The Hypoglycemic Effect of the Kelp on Diabetes Mellitus Model Induced by Alloxan in Rats

Abstract
:1. Introduction
2. Results and Discussion
2.1. General Situation
2.2. The Level of Fasting Blood Glucose (FBG)
2.3. The Serum Level of Insulin
2.4. The Levels of MDA and NO and the Activities of SOD and GSH-Px
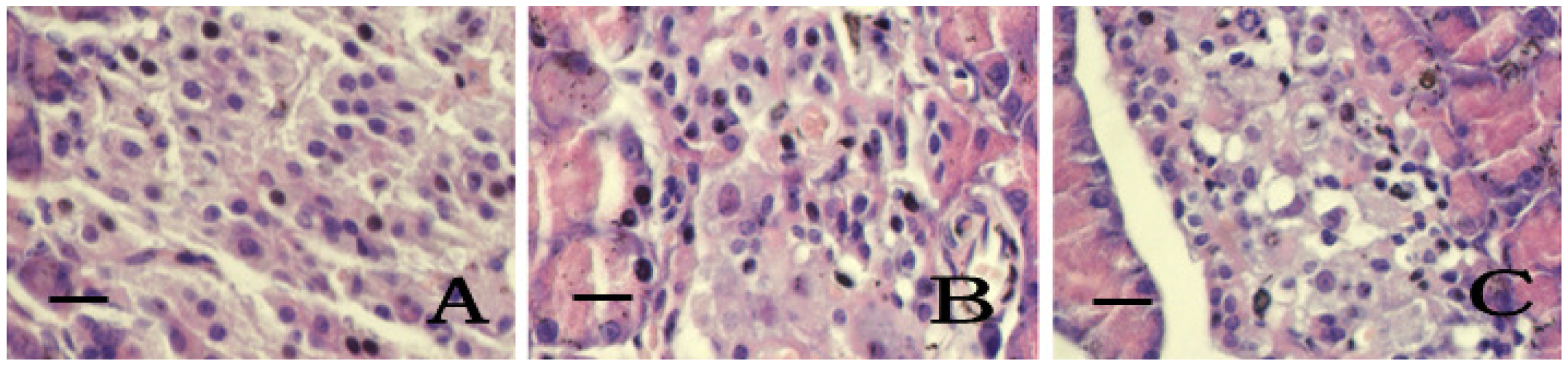
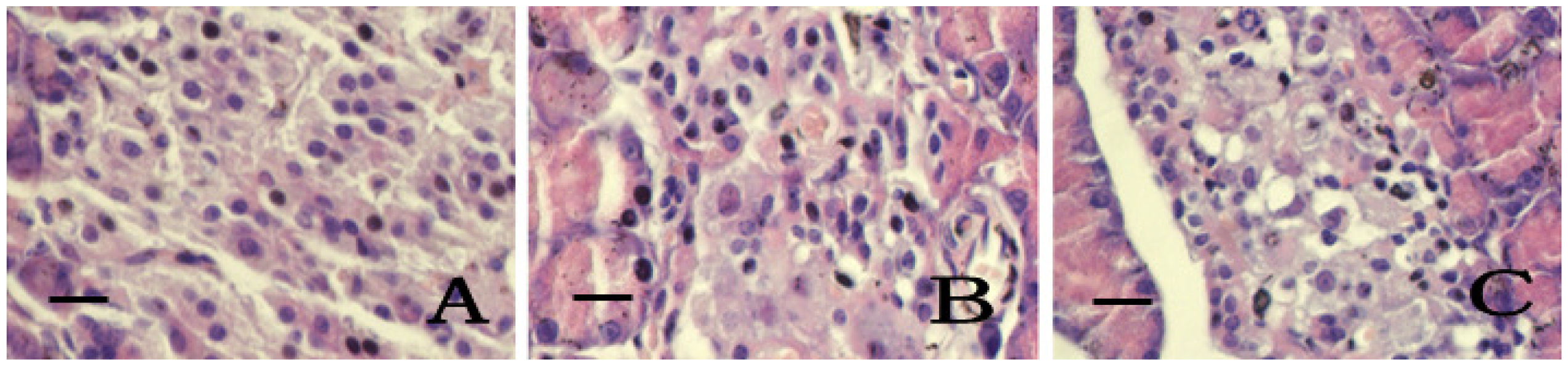
2.5. Pancreatic Islets Tissue Pathology
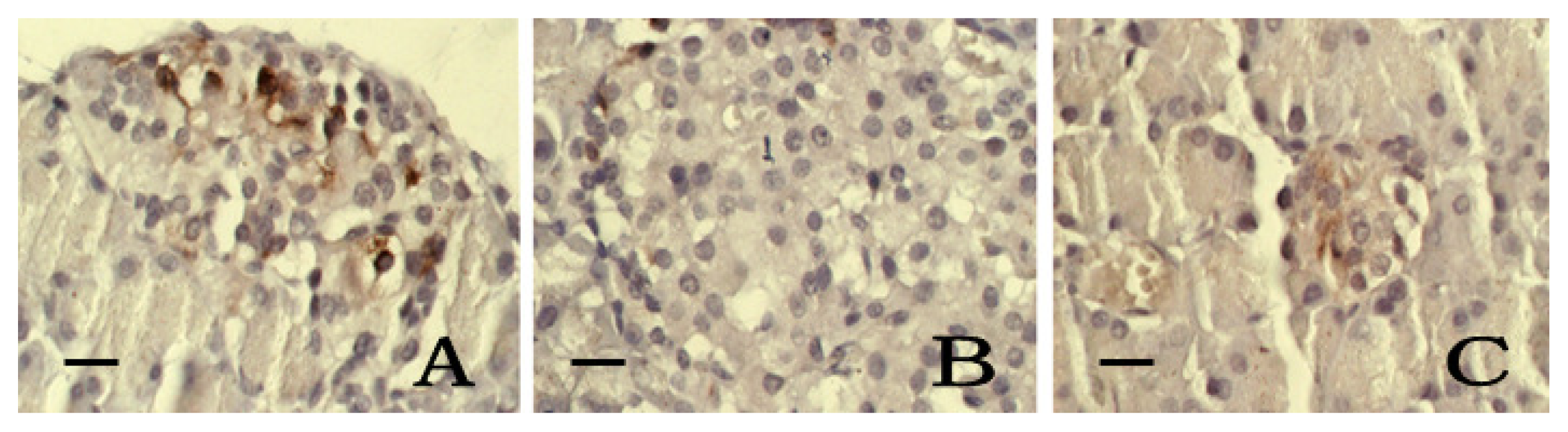
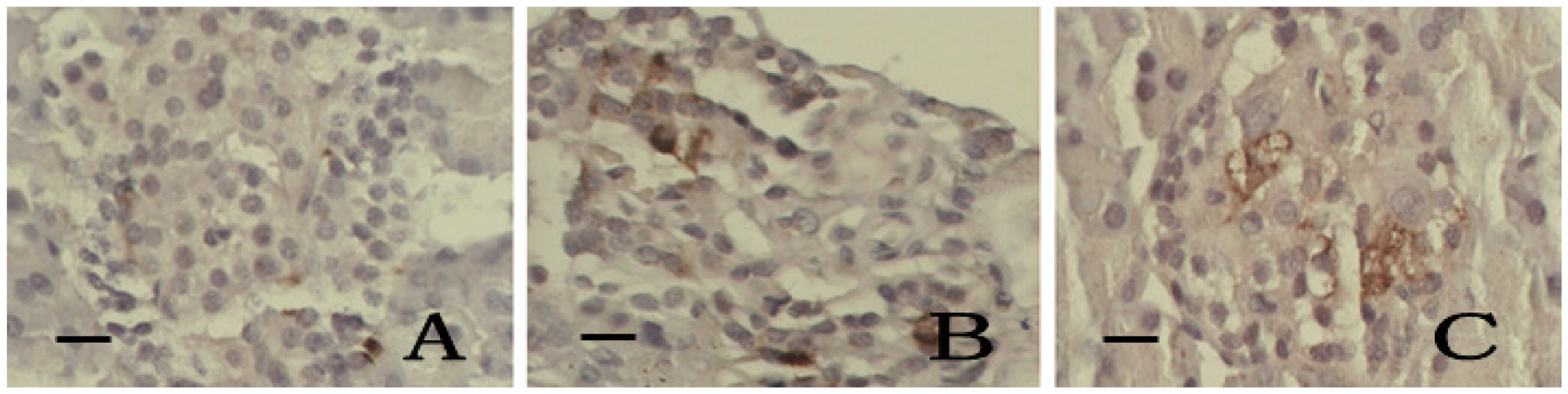
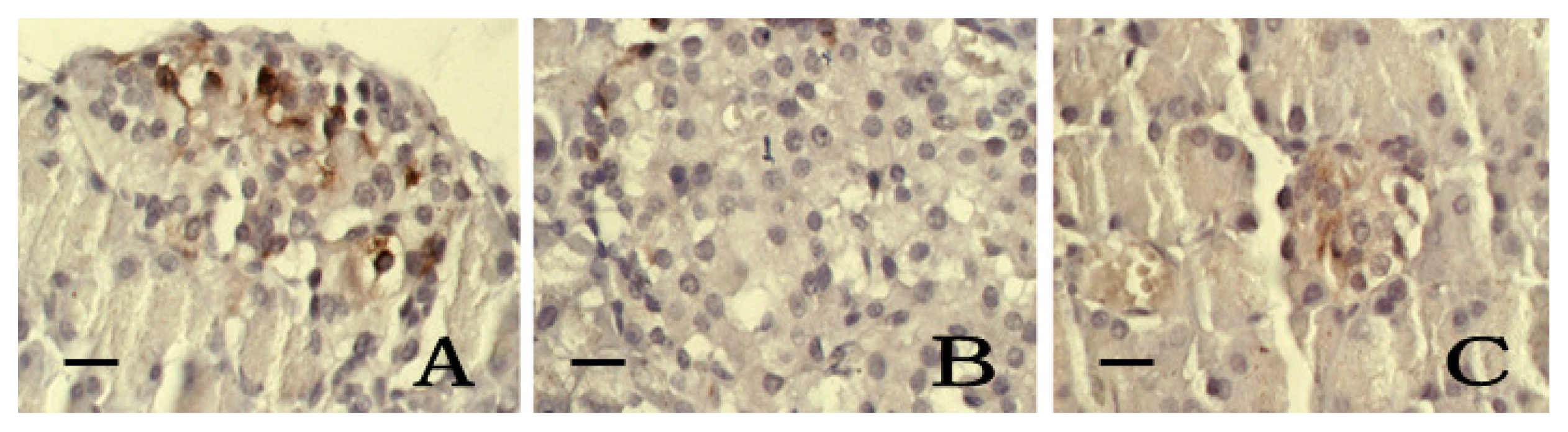
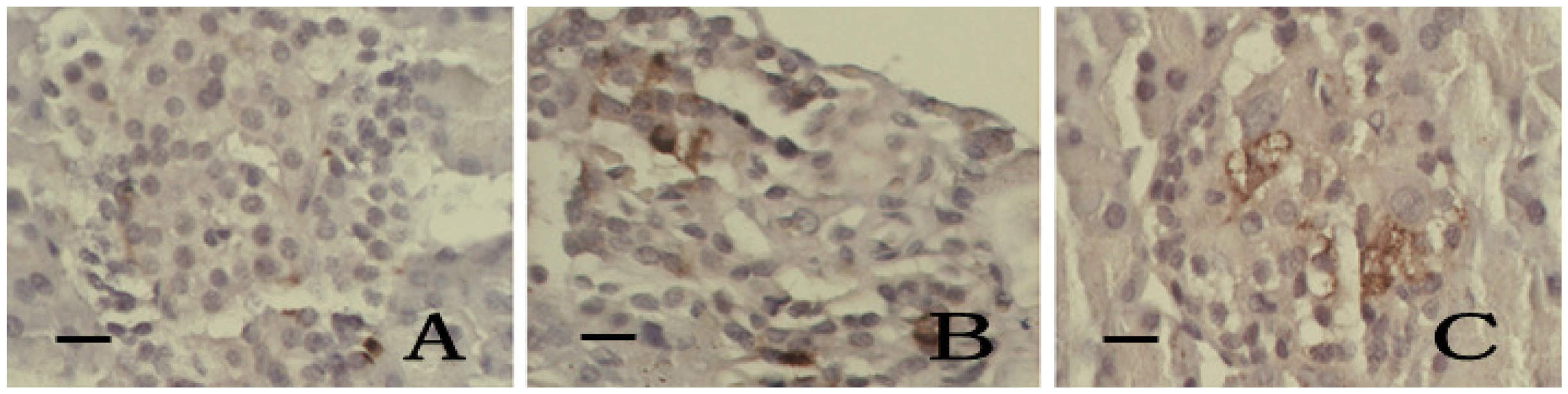
2.6. Immunohistochemistry of SOD and iNOS
2.7. Discussion
3. Experimental Section
3.1. The Creation of Diabetic Models
3.1. The General Forage and Kelp Powder Forage
3.2. Inference of Tests
3.3. Preparation of Samples
3.3.1. Serum preparation
3.3.2. Pancreatic tissue
3.4. Index of Determinations
3.4.1. FBG Level
3.4.2. The Serum Level of Insulin
3.4.3. MDA and NO Values
3.4.4. SOD and GSH-Px Activities
3.4.5. Histopathological Assay
3.4.6. Immunohistochemical Assay
3.5. Statistical Analysis
4. Conclusions
Acknowledgements
- Conflict of InterestThis article does not compromise our adherence to Int. J. Mol. Sci. policies on sharing data and materials.
References
- Shih, C.C.; Wu, Y.W.; Lin, W.C. Anti-hyperglycaemic and anti-oxidant properties of anoectochilus formosanus in diabetic rats. Clin. Exp. pharmacol. Physiol 2002, 29, 684–688. [Google Scholar]
- Surapaneni, K.M.; Venkataramana, G. Status of lipid peroxidation, glutathione, ascorbic acid, vitamin E and antioxidant enzymes in patients with osteoarthritis. Indian J. Med. Sci 2007, 61, 9–14. [Google Scholar]
- Bian, K.; Ke, Y.; Kamisaki, Y.; Murad, F. Proteomic modification by nitric oxide. Pharmacol. Sci 2006, 101, 271–279. [Google Scholar]
- Challa, S.R.; Akula, A.; Metla, S.; Gopal, P.N. Partial role of nitric oxide in infarct size limiting effect of quercetin and rutin against ischemia-reperfusion injury in normal and diabetic rats. Indian J. Exp. Biol 2011, 49, 207–210. [Google Scholar]
- Aguilar, A.; Alvarez-Vijande, R.; Capdevila, S.; Alcoberro, J; Alcaraz, A. Antioxidant patterns (superoxide dismutase, glutathione reductase and glutathione peroxidase) in kidneys from non-heart-beating-donors: Experimental study. Transpl. Proc 2007, 39, 249–252. [Google Scholar]
- Chung, S.S.; Kim, M.; Youn, B.S.; Lee, N.S.; Park, J.W.; Lee, I.K.; Lee, Y.S.; Kim, J.B.; Cho, Y.M.; Lee, H.K.; et al. Glutathione peroxidase mediates the antioxidant effect of peroxisome proliferator-activated receptor in human skeletal muscle cells. Mol. Cell Biol 2009, 29, 20–30. [Google Scholar]
- Tseng, CK. Algal biotechnology industries and research activities in China. J. Appl. Phycol 2001, 13, 375–380. [Google Scholar]
- Huang, L.; Guo, H.W.; Huang, Y.; Xue, K. The association of Lipoprotein lipase gene polymorphism with hyperlipidemia and dietary predisposition of obesity. Acta Nutr. Sin 2007, 29, 228–231. [Google Scholar]
- Chinese Pharmcopoeia Commission, Chinese Pharmacopoeia, Part I, 2000 ed; Chemical Industry Press: Beijing, China, 2000; pp. 168–170.
- Zhu, L.; Zhang, Q.; Wang, Y.F.; Kang, Y.Y.; Cen, Y.Z. Determination of polysaccharide from Ecklonia kurome. Chin. J. Mar. Drugs 2005, 24, 47–48. [Google Scholar]
- Zhou, Q.F.; Li, M.Y.; Na, G.S.; Li, C.W. Progress in research of anti-tumor mechanisms of marine polysaccharides. Chin. Pharmacol. Bull 2009, 25, 995–997. [Google Scholar]
- Xu, X.Y.; Shuai, L.; Guo, Y.L.; Duan, D.L. Regulating effects and mechanism of Laminaria Japonica on serum lipid of hyperlipidemia in rats. Chin. J. Mar. Drugs 2009, 28, 1–4. [Google Scholar]
- Shuai, L.; Xu, X.Y.; Guo, Y.L.; Duan, D.L. Study on the antioxidant effects of Laminaria japonica in hyperlipemia rats. Chin. J. Mar. Drugs 2010, 29, 1–4. [Google Scholar]
- Wang, T.X.; Wang, T.X.; Pang, J.H. Study on the hypoglycemic and hypolipidemic effect of Laminarina japonica polysaccharides. Acta Nutr. Sin 2007, 29, 99–100. [Google Scholar]
- Ceriello, A.; Motz, E. Is oxidative stress the pathogenic mechanism underlying insulin resistance, diabetes, and cardiovascular disease? The common soil hypothesis revisited. Arterioscler. Thromb. Vasc. Biol 2004, 24, 816–823. [Google Scholar]
- Capellini, V.K.; Baldo, C.F.; Celotto, A.C.; Batalhão, M.E.; Cárnio, E.C.; Rodrigues, A.J.; Evora, P.R. Oxidative stress is not associated with vascular dysfunction in a model of alloxan-induced diabetic rats. Arq. Bras. Endocrinol. Metabol 2010, 54, 530–539. [Google Scholar] [Green Version]
- Van Guilder, H.D.; Bixler, G.V.; Kutzler, L.; Brucklacher, R.M.; Bronson, S.K.; Kimball, S.R.; Freeman, W.M. Multi-modal proteomic analysis of retinal protein expression alterations in a rat model of diabetic retinopathy. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Mircescu, G. Oxidative stress: An accomplice to uremic toxicity? Ren Nutr 2006, 16, 194–198. [Google Scholar]
- Lushchak, V.I. Free radical oxidation of proteins and its relationship with functional state of organisms. Biol. Chem. (Mosc.) 2007, 72, 809–827. [Google Scholar]
- Zhou, T.; Zhou, K.K.; Lee, K.; Gao, G.; Lyons, T.J.; Kowluru, R.; Ma, J.-X. The role of lipid peroxidation products and oxidative stress in activation of the canonical wingless-type MMTV integration site (WNT) pathway in a rat model of diabetic retinopathy. Diabetologia 2011, 54, 459–468. [Google Scholar]
- Chaiyasut, C.; Kusirisin, W.; Lailerd, N.; Lerttrakarnnon, P.; Suttajit, M.; Srichairatanakool, S. Effects of phenolic compounds of fermented thai indigenous plants on oxidative stress in streptozotocin-induced diabetic rats. Evid. Based Complement. Altern 2011, 2011, 74–77. [Google Scholar]
- Kröncke, K.D.; Fehsel, K.; Suschek, C.; Kolb-Bachofen, V. Inducible nitric oxide synthesis-derived nitric oxide in gene regulation, cell death and cell survival. Int. Immunopharmacol 2001, 1, 1407–1420. [Google Scholar]
- Chapman, J.; Miles, P.D.; Ofrecio, J.M.; Neels, J.G.; Yu, J.G.; Resnik, J.L.; Wilkes, J.; Talukdar, S.; Thapar, D.; Johnson, K.; et al. Osteopontin is required for the early onset of high fat diet-induced insulin resistance in mice. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Soto, C.; Pérez, J.; García, V.; Uría, E.; Vadillo, M.; Raya, L. Effect of silymarin on kidneys of rats suffering from alloxan-induced diabetes mellitus. Phytomedicine 2010, 17, 1090–1094. [Google Scholar]
- Silva, K.C.; Rosales, M.A.; Biswas, S.K.; Lopes de Faria, J.B.; Lopes de Faria, J.M. Diabetic retinal neurodegeneration is associated with mitochondrial oxidative stress and is improved by an angiotensin receptor blocker in a model combining hypertension and diabetes. Diabetes 2009, 58, 1382–1390. [Google Scholar]
- Ouslimani, N.; Peynet, J.; Bonnefont-Rousselot, D.; Thérond, P.; Legrand, A.; Beaudeux, J.L. Metfrom in decreases intracellular production of reactive oxygen species aortic endothelial cells. Metabolism 2005, 54, 829–834. [Google Scholar]
- Bottino, R.; Balamurugan, A.N.; Tse, H.; Thirunavukkarasu, C.; Ge, X.; Profozich, J.; Milton, M.; Ziegenfuss, A.; Trucco, M.; Piganelli, J.D. Response of human islets to isolation stress and the effect of antioxidant treatment. Diabetes 2004, 53, 2559–2568. [Google Scholar]
- Liu, L.P.; Huang, J.; Chen, B.L.; Wang, J.; Li, M. Research on the hypoglycemic effect of marine alga Porphyridium cruentum in experimental diabetic mice. Chin. J. Mar. Drugs 2005, 24, 18–20. [Google Scholar]
- Owu, D.U.; Antai, A.B.; Udofia, K.H.; Obembe, A.O.; Obasi, K.O.; Eteng, M.U. Vitamin C improves basal metabolic rate and lipid profile in alloxan-induced diabetes mellitus in rats. J. Biosci 2006, 31, 575–579. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Groups | n | Dose | Before experiment | Before kelp-treated | After kelp-treated |
|---|---|---|---|---|---|
| Control group | 10 | General forage | 151.76 ± 3.45 | 168.50 ± 4.22 | 189.69 ± 4.55 |
| DM-model group | 10 | General forage | 151.85 ± 3.67 | 133.62 ± 5.35 a | 139.46 ± 5.36 |
| Low-dose group | 10 | 1.25 g/kg kelp | 151.68 ± 3.38 | 134.46 ± 5.23 a | 141.24 ± 5.32 bc |
| Medium-dose group | 10 | 5.0 g/kg kelp | 152.26 ± 3.51 | 135.27 ± 5.18 a | 150.24 ± 5.45 b,c,d |
| High-dose group | 10 | 12.5 g/kg kelp | 151.65 ± 3.43 | 133.55 ± 5.27 a | 151.56 ± 5.67 b,c,d |
| Groups | n | Dose | Before experiment | Before kelp-treated | After kelp-treated |
|---|---|---|---|---|---|
| Control group | 10 | General forage | 4.78 ± 0.39 | 4.95 ± 0.34 | 4.97 ± 0.33 |
| DM-model group | 10 | General forage | 4.82 ± 0.33 | 17.86 ± 2.26 a | 13.32 ± 1.40 b |
| Low-dose group | 10 | 1.25 g/kg kelp | 4.55 ± 0.35 | 18.12 ± 2.28 a | 12.63 ± 1.67 b |
| Medium-dose group | 10 | 5.0 g/kg kelp | 4.81 ± 0.37 | 17.79 ± 2.31 a | 9.37 ± 1.70b,c,d |
| High-dose group | 10 | 12.5 g/kg kelp | 4.65 ± 0.34 | 18.05 ± 2.35 a | 9.18 ± 1.65b,c,d |
| Groups | n | Dose | MDA(mmol/L) | NO(μmol/L) | SOD(U/mL) | GSH-Px(U/mL) |
|---|---|---|---|---|---|---|
| Control group | 10 | General forage | 7.15 ± 0.68 | 14.96 ± 1.56 | 156 ± 14.02 | 922 ± 22.16 |
| DM-model group | 10 | General forage | 9.38 ± 1.24a | 23.86 ± 2.17 a | 122 ± 11.26 a | 828 ± 15.46 a |
| Low-dose group | 10 | 1.25 g/kg kelp | 8.93 ± 1.02 | 21.50 ± 2.24 | 127 ± 18.35 | 837 ± 24.82 |
| Medium-dose group | 10 | 5.0 g/kg kelp | 8.02 ± 0.45 b,c | 17.13 ± 1.41 b,c | 143 ± 22.26 b,c | 890 ± 24.58 b,c |
| High-dose group | 10 | 12.5 g/kg kelp | 7.83 ± 0.51b,c | 16.32 ± 1.73 b,c | 145 ± 19.38 b,c | 886 ± 25.72 b,c |
| Groups | n | Dose | B cell index(%) | SOD (A) | iNOS(A) |
|---|---|---|---|---|---|
| Control group | 10 | General forage | 61.48 ± 9.13 | 0.48 ± 0.15 | 0.16 ± 0.05 |
| DM-model group | 10 | General forage | 28.16 ± 5.64 a | 0.41 ± 0.12 a | 0.41 ± 0.12 a |
| Low-dose group | 10 | 1.25g/kg kelp | 31.49 ± 6.28 | 0.22 ± 0.08 | 0.35 ± 0.10 |
| Medium-dose group | 10 | 5.0g/kg kelp | 45.37 ± 6.82 b | 0.31 ± 0.10 b | 0.24 ± 0.09 b |
| High-dose group | 10 | 12.5g/kg kelp | 46.71 ± 7.36 b | 0.33 ± 0.12 b | 0.23 ± 0.08 b |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Long, S.-H.; Yu, Z.-Q.; Shuai, L.; Guo, Y.-L.; Duan, D.-L.; Xu, X.-Y.; Li, X.-D. The Hypoglycemic Effect of the Kelp on Diabetes Mellitus Model Induced by Alloxan in Rats. Int. J. Mol. Sci. 2012, 13, 3354-3365. https://doi.org/10.3390/ijms13033354
Long S-H, Yu Z-Q, Shuai L, Guo Y-L, Duan D-L, Xu X-Y, Li X-D. The Hypoglycemic Effect of the Kelp on Diabetes Mellitus Model Induced by Alloxan in Rats. International Journal of Molecular Sciences. 2012; 13(3):3354-3365. https://doi.org/10.3390/ijms13033354
Chicago/Turabian StyleLong, Shao-Hua, Zhu-Qin Yu, Li Shuai, Yun-Liang Guo, De-Lin Duan, Xin-Ying Xu, and Xiao-Dan Li. 2012. "The Hypoglycemic Effect of the Kelp on Diabetes Mellitus Model Induced by Alloxan in Rats" International Journal of Molecular Sciences 13, no. 3: 3354-3365. https://doi.org/10.3390/ijms13033354
APA StyleLong, S.-H., Yu, Z.-Q., Shuai, L., Guo, Y.-L., Duan, D.-L., Xu, X.-Y., & Li, X.-D. (2012). The Hypoglycemic Effect of the Kelp on Diabetes Mellitus Model Induced by Alloxan in Rats. International Journal of Molecular Sciences, 13(3), 3354-3365. https://doi.org/10.3390/ijms13033354