Bioactivities from Marine Algae of the Genus Gracilaria

Abstract
:1. Introduction
2. Results and Discussion
2.1. Studies of Toxicity
2.2. Effects on the Nervous System
2.3. Contraception Activity
2.4. Anti-Inflammatory and Antioxidant Activities
2.5. Gastrointestinal Effects
2.6. Cardiovascular Effects
2.7. Antibiotic Activity
2.8. Antivirial Activity
3. Material and Methods
4. Conclusions
Acknowledgments
References
- Jha, RK; Zi-rong, X. Biomedical compounds from marine organisms. Mar. Drugs 2004, 2, 123–146. [Google Scholar]
- Falcão, VR. Aspectos moleculares de nitrato redutase da macroalga marinha Gracilaria tenuistipitata (Rhodophyta): Seqüenciamento do gene e estudo da expressão do RNA mensageiro. PhD Thesis, Institute of Chemical, University of São Paulo, São Paulo, Brazil, 2006; pp. 1–187. [Google Scholar]
- Babuselvam, M; Ravikumar, S. Screening of Male Anti-Fertility Compounds from Marine Seaweed Macro Algae; Division of Marine Microbiology and Medicine, Manonmaniam Sundaranar University: Rajakkamangalam, India, 1993; pp. 1–14. Available online: http://www.scisoc.or.th/stt/32/sec_h/paper/stt32_H_H0001.pdf accessed on 14 May 2011.
- Smit, AJ. Medicinal and pharmaceutical uses of seaweed natural products: A review. J. Appl. Phycol 2004, 16, 245–262. [Google Scholar]
- Ito, K; Hori, K. Seaweed: Chemical composition and potential uses. Food Rev. Int 1989, 5, 101–1144. [Google Scholar]
- Darcy-Vrillon, B. Nutritional aspects of the developing use of marine macroalgae for the human food industry. Int. J. Food Sci. Nutr 1993, 44, 523–535. [Google Scholar]
- Lahaye, M. Marine algae as a source of dietary fibers: Determination of soluble and insoluble dietary fiber contents in some ‘sea vegetable’. J. Sci. Food Agric 1993, 54, 523–535. [Google Scholar]
- Elena, M; Francisco, Y; Erickson, KL. Mailiohydrin, a cytotoxic chamigrene dibromohydrin from a Phillippine Laurencia species. J. Nat. Prod 2001, 64, 790–791. [Google Scholar]
- Kim, JB; Hudson, AM; Huang, K; Bannistes, A; Jin, TJ; Choi, GHN; Towers, YK; Wreede, RE. Biological activity of seaweed extracts from British, Colombia, Canada and Korea. I. Antiviral activity. Can. J. Bot 1997, 75, 1656–1660. [Google Scholar]
- Okai, Y; Highasi, OK; Ishizaka, S; Yamashita, U. Enhancing effect of polysaccharides from a edible brown algae, Hijikia furiform (Hijki) on release of tumour necrosis factor alpha from macrophages of endotoxin non responder C3H/HCl mice. Nutr. Cancer 1997, 27, 381–386. [Google Scholar]
- Premila, JC; Raviraja, NS; Sridhar, KR. Antimicrobial activity of some marine algae of south-west coast of India. Indian J. Mar. Sci 1996, 26, 201–205. [Google Scholar]
- Vallinayagam, K; Arumugan, R; Ragupathi Raja Kannan, RRR; Thirumaram, G; Anantharaman, P. Antibacterial activity of some selected seaweeds from Pudumadam coastal regions. Glob. J. Pharmacol 2009, 3, 50–52. [Google Scholar]
- Faulkner, DJ. Marine natural products. Nat. Prod. Rep 2002, 19, 1–48. [Google Scholar]
- Cardozo, KHM; Guaratini, T; Barros, MP; Falcão, VR; Tonon, AP; Lopes, NP; Campos, S; Torres, MA; Souza, AO; Colepicolo, P; et al. Metabolites from algae with economical impact. Comp. Biochem. Physiol. C: Comp. Pharmacol 2006, 146, 60–78. [Google Scholar]
- O’Sullivan, L; Murphy, B; McLoughlin, P; Duggan, P; Lawlor, PG; Hughes, H; Gardiner, GE. Prebiotics from marine macroalgae for human and animal health application. Mar. Drugs 2010, 8, 2038–2064. [Google Scholar]
- Paniagua-Michel, J; Capa-Robles, W; Olmos-Soto, J; Gutierrez-Millan, LE. The carotenogenesis pathway via the isoprenoid-beta-carotene interference approach in a new strain of Dunaliella salina isolated from Baja California Mexico. Mar. Drugs 2009, 7, 45–56. [Google Scholar]
- Cen-Pacheco, F; Nordstrom, L; Souto, ML; Martin, MN; Fernandez, JJ; Daranas, AH. Studies on polyethers produced by red algae. Mar. Drugs 2010, 8, 1178–1188. [Google Scholar]
- Klisch, M; Hader, DP. Mycosporine-like amino acids and marine toxins—The common and the different. Mar. Drugs 2008, 6, 147–163. [Google Scholar]
- Pallela, R; Na-Young, Y; Kim, SK. Anti-photoaging and photoprotective compounds derived from marine organisms. Mar. Drugs 2010, 8, 1189–1202. [Google Scholar]
- D’Ayala, GG; Malinconico, M; Laurienzo, P. Marine derived polysaccharides for biomedical applications: Chemical modification approaches. Molecules 2008, 13, 2069–2106. [Google Scholar]
- Kellmann, R; Stuken, A; Orr, RJS; Svendsen, HM; Jakobsen, KS. Biosynthesis and molecular genetics of polyketides in marine Dinoflagellates. Mar. Drugs 2010, 8, 1011–1048. [Google Scholar]
- Souza, ET; Lira, DP; Queiroz, AC; Silva, DJC; Aquino, AB; Mella, EAC; Lorenzo, VP; Miranda, GEC; Araújo-Júnior, JX; Chaves, MCO; et al. The antinociceptive and anti-inflammatory activities of caulerpin, a bisindole alkaloid isolated from seaweeds of the genus Caulerpa. Mar. Drugs 2009, 7, 689–704. [Google Scholar]
- Guven, KC; Percot, A; Sezik, E. Alkaloids in marine algae. Mar. Drugs 2010, 8, 269–284. [Google Scholar]
- Cabrita, MT; Vale, C; Rauter, AP. Halogenated compounds from marine algae. Mar. Drugs 2010, 8, 2301–2317. [Google Scholar]
- La Barre, S; Potin, P; Leblanc, C; Delage, L. The halogenated metabolism of brown algae (Phaeophyta), its biological importance and its environmental significance. Mar. Drugs 2010, 8, 988–1010. [Google Scholar]
- Ianora, A; Boersma, M; Casotti, R; Fontana, A; Harder, J; Hoffmann, F; Pavia, H; Potin, P; Poulet, SA; Toth, G. New trends in marine chemical ecology. Estuaries Coasts 2006, 29, 531–551. [Google Scholar]
- Deig, EF; Ehresmann, DW; Hatch, MT; Riedlinger, DJ. Inhibition of herpesvirus replication by marine algae extracts. Antimicrob. Agents Chemother 1974, 6, 524–525. [Google Scholar]
- Jha, RK; Zi-rong, X. Biomedical compounds from marine organisms. Mar Drugs 2004, 2, 123–146. [Google Scholar]
- Newman, DJ; Cragg, GM; Snader, KM. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod 2003, 66, 1022–1037. [Google Scholar]
- Lindequest, U; Schweder, T. Marine biotechnology. In Biotechnology; Rehm, HJ, Reed, G, Eds.; Wliey-VHC: Weinheim, Germany, 2001; Volume 10, pp. 441–484. [Google Scholar]
- Linsert, P. Revolution in infant formula brewing in Herman’s calves. DNA algae. Genet. Eng. Biotechnol. Monit 1994, 1, 45–46. [Google Scholar]
- Khotimchenko, SV; Vaskovsky, VE; Titlyanavo, TV. Fatty acids of marine algae from the 12 substances from marine algae Puerto Rico. Antimicrob. Agents Chemother 1963, 161, 68–72. [Google Scholar]
- Ryther, JH; Goldman, JC; Gifford, CE. Physical models of integrated waste recycling marine polyculture systems. Aquaculture 1975, 5, 163–177. [Google Scholar]
- Capo, TR; Jaramillo, JC; Boyd, AE; Lapointe, BE; Serafy, JE. Sustained high yields of Gracilaria (Rodophyta) grown in intensive large-scale culture. J. Appl. Phycol 1999, 11, 143–147. [Google Scholar]
- Guiry, MD. AlgaeBase; Martin Ryan Institute, National University of Ireland: Galway, Ireland, 1996–2011. Available online: http://www.algaebase.org accessed on 14 May 2011.
- Skriptsova, AV; Titlyanova, TV; Titlyanov, EA. Red algae of the genus Gracilaria in south of the Russian far east. Russ. J. Mar. Biol 2001, 27, S38–S52. [Google Scholar]
- Kain, JM; Destombe, C. A review of the life history, reproduction and phenology of Gracilaria. J. Appl. Phycol 1995, 7, 69–281. [Google Scholar]
- Misawa, M. Production of natural substances by plant cell cultures described in japanese patents. In Plant Tissue Culture Its Bio-Technol; Barz, W, Reinhard, E, Zenk, MH, Eds.; Springer-Verlag: Berlin, Germany, 1977; pp. 17–26. [Google Scholar]
- Murano, E; Toffanin, R; Paoletti, S; Rizzo, R. Pyruvate-rich agarose from the red alga Gracilaria dura. Planta Med 1992, 58, A588–A589. [Google Scholar]
- Hemmingson, JA; Furneaux, RX; Murray-brown, VH. Biosynthesis of agar polysaccharides in Gracilaria chilensis bird. Carbohydr. Res 1996, 287, 101–115. [Google Scholar]
- Brasch, DJ; Chuah, CT; Melton, LD. Marine algal polysaccharides, Part 2. The agar-type polysaccharide from the red alga Gracilaria secundata. Carbohydr. Res 1983, 115, 191–198. [Google Scholar]
- Glombitza, KW. Marine Algae in Pharmaceutical Science; Hoppe, HA, Levring, T, Eds.; Walter de Gruyter: New York, NY, USA, 1979; Volume 1, pp. 303–342. [Google Scholar]
- Glickman, M. Utilisation of seaweed hydrocolloids in the food industry. Hydrobiology 1987, 151/152, 31–47. [Google Scholar]
- Imbs, AB; Vologodskaya, AV; Nevshupova, NV; Khotimchenko, SV; Titlyanoy, EA. Response of prostaglandin content in the red alga Gracilaria verrucosa to season and solar irradiance. Phytochemistry 2001, 58, 1067–1072. [Google Scholar]
- Chapman, VJ; Chapman, DJ. Seaweeds and Their Uses; Springer-Verlag: Berlin, Germany, 1980. [Google Scholar]
- Sajiki, J. Effect of acetic acid treatment on the concentrations of arachidonic acid and prostaglandin E2 in the red algae, Gracilaria asiatica and G. rhodocaudata. Fish Sci 1997, 63, 128–131. [Google Scholar]
- Fusetani, N; Hashimoto, K. Prostaglandin E2: A candidate for causative agent of “Ogonori” poisoning. Bull. Jpn. Soc. Sci. Fish 1984, 50, 465–469. [Google Scholar]
- Yotsu-yamashita, M; Haddock, RL; Yasumoto, T. Polycavernoside A: A novel glycosidic macrolide from the red alga Polycavernosa tsudai (Gracilaria edulis). J. Am. Chem. Soc 1993, 115, 1147–1148. [Google Scholar]
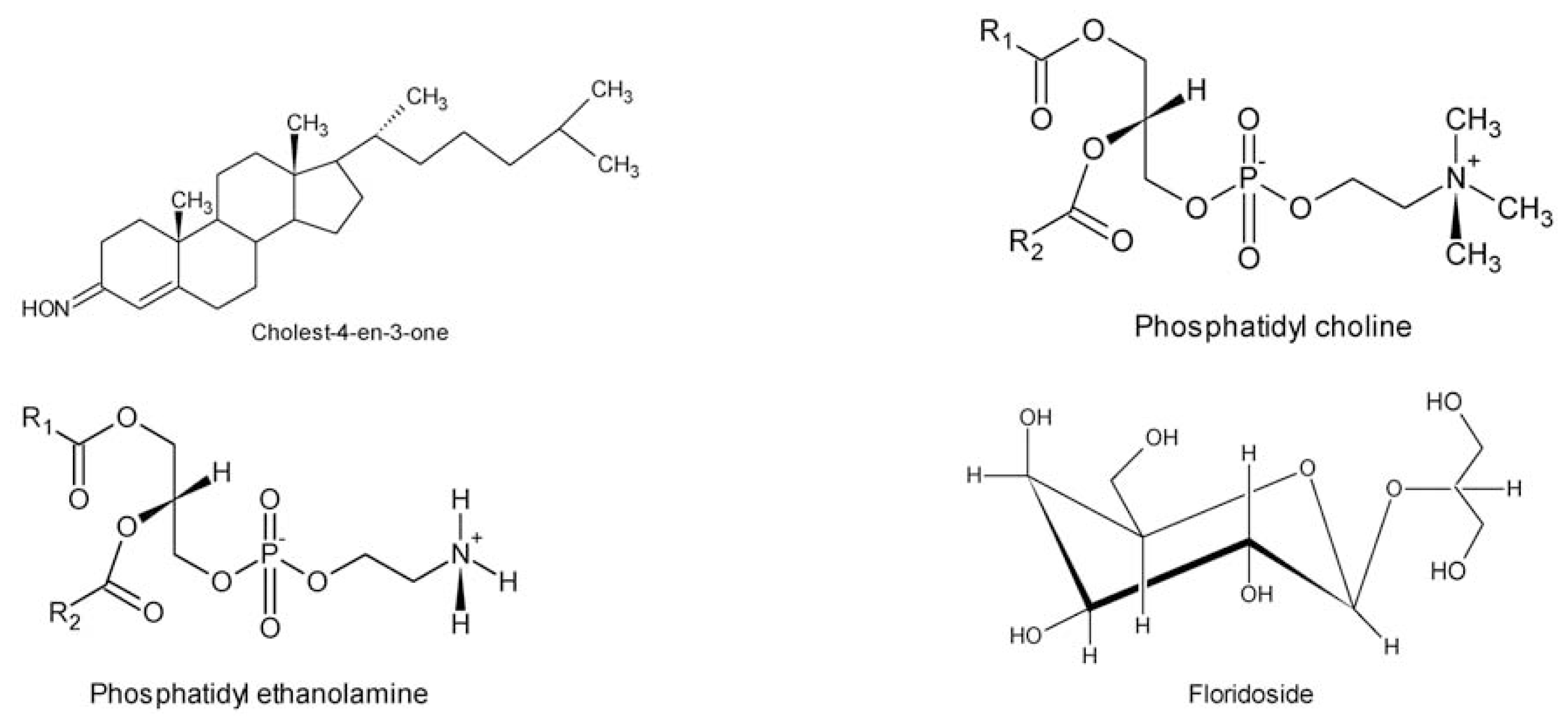
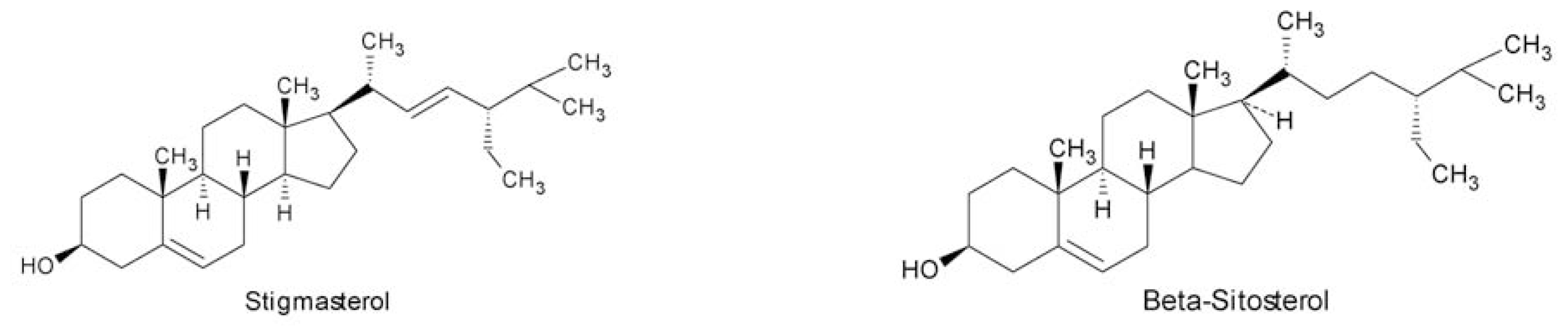
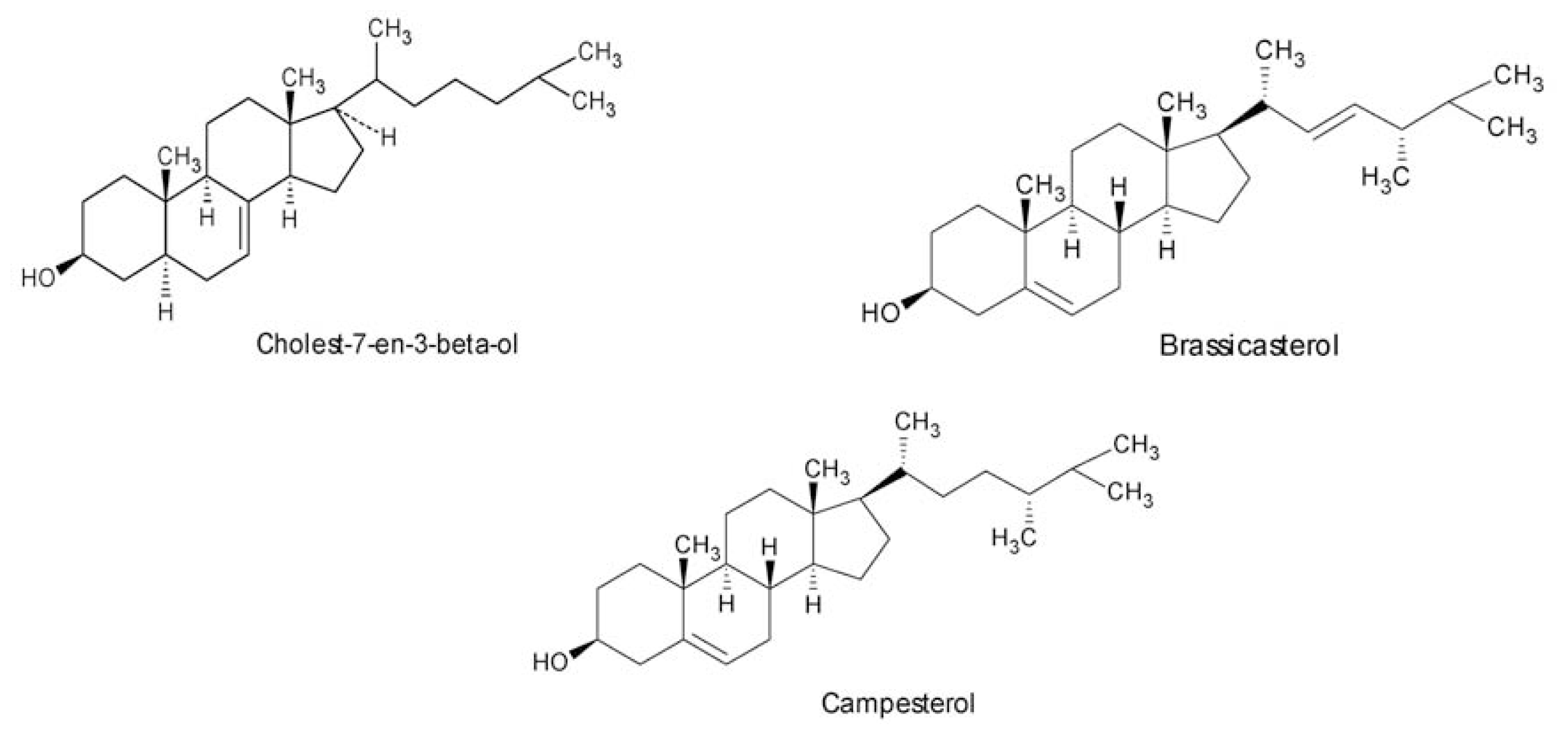
- Das, B; Srinivas, KVNS. Minor C29-steroids from the marine red alga, Gracilaria edulis. Phytochemistry 1992, 31, 2427–2429. [Google Scholar]
- Das, B; Srinivas, KVNS. Dihydroxysterols from the marine red alga, Gracilaria edulis. Phytochemistry 1992, 31, 4731–4373. [Google Scholar]
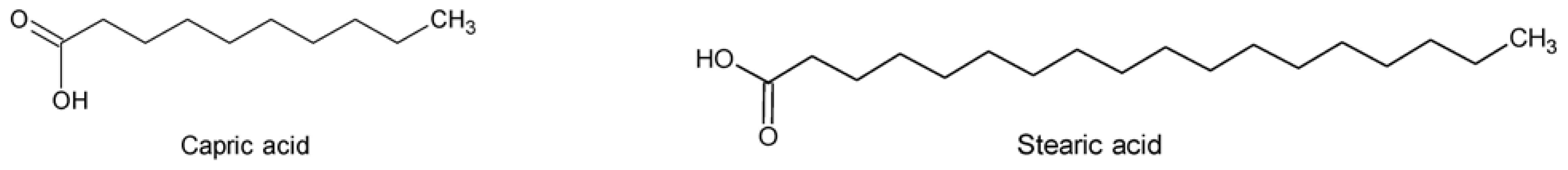
- Combres, A; Bianchini, JP; Gaydou, EM. Fatty acid and sterol composition of red algae of the Indian ocean. Oceanol. Acta 1986, 9, 339–342. [Google Scholar]
- Toffanin, R; Murano, E; Modricky, C; Kvam, BJ; Paoletti, S; Rizzo, R; Pollesello, P. Free and acylated cholesterol in the red alga Gracilaria longa: detection and quantification by 1H- and 13C-NMR on lipid extracts. Planta Med 1992, 58, A589–A590. [Google Scholar]
- Castedo, L; Quintela, JM; Vilalta, R. Sterols from red and brown algae from the Galician coast. An. Quim. Ser. C 1985, 8, 113–115. [Google Scholar]
- Vilalta, R; Quintela, JM; Riguera, R; Castedo, L. Natural marine products from algae and cnidaria of the Galician estuaries. Cuad. Area Cienc. Mar 1984, 2, 617–625. [Google Scholar]
- Henriquez, P; Trucco, R; Silva, M; Sammes, PG. Cholesterol in Iridaea laminarioides and Gracilaria verrucosa. Phytochemistry 1972, 11, 1171. [Google Scholar]
- Anonymous. Prostaglandin E-2. Patent-Japan Kokai Tokkyo Koho 1982, 59, 73–565. [Google Scholar]
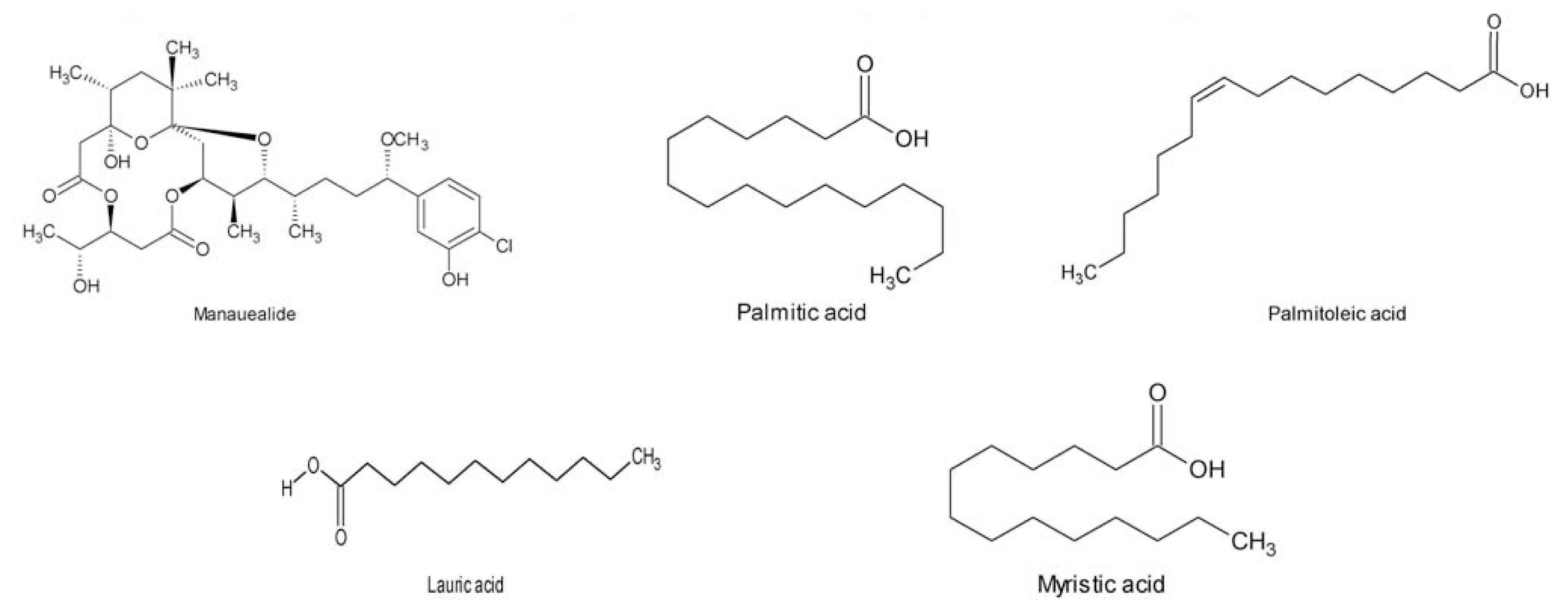
- Araki, S; Sakurai, T; Oohusa, T; Kayama, M. Component fatty acid of lipid from Gracilaria verrucosa. Bull. Jpn. Soc. Sci. Fish 1986, 52, 1871–1874. [Google Scholar]
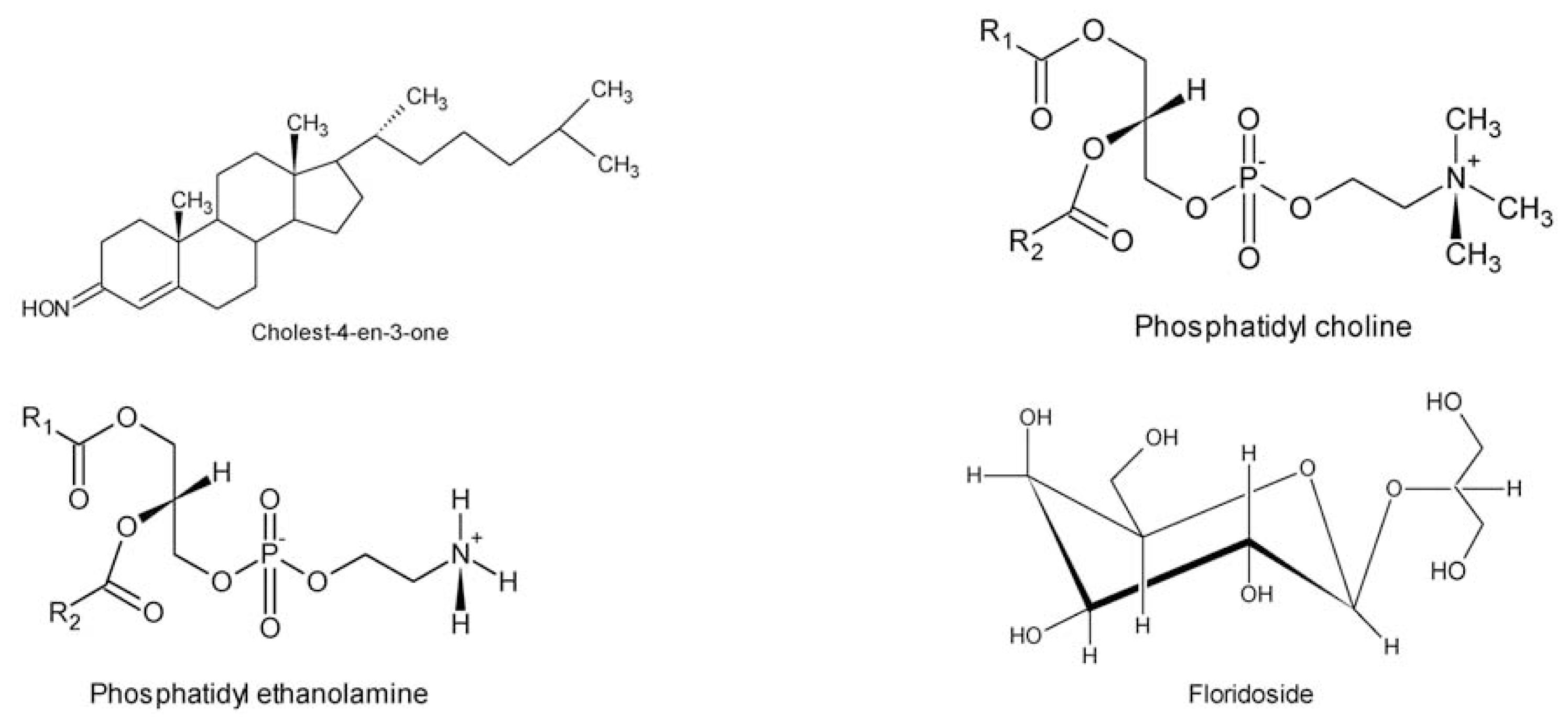
- Son, BW. Glycolipids from Gracilaria verrucosa. Phytochemistry 1990, 29, 307–309. [Google Scholar]
- Das, B; Srinivas, KVNS. Two new sterols from the marine red alga Gracilaria edulis. Planta Med 1993, 59, 572–573. [Google Scholar]
- Sajiki, J; Hakimi, H. Identification of eicosanoids in the red algae, Gracilaria asiatica, using high-performance liquid chromatography and electrospray ionization mass spectrometry. J. Chromatogr 1998, 795, 227–237. [Google Scholar]
- Ravi, BN; Faulkner, DJ. Acyclic diterpenes from the marine sponge Didiscus sp. J. Org. Chem 1979, 44, 968–970. [Google Scholar]
- Sims, JJ; Pettus, JA, Jr. Isolation of free cis and trans-phytol from the red alga Gracilaria andersoniana. Phytochemistry 1976, 15, 1076–1077. [Google Scholar]
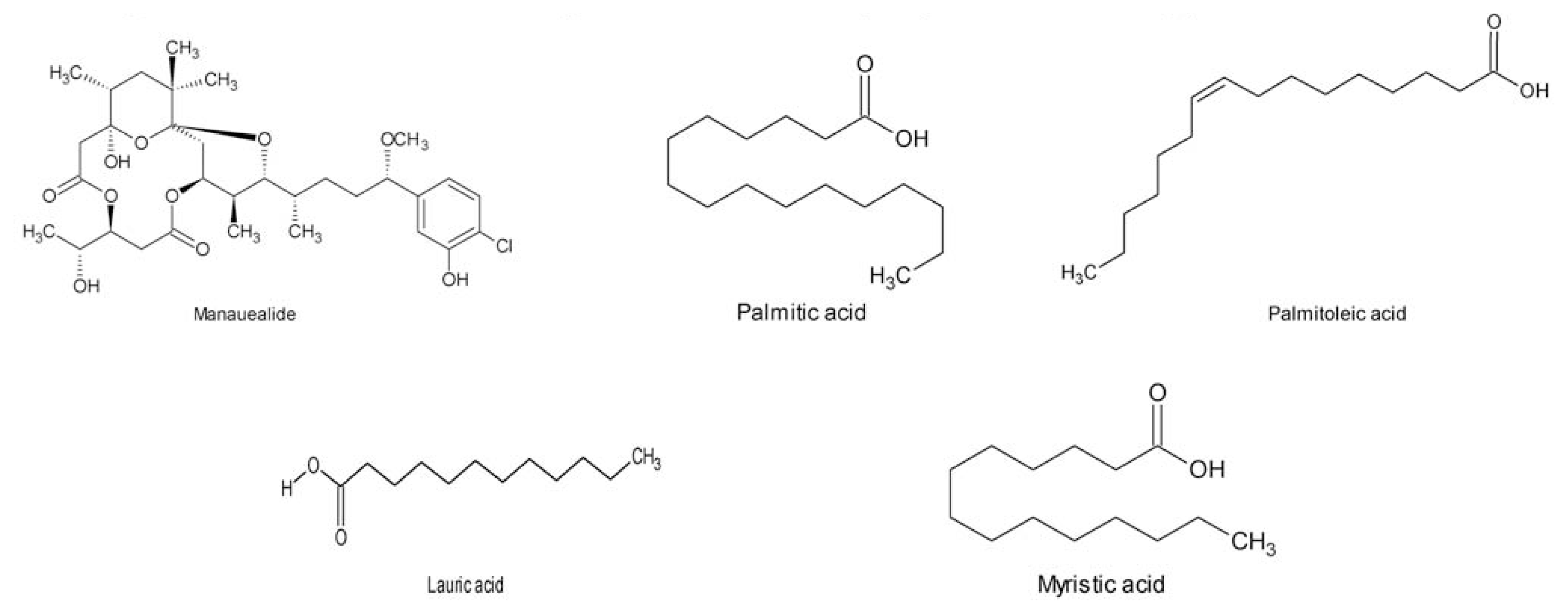
- Nagai, H; Yasumoto, T; Hokama, Y. Manauealides, some of the causative agents of a red alga Gracilaria coronopifolia poisoning in Hawaii. J. Nat. Prod 1997, 60, 925–928. [Google Scholar]
- Nagai, H; Yasumoto, T; Hokama, Y. Aplysiatoxin and debromoaplysiatoxin as the causative agents of a red alga Gracilaria coronopifolia poisoning in Hawaii. Toxicon 1996, 34, 753–761. [Google Scholar]
- Yotsu-yamashita, M; Seki, T; Paul, VJ; Naoki, H; Yasumoto, T. Four new analogs of polycavernoside A. Tetrahedron Lett 1995, 36, 5563–5566. [Google Scholar]
- D’agnolo, E; Rizzo, R; Paoletti, S; Murano, E. R-phycoerythrin from the red alga Gracilaria longa. Phytochemistry 1994, 35, 693–696. [Google Scholar]
- Talarico, L; Kosovel, V. Properties and ultrastructures of r-phycoerythrin from Gracilaria verrucosa. Photosynthetica 1978, 12, 369–374. [Google Scholar]
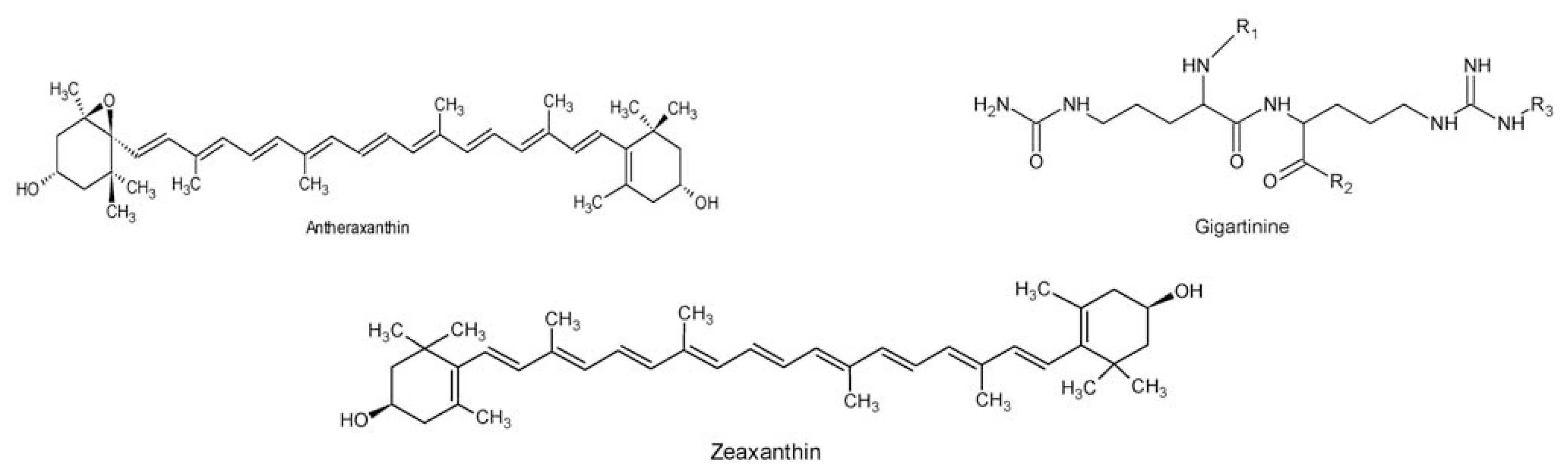
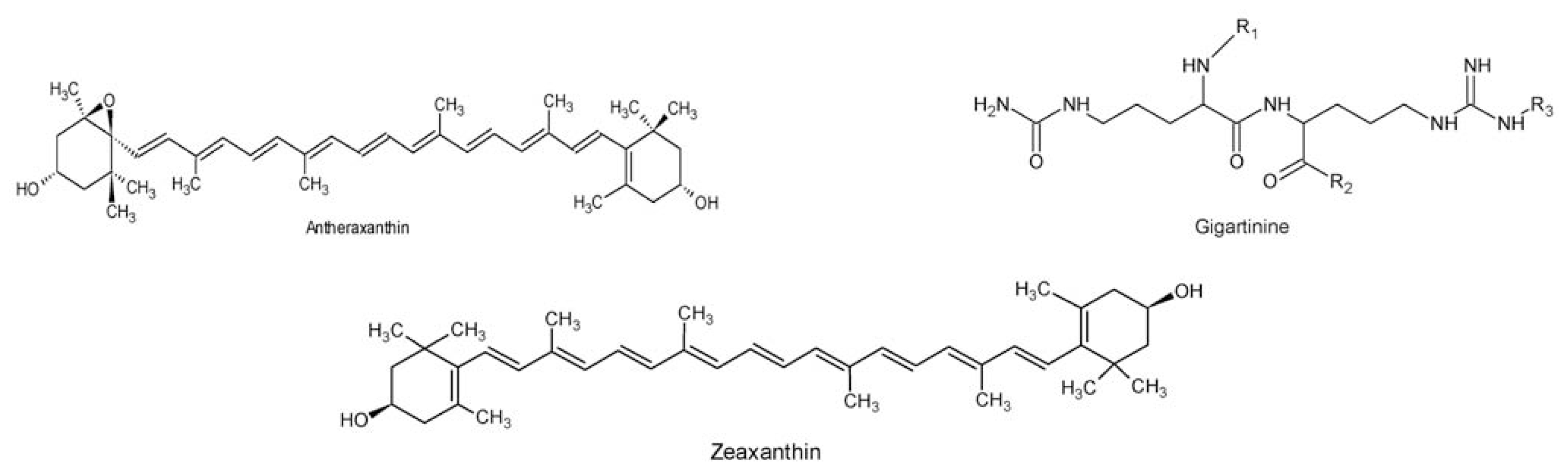
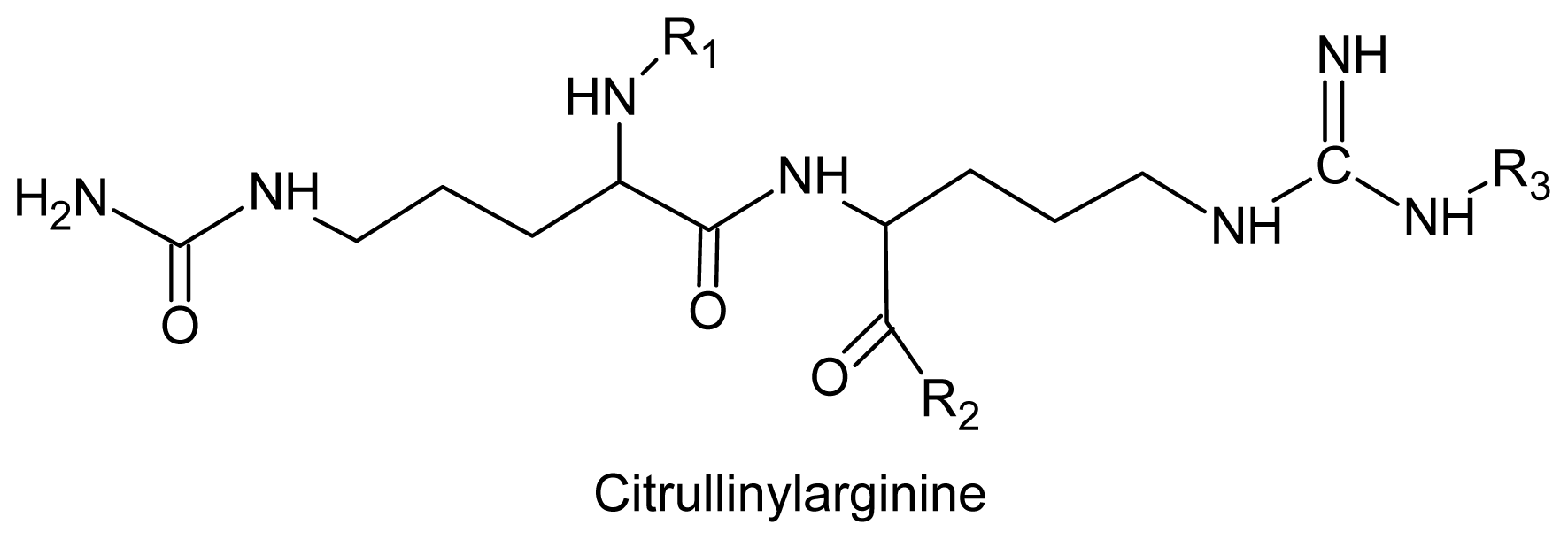
- Wilcox, SJ; Bloor, SJ; Hemmingson, JA; Furneaux, RH; Nelson, WA. The presence of gigartinine in a New Zealand Gracilaria species. J. Appl. Phycol 2001, 13, 409–413. [Google Scholar]
- Kanoh, H; Kitamura, T; Kaboyashi, Y. A sulfated proteoglycan from the red alga Gracilaria verrucosa is a hemagglutinin. Comp. Biochem. Physiol. B 1992, 102, 445–449. [Google Scholar]
- Kirkpatrick, P. Antibacterial drugs. Stitching together naturally. Nat. Rev. Drug Discovery 2002, 1, 748–748. [Google Scholar]
- Kosovel, V; Avanzini, A; Scarcia, V; Furlani, A. Algae as possible sources of antitumoural agents: Preliminary evaluation of the “in vitro” cytostatic activity of crude extracts. Pharmacol. Res. Commun 1988, 20, 27–31. [Google Scholar]
- Kamat, SY; Wahidulla, S; D’Souza, L; Naik, CG; Ambiye, V; Bhakuni, DS; Goel, AK; Garg, HS; Srimal, RC. Bioactivity of marine organisms. VI. Antiviral evaluation of marine algal extracts from the Indian Coast. Bot. Mar 1992, 35, 161–164. [Google Scholar]
- Lhullier, C; Horta, PA; Falkenberg, M. Avaliação de extratos de macroalgas bênticas do litoral catarinense utilizando o teste de letalidade para Artemia salina. Rev. Bras. Farmacogn 2006, 16, 158–163. [Google Scholar]
- Bhakuni, DS; Dhawan, BN; Garg, HS; Goel, AK; Mehrotra, BN; Srimal, RC; Srivastava, MN. Bioactivity of marine organisms: Part VI-Screening of some marine flora from Indian coasts. Indian J. Exp. Biol 1992, 30, 512–517. [Google Scholar]
- Chenieux, JC; Verbist, JF; Biard, JF; Clement, E; Le Boterff, J; Maupas, P; Lecocq, M. Seaweeds of French atlantic coast with antimitotic activity. Planta Med 1980, 40, 152–162. [Google Scholar]
- Arisawa, M; Hayashi, K; Nikaido, T; Koike, K; Fujita, D; Nunomura, N; Tanaka, M; Sasaki, T. Screening of some marine organism extracts for camp phosphodiesterase inhibition, cytotoxicity, and antiviral activity against HSV-1. Int. J. Pharmacogn 1997, 35, 6–11. [Google Scholar]
- Numata, A; Kanbara, S; Takahashi, C; Fujiki, R; Yoneda, M; Fujita, E; Nabeshima, Y. Cytotoxic activity of marine algae and a cytotoxic principle of the brown alga Sargassum tortile. Chem. Pharm. Bull 1991, 39, 2129–2131. [Google Scholar]
- Avanzini, A; Kosovel, V; Scarcia, V; Furlani, A; Ravalico, L. Green, red and brown algae from North Adriatic sea as source of possible cytotoxic products. Fitoterapia 1987, 58, 391–394. [Google Scholar]
- Ratnasooriya, WD; Premakumara, GAS; Tillekeratne, LMV. Post-coital contraceptive activity of crude extracts of Sri Lankan marine red algae. Contraception 1994, 50, 291–299. [Google Scholar]
- Okada, Y; Miyauch, N; Suzuki, K; Kobayashi, T; Tsutsui, C; Mayuzumi, K; Okuyama, T. Search for naturally occurring substances for prevention against the complications of diabetes; inhibitory effect on aldose reductase and platelet aggregation. Nat. Med 1994, 48, 324–329. [Google Scholar]
- Yoshizawa, Y; Tsunehiro, J; Nomura, K; Itoh, M; Fukui, F; Ametani, A; Kaminogawa, S. In vivo macrophage-stimulation activity of the enzyme-degraded water-soluble polysaccharide fraction from a marine alga (Gracilaria verrucosa). Biosci. Biotechnol. Biochem 1996, 60, 1667–1671. [Google Scholar]
- Choi, JS; Lee, JH; Park, HJ; Kim, HG; Young, HS; Mun, SI. Screening for antioxidant activity of plants and marine algae and its active principles from Prunus davidiana. Korean J. Pharmacogn 1993, 24, 299–303. [Google Scholar]
- Funayama, S; Hikino, H. Hypotensive principles from plants. Heterocycles 1981, 15, 1239–1256. [Google Scholar]
- Gregson, RP; Marwood, JF; Quinn, RJ. The occurrence of prostaglandins PGE2 and PGF2α in a plant-the red alga Gracilaria lichenoides. Tetrahedron Lett 1979, 20, 4505–4506. [Google Scholar]
- Choi, BW; Lee, BH; Kang, KJ; Lee, ES; Lee, NH. Screening of the tyrosinase inhibitors from marine algae and medicinal plants. Korean J. Pharmacogn 1998, 29, 237–242. [Google Scholar]
- Sridhar, P; Lakshmi, VV; Polasa, H; Reddy, VS; Rao, CHP; Srimannarayana, G. Biological activity of some marine algal extracts. Indian J. Mar. Sci 1984, 13, 90–91. [Google Scholar]
- Bitou, N; Inomiya, M; Tsujita, T; Okuda, H. Screenning of lipase inhibitors from marine algae. Lipids 1999, 34, 441–445. [Google Scholar]
- Perez, RM; Avila, JG; Perez, S; Martinez, A; Martinez, G. Antimicrobial activity of some American algae. J. Ethnopharmacol 1990, 29, 111–116. [Google Scholar]
- Salleh, A; Wati Haron, N; Mahmud, N; Mohammad, J. Distribution of pyrimidine derivatives in algae. Biochem. Syst. Ecol 1994, 22, 860. [Google Scholar]
- Usmanghani, K; Shameel, M; Sualeh, M; Khan, KH; Mahmood, ZA. Antibacterial and antifungal activities of marine algae from Karachi seashore of Pakistan. Fitoterapia 1984, 55, 73–77. [Google Scholar]
- Albuquerque, MR; Campos-Takaki; Koening, ML. Detection of antimicrobial activity in marine seaweeds. Rev. Inst. Antibiot. Univ. Fed Pernambuco Recife 1983, 21, 127–138. [Google Scholar]
- Oranday, MA; Verde, MSJ; Martínez-Lozano, NH; Waksman, J. Active fractions from four species of marine algae. Phyton 2004, 73, 165–170. [Google Scholar]
- Lustigman, B; Lee, LH; Thees, N; Masucci, J. Production of antibacterial substances by macroalgae of the New York/New Jersey coast, USA. Bull. Environ. Contam. Toxicol 1992, 49, 743–749. [Google Scholar]
- Couladis, M; Vagias, C; Roussis, V; Verykokidou, E; Skaltsa, H. Antiphage properties of some Greek algae extracts. Phytomedicine 1998, 5, 479–483. [Google Scholar]
- Hayashi, K; Hamada, J; Hayashi, T. A screening strategy for selection of anti-HSV-1 and anti- HIV extracts from algae. Phytother. Res 1996, 10, 233–237. [Google Scholar]
- Kamat, SY; Wahidulla, S; D’Souza, L; Naik, CG; Ambiye, V; Bhakuni, DS; Goel, AK; Garg, HS; Srimal, RC. Bioactivity of marine organisms. VI. Antiviral evaluation of marine algal extracts from the Indian Coast. Bot. Mar 1992, 35, 161–164. [Google Scholar]
- Nakamura, H; Ohnuki, N; Sadamasu, K; Sekine, H; Tanaka, J; Okada, Y; Okuyama, T. Anti-human immunodeficiency virus (HIV) activities of aqueous extracts from marine algae. Nat. Med 1994, 48, 173–179. [Google Scholar]
- Ohigashi, H; Sakai, Y; Yamaguchi, K; Umezaki, I; Koshimizu, K. Possible anti-tumor promoting properties of marine algae and in vivo activity of wakame seaweed extract. Biosci. Biotechnol. Biochem 1992, 56, 994–995. [Google Scholar]
- Hsu, B-Y; Tsao, C-Y; Chiou, T-K; Hwang, D-F. Factors affeecting PGE2 production in seaweed Gracilaria tenuistipitata. J. Food Drug Anal 2008, 16, 59–65. [Google Scholar]
- Laserna, EC; Veroy, RL; Luistro, AH; Cajipe, GJB. Extracts from some red and brown seaweeds of the Philippines. In Tenth International Seaweed Symposium; Levring, T, Ed.; Walter de Gruyter & Co: Berlin, Germany, 1980; pp. 443–448. [Google Scholar]
- Nagai, H; Kan, Y; Fujita, T; Sakamoto, B; Kokama, Y. Manauealide C and anhydrodebromoaplysiatoxin, toxic constituents of the hawaiian red alga, Gracilaria coronopifolia. Biosci. Biotechnol. Biochem 1998, 62, 1011–1013. [Google Scholar]
- Parekh, KS; Parekh, HH; Rao, PS. Fatty acid content of some Indian marine algae. Indian J. Mar. Sci 1984, 13, 45–46. [Google Scholar]
- Kan, Y; Fujita, T; Nagai, H; Sakamoto, B; Hokama, Y. Malyngamides M and N from the hawaiian red alga Gracilaria coronopifolia. J. Nat. Prod 1998, 61, 152–155. [Google Scholar]
- Parra, AL; Yhebra, RS; Sardiñas, IG; Buela, LI. Comparative study of the assay of Artemia salina L. and the estimate of the medium lethal dose (LD50 value) in mice, to determine oral acute toxicity of plant extracts. Phytomedicine 2001, 8, 395–400. [Google Scholar]
- Meyer, BN; Ferrigni, NR; Putnam, JE; Jacobsen, LB; Nichols, DE; McLaughlin, JL. Brine shrimp: A convenient general bioassay for active plant constituints. Planta Med 1982, 45, 31–34. [Google Scholar]
- Kanazawa, A; Yoshioka, M. Occurrence of Cholest-4-en-3-one in the Red Alga. Gracilaria textorii Proceedings of the Seventh International Seaweed Symposium, Sapporo, Japan, 1–12 August 1971; Wiley: New York, NY, USA, 1972; 7, pp. 502–505. [Google Scholar]
- Kunzler, K; Eichenberger, W. Betaine lipids and zwitterionic phospholipids in plants and fungi. Phytochemistry 1997, 46, 883–892. [Google Scholar]
- Roh, YS; Son, W; Im, KS; Choi, HD. Structure of floridoside, a glycerol glycoside from the marine red alga Gracilaria verrucosa. Korean J. Pharmacogn 1994, 25, 117–120. [Google Scholar]
- Laserna, EC; Veroy, RL; Luistro, AH; Cajipe, GJB. Extracts from some red and brown seaweeds of the Philippines. In Tenth International Seaweed Symposium; Levring, T, Ed.; Walter de Gruyter & Co: Berlin, Germany, 1980; pp. 443–448. [Google Scholar]
- Matsuhashi, T. Effects of the freezing and drying method and the mechanical dehydration method by pressure on gel properties of dried agar. Reito 1974, 49, 756–760. [Google Scholar]
- Shi, SY; Zhang, YX; Liu, WQ; Li, z. Seasonal variation in yield, physical properties and chemical composition of agar from Gracilaria verrucosa. Oceanol. Limnol. Sin 1983, 14, 272–278. [Google Scholar]
- Friedlander, M; Lipkin, Y; Yaphe, W. Composition of agars from Gracilaria verrucosa and Pterocladia capillacea. Bot. Mar 1981, 24, 595–598. [Google Scholar]
- Minghetti, L; Levi, G. Microglia as effector cells in brain damage and repair: Focus on prostanoids and nitric oxide. Prog. Neurobiol 1998, 54, 99–125. [Google Scholar]
- Nishihara, I; Minami, T; Uda, R; Ito, S; Hyodo, M; Hayaishi, O. Effect of NMDA receptor antagonists on prostaglandin E2-induced hyperalgesia in conscious mice. Brain Res 1995, 677, 138–144. [Google Scholar]
- Aihara, MS; Yamamoto, H. Occurrence of antheraxanthin in two Rhodophyceae Acanthophora spicifera and Gracilaria lichenoides. Phytochemistry 1968, 7, 497–499. [Google Scholar]
- Tuney, I; Çadirci, BH; Unal, D; Sukatar, A. Antimicrobial activities of the extracts of marine algae from the coast of Urla. Turk. J. Biol 2006, 30, 171–175. [Google Scholar]
- Aihara, MS; Yamamoto, H. Occurrence of antheraxanthin in two Rhodophyceae Acanthophora spicifera and Gracilaria lichenoides. Phytochemistry 1968, 7, 497–499. [Google Scholar]
- Panossian, AG. Search of prostaglandins and related compounds in plants: A review of the occurrence of prostaglandins and prostaglandin-like compounds in plants. prostaglandins 1987, 33, 363–381. [Google Scholar]
- Mahasneh, I; Jamal, M; Kashashneh, M; Zibdeh, M. Antibiotic-activity of marine-algae against multi-antibiotic resistant-bacteria. Microbios 1995, 83, 23–26. [Google Scholar]
- Parekh, KS; Parekh, HH; Rao, PS. Fatty acid content of some Indian marine algae. Indian J. Mar. Sci 1984, 13, 45–46. [Google Scholar]
- Fernandez, LE; Valiente, OG; Mainardi, V; Bello, JL; Velez, H; Rosado, A. Isolation and characterization of an antitumor active agar-type polysaccharide of Gracilaria dominguensis. Carbohydr. Res 1989, 190, 77–83. [Google Scholar]
- Okamoto, R; Hori, K; Miyazawa, K; Ito, K. Isolation and charcterization of a new hemagglutinin from the red alga Gracilaria bursa-pastoris. Experientia 1990, 46, 975–977. [Google Scholar]
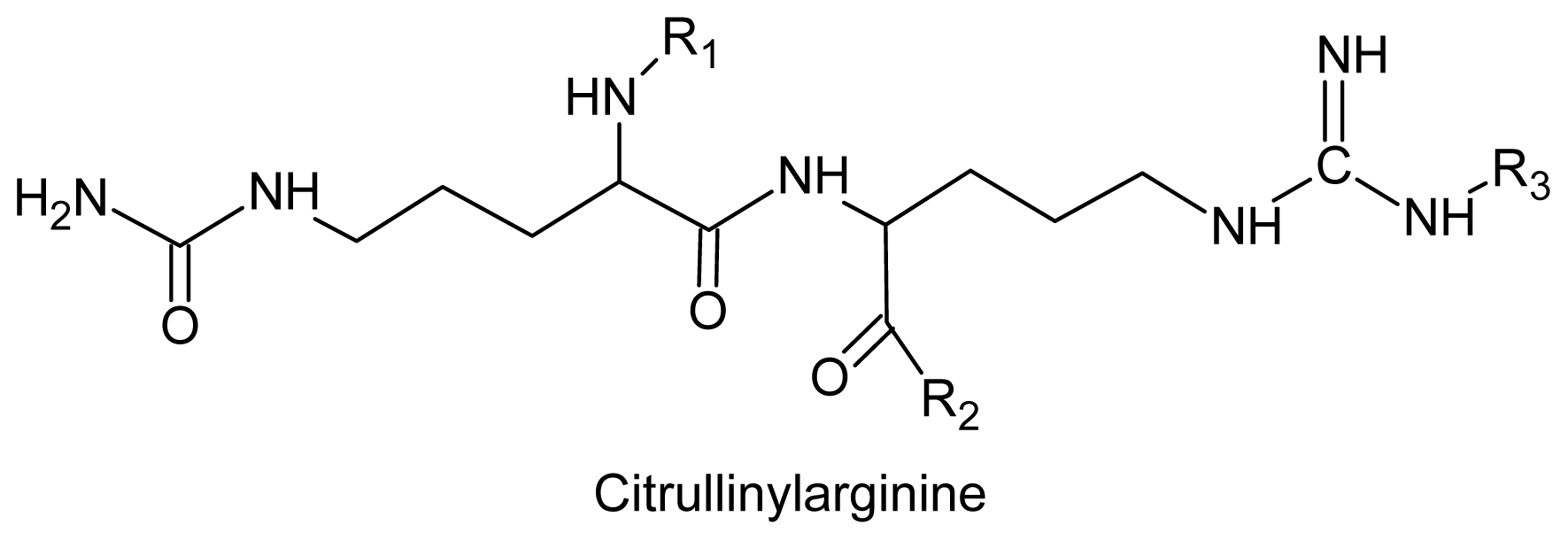
- Laycock, MV; Craigie, JS. The occurrence and seasonal variation of gigartinine and l-citrullinyl-l-arginine in Chondrus crispus stackh. Can. J. Biochem 1977, 55, 27–30. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Botanical Name | Part used | Type of extract | Bioassays models, organism, dose or route of administration | Result |
|---|---|---|---|---|
| Studies of toxicity | ||||
| Gracilaria bursa-pastoris (S.G.Gmelin) P.C.Silva | FzDTh | H2O Ext. | Cytotoxic activity-cell culture-10.0 μg/mL | Inactive [72] |
| FTh | 95% EtOH Ext. or CHCl3 Ext. | Cytotoxic activity-cell culture-10.0 μg/mL | Inactive [72] | |
| Gracilaria chorda (Holmes) | FsO | H2O Ext. | Toxicity assessment-mouse-1.2 mg/animal-i.p. | Active [48] |
| Gracilaria coronopifolia (J. Agardh) | FTh | Plant | Toxic effect-human adult-oral | Active [65] |
| Gracilaria corticata (J.Agardh) J.Agardh | Th | 50% EtOH-H2O Ext. | Toxicity assessment-mouse-DL50 1000 mg/kg-ip | Active [73] |
| Gracilaria domingensis (Kützing) Sonder ex Dickie | DO | 90% EtOH Ext. | Cytotoxity-Artemia salina L.-200 μg/mL | Active [74] |
| Gracilaria edulis (S.G.Gmelin) P.C.Silva | DTh | Plant | Toxicity effect (death)-human adult-oral | Active [49] |
| SDTh | 90% EtOH Ext. | Toxicity assessment-mouse-DL50 0.825 mg/kg-i.p. | Active [75] | |
| Gracilaria foliifera (Forsskål) Borgesen | DEP | (1:1) EtOH-H2O Ext. | Cytotoxic activity-cell culture-dose: dry weight of plant | Active [76] |
| Gracilaria textorii (Suringar) De Toni | FzDO | MeOH Ext. | Cytotoxic activity-cell culture (CA 9 KB) | Inactive [77] |
| FsTh | Hexane Ext. | Cytotoxic activity-culture cell (LEUK P 388)-ED 50 > 100 μg/mL | Equivocal [78] | |
| CCl4 Ext. | Cytotoxic activity-culture cell (LEUK P 388)-ED 50 22.2 μg/mL | Equivocal [78] | ||
| CHCl3 Ext. | Cytotoxic activity-culture cell (LEUK P 388)-ED 50 32.2 μg/mL | Inactive [78] | ||
| Gracilaria verrucosa (Hudson) Papenfuss | DO | H2O Ext. | Toxicity assessment-mouse-1.2 mg/animal-i.p. | Active [48] |
| FzDO | MeOH Ext. | Cytotoxic activity-cell culture (CA 9 KB) | Inactive [77] | |
| FO | 30% EtOH Ext. | Cytotoxic activity-cell culture (CA 9 KB)-10.0 μg/mL | Inactive [79] | |
| (1:1) CHCl3-MeOH Ext. | Cytotoxic activity-cell culture (CA 9 KB)-1.0 μg/mL | Equivocal [79] | ||
| FTh | H2O Ext. and 95% EtOH Ext. | Cytotoxic activity-cell culture (LEUK P 388-P 3)-10.0 μg/μL | Inactive [72] | |
| Effects on the nervous system | ||||
| Gracilaria corticata J.Agardh | SDTh | 90% EtOH Ext. | Autonomic effects-dog-50 mg/kg-iv | Inactive [75] |
| CNS effects-mouse | Inactive [75] | |||
| Analgesic activity-mouse | Inactive [75] | |||
| Anticonvulsant activity-mouse | Inactive [75] | |||
| Gracilaria edulis (S.G.Gmelin) P.C.Silva | SDTh | 90% EtOH Ext. | Autonomic effects-dog-50 mg/kg-iv | Inactive [75] |
| CNS effects-mouse | Inactive [75] | |||
| Analgesic activity-mouse | Inactive [75] | |||
| Anticonvulsant activity-mouse | Inactive [75] | |||
| Gracilaria verrucosa (Hudson) Papenfuss | SDTh | 90% EtOH Ext. | CNS effects-mouse | Inactive [75] |
| Contraception activity | ||||
| Gracilaria corticata J.Agardh | DTh | (1:1) MeOH-CH2Cl2 Ext. | Embryotoxic effect-pregnant rat-1.0 mg/kg-intragastric | Inactive [80] |
| SDTh | 90% EtOH Ext. | Antiimplantation effect-pregnant rat-100.0 mg/kg | Inactive [75] | |
| Spermicidal effect-rat-2.0 % | Inactive [75] | |||
| Gracilaria edulis (S.G.Gmelin) P.C.Silva | SDTh | 90% EtOH Ext. | Antiimplantation effect-pregnant rat-100.0 mg/kg | Inactive [75] |
| Spermicidal effect-rat-2.0% | Inactive [75] | |||
| Gracilaria verrucosa (Hudson) Papenfuss | SDTh | 90% EtOH Ext. | Spermicidal effect-rat-2.0% | Inactive [75] |
| Anti-inflammatory activity | ||||
| Gracilaria textorii (Suringar) De Toni | EP | H2O Ext. | Platelet aggregation inhibition (adenosine diphosphate; arachidonic acid or collagen stimulation)-100.0 μg/mL | Inactive [81] |
| Venotonic activity (platelet aggregating factor stimulation)-100.0 μg/mL | Inactive [81] | |||
| Gracilaria verrucosa (Hudson) Papenfuss | DTh | Polysaccharide fraction | Immunostimulant activity-mouse-4.0 mg/animal-i.p. | Active [82] |
| Phagocytosis stimulation-mouse-4.0 mg/animal-i.p. | Active [82] | |||
| SDTh | 90% EtOH Ext. | Antiinflammatory activity-rat-intragastric | Inactive [75] | |
| Antioxidant activity | ||||
| Gracilaria verrucosa (Hudson) Papenfuss | Plant | MeOH Ext. | Radical scavenging effect (DPPH radicals)-IC50 480.0 μg | Active [83] |
| DTh | Polysaccharide fraction | Oxygen radical formation induction-mouse-4.0 mg/animal-i.p. | Active [82] | |
| Gastrointestinal effects | ||||
| Gracilaria chorda (Holmes) | FsO | H2O Ext. | Mouse-0.5 mg/animal-gastric intubation and dose 0.5 mg/loop-i.p. | Active [48] |
| Gracilaria verrucosa (Hudson) Papenfuss | DO | H2O Ext. | Mouse-0.5 mg/animal-gastric intubation | Active [48] |
| Cardiovascular effects | ||||
| Gracilaria corticata (J.Agardh) J.Agardh | SDTh | 90% EtOH Ext. | Cardiovascular effects-dog-50 mg/kg-iv | Inactive [75] |
| Gracilaria edulis (S.G.Gmelin) P.C.Silva | SDTh | 90% EtOH Ext. | Cardiovascular effects-dog-50 mg/kg-iv | Inactive [75] |
| Diuretic activity-rat-intragastric | Active [75] | |||
| Gracilaria lichenoides (Greville) | EP | H2O Ext. | Antihypertensive activity-rat-iv | Active [84] |
| FsTh | H2O Ext. | Antihypertensive activity-rat-iv | Active [85] | |
| Gracilaria verrucosa (Hudson) Papenfuss | SDTh | 90%EtOH Ext. | Cardiovascular effects-dog-50 mg/kg-iv | Inactive [75] |
| Hypoglycemic activity | ||||
| Gracilaria corticata J.Agardh | SDTh | 90% EtOH Ext. | Rat-250 mg/kg – intragastric | Inactive [75] |
| Gracilaria edulis (S.G.Gmelin) P.C.Silva | SDTh | 90% EtOH Ext. | Rat-250.0 mg/kg – intragastric | Inactive [75] |
| Anti-enzymes activity | ||||
| Gracilaria arcuata (Zanardini) | DTh | MeOH Ext. | Tyrosinase inhibition-0.33 mg/mL | Inactive [86] |
| Gracilaria corticata (J.Agardh) J.Agardh | DO | PET Ether Ext.; CHCl3 Ext. or MeOH Ext. | Penicillinase inhibition-1.0 μg/units | Inactive [87] |
| Gracilaria textorii (Suringar) De Toni | EP | H2O Ext. | Aldose reductase inhibition-10.0 μg/mL | Inactive [81] |
| FzDO | MeOH Ext. | Cyclic AMP phosphodiesterase inhibition | Inactive [77] | |
| Gracilaria verrucosa (Hudson) Papenfuss | FzDO | EtOAc Ext. | Lipase inhibition | Equivocal [88] |
| MeOH Ext. | Cyclic AMP phosphodiesterase inhibition | Inactive [77] | ||
| Respiratory effects | ||||
| Gracilaria corticata (J.Agardh) J.Agardh | SDTh | 90% EtOH Ext. | Respiratory depressant-dog-50 mg/kg-iv | Inactive [75] |
| Gracilaria edulis (S.G.Gmelin) P.C.Silva | SDTh | 90% EtOH Ext. | Respiratory depressant-dog-50 mg/kg-iv | Inactive [75] |
| Gracilaria verrucosa (Hudson) Papenfuss | SDTh | 90% EtOH Ext. | Respiratory depressant-dog-50.0 mg/kg-iv | Inactive [75] |
| Spasmolytic activity | ||||
| Gracilaria corticata (J.Agardh) J.Agardh | SDTh | 90% EtOH Ext. | Spasmolytic activity-guinea pig | Inactive [75] |
| Gracilaria edulis (S.G.Gmelin) P.C.Silva | SDTh | 90% EtOH Ext. | Negative chronotropic effect-dog-50.0 mg/kg-iv | Inactive [75] |
| Antibacterial activity | ||||
| Gracilaria cervicornis (Turner) J.Agardh | DEP | 95% EtOH Ext. | Agar plate-Staphylococcus aureus-5.0 mg/mL | Active [89] |
| Agar plate-Proteus vulgaris; Escherichia coli; Aspergillus fumigates; Candida albicans; Pseudomonas aeruginosa; Streptococcus pyogenes- 50.0 mg/mL | Inactive [89] | |||
| Gracilaria corticata (J.Agardh) J.Agardh | DO | PET Ether Ext.; CHCl3 Ext. or MeOH Ext. | Agar plate-Staphylococcus aureus; Escherichia coli-MIC >200 μg/mL | Inactive [90] |
| FsO | MeOH Ext. | Agar plate-Escherichia coli; Salmonella paratyphi A; Salmonella paratyphi B; Shigella sonnei | Inactive [91] | |
| Agar plate-Bacillus subtilis; Staphylococcus aureus; Bacillus megaterium; Streptococcus viridans | Active [91] | |||
| SDTh | 90% EtOH Ext. | Agar plate-Klebsiella pneumonia; Pseudomonas aeruginosa; Staphylococcus aureus; Escherichia coli; Streptococcus faecalis | Inactive [75] | |
| Gracilaria debilis (Forsskål) Borgesen | DO | 95% EtOH Ext. | Agar plate-Escherichia coli; Staphylococcus aureus | Active [92] |
| Agar plate-Mycobacterium smegmatis Inactive | [92] | |||
| Gracilaria domingensis (Kützing) Sonder ex Dickie | DO | 95% EtOH Ext. | Agar plate-Escherichia coli; Staphylococcus aureus | Active [92] |
| Acetone Ext. or Ether Ext. | Agar plate-Escherichia coli; Staphylococcus aureus | Inactive [92] | ||
| 95% EtOH Ext. or Acetone Ext. | Agar plate-Mycobacterium smegmatis | Active [92] | ||
| Gracilaria edulis (S.G.Gmelin) P.C.Silva | SDTh | 90% EtOH Ext. | Agar plate-Escherichia coli; Streptococcus faecalis; Staphylococcus aureus; Pseudomonas aeruginosa; Klebsiella pneumoniae | Inactive [75] |
| Gracilaria pygmea (Borgesen) | FsO | MeOH Ext. | Agar plate-Bacillus subtilis; Staphylococcus aureus; Escherichia coli; Salmonella paratyphi A; Streptococcus viridans; Shigella sonnei; Salmonella paratyphi B | Inactive [91] |
| Agar plate-Bacillus megaterium | Active [91] | |||
| Gracilaria sjoestedii (Kylin) | DO | 95% EtOH Ext. | Agar plate-Escherichia coli; Staphylococcus aureus | Active [92] |
| Agar plate-Mycobacterium smegmatis | Inactive [92] | |||
| Gracilaria tikvahiae McLachlan | DEP | CHCl3 Ext. or EtOH Ext. | Agar plate -Staphylococcus aureus | Active [93] |
| Agar plate-Streptococcus faecalis; Pseudomonas aeruginosa | Inactive [93] | |||
| Gracilaria verrucosa (Hudson) Papenfuss | FTh | ** | Agar plate-Vibrio marinofulvis; Micrococcus imfimus; Pseudomonas atlantica-40.0 μg/μL | Inactive [94] |
| Th | 70% EtOH Ext. | Antiphage activity-agar plate-Bacteriophage T 1; Bacteriophage T 2; Bacteriophage T 4; Bacteriophage T 7; Bacteriophage MS 2; Bacteriophage PHI-CHI 174-0.50 μg/mL | Inactive [95] | |
| Antifungal activity | ||||
| Gracilaria corticata (J.Agardh) J.Agardh | FsO | MeOH Ext. | Agar plate-Aspergillus niger; Fusarium solani; Alternaria solani; Penicillium funiculosum | Inactive [91] |
| SDTh | 90% EtOH Ext. | Agar plate-Sporotrichum schenckii; Cryptococcun neoformans; Candida albicans; Trichophyton mentagrophytes; Aspergillus fumigates | Inactive [75] | |
| Gracilaria debilis (Forsskål) Borgesen | DO | 95% EtOH Ext. | Agar plate-Candida albicans | Active [92] |
| Agar plate-Neurospora crassa | Inactive [92] | |||
| Gracilaria domingensis (Kützing) Sonder ex Dickie | DO | 95% EtOH Ext. and Acetone Ext. | Agar plate-Candida albicans; Neurospora crassa | Active [92] |
| Ether Ext. | Agar plate-Candida albicans | Inactive [92] | ||
| Gracilaria edulis (S.G.Gmelin) P.C.Silva | SDTh | 90% EtOH Ext. | Agar plate-Sporotrichum schenckii; Candida albicans; Cryptococcus neoformans; Trichophyton mentagrophytes; Aspergillus fumigates | Inactive [75] |
| Gracilaria pygmea (Borgesen) | FsO | MeOH Ext. | Agar plate-Aspergillus niger; Fusarium solani; Alternaria solani; Penicillium funiculosum | Inactive [91] |
| Gracilaria sjoestedii (Kylin) | DO | 95% EtOH Ext. | Agar plate-Candida albicans | Active [92] |
| Agar plate-Neurospora crassa | Inactive [92] | |||
| Gracilaria tikvahiae McLachlan | DEP | CHCl3 Ext. and EtOH Ext. | Agar plate-Candida albicans | Active [93] |
| Antiviral activity | ||||
| Gracilaria bursa-pastoris (S.G.Gmelin) P.C.Silva | FzDTh | ** | Cell culture-Herpes simplex 1 and HIV Virus | Inactive [96] |
| Gracilaria corticata (J.Agardh) J.Agardh | Th | 50% EtOH-H2O Ext. | Cell culture-Semlicki-forest Virus-0.05 mg/mL | Equivocal [73] |
| Cell culture-Ranikhet and Vaccinia Virus-0.05 mg/mL | Inactive [73] | |||
| SDTh | 90% EtOH Ext. | Cell culture-Ranikhet Virus | Inactive [75] | |
| Gracilaria edulis (S.G.Gmelin) P.C.Silva | SDTh | 90% EtOH Ext. | Cell culture-Semlicki-forest and Ranikhet Virus | Inactive [75] |
| Gracilaria pacifica (I. A. Abbott) | DO | MeOH Ext. | Cell culture-Herpes simplex 1 Virus-400.0 μg/mL | Inactive [97] |
| Cell culture-Virus sindbis-200.0 μg/mL | Active [97] | |||
| Gracilaria species | FzDTh | ** | Cell culture-Herpes simplex 1 and HIV Virus | Inactive [96] |
| Gracilaria textorii (Suringar) De Toni | FzDO | MeOH Ext. | Cell culture-Herpes simplex 1 Virus | Inactive [77] |
| Th | H2O Ext. | Cell culture-HIV Virus-MIC > 1000 μg/mL | Inactive [98] | |
| Fresh | MeOH Ext. | Epstein-Barr virus early antigen activation inhibition (telocidin b-4 induced | ** [99] | |
| Th | Epstein-Barr virus induced activation)-4.0 μg/mL | |||
| Gracilaria verrucosa (Hudson) Papenfuss | FzDO | MeOH Ext. | Cell culture-Herpes simplex 1 Virus | Inactive [77] |
| Antiprotozoal activity | ||||
| Gracilaria corticata (J.Agardh) J.Agardh | SDTh | 90% EtOH Ext. | Agar plate-Entamoeba histolytica; Plasmodium berghei | Inactive [75] |
| Gracilaria edulis (S.G.Gmelin) P.C.Silva | SDTh | 90% EtOH Ext. | Agar plate-Entamoeba histolytica; Plasmodium berghei | Inactive [75] |
| Allelophatic activity | ||||
| Gracilaria compressa (C.Agardh) Greville | DEP | 95% EtOH Ext. | Agar plate-Helianthus tuberosus-dose: dry weight of plant | Active [76] |
| Gracilaria foliifera (Forsskål) Borgesen | DEP | H2O Ext. | Agar plate-Helianthus tuberosus-dose: dry weight of plant | Active [76] |
| Gracilaria verrucosa (Hudson) Papenfuss | DEP | 95% EtOH Ext. | Agar plate-Helianthus tuberosus-dose: dry weight of plant | Active [76] |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
De Almeida, C.L.F.; De S. Falcão, H.; De M. Lima, G.R.; De A. Montenegro, C.; Lira, N.S.; De Athayde-Filho, P.F.; Rodrigues, L.C.; De Souza, M.d.F.V.; Barbosa-Filho, J.M.; Batista, L.M. Bioactivities from Marine Algae of the Genus Gracilaria. Int. J. Mol. Sci. 2011, 12, 4550-4573. https://doi.org/10.3390/ijms12074550
De Almeida CLF, De S. Falcão H, De M. Lima GR, De A. Montenegro C, Lira NS, De Athayde-Filho PF, Rodrigues LC, De Souza MdFV, Barbosa-Filho JM, Batista LM. Bioactivities from Marine Algae of the Genus Gracilaria. International Journal of Molecular Sciences. 2011; 12(7):4550-4573. https://doi.org/10.3390/ijms12074550
Chicago/Turabian StyleDe Almeida, Cynthia Layse F., Heloina De S. Falcão, Gedson R. De M. Lima, Camila De A. Montenegro, Narlize S. Lira, Petrônio F. De Athayde-Filho, Luis C. Rodrigues, Maria de Fátima V. De Souza, José M. Barbosa-Filho, and Leônia M. Batista. 2011. "Bioactivities from Marine Algae of the Genus Gracilaria" International Journal of Molecular Sciences 12, no. 7: 4550-4573. https://doi.org/10.3390/ijms12074550
APA StyleDe Almeida, C. L. F., De S. Falcão, H., De M. Lima, G. R., De A. Montenegro, C., Lira, N. S., De Athayde-Filho, P. F., Rodrigues, L. C., De Souza, M. d. F. V., Barbosa-Filho, J. M., & Batista, L. M. (2011). Bioactivities from Marine Algae of the Genus Gracilaria. International Journal of Molecular Sciences, 12(7), 4550-4573. https://doi.org/10.3390/ijms12074550