Antifungal Activity of (KW)n or (RW)n Peptide against Fusarium solani and Fusarium oxysporum


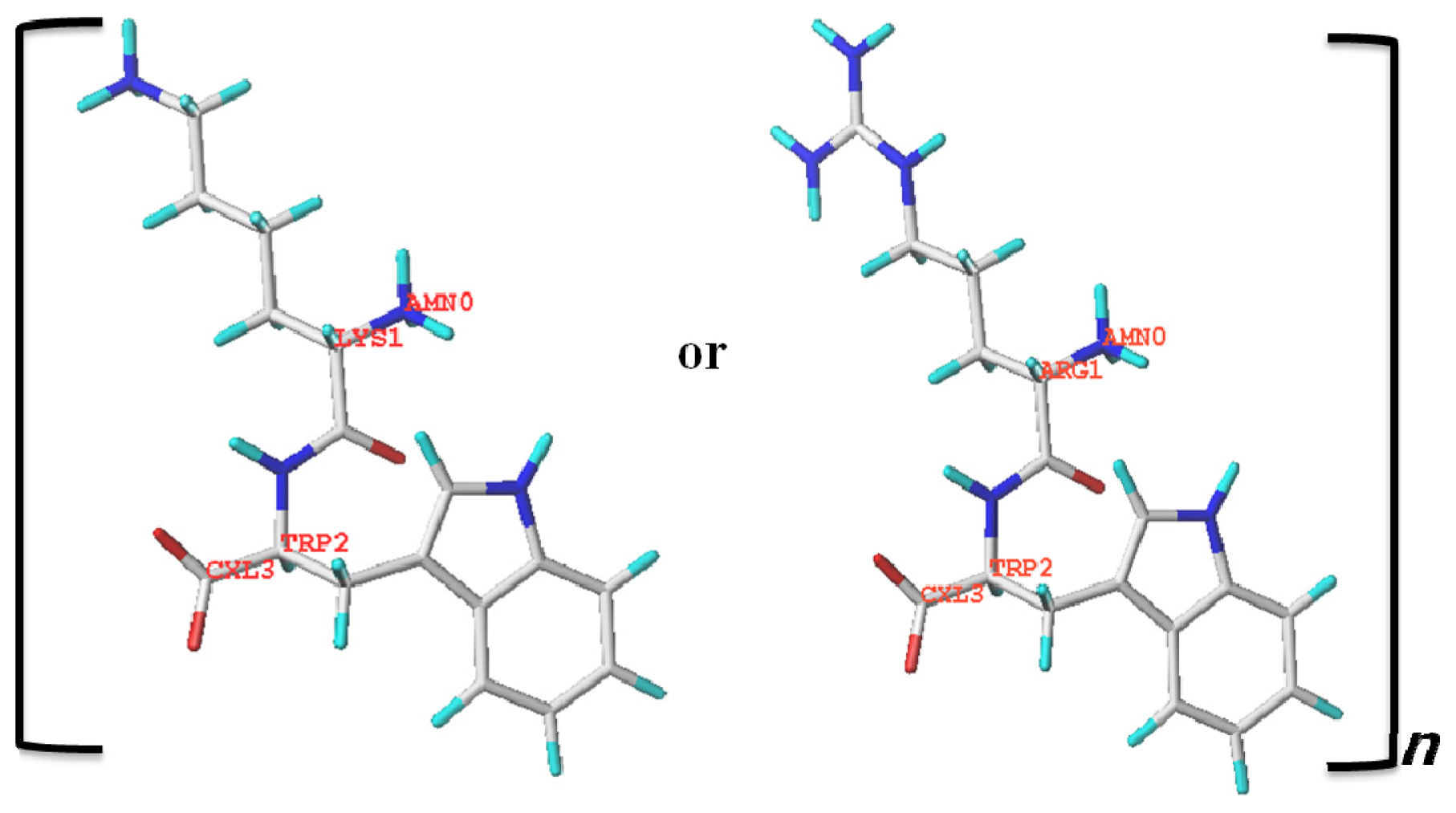{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
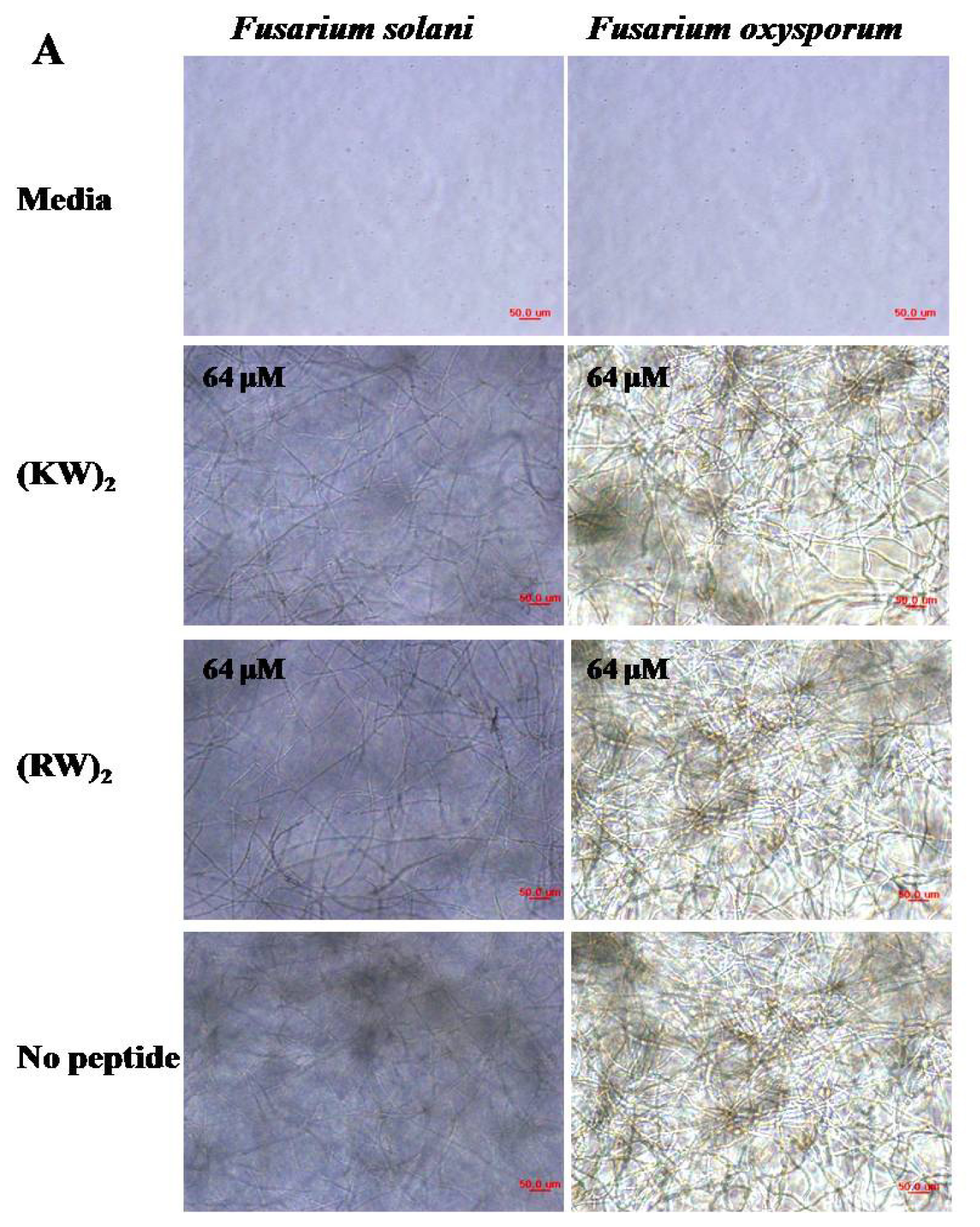
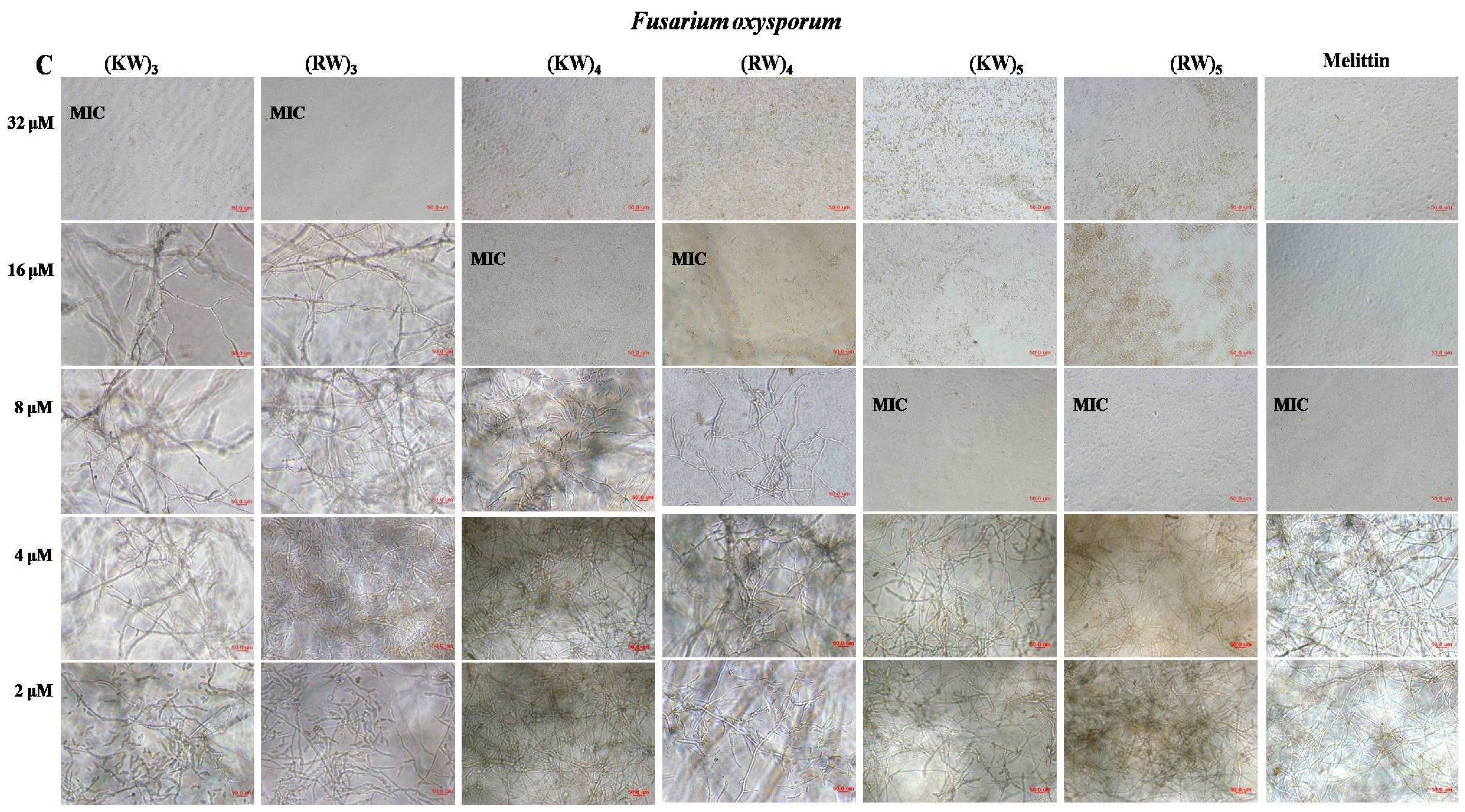
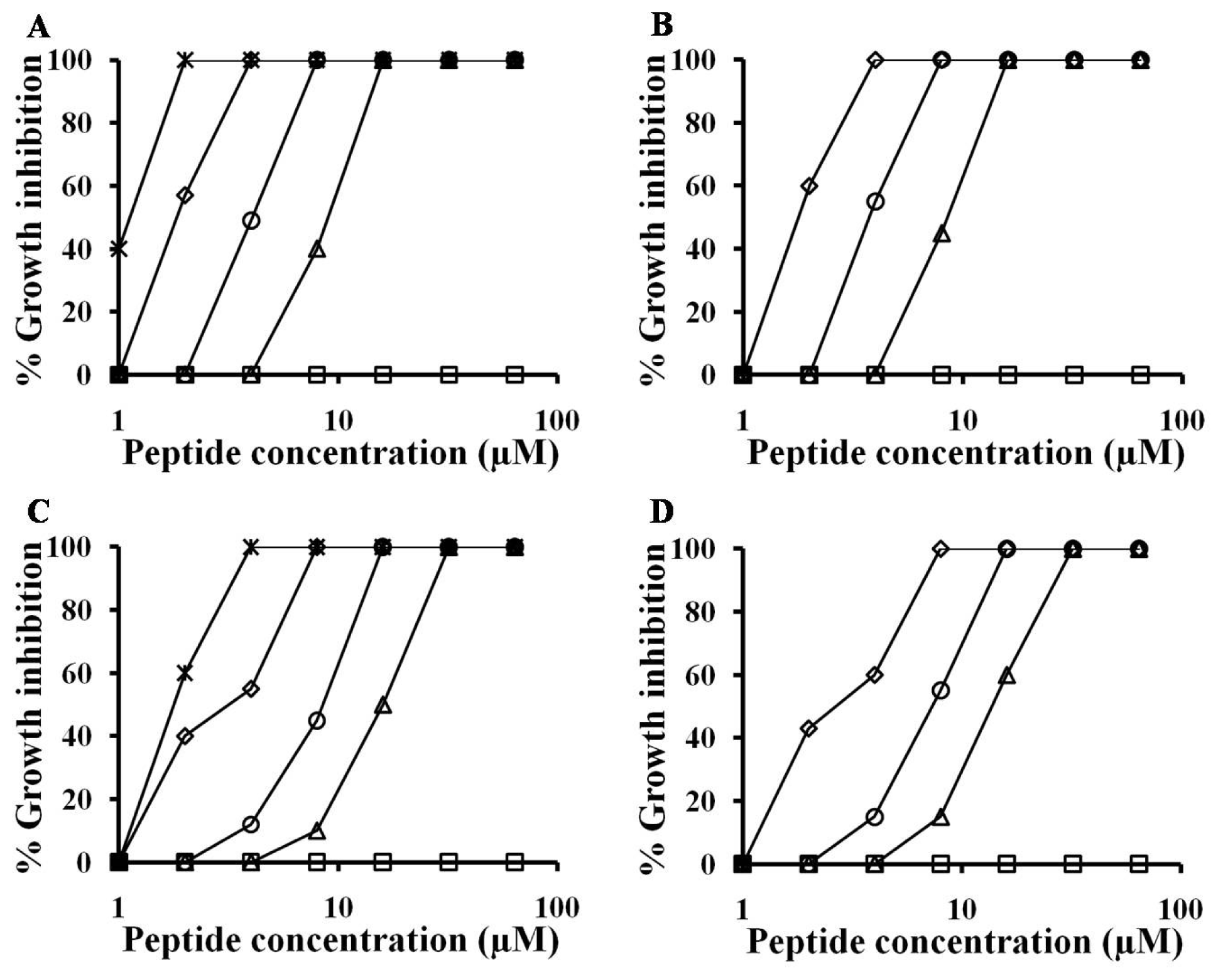
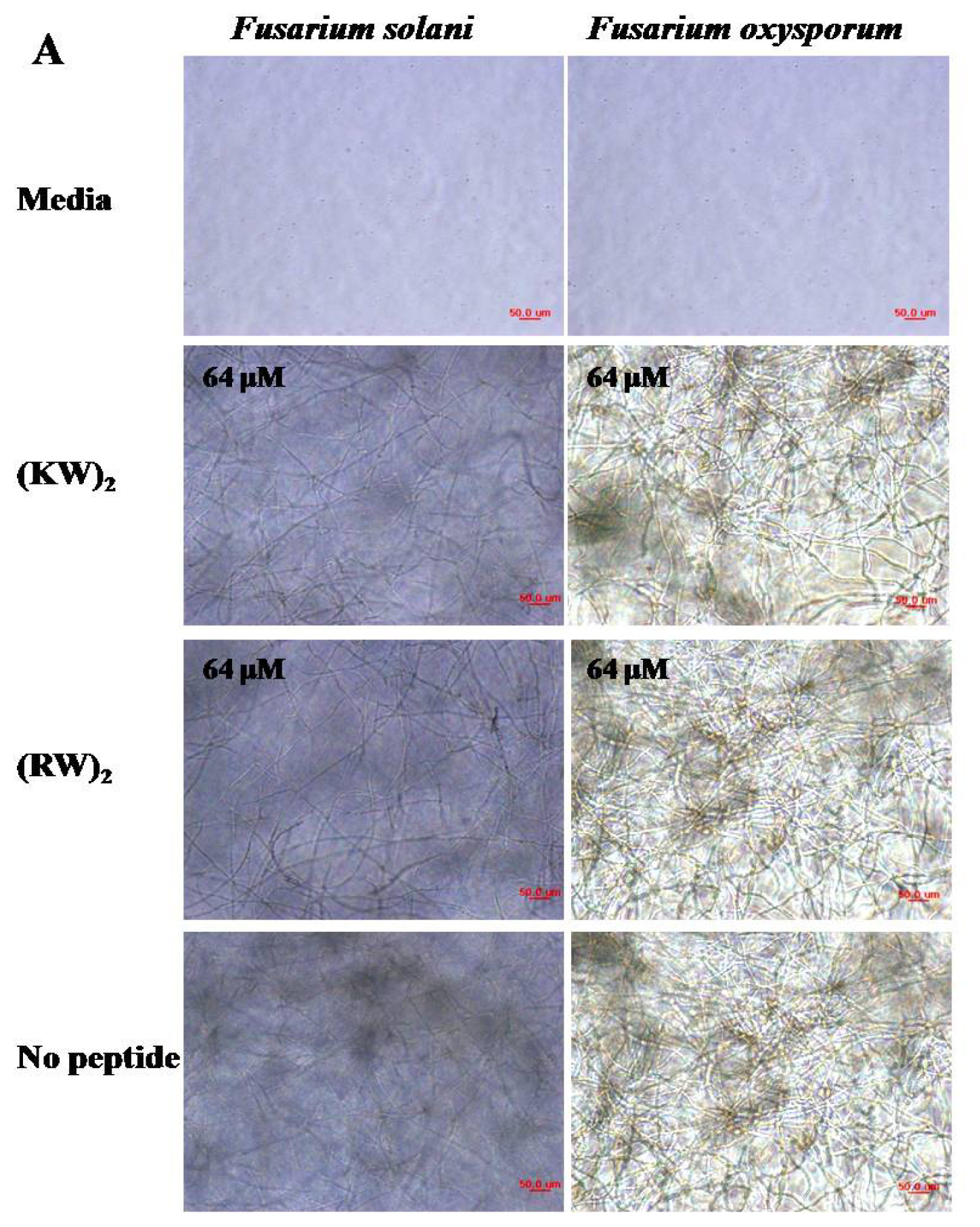
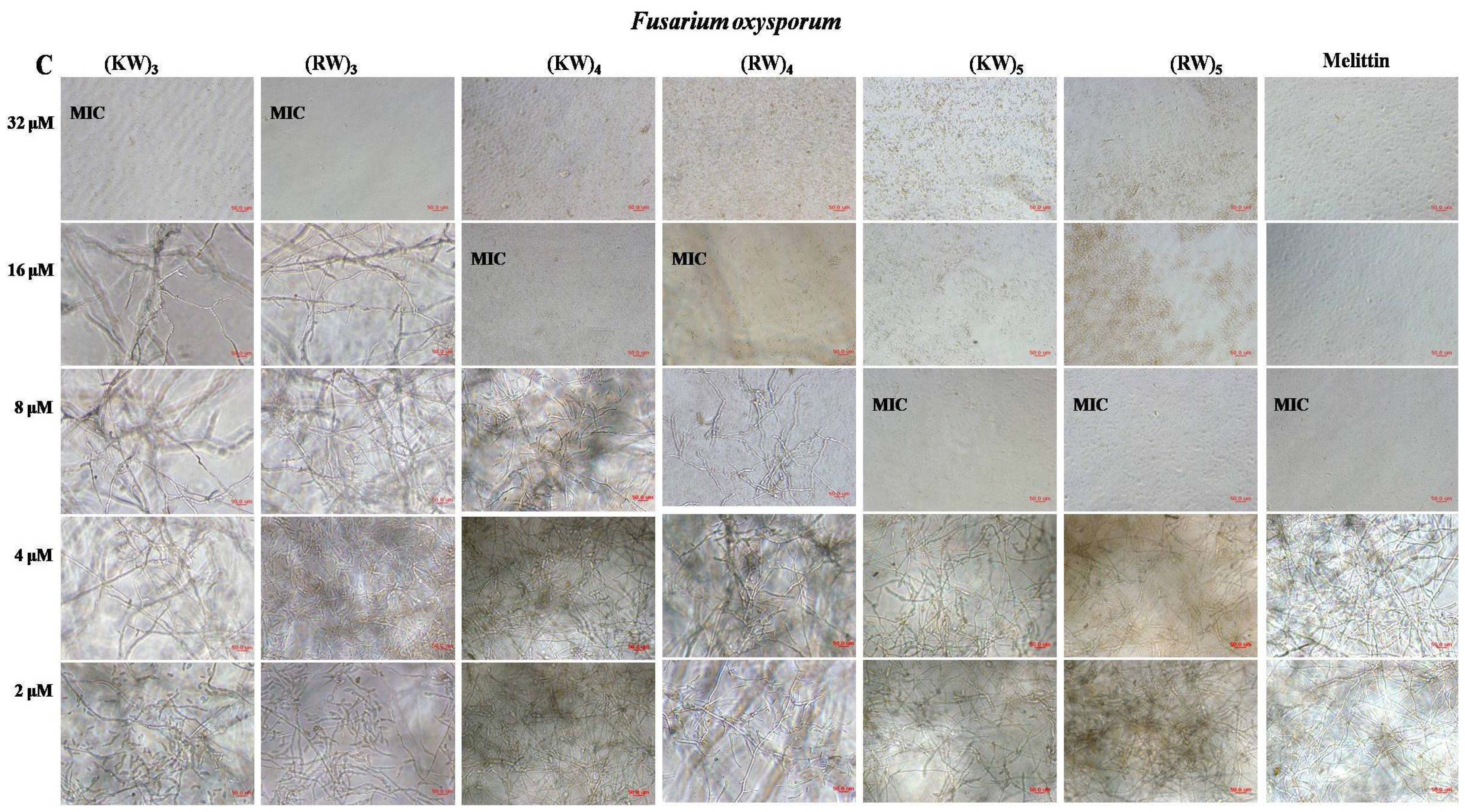
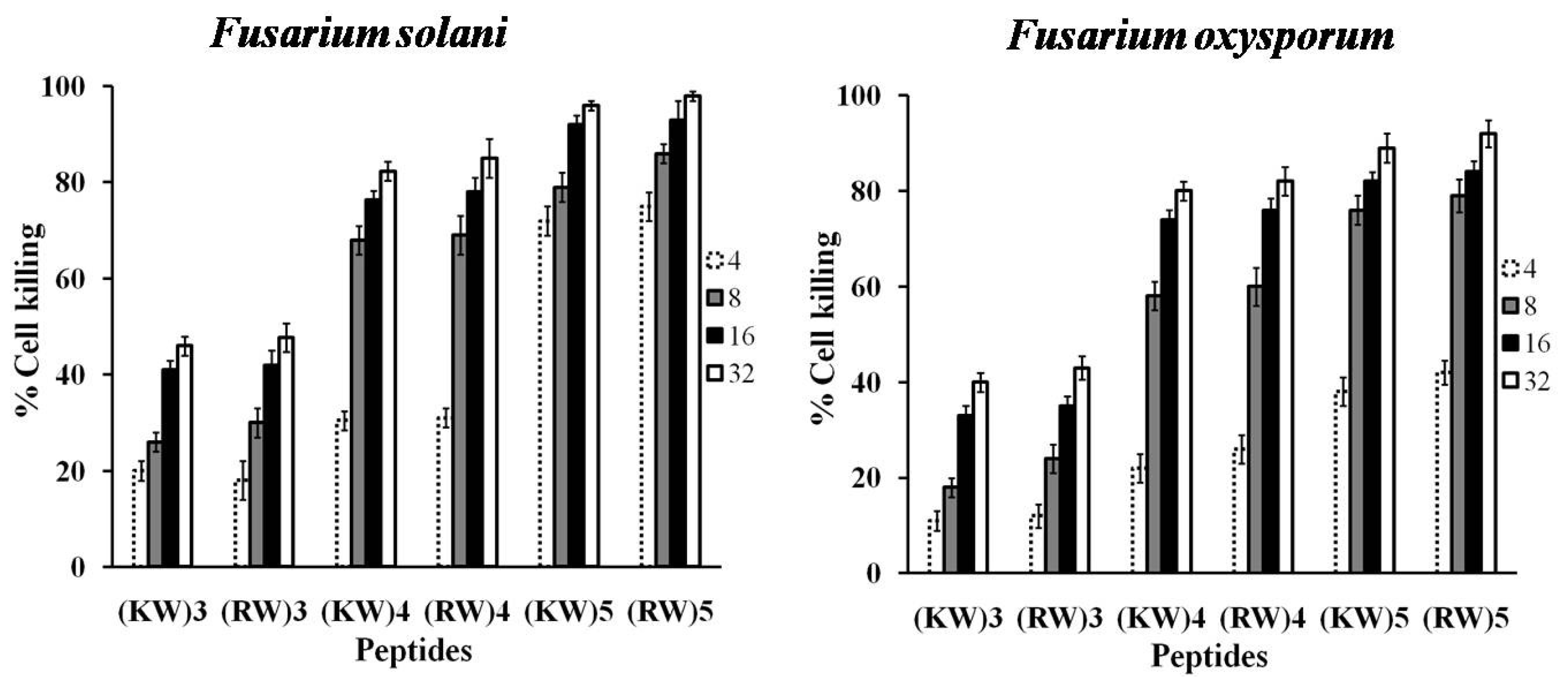
2.1. Antifungal Activities against Hyphal Growth
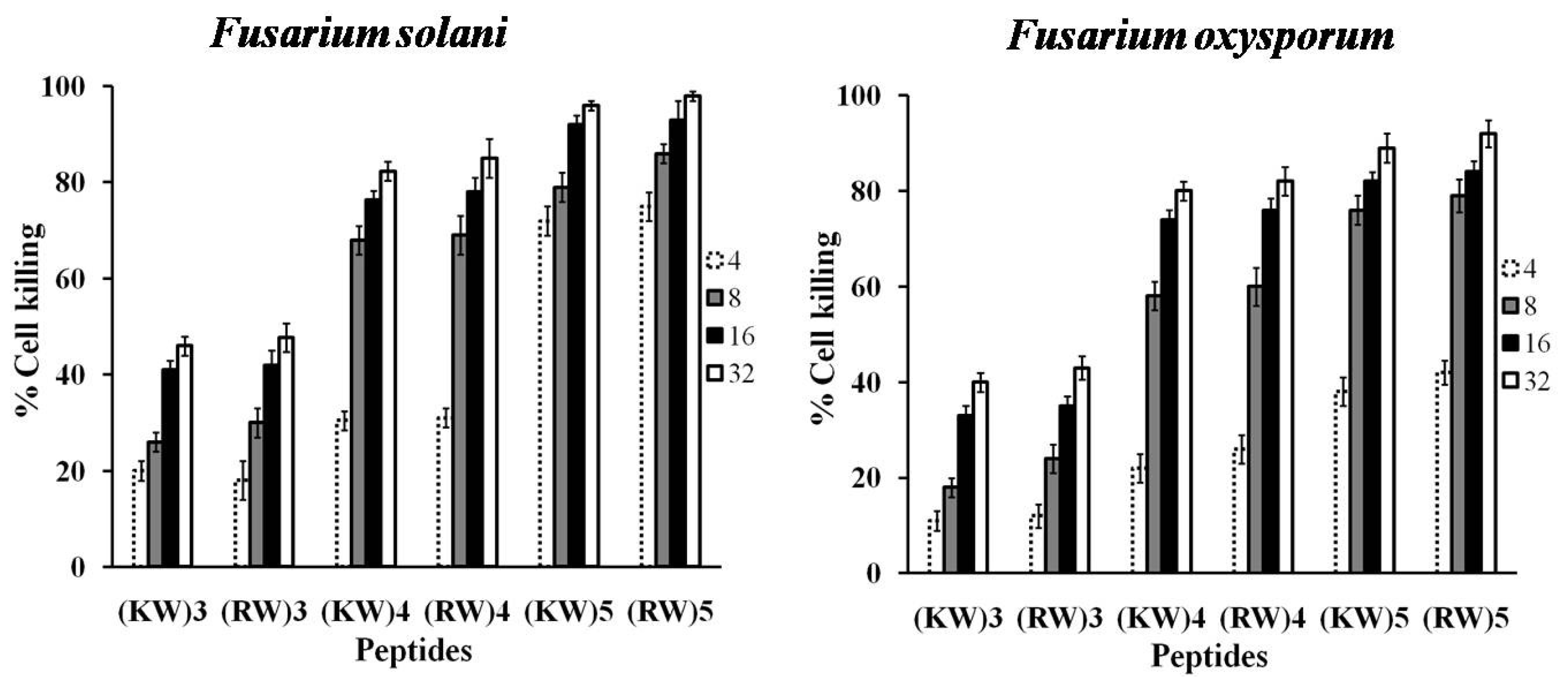
2.2. Effects of Longer Peptides on Cell Viability
3. Experimental Section
3.1. Materials
3.2. Peptide Synthesis and Purifications
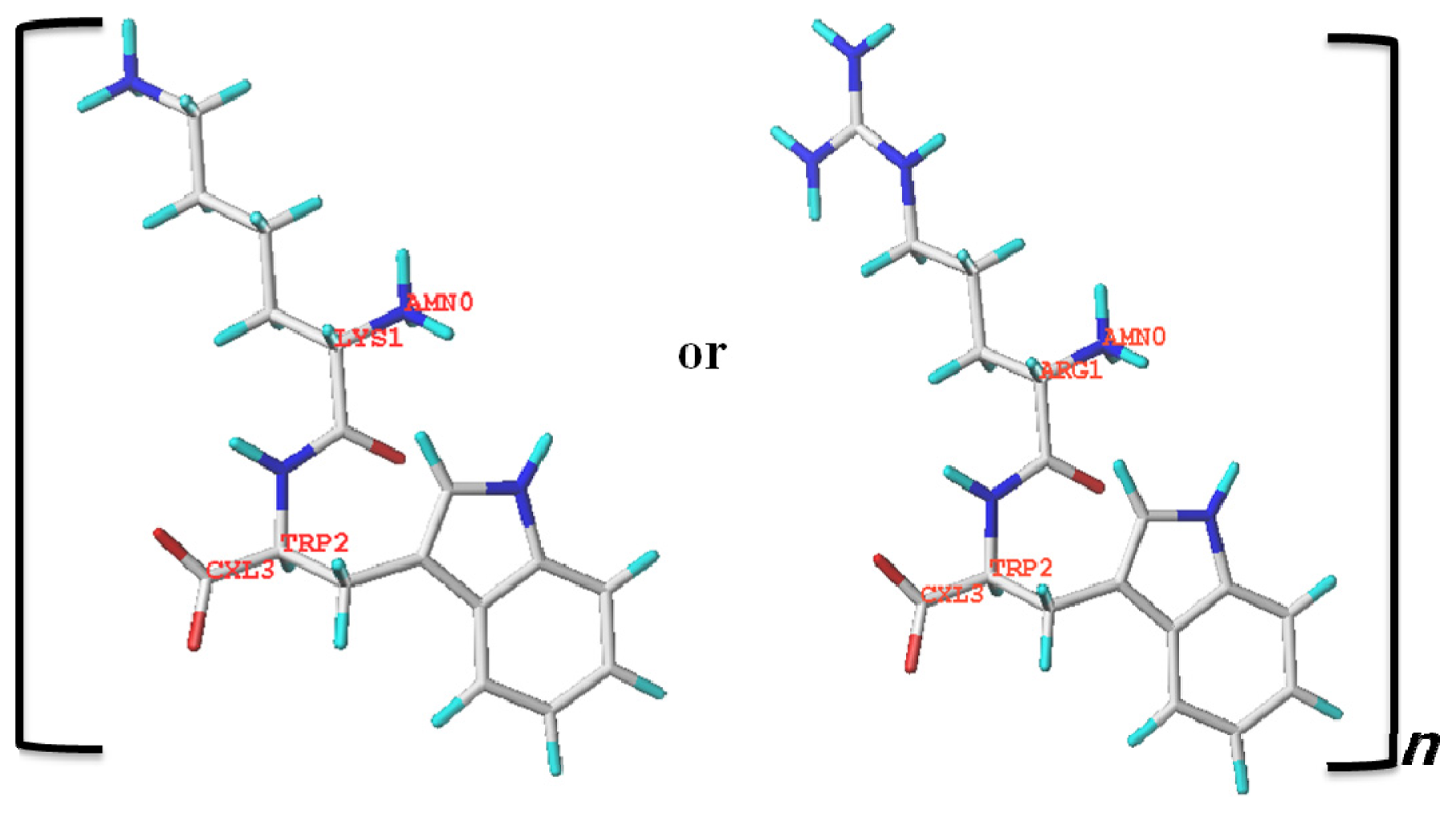
3.3. Computational Modeling
3.4. Antifungal Assay
3.5. Cell Viability Assay
4. Conclusions
Acknowledgments
References
- Laverty, G.; Gorman, S.P.; Gilmore, B.F. The potential of antimicrobial peptides as biocides. Int. J. Mol. Sci 2011, 12, 6566–6596. [Google Scholar]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol 2005, 3, 238–250. [Google Scholar]
- Hancock, R.E.W.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol 2006, 24, 1551–1557. [Google Scholar]
- Strom, M.B.; Rekdal, O.; Svendsen, J.S. Antimicrobial activity of short arginine- and tryptophan-rich peptides. J. Pept. Sci 2002, 8, 431–437. [Google Scholar]
- Storm, M.B.; Haug, B.E.; Skar, M.L.; Stensen, W.; Stiberg, T.; Svendsen, J.S. The pharmacophore of short cationic antibacterial peptides. J. Med. Chem 2003, 46, 1567–1570. [Google Scholar]
- Liu, Z.; Brady, A.; Young, A.; Rasimick, B.; Chen, K.; Zhou, C.; Kallenbach, N.R. Length effects in antimicrobial peptides of the (RW)n series. Antimicrob. Agents Chemother 2007, 51, 597–603. [Google Scholar]
- Jing, W.; Hunter, H.N.; Hagel, J.; Vogel, H.J. The structure of the antimicrobial peptide Ac-RRWWRF-NH2 bound to micelles and its interactions with phospholipids bilayers. J. Pept. Res 2003, 61, 219–229. [Google Scholar]
- Dathe, M.; Nikolenko, H.; Klose, J.; Bienert, M. Cyclization increases the antimicrobial activity and selectivity of arginine- and tryptophan-containing hexapeptides. Biochemistry 2004, 43, 9140–9150. [Google Scholar]
- Sagaram, U.S.; Pandurangi, R.; Kaur, J.; Smith, T.J.; Shah, D.M. Structure-activity determinants in antifungal plant defensins MsDef1 and MtDef4 with different modes of action against fusarium graminearum. PLoS One 2011, 6. [Google Scholar] [CrossRef]
- Powell, W.A.; Catranis, C.M.; Maynard, C.A. Synthetic antimicrobial peptide design. Mol. Plant Microb. Interact 1995, 8, 792–794. [Google Scholar]
- Muñoz, A.; López-Garcia, B.; Marcos, J.F. Comparative study of antimicrobial peptides to control citrus postharvest decay caused by penicillium digitatum. J. Agric. Food Chem 2007, 55, 8170–8176. [Google Scholar]
- Muñoz, A.; López-Garcia, B.; Pérez-Payá, E.; Marcos, J.F. Antimicrobial properties of derivatives of the cationic tryptophan-rich hexapeptide PAF26. Biochem. Biophys. Res. Commun 2007, 354, 172–177. [Google Scholar]
- Chan, D.I.; Prenner, E.J.; Vogel, H.J. Tryptophan and arginine rich antimicrobial peptides: Structures and mechanism of action. Biochim. Biophys. Acta 2006, 1758, 1184–1202. [Google Scholar]
- Falla, T.J.; Karunaratne, D.N.; Hancock, R.E.W. Mode of action of the antimicrobial peptide indolicidin. J. Biol. Chem 1996, 271, 19298–19303. [Google Scholar]
- Lay, F.T.; Anderson, M.A. Defensins–components of the innate immune system in plants. Curr. Protein Pept. Sci 2005, 6, 85–101. [Google Scholar]
- Thomma, B.P.H.J.; Cammue, B.P.A.; Thevissen, K. Plant defensins. Planta 2002, 216, 193–202. [Google Scholar]
- Carvalho, A.O.; Gomes, V.M. Plant defensins–prospects for the biological functions and biotechnological properties. Peptides 2009, 30, 1007–1020. [Google Scholar]
- Murad, A.M.; Pelegrini, P.B.; Neto, S.M.; Franco, O.L. Novel findings of defensins and their utilization in construction of transgenic plants. Transgenic Plant J 2007, 1, 39–48. [Google Scholar]
- Stotz, H.U.; Thomson, J.G.; Wang, Y. Plant defensins: Defense, development and application. Plant Signal. Behav 2009, 4, 1010–1012. [Google Scholar]
- Tavares, L.S.; Santos Mde, O.; Viccini, L.F.; Moreira, J.S.; Miller, R.N.; Franco, O.L. Biotechnological potential of antimicrobial peptides from flowers. Peptides 2008, 29, 1842–1851. [Google Scholar]
- López-García, B.; Pérez-Payá, E.; Marcos, J.F. Identification of novel hexapeptide bioactive against phytopathogenic fungi through screening of a synthetic peptide combinatorial library. Appl. Environ. Microbiol 2002, 68, 2453–2460. [Google Scholar]
- Choi, J.; Moon, E. Identification of novel bioactive hexapeptides against phytopathogenic bacteria through rapid screening of a synthetic combinatorial library. J. Microbiol. Biotechnol 2009, 19, 792–802. [Google Scholar]
- Lóopez-Garcia, B.; González-Candelas, L.; Pérez-payá, E.; Marcos, J.F. Identification and characterization of a hexapeptide with activity against phytopathogenic fungi that cause postharvest decay in fruits. Mol. Plant Microbe Interact 2000, 13, 837–846. [Google Scholar]
- Chen, P.W.; Shyu, C.L.; Mao, F.C. Antimicrobial activity of short hydrophobic and basic-rich peptides. Am. J. Vet. Res 2003, 64, 1088–1092. [Google Scholar]
- Wessolowski, A.; Bienert, M.; Dathe, M. Antimicrobial activity of arginine and tryptophan rich hexapeptides: The effects of aromatic clusters, d-amino acid substitution and cyclization. J. Pept. Res 2004, 64, 159–169. [Google Scholar]
- Gopal, R.; Kim, Y.J.; Seo, C.H.; Hahm, K.S.; Park, Y. Reversed sequence enhances antimicrobial activity of a synthetic peptide. J. Pept. Sci 2011, 17, 329–334. [Google Scholar]
- Cochrane, V.W.; Cochrane, J.C. Chlamydospore induction in pure culture in Fusarium solani. Mycologia 1971, 63, 462–477. [Google Scholar]
- Cho, J.H.; Rupe, J.C.; Cummings, M.S.; Gbur, E.E., Jr. Isolation and identification of Fusarium solani f. sp. glycines from soil on modified Nash and Snyder’s medium. Plant Dis. 2001, 85, 256–260. [Google Scholar]
- Aoki, T.; O’Donnell, K.; Homma, Y.; Lattanzi, A. Sudden-death syndrome of soybean is caused by two morphologically and phylogenetically distinct species within the Fusarium solani species complex—F. virguliforme in North America and F. tucumaniae in South America. Mycologia 2003, 95, 660–684. [Google Scholar]
- Zaccardelli, M.; Vitale, S.; Luongo, L.; Merighi, M.; Corazza, L. Morphological and Molecular Characterization of Fusarium solani Isolates. J. Phytopathol 2008, 156, 534–541. [Google Scholar]
- Gómez, Y.; Gil, K.; González, E.; Farías, L.M. Anti-fungi activity of organic extracts from the tree Fagara monophylla (Rutaceae) in Venezuela. Rev. Biol. Trop 2007, 55, 767–775. [Google Scholar]
- Blondelle, S.E.; Takahashi, E.; Dinh, K.T.; Houghten, R.A. The antimicrobial activity of hexapeptides derived from synthetic combinatorial libraries. J. Appl. Bacteriol 1995, 78, 39–46. [Google Scholar]
- Knight, S.C.; Anthony, V.M.; Brady, A.M.; Greenland, A.J.; Heaney, S.P.; Murray, D.C.; Powell, K.A.; Schulz, M.A.; Spinks, C.A.; Worthington, P.A.; et al. Rationale and perspectives in the development of fungicides. Annu. Rev. Phytopathol 1997, 35, 349–372. [Google Scholar]
- Terras, F.R.G.; Eggermont, K.; Kovaleva, V.; Raikhel, N.V.; Osborn, R.W.; Kester, A.; Rees, S.B.; Torrekens, S.; van Leuven, F.; Vanderleyden, J.; et al. Small cysteine rich antifungal proteins from radish: Their role in host defence. Plant Cell 1995, 7, 573–588. [Google Scholar]
- Ponti, D.; Mangoni, M.L.; Mignogna, G.; Simmaco, M.; Barra, D. An amphibian antimicrobial peptide variant expressed in Nicotiana tabacum confers resistance to phytopathogens. Biochem. J 2003, 370, 121–127. [Google Scholar]
- Huang, Y.; Nordeen, R.O.; Di, M.; Owens, L.D.; Mcbeath, J.H. Expression of an engineered cecropin gene cassette in transgenic tobacco plants confers resistance to Pseudomonas syringae pv. tabaci. Phytopathology 1997, 87, 494–499. [Google Scholar]
- Cary, J.W.; Rajasekaran, K.; Jaynes, J.M.; Cleveland, T.E. Transgenic expression of a gene encoding a synthetic antimicrobial peptide result in inhibition of fungal growth in vitro and in planta. Plant Sci 2000, 154, 171–181. [Google Scholar]
- Osusky, M.; Zhou, G.; Osuska, L.; Hancock, R.E.; Kay, W.W.; Misra, S. Transgenic plants expressing cationic peptide chimeras exhibit broad spectrum resistance to phytopathogens. Nat. Biotechnol 2000, 18, 1162–1166. [Google Scholar]
- Montesinos, E. Antimicrobial peptides and plant disease control. FEMS Microbiol. Lett 2007, 270, 1–11. [Google Scholar]
- Xing, H.Y.; Lawrence, C.B.; Chambers, O.; Davies, H.M.; Everett, N.P.; Li, Q.Q. Increased pathogen resistance and yield in transgenic plants expressing combinations of the modified antimicrobial peptides based on indolicidin and magainin. Planta 2006, 223, 1024–1032. [Google Scholar]
- Bhargava, A.; Osusky, M.; Hancock, R.E.; Forward, B.S.; Kay, W.W.; Misra, S. Antiviral indolicidin variant peptides: Evaluation for broad-spectrum disease resistance in transgenic Nicotiana tabacum. Plant Sci 2007, 172, 515–523. [Google Scholar]
- López-Garcia, B.; Veyrat, A.; Pérez-Payá, E.; González-candelas, L.; Marcos, J.F. Comparison of the activity of antifungal hexapeptides and the fungicides thiabendazole and imazalil against postharvest fungal pathogens. Int. J. Food Microbiol 2003, 89, 163–170. [Google Scholar]
- Gopal, R.; Park, J.S.; Seo, C.H.; Park, Y. Applications of circular dichroism for structural analysis of gelatin and antimicrobial peptides. Int. J. Mol. Sci 2012, 13, 3229–3244. [Google Scholar]
- Nan, Y.H.; Lee, S.H.; Kim, H.J.; Shin, S.Y. Mammalian cell toxicity and candidacidal mechanism of Arg- or Lys-containing Trp-rich model antimicrobial peptides and their d-enantiomeric peptides. Peptides 2010, 31, 1826–1831. [Google Scholar]
- Muñoz, A.; López-Garcia, B.; Marcos, J.F. Studies on the mode of action of the antifungal hexapeptide PAF26. Antimicrob. Agents Chemother 2006, 50, 3847–3855. [Google Scholar]
- Munoz, A.; Marcos, J.F.; Read, N.D. Concentration-dependent mechanisms of cell penetration and killing by the de novo designed antifungal hexapeptide PAF26. Mol. Microbiol 2012. [Google Scholar] [CrossRef]
- Atherton, E.; Sheppard, R.C. Solid Phase Peptide Synthesis: A Practical Approach; IRL Press: Oxford, UK; p. 1989.
- Van der Weerden, N.L.; Hancock, R.E.; Anderson, M.A. Permeabilization of fungal hyphae by the plant defensin NAD1 occurs through a cell wall dependent process. J. Biol. Chem 2010, 285, 37513–37520. [Google Scholar]
- Park, S.C.; Lee, J.R.; Kim, J.Y.; Hwang, I.; Nah, J.W.; Cheong, H.; Park, Y.; Hahm, K.S. Pr-1, a novel antifungal protein from pumpkin rinds. Biotechnol. Lett 2010, 32, 125–130. [Google Scholar]

© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gopal, R.; Na, H.; Seo, C.H.; Park, Y. Antifungal Activity of (KW)n or (RW)n Peptide against Fusarium solani and Fusarium oxysporum. Int. J. Mol. Sci. 2012, 13, 15042-15053. https://doi.org/10.3390/ijms131115042
Gopal R, Na H, Seo CH, Park Y. Antifungal Activity of (KW)n or (RW)n Peptide against Fusarium solani and Fusarium oxysporum. International Journal of Molecular Sciences. 2012; 13(11):15042-15053. https://doi.org/10.3390/ijms131115042
Chicago/Turabian StyleGopal, Ramamourthy, Hyungjong Na, Chang Ho Seo, and Yoonkyung Park. 2012. "Antifungal Activity of (KW)n or (RW)n Peptide against Fusarium solani and Fusarium oxysporum" International Journal of Molecular Sciences 13, no. 11: 15042-15053. https://doi.org/10.3390/ijms131115042
APA StyleGopal, R., Na, H., Seo, C. H., & Park, Y. (2012). Antifungal Activity of (KW)n or (RW)n Peptide against Fusarium solani and Fusarium oxysporum. International Journal of Molecular Sciences, 13(11), 15042-15053. https://doi.org/10.3390/ijms131115042