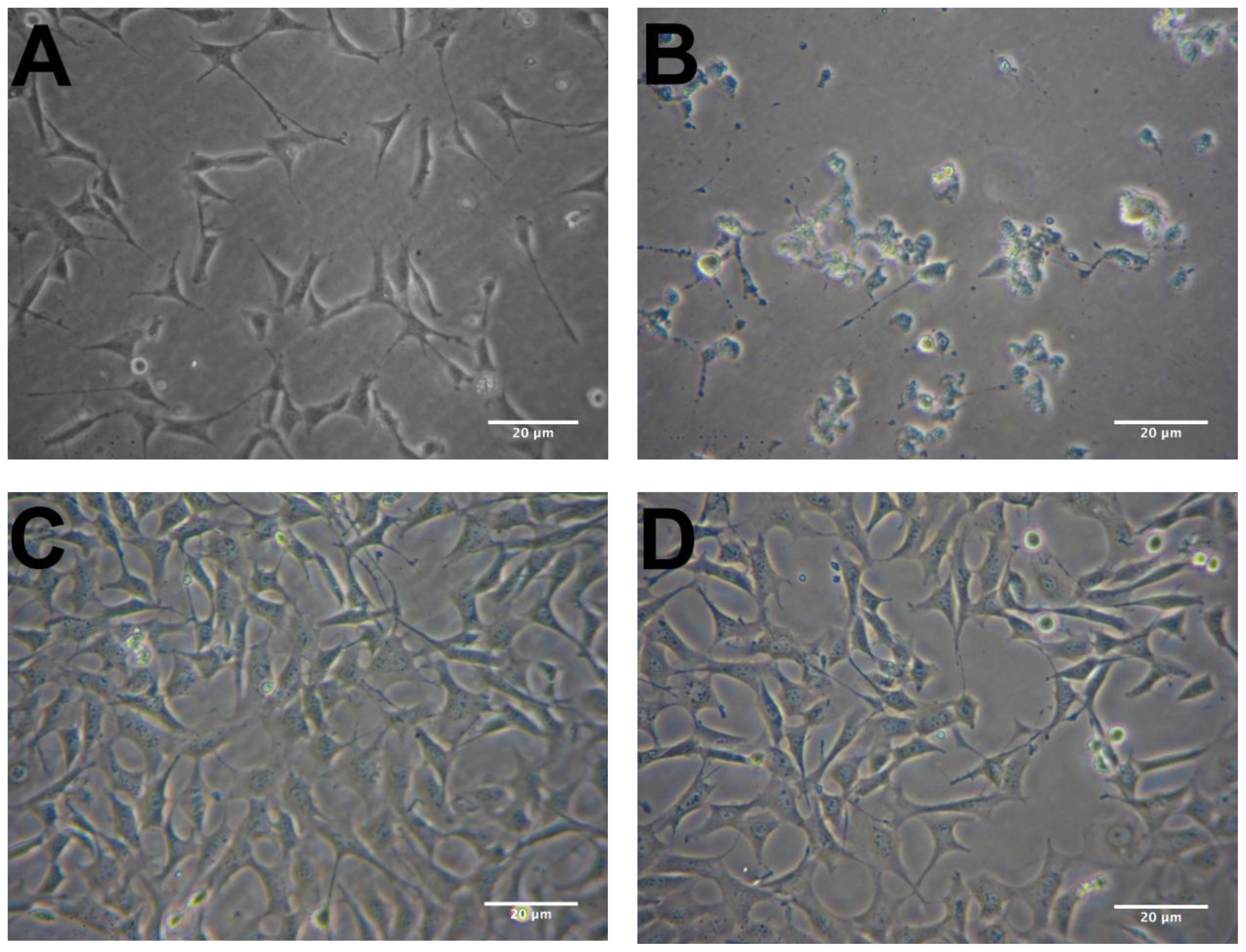
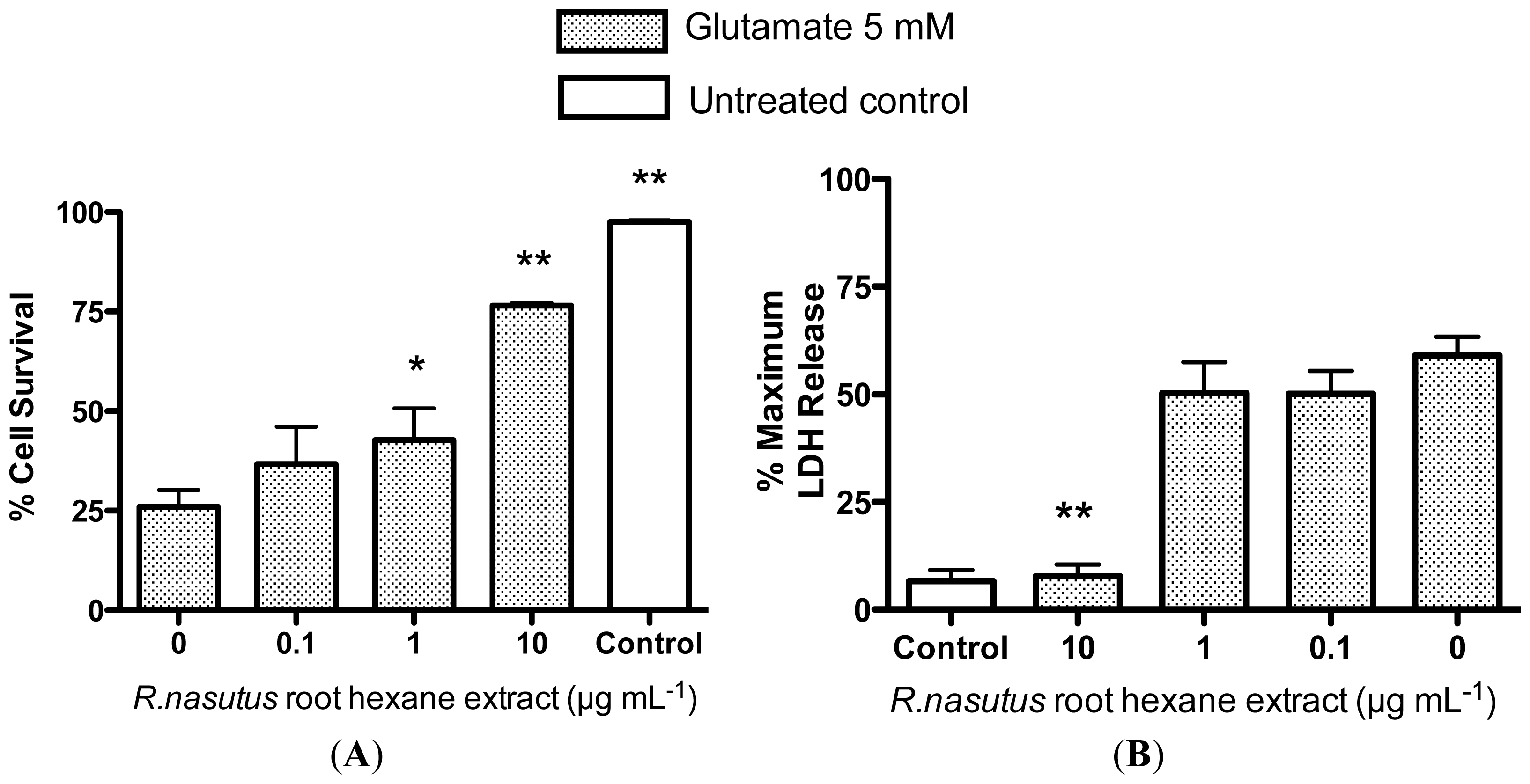
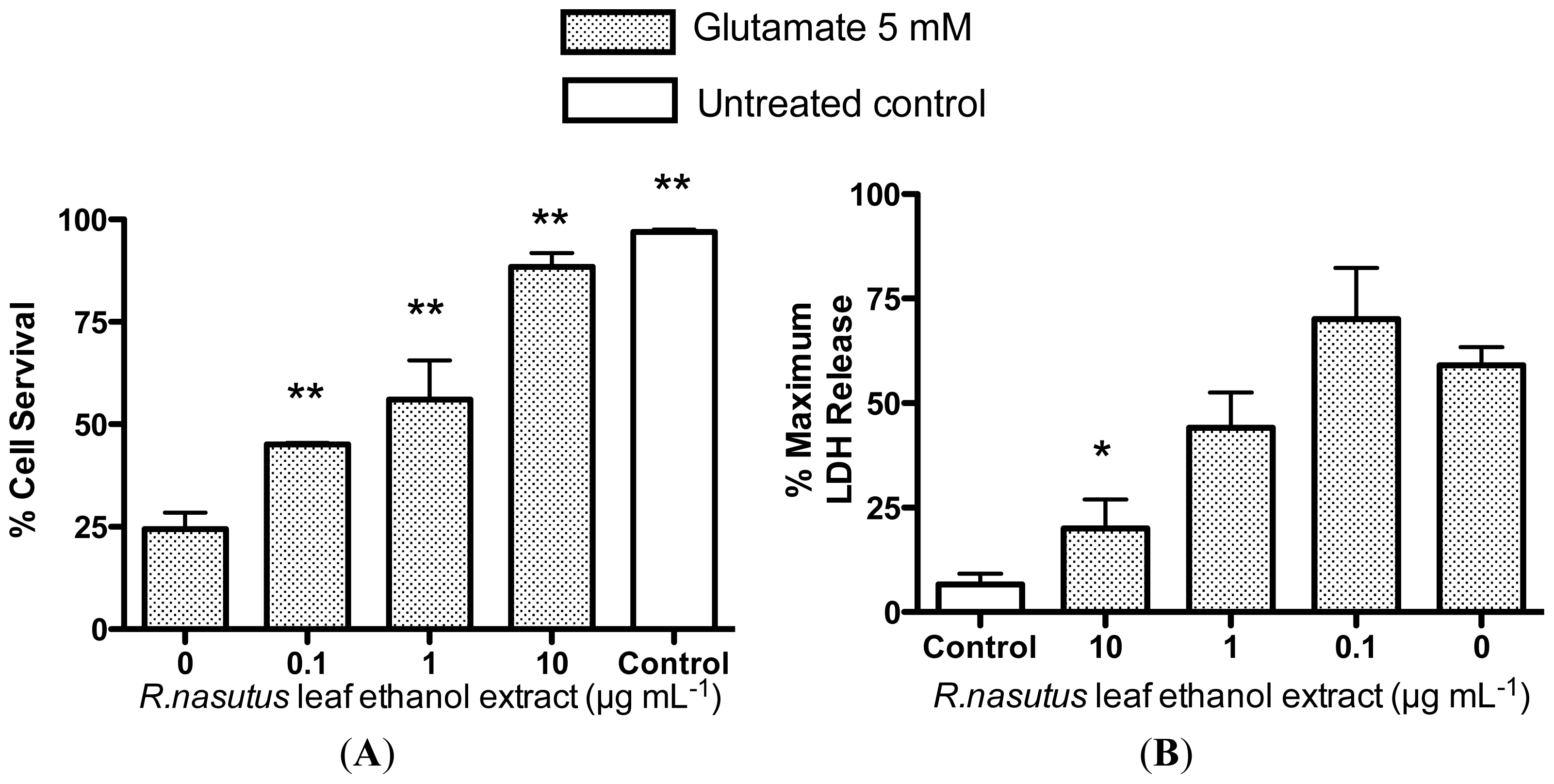
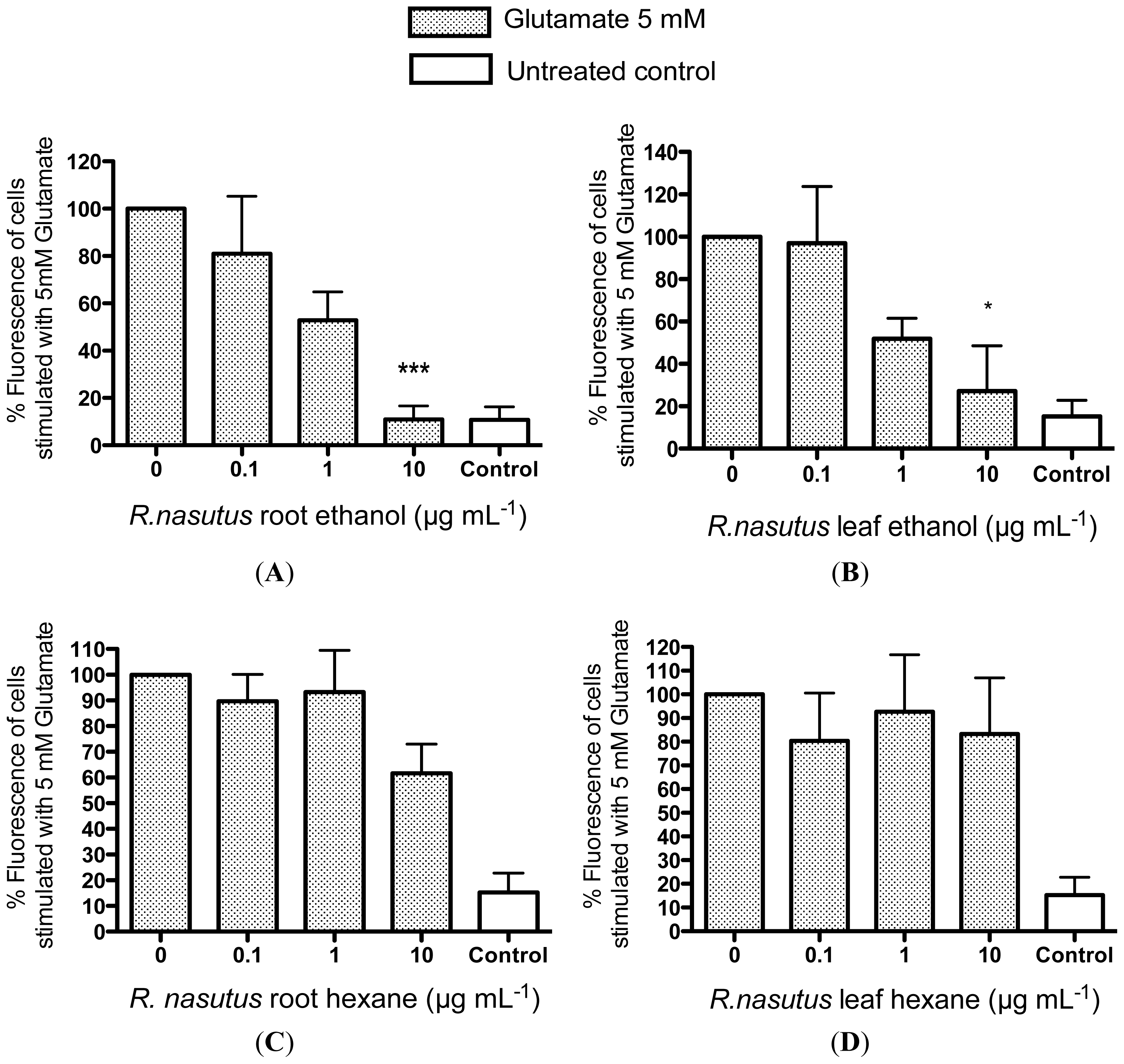
2.2. Protective Effects of R. nasutus Against Glutamate Toxicity in Mouse Hippocampus (HT-22) Cells
3. Experimental Section
3.1. Chemicals
3.2. Amyloid-β25–35
3.3. Cell Culture
3.4. Plant Material
3.5. Trypan Blue Exclusion Assay
3.6. 3-(4,5-Ethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide (MTT) Assay
3.7. Lactate Dehydrogenase Release (LDH) Assay
3.8. Total Phenolic Content Assay
3.9. Total Flavonoid Content Assay
3.10. 2,2′-Azino-bis(3-ethyl)benzothiazoline-6-sulphonic Acid (ABTS) Decolorization Assay
3.11. Hydroxyl Radical Scavenging Assay
3.12. Nitric Oxide Scavenging Assay
3.13. Superoxide Scavenging Assay
3.14. 2,2-diphenyl-1-Picrylhydrazyl (DPPH) Assay
3.15. Carboxy-H2-DFCDA Free Radical Assay
3.16. High Performance Liquid Chromatography
3.17. Data Analysis
4. Conclusions
Acknowledgments
References
- Thongrakard, V.; Tencomnao, T. Modulatory effects of Thai medicinal plant extract on proinflammatory cytokines-induced apoptosis in human keratinocyte HaCaT cells. Afr. J. Biotechnol 2010, 9, 4999–5003. [Google Scholar]
- Shyamal, S.; Latha, P.G.; Suja, S.R.; Shine, V.J.; Anuja, G.I.; Sini, S.; Pradeep, S.; Shikha, P.; Rajasekharan, S. Hepatoprotective effect of three herbal extracts on aflatoxin B1-intoxicated rat liver. Singap. Med. J 2010, 51, 326–331. [Google Scholar]
- Brimson, J.M.; Tencomnao, T. Rhinacanthus nasutus protects cultured neuronal cells against hypoxia induced cell death. Mol. Basel Switz 2011, 16, 6322–6338. [Google Scholar]
- Choi, D.W. Excitotoxic cell death. Dev. Neurobiol 1992, 23, 1261–1276. [Google Scholar]
- Patel, V.P.; Chu, C.T. Review Article Nuclear transport, oxidative stress, and neurodegeneration. Int. J. Clin. Exp. Pathol 2011, 4, 215–229. [Google Scholar]
- Coyle, J.T.; Puttfarcken, P. Oxidative stress, glutamate, and neurodegenerative disorders. Science 1993, 262, 689–695. [Google Scholar]
- Luo, H.; Huang, J.; Liao, W.-G.; Huang, Q.-Y.; Gao, Y.-Q. The antioxidant effects of garlic saponins protect PC12 cells from hypoxia-induced damage. Br. J. Nutr 2011, 105, 1164–1172. [Google Scholar]
- Jin, D.-Q.; Lim, C.S.; Hwang, J.K.; Ha, I.; Han, J.-S. Anti-oxidant and anti-inflammatory activities of macelignan in murine hippocampal cell line and primary culture of rat microglial cells. Biochem. Biophys. Res. Commun 2005, 331, 1264–1269. [Google Scholar]
- Sucontphunt, A.; De-Eknamkul, W.; Nimmannit, U.; Dan Dimitrijevich, S.; Gracy, R.W. Protection of HT22 neuronal cells against glutamate toxicity mediated by the antioxidant activity of Pueraria candollei var. mirifica extracts. J. Nat. Med 2011, 65, 1–8. [Google Scholar]
- McEntee, W.J.; Crook, T.H. Glutamate: Its role in learning, memory, and the aging brain. Psychopharmacology 1993, 111, 391–401. [Google Scholar]
- Hynd, M.R.; Scott, H.L.; Dodd, P.R. Glutamate-mediated excitotoxicity and neurodegeneration in Alzheimer’s disease. Neurochem. Int 2004, 45, 583–595. [Google Scholar]
- Danbolt, N.C. Glutamate uptake. Prog. Neurobiol 2001, 65, 1–105. [Google Scholar]
- Murphy, T.H.; Miyamoto, M.; Sastre, A.; Schnaar, R.L.; Coyle, J.T. Glutamate toxicity in a neuronal cell line involves inhibition of cystine transport leading to oxidative stress. Neuron 1989, 2, 1547–1558. [Google Scholar]
- Brookmeyer, R.; Gray, S.; Kawas, C. Projections of Alzheimer’s disease in the United States and the public health impact of delaying disease onset. Am. J. Public Health 1998, 88, 1337–1342. [Google Scholar]
- Ansari, M.A.; Abdul, H.M.; Joshi, G.; Opii, W.O.; Butterfield, D.A. Protective effect of quercetin in primary neurons against Abeta(1–42): Relevance to Alzheimer’s disease. J. Nutr. Biochem 2009, 20, 269–275. [Google Scholar]
- Huang, X.; Cuajungco, M.P.; Atwood, C.S.; Hartshorn, M.A.; Tyndall, J.D.; Hanson, G.R.; Stokes, K.C.; Leopold, M.; Multhaup, G.; Goldstein, L.E.; et al. Cu(II) potentiation of alzheimer abeta neurotoxicity. Correlation with cell-free hydrogen peroxide production and metal reduction. J. Biol. Chem 1999, 274, 37111–37116. [Google Scholar]
- Huang, X.; Atwood, C.S.; Hartshorn, M.A.; Multhaup, G.; Goldstein, L.E.; Scarpa, R.C.; Cuajungco, M.P.; Gray, D.N.; Lim, J.; Moir, R.D.; et al. The A beta peptide of alzheimer’s disease directly produces hydrogen peroxide through metal ion reduction. Biochemistry 1999, 38, 7609–7616. [Google Scholar]
- Subarnas, A.; Tadano, T.; Nakahata, N.; Arai, Y.; Kinemuchi, H.; Oshima, Y.; Kisara, K.; Ohizumi, Y. A possible mechanism of antidepressant activity of beta-amyrin palmitate isolated from Lobelia inflata leaves in the forced swimming test. Life Sci 1993, 52, 289–296. [Google Scholar]
- Subarnas, A.; Tadano, T.; Oshima, Y.; Kisara, K.; Ohizumi, Y. Pharmacological properties of beta-amyrin palmitate, a novel centrally acting compound, isolated from Lobelia inflata leaves. J. Pharm. Pharmacol 1993, 45, 545–550. [Google Scholar]
- Subarnas, A.; Oshima, Y.; Sidik; Ohizumi, Y. An antidepressant principle of Lobelia inflata L. (Campanulaceae). J. Pharm. Sci 1992, 81, 620–621. [Google Scholar]
- Aragão, G.F.; Carneiro, L.M.; Junior, A.P.; Vieira, L.C.; Bandeira, P.N.; Lemos, T.L.; Viana, G.S. A possible mechanism for anxiolytic and antidepressant effects of alpha- and beta-amyrin from Protium heptaphyllum (Aubl.) March. Pharmacol. Biochem. Behav 2006, 85, 827–834. [Google Scholar]
- Davis, J.B.; Maher, P. Protein kinase C activation inhibits glutamate-induced cytotoxicity in a neuronal cell line. Brain Res 1994, 652, 169–173. [Google Scholar]
- Fukui, M.; Song, J.-H.; Choi, J.; Choi, H.J.; Zhu, B.T. Mechanism of glutamate-induced neurotoxicity in HT22 mouse hippocampal cells. Eur. J. Pharmacol 2009, 617, 1–11. [Google Scholar]
- Behl, C. Vitamin E protects neurons against oxidative cell death in vitro more effectively than 17-beta estradiol and induces the activity of the transcription factor NF-kappaB. J. Neural Transm 2000, 107, 393–407. [Google Scholar]
- Celsi, F.; Ferri, A.; Casciati, A.; D’Ambrosi, N.; Rotilio, G.; Costa, A.; Volonté, C.; Carrì, M.T. Overexpression of superoxide dismutase 1 protects against beta-amyloid peptide toxicity: Effect of estrogen and copper chelators. Neurochem. Int 2004, 44, 25–33. [Google Scholar]
- Tewtrakul, S.; Tansakul, P.; Panichayupakaranant, P. Effects of rhinacanthins from Rhinacanthus nasutus on nitric oxide, prostaglandin E2 and tumor necrosis factor-alpha releases using RAW264.7 macrophage cells. Phytomed. Int. J. Phytother. Phytopharmacol 2009, 16, 581–585. [Google Scholar]
- Tewtrakul, S.; Tansakul, P.; Panichayupakaranant, P. Anti-allergic principles of Rhinacanthus nasutus leaves. Phytomed. Int. J. Phytother. Phytopharmacol 2009, 16, 929–934. [Google Scholar]
- Behl, C.; Davis, J.B.; Klier, F.G.; Schubert, D. Amyloid β peptide induces necrosis rather than apoptosis. Brain Res 1994, 645, 253–264. [Google Scholar]
- Mook-Jung, I.; Joo, I.; Sohn, S.; Kwon, H.J.; Huh, K.; Jung, M.W. Estrogen blocks neurotoxic effects of beta-amyloid (1–42) and induces neurite extension on B103 cells. Neurosci. Lett 1997, 235, 101–104. [Google Scholar]
- Behl, C.; Trapp, T.; Skutella, T.; Holsboer, F.; Behll, C.; Trappi, T.; Holsboerl, F. Protection against oxidative stress-induced neuronal cell death-a novel role for RU486. Eur. J. Neurosci 1997, 9, 912–920. [Google Scholar]
- Behl, C.; Skutella, T.; Lezoualc’h, F.; Post, A.; Widmann, M.; Newton, C.J.; Holsboer, F. Neuroprotection against oxidative stress by estrogens: Structure-activity relationship. Mol. Pharmacol 1997, 51, 535–541. [Google Scholar]
- Gursoy, E.; Cardounel, A.; Al-Khlaiwi, T.; Al-Drees, A.; Kalimi, M. Tamoxifen protects clonal mouse hippocampal (HT-22) cells against neurotoxins-induced cell death. Neurochem. Int 2002, 40, 405–412. [Google Scholar]
- Behl, C. Oestrogen as a neuroprotective hormone. Nat. Rev. Neurosci 2002, 3, 433–442. [Google Scholar]
- Wu, T.S.; Hsu, H.C.; Wu, P.L.; Teng, C.M.; Wu, Y.C. Rhinacanthin-Q, a naphthoquinone from Rhinacanthus nasutus and its biological activity. Phytochemistry 1998, 49, 2001–2003. [Google Scholar]
- Wu, T.S.; Yang, C.C.; Wu, P.L.; Liu, L.K. A quinol and steroids from the leaves and stems of Rhinacanthus nasutus. Phytochemistry 1995, 40, 1247–1249. [Google Scholar]
- Subarnas, A.; Tadano, T.; Kisara, K.; Ohizumi, Y. An alpha-adrenoceptor-mediated mechanism of hypoactivity induced by beta-amyrin palmitate. J. Pharm. Pharmacol 1993, 45, 1006–1008. [Google Scholar]
- Madrigal, J.L.M.; Kalinin, S.; Richardson, J.C.; Feinstein, D.L. Neuroprotective actions of noradrenaline: Effects on glutathione synthesis and activation of peroxisome proliferator activated receptor delta. J. Neurochem 2007, 103, 2092–2101. [Google Scholar]
- Gallo, M.B.C.; Sarachine, M.J. Biological activities of Lupeol. Int. J. Biomed. Pharm. Sci 2009, 2, 46–66. [Google Scholar]
- Pike, C.J.; Burdick, D.; Walencewicz, A.J.; Glabe, C.G.; Cotman, C.W. Neurodegeneration Induced by beta Amyloid Peptide in vitro: The role of peptide assembly state. J. Neurosci 1993, 13, 1676–1687. [Google Scholar]
- Stanojević, L.; Stanković, M.; Nikolić, V.; Nikolić, L.; Ristić, D.; Čanadanovic-Brunet, J.; Tumbas, V. Antioxidant activity and total phenolic and flavonoid contents of Hieracium pilosella L. extracts. Sensors 2009, 9, 5702–5714. [Google Scholar]
- Murali, A.; Ashok, P.; Madhavan, V. In vitro antiocidant activity and HPTLC studies on the roots and rhizomes of Smilax zeylanica L. (Smilacaceae). Int. J. Pharm. Pharm. Sci 2011, 3, 1–4. [Google Scholar]
- Jiao, Z.; Liu, J. Antioxidant activities of total pigment extract from blackberries. Food Technol. Biotechnol 2005, 43, 97–102. [Google Scholar]
- Balakrishnan, N.; Panda, A.B.; Raj, N.R.; Shrivastava, A.; Prathani, R. The evaluation of nitric oxide scavenging activity of acalypha indica linn root. Asian J. Res. Chem 2009, 2, 148–150. [Google Scholar]
- Murali, A.; Ashok, P.; Madhavan, V. Free radical scavenging effect of leaf of Hermidesmus indicus (L.) R. BR. Var (W. & A.) HK.F. (Periplocaceae)—An in vitro analysis: A research. Int. J. Drug Formul. Res 2010, 1, 162–175. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| R. nasutus Extract | Hydroxyl Scavenging Assay | n | ABTS+ Decolorization Assay | n | DPPH Scavenging Assay | N | NO− Scavenging Assay | n | SO Scavenging Assay | n |
|---|---|---|---|---|---|---|---|---|---|---|
| Root Ethanol | 4.5 ± 0.38 | 3 | 3.49 ± 0.07 | 3 | 3.5 ± 0.15 | 4 | <2 | 2 | 3.25 ± 0.22 | 3 |
| Root Hexane | ND | 3 | 3.28 ± 0.43 | 3 | 1.9 ± 0.01 | 5 | <2 | 2 | 2.81 ± 0.03 | 3 |
| Leaf Ethanol | 4.4 ± 0.27 | 3 | 4.04 ± 0.06 | 3 | 4.4 ± 0.05 | 4 | <2 | 2 | 4.49 ± 0.07 | 3 |
| Leaf Hexane | ND | 3 | 2.30 ± 2.40 | 3 | 0.5 ± 0.10 | 3 | <2 | 2 | 2.31 ± 0.20 | 3 |
| Trolox | 3.2 ± 0.08 | 3 | 3.60 ± 0.02 | 3 | 4.9 ± 0.02 | 5 | NA | 0 | NA | 0 |
| L-Ascorbic Acid | 4.3 ± 0.15 | 3 | 3.86 ± 0.10 | 3 | 6.1 ± 0.11 | 3 | ± | 2 | 6.14 ± 0.05 | 3 |
| R. nasutus Extract | Total Phenolic Content GAE mg·g−1 Dry Extract | n | Total Flavonoid Content RE mg·g−1 Dry Extract | n |
|---|---|---|---|---|
| Root Ethanol | 2.17 ± 0.15 | 5 | 1.13 ± 0.17 | 4 |
| Root Hexane | 0.36 ± 0.11 | 5 | ND | 4 |
| Leaf Ethanol | 6.50 ± 0.12 | 5 | 1.15 ± 0.14 | 4 |
| Leaf Hexane | 0.40 ± 0.01 | 5 | ND | 4 |
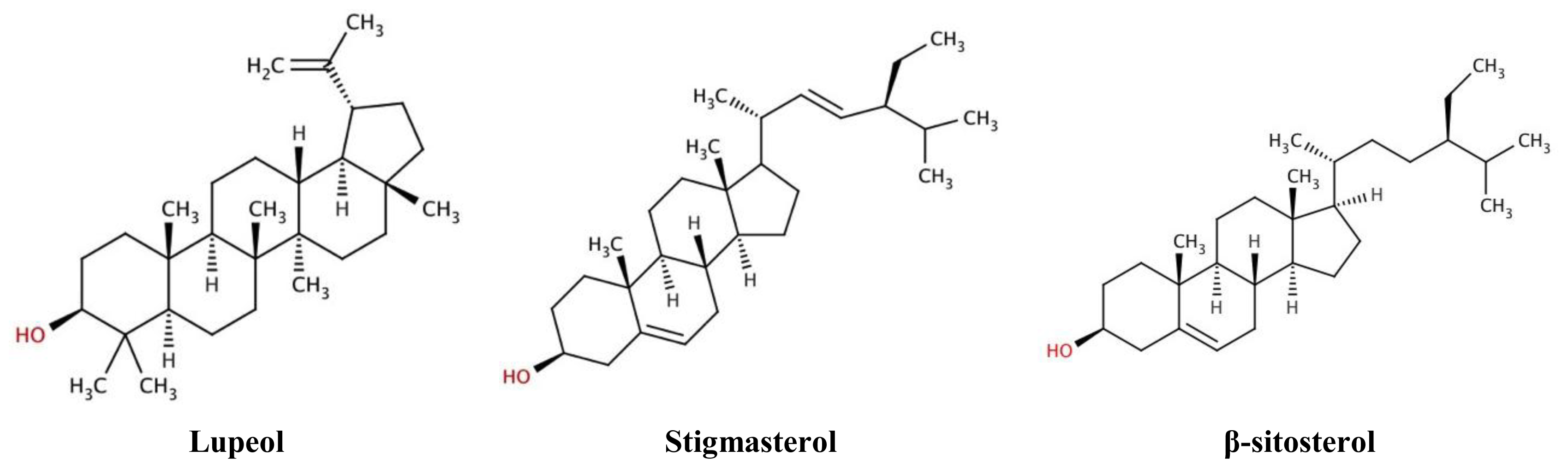
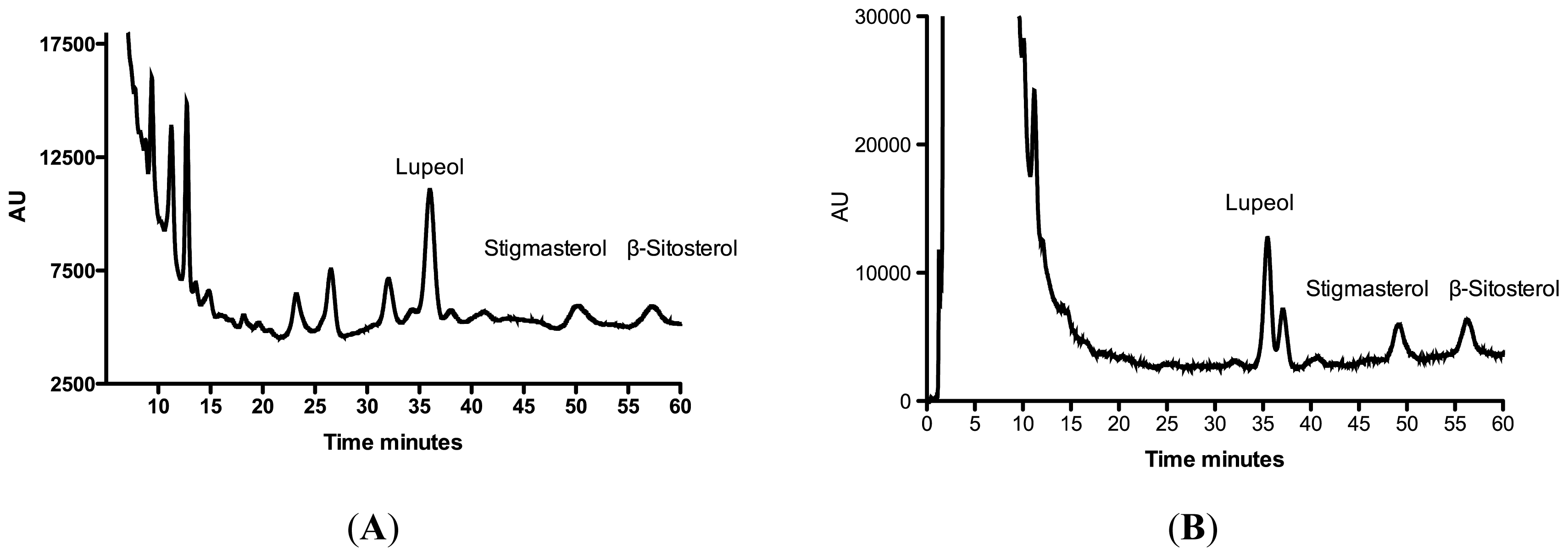
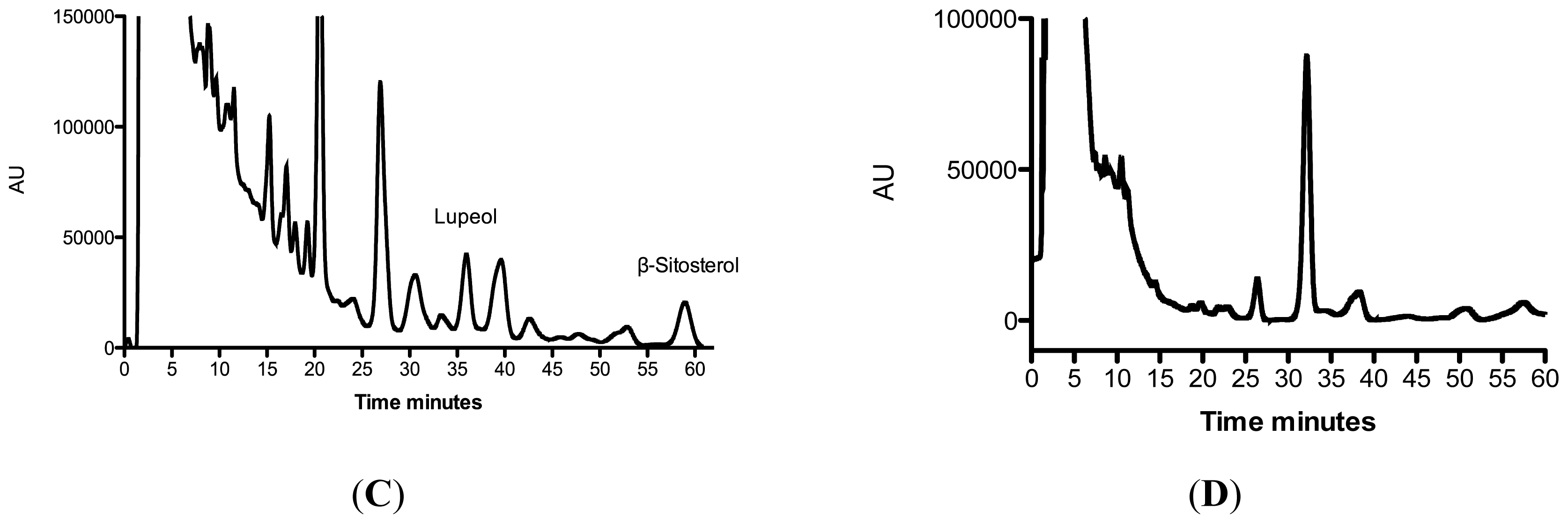
| R. nasutus Extract | Lupeol μg·mg−1 Extract | Stigmasterol μg·mg−1 Extract | β-sitosterol μg·mg−1 Extract |
|---|---|---|---|
| Root ethanol | 5.4 ± 0.3 | 13.7 ± 0.1 | 0.5 ± 0.1 |
| Root hexane | 1.5 ± 0.3 | 2.91 ± 0.03 | 1.3 ± 0.2 |
| Leaf ethanol | 12.4 ± 2.5 | ND | 17.4 ± 3.2 |
| Leaf hexane | ND | ND | ND |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Brimson, J.M.; Brimson, S.J.; Brimson, C.A.; Rakkhitawatthana, V.; Tencomnao, T. Rhinacanthus nasutus Extracts Prevent Glutamate and Amyloid-β Neurotoxicity in HT-22 Mouse Hippocampal Cells: Possible Active Compounds Include Lupeol, Stigmasterol and β-Sitosterol. Int. J. Mol. Sci. 2012, 13, 5074-5097. https://doi.org/10.3390/ijms13045074
Brimson JM, Brimson SJ, Brimson CA, Rakkhitawatthana V, Tencomnao T. Rhinacanthus nasutus Extracts Prevent Glutamate and Amyloid-β Neurotoxicity in HT-22 Mouse Hippocampal Cells: Possible Active Compounds Include Lupeol, Stigmasterol and β-Sitosterol. International Journal of Molecular Sciences. 2012; 13(4):5074-5097. https://doi.org/10.3390/ijms13045074
Chicago/Turabian StyleBrimson, James M., Sirikalaya J. Brimson, Christopher A. Brimson, Varaporn Rakkhitawatthana, and Tewin Tencomnao. 2012. "Rhinacanthus nasutus Extracts Prevent Glutamate and Amyloid-β Neurotoxicity in HT-22 Mouse Hippocampal Cells: Possible Active Compounds Include Lupeol, Stigmasterol and β-Sitosterol" International Journal of Molecular Sciences 13, no. 4: 5074-5097. https://doi.org/10.3390/ijms13045074
APA StyleBrimson, J. M., Brimson, S. J., Brimson, C. A., Rakkhitawatthana, V., & Tencomnao, T. (2012). Rhinacanthus nasutus Extracts Prevent Glutamate and Amyloid-β Neurotoxicity in HT-22 Mouse Hippocampal Cells: Possible Active Compounds Include Lupeol, Stigmasterol and β-Sitosterol. International Journal of Molecular Sciences, 13(4), 5074-5097. https://doi.org/10.3390/ijms13045074