Assessment of Genetic Diversity of Bermudagrass (Cynodon dactylon) Using ISSR Markers

Abstract
:1. Introduction
2. Results
2.1. ISSR Amplification
2.2. Genetic Variation and Similarity among Genotypes
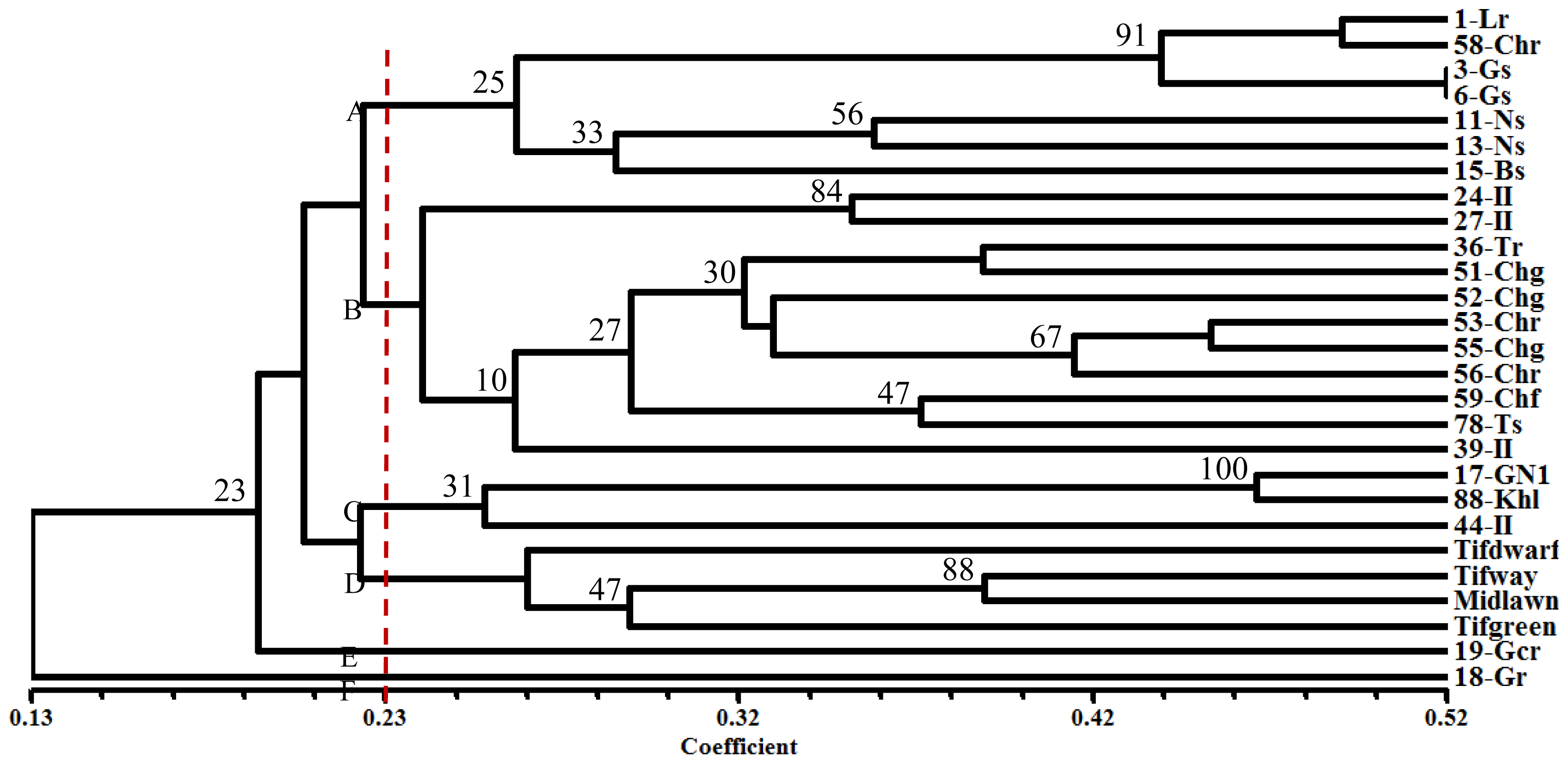
2.3. Cluster Analysis
3. Discussion
4. Experimental Section
4.1. Plant Materials and DNA Extraction
4.2. ISSR Assays
4.3. Data Analysis
5. Conclusion
References
- Harlan, J.R. Cynodon species and their value for grazing or hay. Herb. Abstr 1970, 40, 233–238. [Google Scholar]
- Harlan, J.R.; de Wet, J.M.J. Sources of variation in Cynodon dactylon (L.) Pers. Crop Sci 1969, 9, 774–778. [Google Scholar]
- Gupta, P.K.; Varshney, R.K. The development and use of microsatellite markers for genetic analysis and plant breeding with emphasis on bread wheat. Euphytica 2000, 113, 163–185. [Google Scholar]
- Reddy, M.P.; Sarla, N.; Siddiq, E.A. Inter simple sequence repeat (ISSR) polymorphism and its application in plant breeding. Euphytica 2002, 128, 9–17. [Google Scholar]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar]
- Barth, S.; Melchinger, A.E.; Lubberstedt, T.H. Genetic diversity in Arabidopsis thaliana L. Heynh. Investigated by cleaved amplified polymorphic sequence (CAPS) and intersimple sequence repeat (ISSR) markers. Mol. Ecol 2002, 11, 495–505. [Google Scholar]
- Huang, C.Q.; Liu, G.D.; Bai, C.J.; Wang, W.Q.; Zhou, S.Y.; Yu, D.Q. Estimation of genetic variation in Cynodon dactylon accessions using the ISSR technique. Biochem. Syst. Ecol 2010, 38, 463–470. [Google Scholar]
- Reunova, G.D.; Kats, I.L.; Muzarok, T.I.; Zhuravleu, I. Polymerase of RAPD, ISSR and AFLP markers of the Panax ginseng C.A. Meyer (Araliaceae) genome. Genetika 2010, 46, 1057–1066. [Google Scholar]
- Smolik, M.; Ochmian, I.; Grajkowski, J. Genetic variability of Polish and Russian accessions of cultivated blue honeysuckle (Lonicera caerulea). Genetika 2010, 46, 1079–1085. [Google Scholar]
- Zehdi, S.; Trifi, M.; Salem, A.; Rhouma, A.; Marrakchi, M. Survey of inter simple sequence repeat polymorphisms in Tunisian date palms (Phoenix dactylifera L.). J. Genet. Breed 2002, 56, 77–83. [Google Scholar]
- Gulsen, O.; Sever, S.; Mutlu, N.; Tuna, M.; Karaquzel, O.; Shearman, R.C. Ployploidy creates higher diversity among Cynodon accessions as assessed by molecular markers. Theor. Appl. Genet 2009, 118, 1309–1319. [Google Scholar]
- Arcade, A.; Anselin, F.; Faivye, P.; Lesage, M.C.; Paques, L.E.; Prat, D. Application of AFLP, RAPD and ISSR markers to genetic mapping of European and Japanese larch. Theor. Appl. Genet 2004, 100, 627–632. [Google Scholar]
- Budak, H.; Shearman, R.C.; Gulsen, O.; Dweikat, I. Understanding ploidy complex and geographic origin of the Buchloe dactyloides genome using cytoplasmic and nuclear marker systems. Theor. Appl. Genet 2005, 111, 1545–1552. [Google Scholar]
- Ge, X.J.; Yu, Y.; Yuan, Y.; Huang, H.W.; Yan, C. Genetic diversity and geographic differentiation in endangered Ammopiptanthus (Leguminosae) populations in desert regions of northwest China as revealed by ISSR analysis. Ann. Bot 2005, 95, 843–851. [Google Scholar]
- Raina, S.N.; Rani, V.; Kojima, T.; Ogihara, Y.; Singh, K.P.; Devarumath, R.M. RAPD and ISSR fingerprints as useful genetic markers for analysis of genetic diversity, varietal identification, and phylogenetic relationships in peanut (Arachis hypogaea) cultivars and wild species. Genome 2001, 44, 763–772. [Google Scholar]
- Zhang, L.; Li, Q.J.; Li, H.T.; Chen, J.; Li, D.Z. Genetic diversity and geographic differentiation in Tacca chantrieri (Taccaceae) san autonomous selfing plant with showy floral display. Ann. Bot 2006, 98, 449–457. [Google Scholar]
- Anderson, M.P.; Taliaferro, C.M.; Martin, D.L.; Anderson, C.S. Comparative DNA profiling of U-3 turf bermudagrass strains. Crop Sci 2001, 41, 1184–1189. [Google Scholar]
- Etemadi, N.; Sayed-tabatabaei, B.E.; Zamani, Z.; Razmjoo, K.H.; Khalighi, A.; Lessani, H. Evaluation of diversity among Cynodon dactylon (L.) Pers. using RAPD markers. Agric. Biol 2006, 8, 198–202. [Google Scholar]
- Roodt, R.; Spies, J.J.; Burger, T.H. Preliminary DNA fingerprinting of the turf grass Cynodon dactylon (Poaceae: Chloridoideae). Bothalia 2002, 32, 117–122. [Google Scholar]
- Karaca, M.; Saha, S.; Zipf, A.; Jenkins, J.N.; Lang, D.J. Genetic diversity among bermudagrass (Cynodon spp.): Evidence from chloroplast and nuclear DNA fingerprinting. Crop Sci 2002, 42, 2118–2127. [Google Scholar]
- Wu, Y.; Taliaferro, C.M.; Bai, G.H.; Anderson, M.P. AFLP analysis of Cynodon dactylon (L.) Pers. Var. dactylon genetic variation. Genome 2004, 47, 689–696. [Google Scholar]
- Li, H.; Liu, L.; Lou, Y.; Hu, T.; Fu, J. Genetic diversity of Chinese natural bermudagrass (Cynodon dactylon) germplasm using ISSR markers. Sci. Hortic 2011, 127, 555–561. [Google Scholar]
- Kamps, T.L.; Williams, N.R.; Ortega, V.M.; Chamusco, K.C.; Harris-Shultz, K.; Scully, B.T.; Chase, C.D. DNA polymorphisms at bermudagrass microsatellite loci and their use in genotype fingerprinting. Crop Sci 2011, 51, 1122–1131. [Google Scholar]
- Vaiman, D.; Mercier, D.; Moazami-Goudarzi, K.; Eggen, A.; Ciampolini, R.; Lépingle, A.; Velmala, R.; Kaukinen, J.; Varvio, S.L.; Martin, P.; Levéziel, H.; Guérin, G. A set of 99 cattle microsatellite, characterization, synteny mapping and polymorphism. Mamm. Genome 1994, 5, 288–297. [Google Scholar]
- Xie, W.; Zhang, X.; Cai, H.; Liu, W.; Peng, Y. Genetic diversity analysis and transferability of cereal EST-SSR markers to orchardgrass (Dactylis glomerata L.). Biochem. Syst. Ecol 2010, 38, 740–749. [Google Scholar]
- Tajima, F.; Tokunaga, T.; Miyashita, N.T. Statistical methods for estimating the effective number of alleles, expected heterozygosity and genetic distance in self-incompatibility locus. Jpn. J. Genet 1994, 69, 287–295. [Google Scholar]
- Mohammadi, S.A.; Prasanna, B.M. Analysis of genetic diversity in crop plants—Salient statistical tools and considerations. Crop Sci 2003, 43, 1235–1248. [Google Scholar]
- Burton, G.W. African Grasses; Janick, J., Simon, J.E., Eds.; Wiley: New York, NY, USA, 1993; pp. 294–298. [Google Scholar]
- Zhang, L.H.; Ozias-Akins, P.; Kochert, G.; Kresovich, S.; Dean, R.; Hanna, W. Differentiation of bermudagrass (Cynodon spp.) genotypes by AFLP analyses. Theor. Appl. Genet 1999, 98, 895–902. [Google Scholar]
- Caetano-Anolles, G. DNA analysis of turfgrass genetic diversity. Crop Sci 1998, 38, 1415–1424. [Google Scholar]
- Yi, Y.; Zhang, X.; Huang, L.; Ling, Y.; Ma, X.; Liu, W. Genetic diversity of wild Cynodon dactylon germplasm detected by SRAP markers. Yi Chuan 2008, 30, 94–100. [Google Scholar]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep 1983, 4, 19–21. [Google Scholar]
- Smith, J.S.C.; Chin, E.C.L.; Shu, H.; Smith, O.S.; Wall, S.J.; Senior, M.L.; Mitchell, S.E.; Kresovich, S.; Zeigle, J. An evaluation of the utility of SSR loci as molecular markers in maize (Zea mays L.): Comparison with data from RFLPs and pedigree. Theor. Appl. Genet 1997, 95, 163–173. [Google Scholar]
- Michener, C.D.; Sokal, R.R. A quantitative approach to a problem of classification. Evolution 1957, 11, 490–499. [Google Scholar]
- Rohlf, F.J. NTSYSpc. Numerical Taxonomy and Multivariate Analysis System, version 2.02; Exeter software; State University of New York: New York, NY, USA, 1998. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. The comparison of dendrograms by objective methods. Taxon 1962, 11, 30–40. [Google Scholar]
- Yap, I.V.; Nelson, R.J. Winboot: A Program for Performing Bootstrap Analysis of Binary Data to Determine the Confidence Limits of UPGMA-based Dendrograms, International Rice Research Institute: Metro Manila, Philippines, 1996.
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Yeh, F.C.; Yang, R.C.; Boyle, T.; Ye, Z.H.; Mao, J.X. POPGENE, the User Friendly Shareware for Population Genetic Analysis; University of Alberta: Edmonton, AB, Canada, 1999. [Google Scholar]
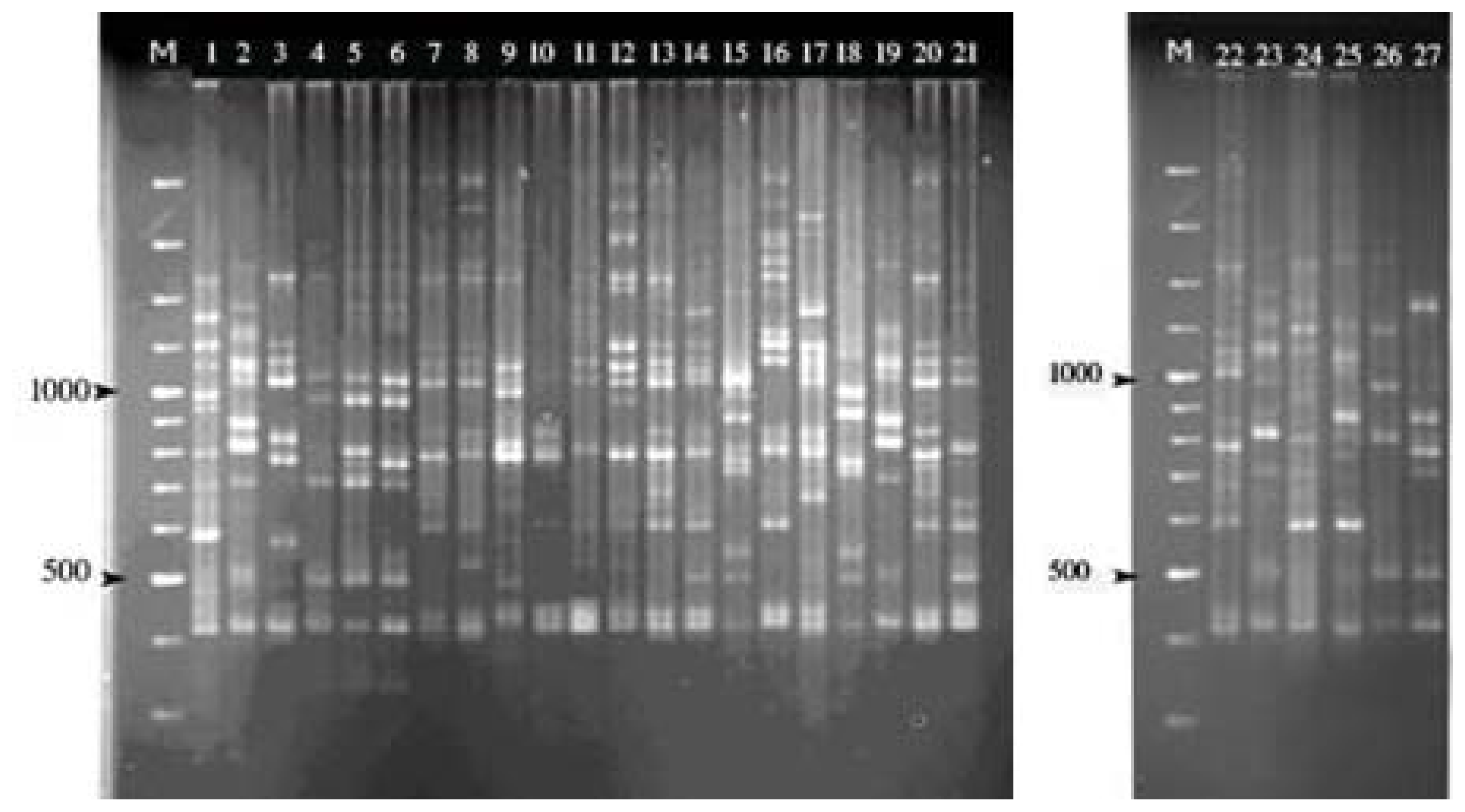

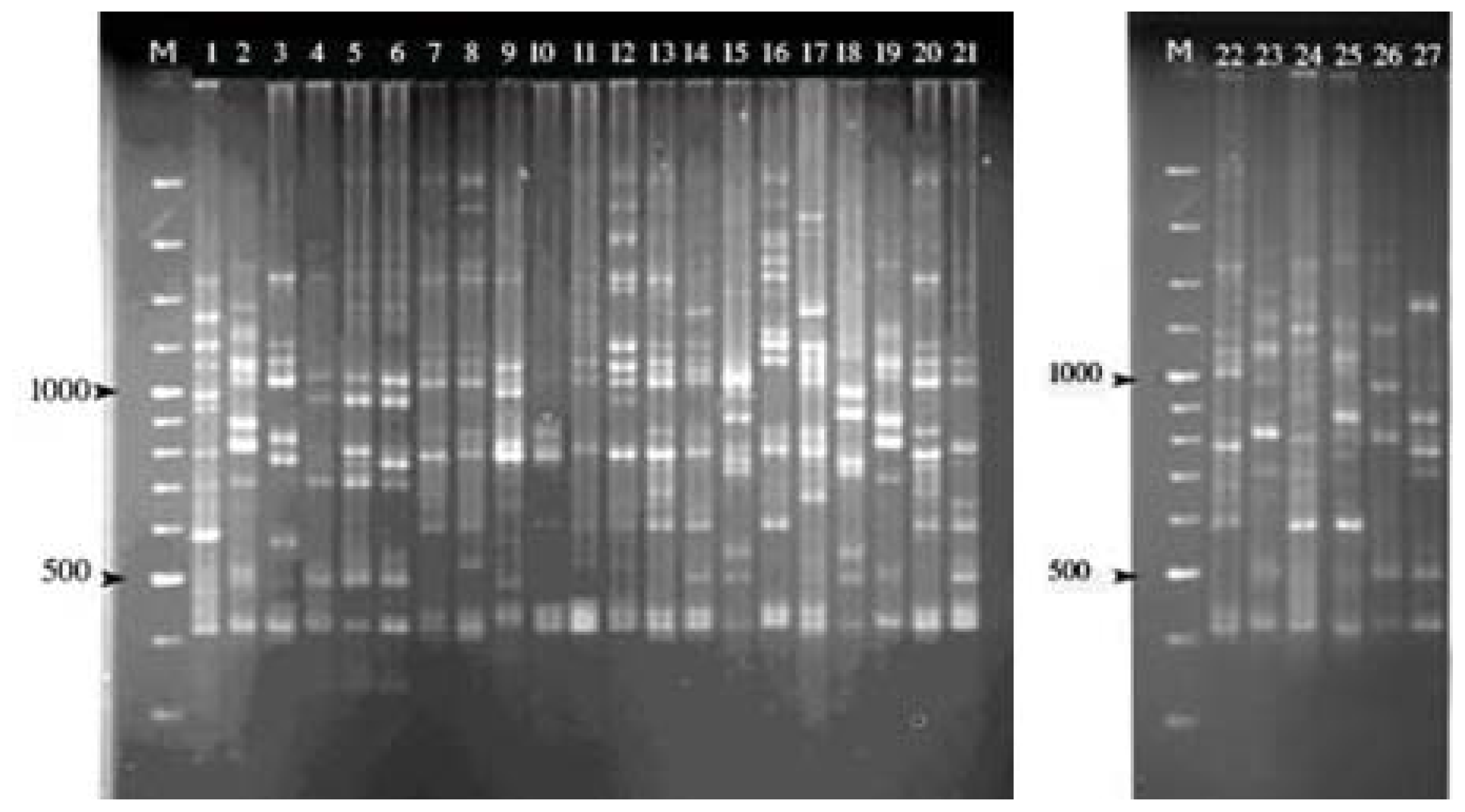{kind=link}
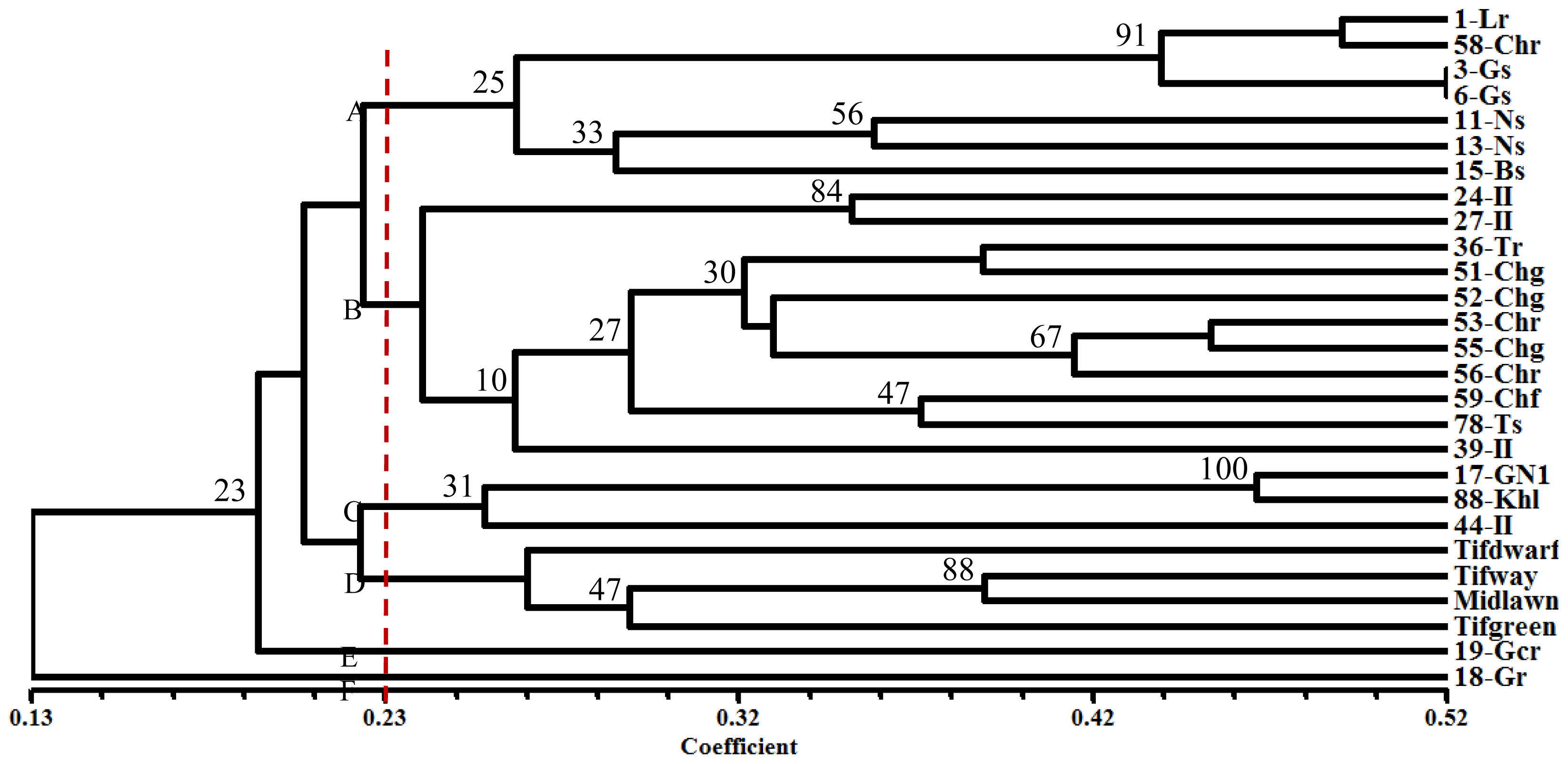{kind=link}
| Primer name | Nucleotide sequence | Tm (°C) | No. of fragments amplified | No. of polymorphic fragments | Percentage of poymorphism | PIC value |
|---|---|---|---|---|---|---|
| ISSR-1 | 5′-(AC)8 G-3′ | 54/9 | 23 | 19 | 82.6 | 0.313 |
| ISSR-2 | 5′-(AC)8T-3′ | 49/2 | 29 | 25 | 86.2 | 0.266 |
| ISSR-3 | 5′-(AC)8C-3′ | 53/3 | 25 | 20 | 80 | 0.291 |
| ISSR-4 | 5′-(AG)8C-3′ | 46/8 | 24 | 20 | 83.3 | 0.280 |
| ISSR-5 | 5′-(AG)8T-3′ | 42/5 | 28 | 26 | 92.8 | 0.205 |
| ISSR-6 | 5′-(GA)8C-3′ | 43/3 | 25 | 22 | 88 | 0.283 |
| ISSR-7 | 5′-(GA)8T-3′ | 42/9 | 36 | 26 | 72.6 | 0.333 |
| ISSR-8 | 5′-(CT)8G-3′ | 44/9 | 30 | 20 | 66.7 | 0.346 |
| ISSR-11 | 5′-(AG)8GT-3′ | 47/1 | 40 | 27 | 67.5 | 0.333 |
| ISSR-14 | 5′-(AC)8GG-3′ | 57 | 20 | 16 | 80 | 0.375 |
| ISSR-15 | 5′-(AC)8GA-3′ | 53/7 | 25 | 20 | 80 | 0.433 |
| RAMP-TAG | 5′-T(AG)9-3′ | 56/1 | 29 | 24 | 82.7 | 0.428 |
| RAMP-GAC | 5′-G(AC)9-3′ | 53/9 | 25 | 22 | 88 | 0.391 |
| LK7 | 5′-CCA(CT)8-3′ | 56/1 | 30 | 26 | 86.7 | 0.321 |
| Total means | - | - | 389 | 313 | 80.5 | 0.328 |
| Accession name | Collected site (province-city) | Altitude (m) | Annual Rainfall (mm) | Main daily temperature (°C) | Choromosome No. |
|---|---|---|---|---|---|
| 1-Lr | Charmahal Bakhtiari-Lordegan | 1900 | 551 | 17/5 | 36 |
| 3-Gs | Charmahal Bakhtiari-Borujen | 2200 | 245 | 12/8 | 36 |
| 6-Gs | Charmahal Bakhtiari-Borujen | 1930 | 245 | 12/8 | 36 |
| 11-Ns | Isfahan-Natanz | 1800 | 198 | 16/4 | 36 |
| 13-Ns | Isfahan-Natanz | 1800 | 198 | 16/4 | 36 |
| 15-Bs | Isfahan-Natanz | 1300 | 198 | 16/4 | 36 |
| 17-GN1 | Mazandaran-Neka | 15 | 895 | 16/8 | 18 |
| 18-Gr | Gilan-Chaboksar | 15- | 1220 | 15/9 | 36 |
| 19-Gcr | Gilan-Chaboksar | 15- | 1220 | 15/9 | 36 |
| 24-II | Isfahan-Isfahan | 2060 | 122 | 16/2 | 36 |
| 27-II | Isfahan-Isfahan | 1577 | 122 | 16/2 | 36 |
| 36-Tr | Isfahan-Isfahan | 1580 | 122 | 16/2 | 36 |
| 39-II | Isfahan-Isfahan | 1573 | 122 | 16/2 | 36 |
| 44-II | Isfahan-Isfahan | 1570 | 122 | 16/2 | 27 |
| 51-Chg | Isfahan-Chadegan | 2150 | 317 | 10/2 | 36 |
| 52-Chg | Isfahan-Chadegan | 2150 | 317 | 10/2 | 36 |
| 53-Chr | Isfahan-Chadegan | 2150 | 317 | 10/2 | 36 |
| 55-Chg | Isfahan-Chadegan | 2180 | 317 | 10/2 | 36 |
| 56-Chr | Isfahan-Chadegan | 2180 | 317 | 10/2 | 36 |
| 58-Chr | Isfahan-Chadegan | 2170 | 317 | 10/2 | 36 |
| 59-Chf | Isfahan-Chadegan | 2160 | 317 | 10/2 | 36 |
| 78-Ts | Isfahan-Tiran | 1900 | 175 | 14/5 | 36 |
| 88-Khl | Isfahan-Khomeini shahr | 1560 | 130 | 15/8 | 18 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Farsani, T.M.; Etemadi, N.; Sayed-Tabatabaei, B.E.; Talebi, M. Assessment of Genetic Diversity of Bermudagrass (Cynodon dactylon) Using ISSR Markers. Int. J. Mol. Sci. 2012, 13, 383-392. https://doi.org/10.3390/ijms13010383
Farsani TM, Etemadi N, Sayed-Tabatabaei BE, Talebi M. Assessment of Genetic Diversity of Bermudagrass (Cynodon dactylon) Using ISSR Markers. International Journal of Molecular Sciences. 2012; 13(1):383-392. https://doi.org/10.3390/ijms13010383
Chicago/Turabian StyleFarsani, Tayebeh Mohammadi, Nematollah Etemadi, Badraldin Ebrahim Sayed-Tabatabaei, and Majid Talebi. 2012. "Assessment of Genetic Diversity of Bermudagrass (Cynodon dactylon) Using ISSR Markers" International Journal of Molecular Sciences 13, no. 1: 383-392. https://doi.org/10.3390/ijms13010383
APA StyleFarsani, T. M., Etemadi, N., Sayed-Tabatabaei, B. E., & Talebi, M. (2012). Assessment of Genetic Diversity of Bermudagrass (Cynodon dactylon) Using ISSR Markers. International Journal of Molecular Sciences, 13(1), 383-392. https://doi.org/10.3390/ijms13010383