Effect of Tween Series on Growth and cis-9, trans-11 Conjugated Linoleic Acid Production of Lactobacillus acidophilus F0221 in the Presence of Bile Salts


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
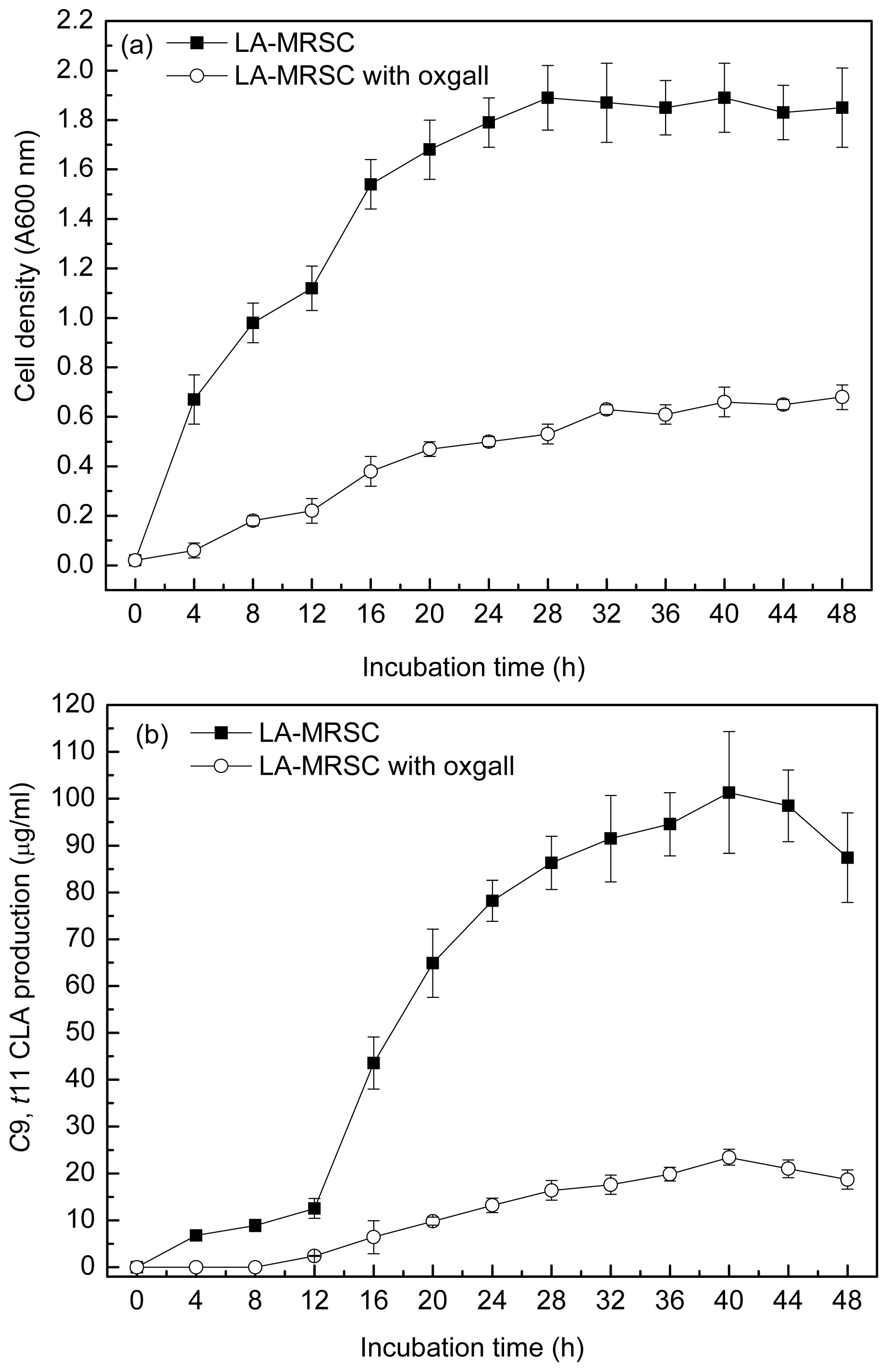
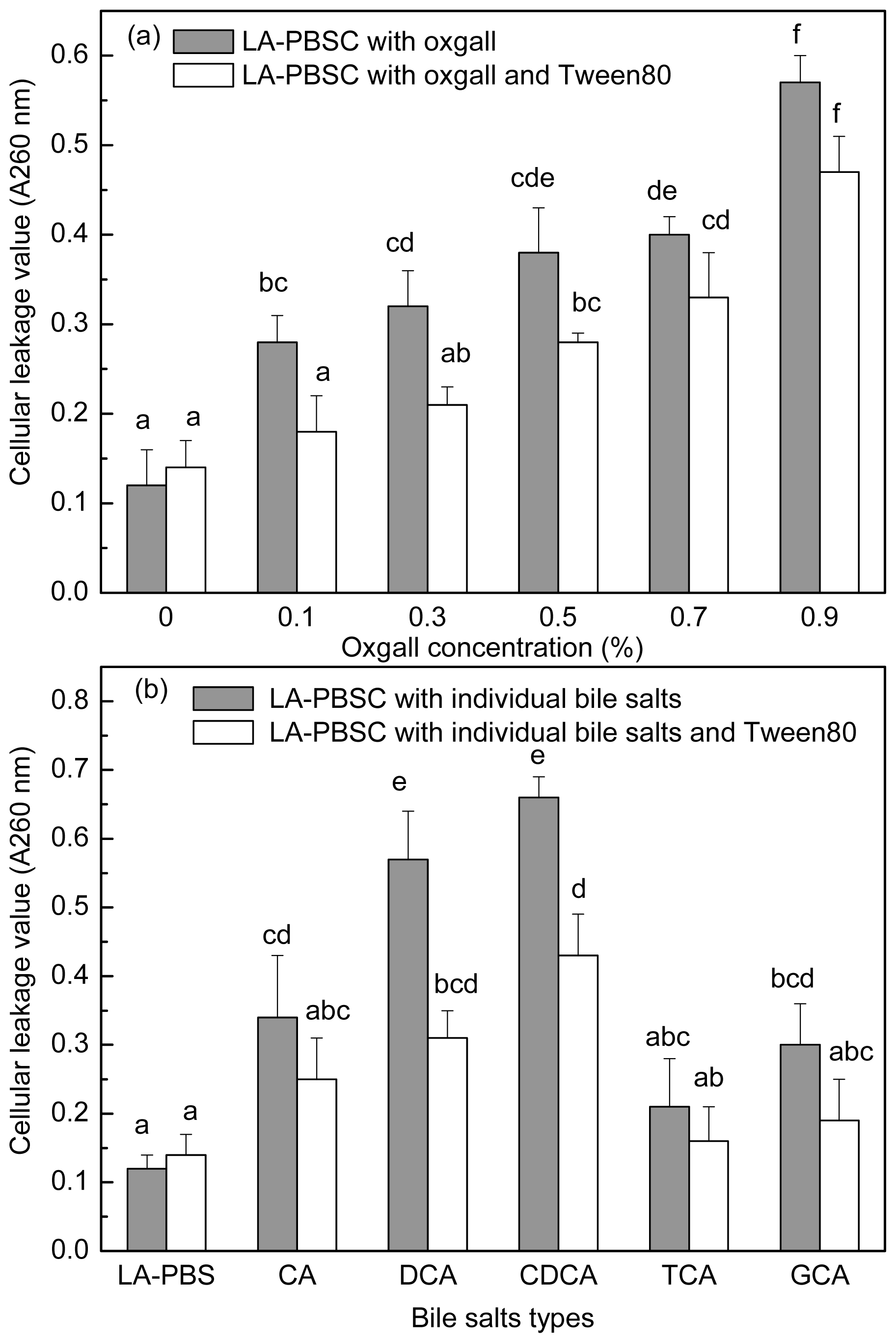
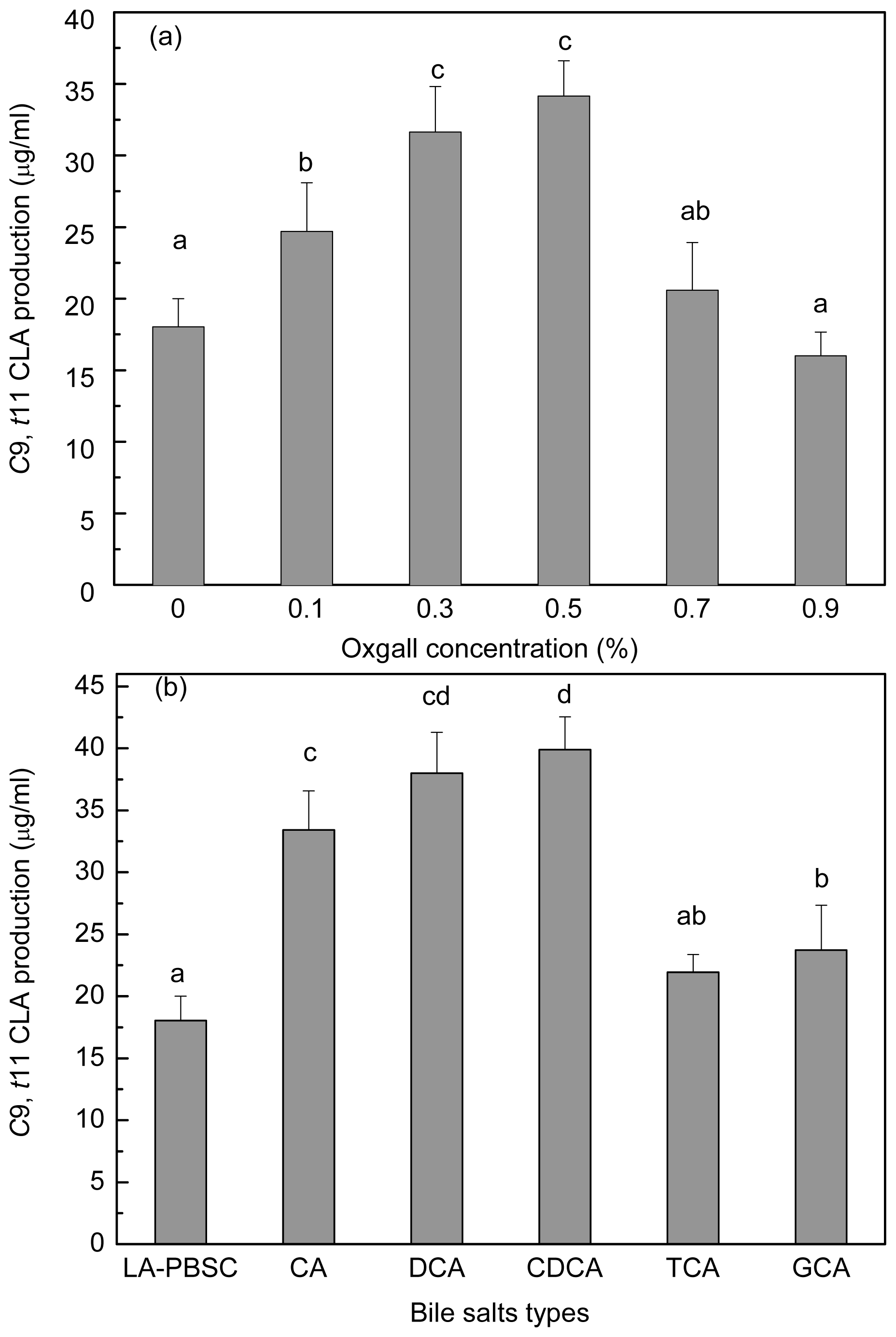
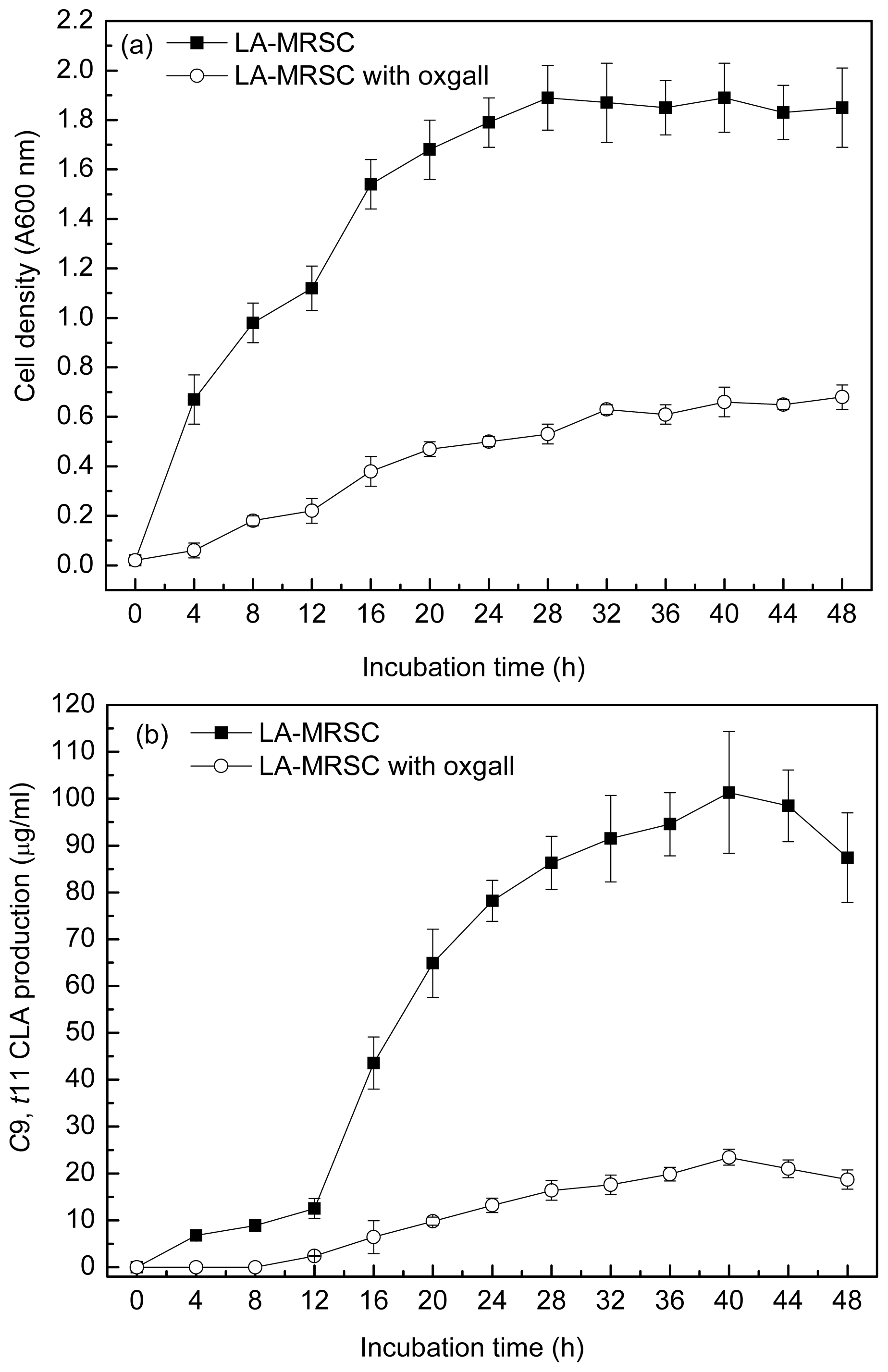
2.1. Effect of Oxgall on Growth and c9, t11 CLA Production
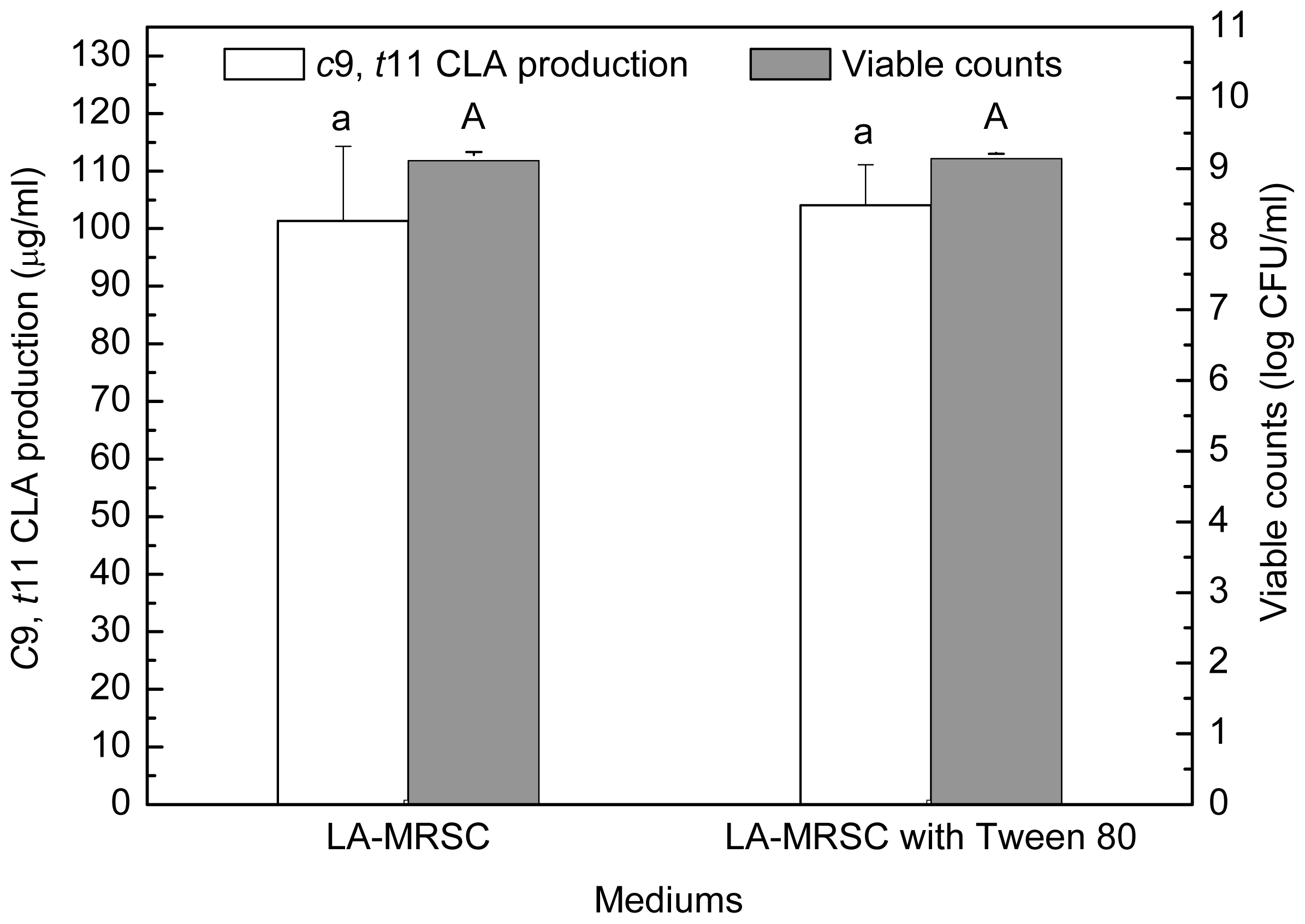
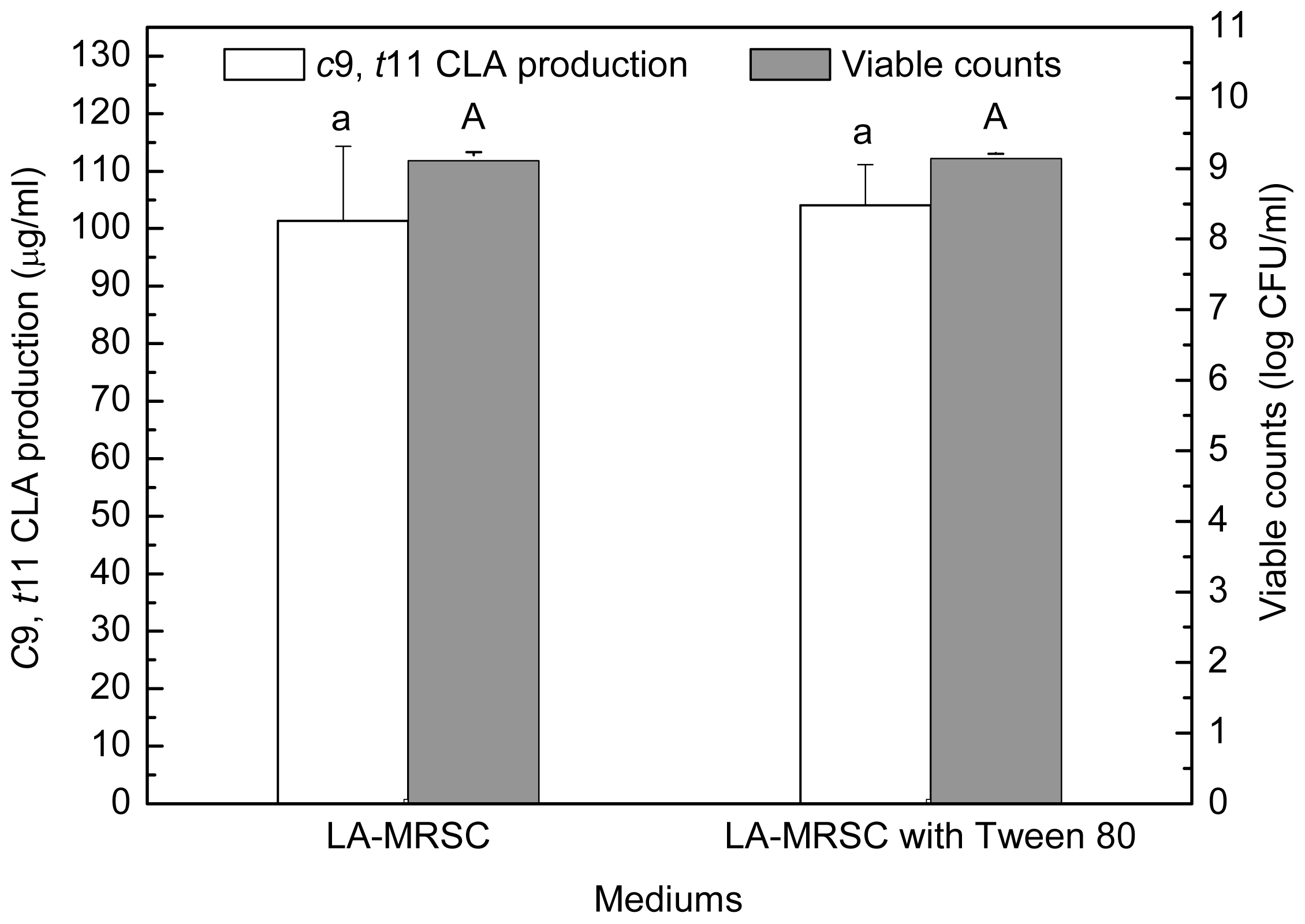
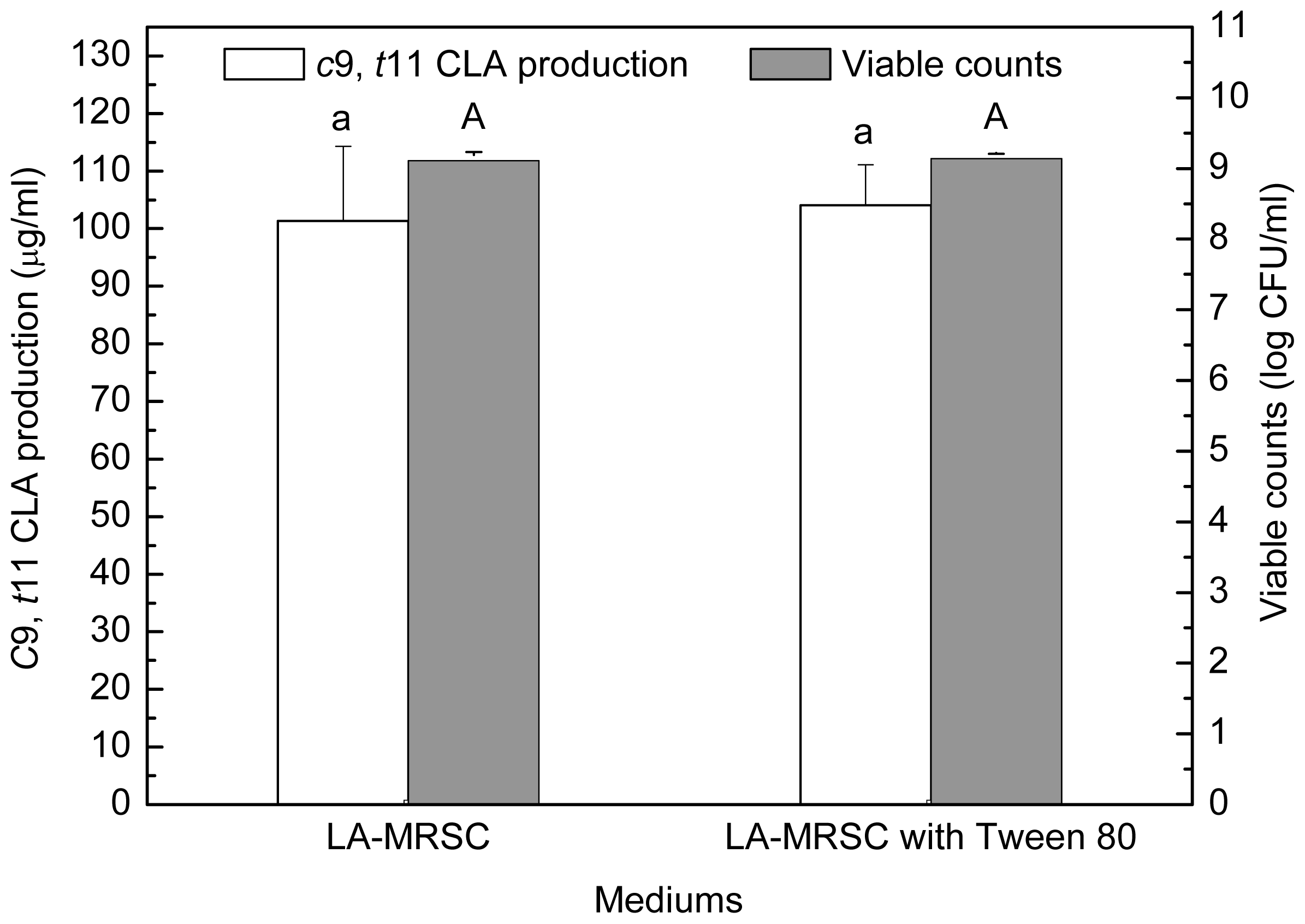
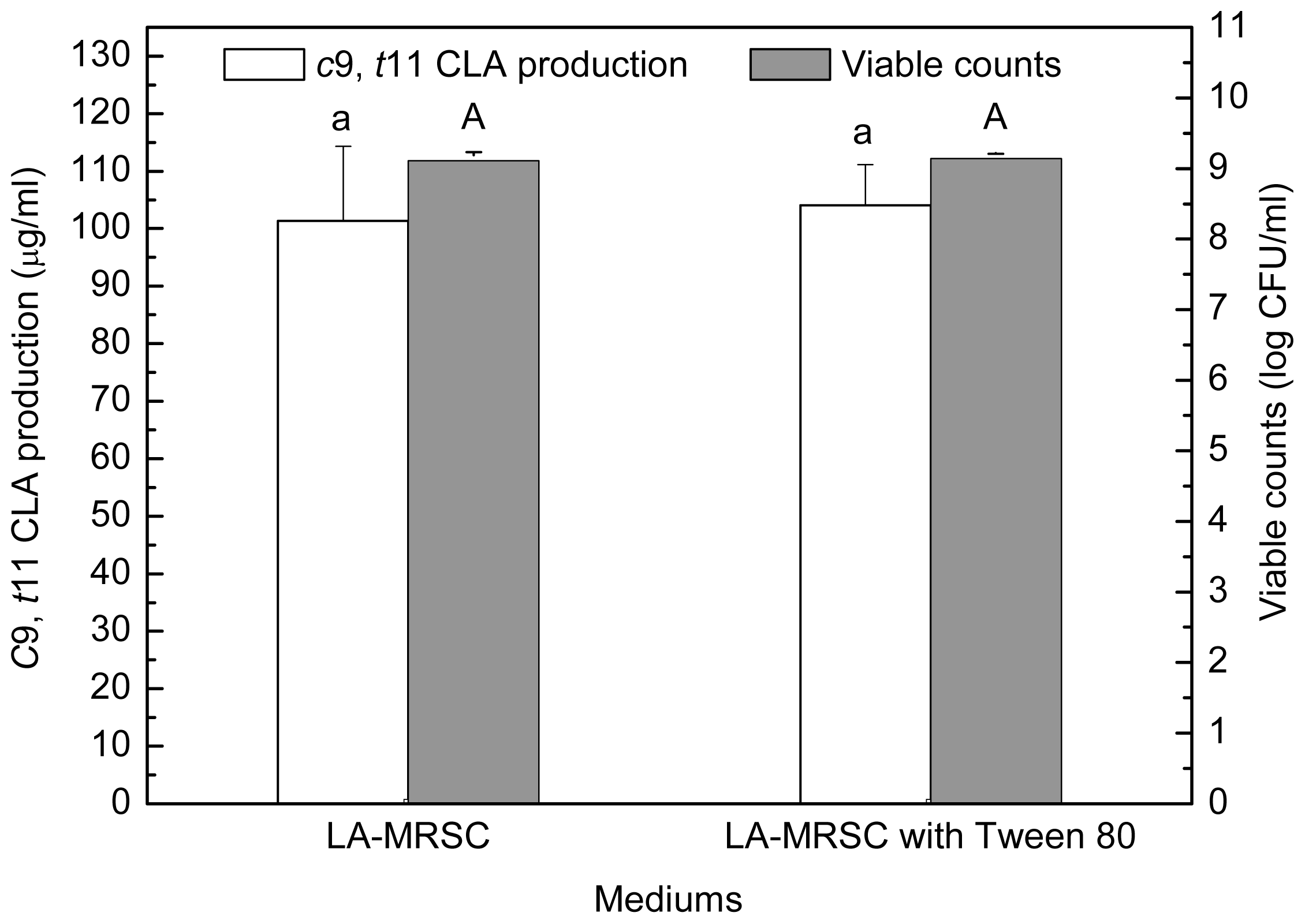
2.2. Effect of Tween Series on Growth and c9, t11 CLA Production in the Presence of Oxgall
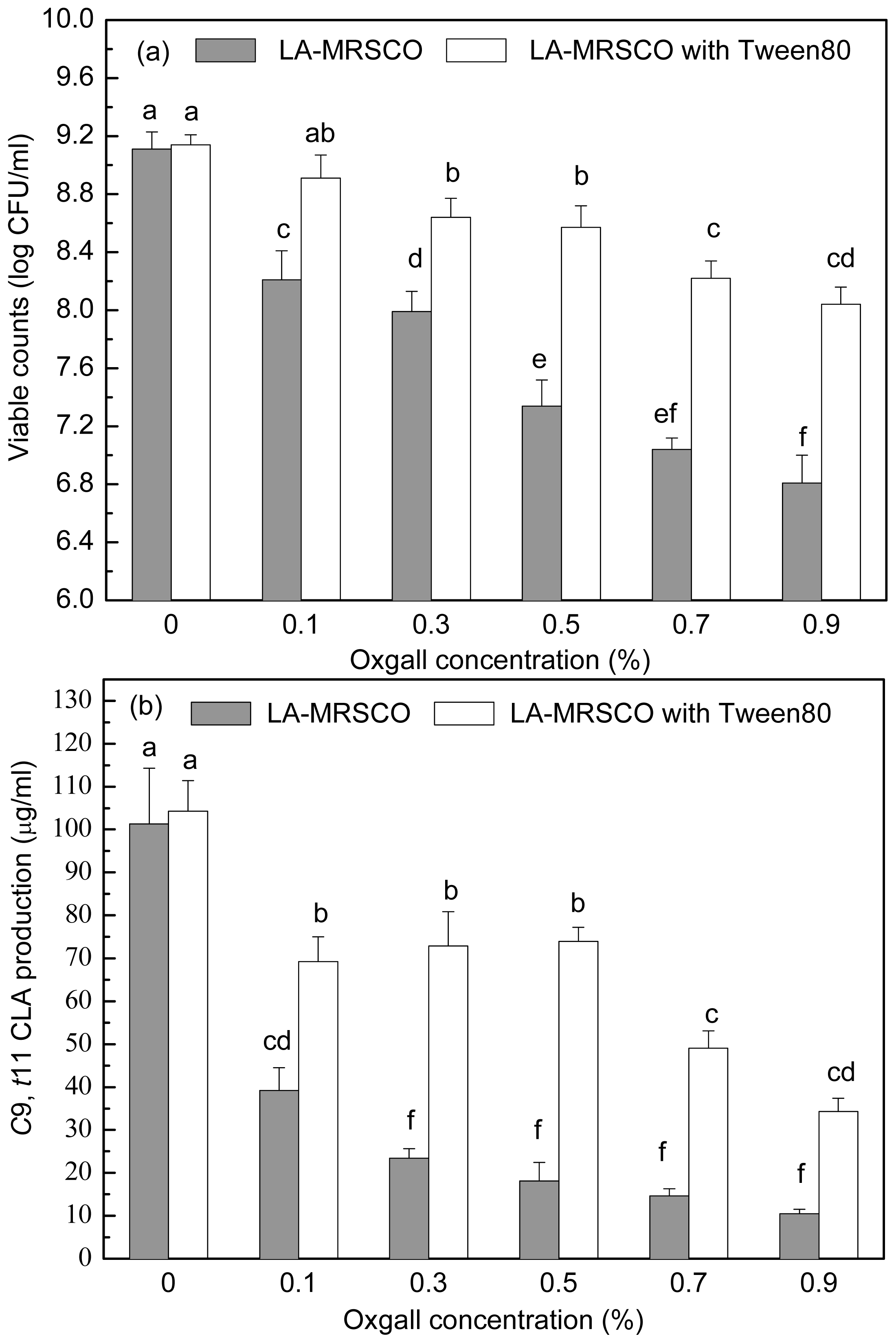
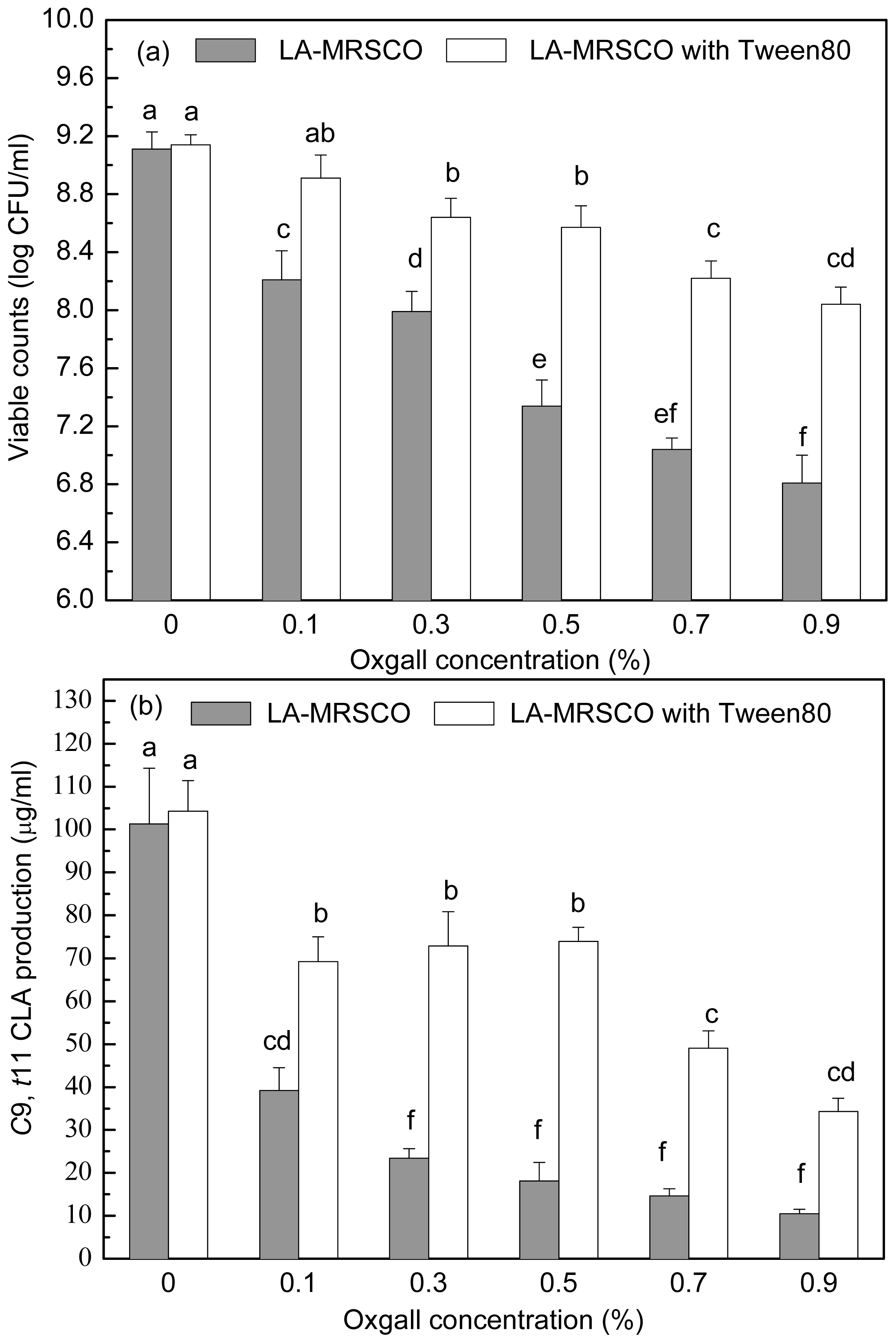
2.3. Effect of Tween 80 on Growth and c9, t11 CLA Production in the Presence of Different Concentrations of Oxgall
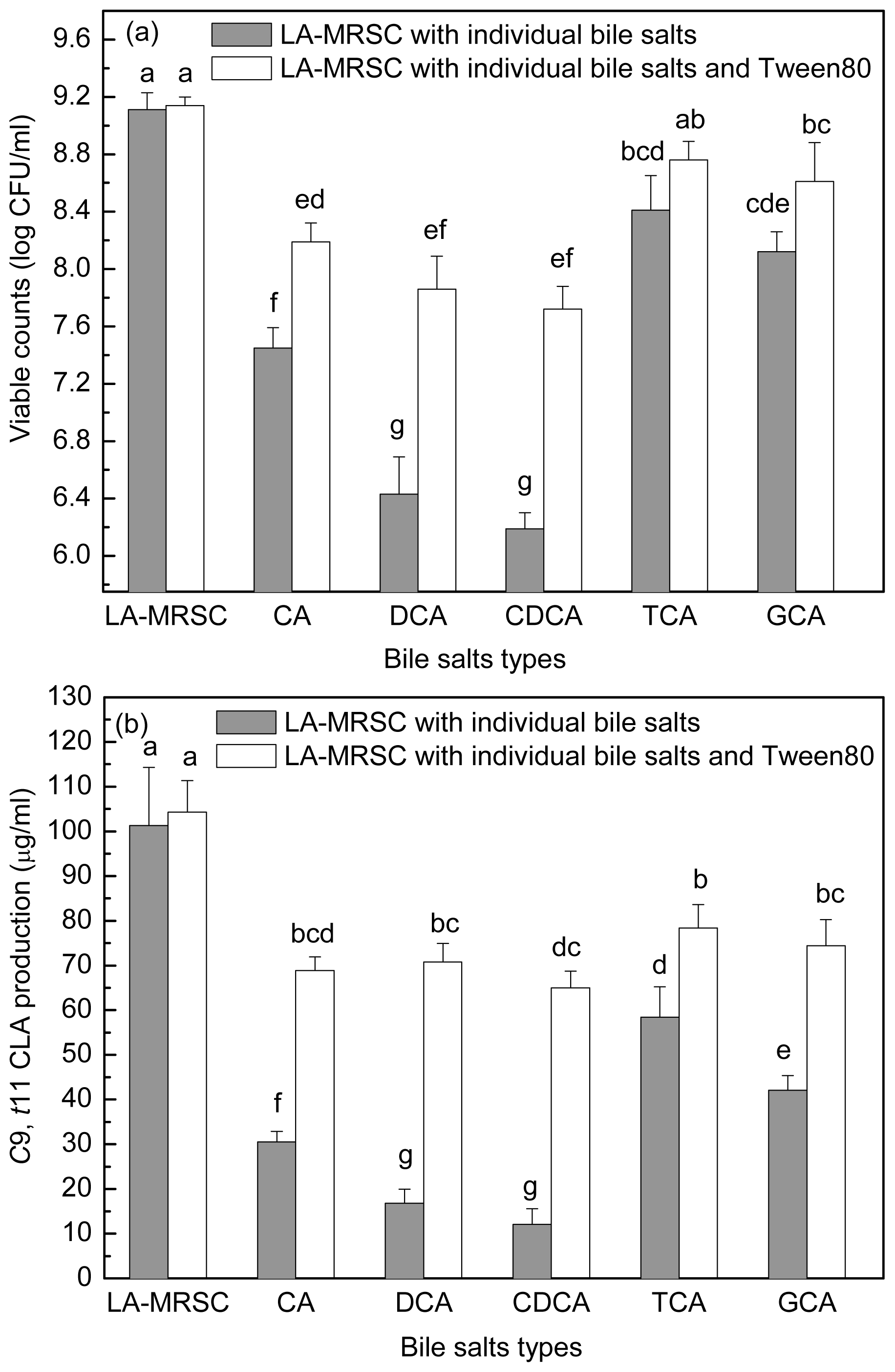
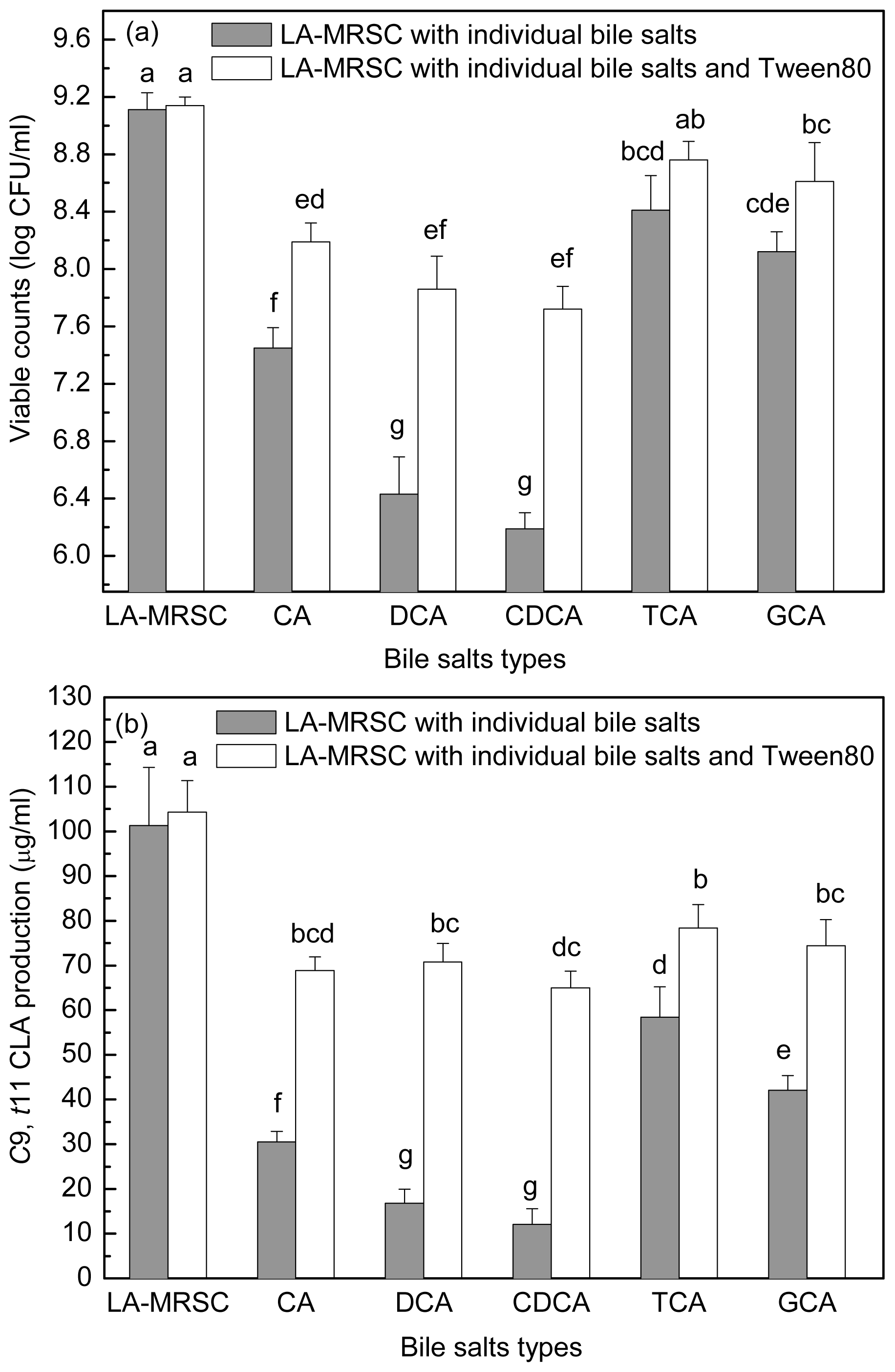
2.4. Effect of Tween 80 on Growth and c9, t11 CLA Production in the Presence of Different Types of Individual Bile Salts
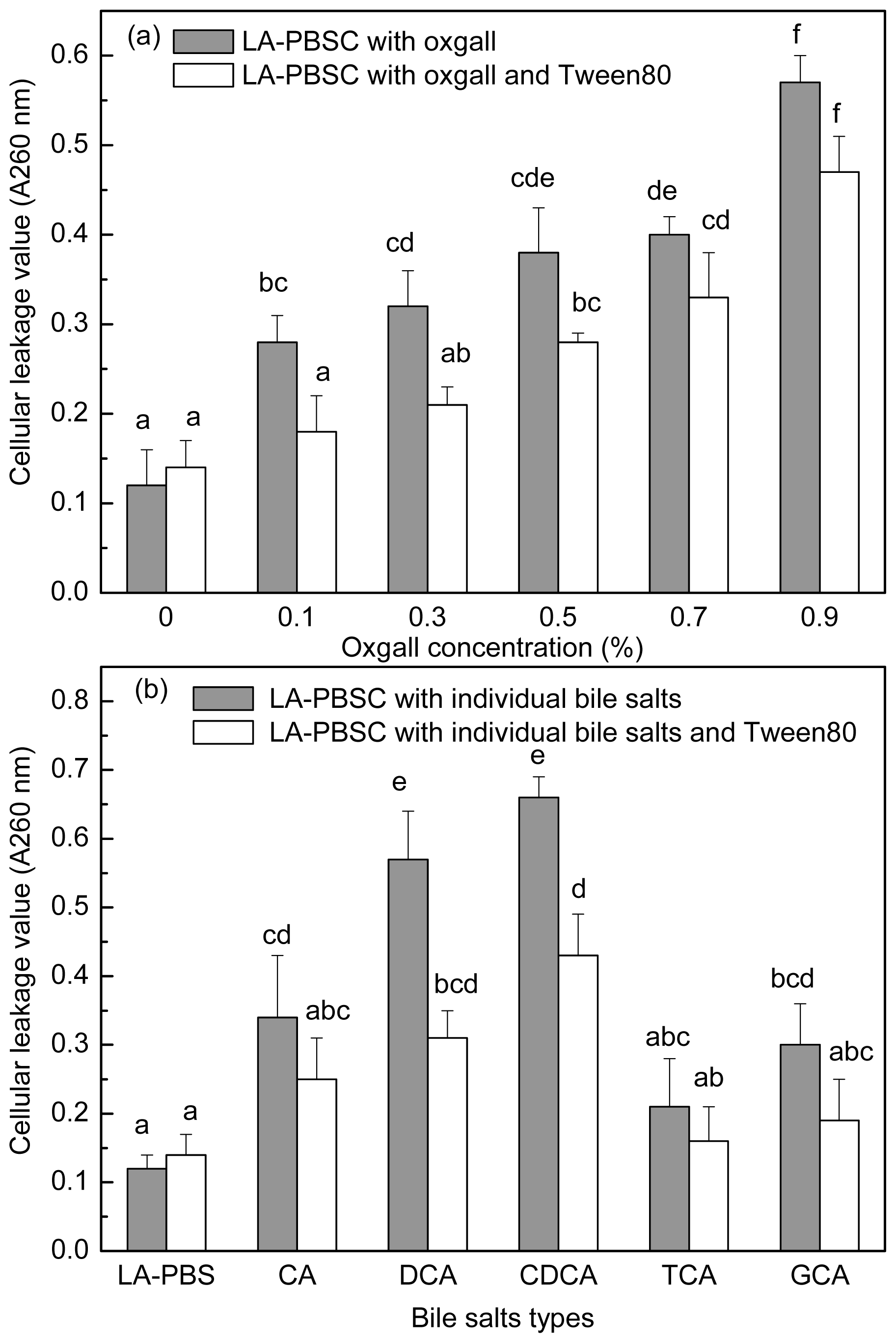
2.5. Effect of Tween 80 on Leakage of Cellular Material in the Presence of Bile Salts by Resting Cell
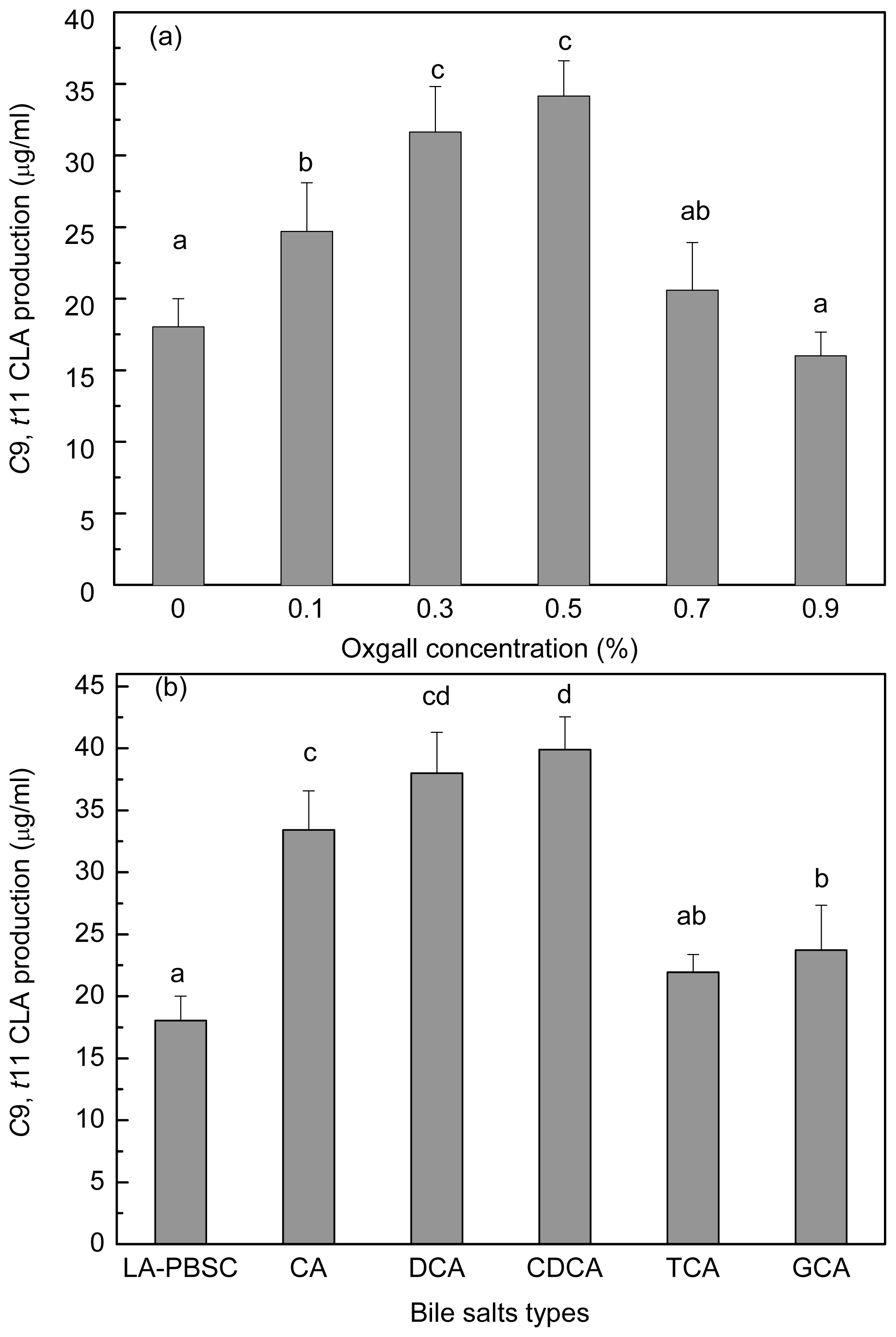
2.6. Effect of Bile Salts on c9, t11 CLA Production by Resting Cell
3. Discussion
4. Experimental Section
4.1. Strain Isolation and Maintenance
4.2. Bacteria Enumeration and Cell Density Measurement
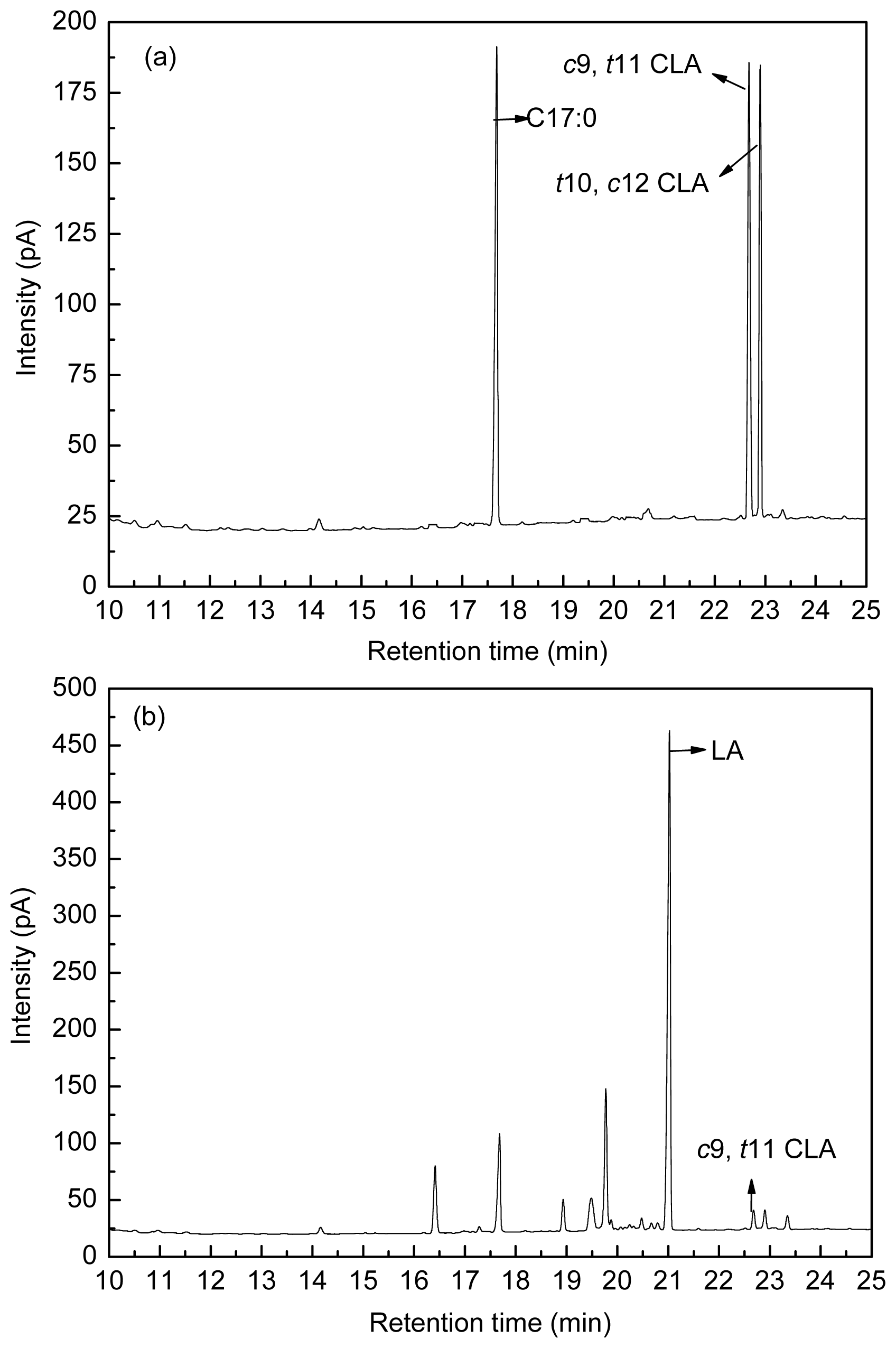
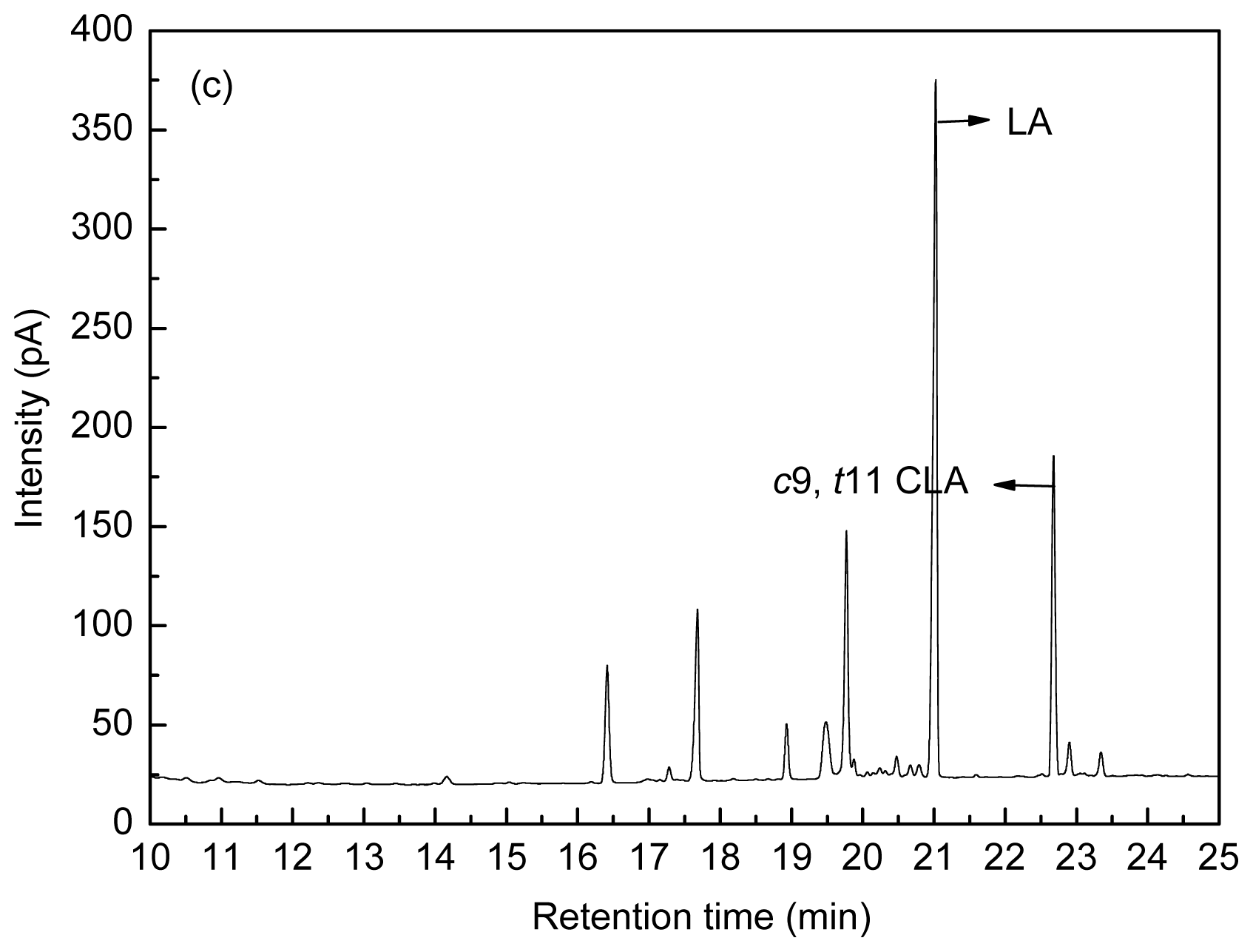
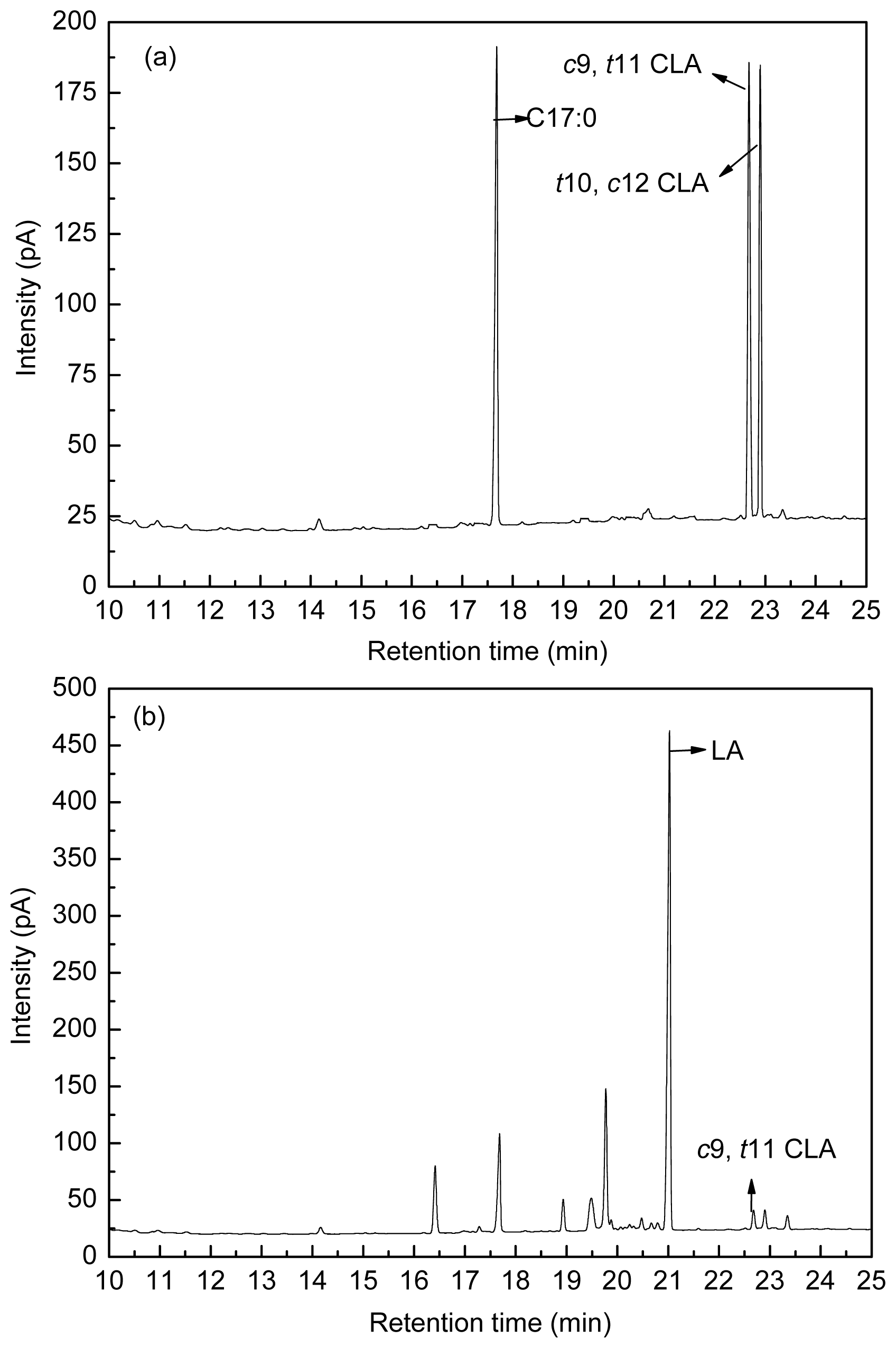
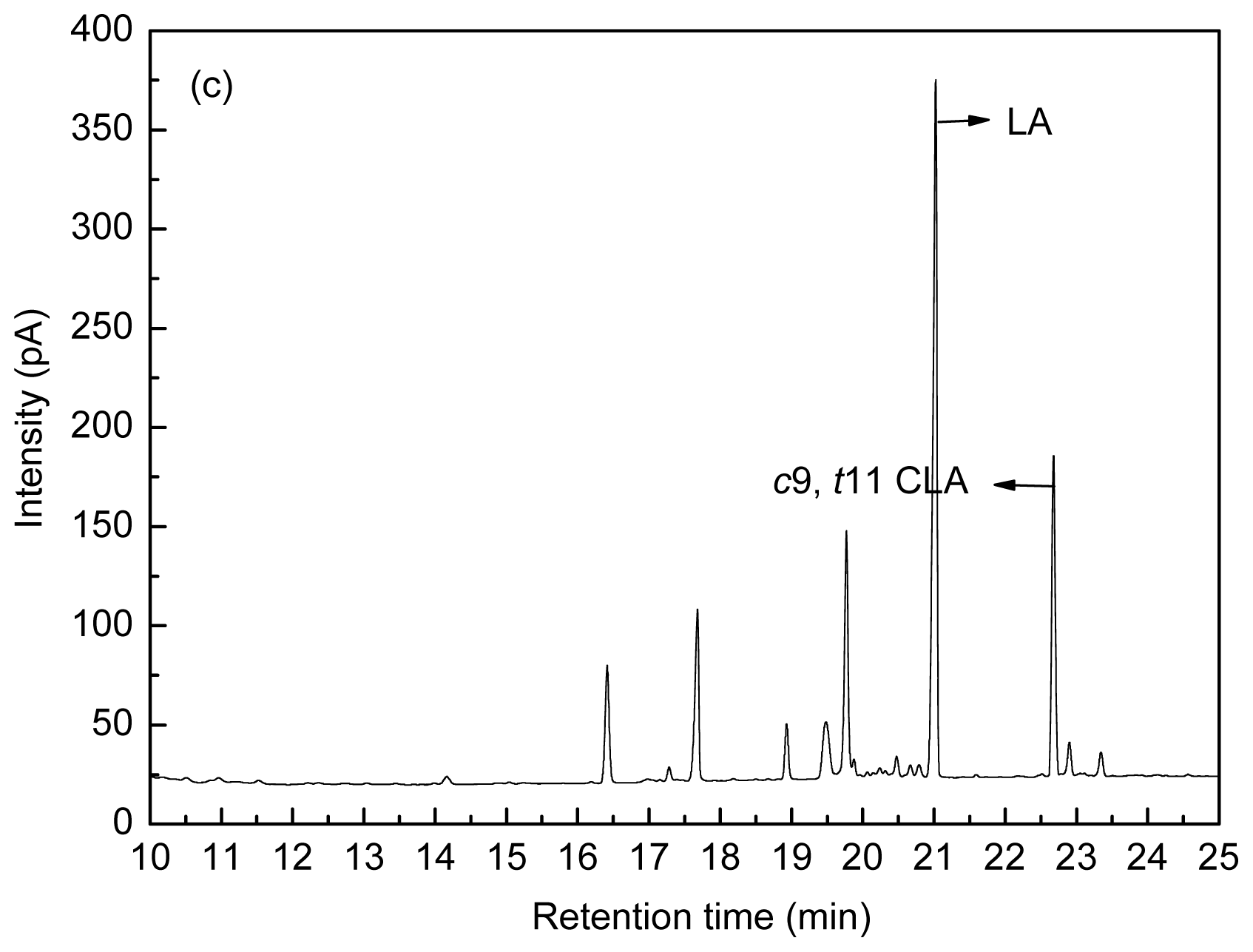
4.3. Fatty Acid Analysis
4.4. Cell Growth and c9, t11 CLA Production in the Presence of Oxgall
4.5. Effect of Tween Series on Cell Growth and c9, t11 CLA Production in the Presence of Oxgall
4.6. Effect of Tween 80 on Cell Growth and c9, t11 CLA Production in the Presence of Different Concentrations and Types of Individual Bile Salts
4.7. Effect of Tween 80 on Leakage of Intracellular Material in the Presence of Bile Salts by Resting Cell
4.8. Effect of Bile Salts on c9, t11 CLA Production by Resting Cell
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
References
- Ha, Y.L.; Grimm, N.K.; Pariza, M.W. Anticarcinogens from fried ground beef: heat-altered derivatives of linoleic acid. Carcinogenesis 1987, 8, 1881–1887. [Google Scholar]
- Ip, C.; Singh, M.; Thompson, H.J.; Scimeca, J.A. Conjugated linoleic acid suppresses mammary carcinogenesis and proliferative activity of the mammary-gland in the rat. Cancer Res 1994, 54, 1212–1215. [Google Scholar]
- Jaudszus, A.; Foerster, M.; Kroegel, C.; Wolf, I.; Jahreis, G. Cis-9, Trans-11-CLA exerts anti-inflammatory effects in human bronchial epithelial cells and eosinophils: Comparison to Trans-10, Cis-12-CLA and to linoleic acid. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2005, 1737, 111–118. [Google Scholar]
- Racine, N.M.; Watras, A.C.; Carrel, A.L.; Allen, D.B.; McVean, J.J.; Clark, R.R.; O’Brien, A.R.; O’Shea, M.; Scott, C.E.; Schoeller, D.A. Effect of conjugated linoleic acid on body fat accretion in overweight or obese children. Am. J. Clin. Nutr 2010, 91, 1157–1164. [Google Scholar]
- Smit, L.A.; Baylin, A.; Campos, H. Conjugated linoleic acid in adipose tissue and risk of myocardial infarction. Am. J. Clin. Nutr 2010, 92, 34–40. [Google Scholar]
- Ryder, J.W.; Portocarrero, C.P.; Song, X.M.; Cui, L.; Yu, M.; Combatsiaris, T.; Galuska, D.; Bauman, D.E.; Barbano, D.M.; Charron, M.J.; et al. Isomer-specific antidiabetic properties of conjugated linoleic acid. Improved glucose tolerance, skeletal muscle insulin action, and UCP-2 gene expression. Diabetes 2001, 50, 1149–1157. [Google Scholar]
- Collomb, M.; Schmid, A.; Sieber, R.; Wechsler, D.; Ryhanen, E.L. Conjugated linoleic acids in milk fat: Variation and physiological effects. Int. Dairy J 2006, 16, 1347–1361. [Google Scholar]
- McGuire, M.K.; Park, Y.; Behre, R.A.; Harrison, L.Y.; Shultz, T.D.; McGuire, M.A. Conjugated linoleic acid concentrations of human milk and infant formula. Nutr. Res 1997, 17, 1277–1283. [Google Scholar]
- Jiang, J.; Wolk, A.; Vessby, B. Relation between the intake of milk fat and the occurrence of conjugated linoleic acid in human adipose tissue. Am. J. Clin. Nutr 1999, 70, 21–27. [Google Scholar]
- Chin, S.F.; Liu, W.; Storkson, J.M.; Ha, Y.L.; Pariza, M.W. Dietary sources of conjugated dienoic isomers of linoleic acid, a newly recognized class of anticarcinogens. J. Food Compos. Anal 1992, 5, 185–197. [Google Scholar]
- Sieber, R.; Collomb, M.; Aeschlimann, A.; Jelen, P.; Eyer, H. Impact of microbial cultures on conjugated linoleic acid in dairy products a review. Int. Dairy J 2004, 14, 1–15. [Google Scholar]
- Albers, R.; van der Wielen, R.P.J.; Brink, E.J.; Hendriks, H.F.J.; Dorovska-Taran, V.N.; Mohede, I.C.M. Effects of cis-9, trans-11 and trans-10, cis-12 conjugated linoleic acid (CLA) isomers on immune function in healthy men. Eur. J. Clin. Nutr 2003, 57, 595–603. [Google Scholar]
- Berdeaux, O.; Voinot, L.; Angioni, E.; Juaneda, P.; Sebedio, J.L. A simple method of preparation of methyl trans-10, cis-12- and cis-9, trans-11-octadecadienoates from methyl linoleate. J. Am. Oil. Chem. Soc 1998, 75, 1749–1755. [Google Scholar]
- Lee, H.Y.; Park, J.H.; Seok, S.H.; Baek, M.W.; Kim, D.J.; Lee, K.E.; Paek, K.S.; Lee, Y. Human originated bacteria, Lactobacillus rhamnosus PL60, produce conjugated linoleic acid and show anti-obesity effects in diet-induced obese mice. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2006, 1761, 736–744. [Google Scholar]
- Lee, K.; Paek, K.; Lee, H.Y.; Park, J.H.; Lee, Y. Antiobesity effect of trans-10, cis-12-conjugated linoleic acid-producing Lactobacillus plantarum PL62 on diet-induced obese mice. J. Appl. Microbiol 2007, 103, 1140–1146. [Google Scholar]
- Wall, R.; Ross, R.P.; Shanahan, F.; O’Mahony, L.; O’Mahony, C.; Coakley, M.; Hart, O.; Lawlor, P.; Quigley, E.M.; Kiely, B.; et al. Metabolic activity of the enteric microbiota influences the fatty acid composition of murine and porcine liver and adipose tissues. Am. J. Clin. Nutr 2009, 89, 1393–1401. [Google Scholar]
- Begley, M.; Gahan, C.G.M.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev 2005, 29, 625–651. [Google Scholar]
- Noh, D.O.; Gilliland, S.E. Influence of bile on cellular integrity and β-galactosidase activity of Lactobacillus acidophilus. J. Dairy Sci 1993, 76, 1253–1259. [Google Scholar]
- Noriega, L.; Gueimonde, M.; Sanchez, B.; Margolles, A.; de los Reyes-Gavilan, C.G. Effect of the adaptation to high bile salts concentrations on glycosidic activity, survival at low pH and cross-resistance to bile salts in Bifidobacterium. Int. J. Food Microbiol 2004, 94, 79–86. [Google Scholar]
- Patel, H.M.; Pandiella, S.S.; Wang, R.H.; Webb, C. Influence of malt, wheat, and barley extracts on the bile tolerance of selected strains of lactobacilli. Food Microbiol 2004, 21, 83–89. [Google Scholar]
- Kimoto, H.; Ohmomo, S.; Okamoto, T. Enhancement of bile tolerance in lactococci by Tween 80. J. Appl. Microbiol. 2002, 92, 41–46. [Google Scholar]
- Lundeen, S.G.; Savage, D.C. Characterization and purification of bile salt hydrolase from Lactobacillus sp. Strain 100-100. J. Bacteriol 1990, 172, 4171–4177. [Google Scholar]
- Cheng, Y.Y.; Yang, J.P. Solubilization of non-steroidal anti-inflammatory drugs in the presence of Tween series surfactants. Phys. Chem. Liq 2006, 44, 249–256. [Google Scholar]
- Partanen, L.; Marttinen, N.; Alatossava, T. Fats and fatty acids as growth factors for Lactobacillus delbrueckii. Syst. Appl. Microbiol 2001, 24, 500–506. [Google Scholar]
- Jacques, N.A. Studies on cyclopropane fatty acid synthesis: Correlation between the state of reduction of respiratory components and the accumulation of methylene hexadecanoic acid by Pseudomonas denitrificans. Biochim. Biophys. Acta 1981, 665, 270–282. [Google Scholar]
- Kim, Y.J.; Liu, R.H.; Bond, D.R.; Russell, J.B. Effect of linoleic acid concentration on conjugated linoleic acid production by Butyrivibrio fibrisolvens A38. Appl. Environ. Microbiol. 2000, 66, 5226–5230. [Google Scholar]
- Marteau, P.; Minekus, M.; Havenaar, R.; Veld, J. Survival of lactic acid bacteria in a dynamic model of the stomach and small intestine: Validation and the effects of bile. J. Dairy Sci 1997, 80, 1031–1037. [Google Scholar]
- Noriega, L.; Cuevas, I.; Margolles, A.; de los Reyes-Gavilan, C.G. Deconjugation and bile salts hydrolase activity by Bifidobacterium strains with acquired resistance to bile. Int. Dairy J 2006, 16, 850–855. [Google Scholar]
- Kurdi, P.; Kawanishi, K.; Mizutani, K.; Yokota, A. Mechanism of growth inhibition by free bile acids in Lactobacilli and Bifidobacteria. J. Bacteriol 2006, 188, 1979–1986. [Google Scholar]
- Suskovic, J.; Kos, B.; Matosic, S.; Besendorfer, V. The effect of bile salts on survival and morphology of a potential probiotic strain Lactobacillus acidophilus M92. World J. Mircobiol. Biotechnol 2000, 16, 673–678. [Google Scholar]
- Taranto, M.P.; Murga, M.L.F.; Lorca, G.; de Valdez, G.F. Bile salts and cholesterol induce changes in the lipid cell membrane of Lactobacillus reuteri. J. Appl. Microbiol 2003, 95, 86–91. [Google Scholar]
- Fujisawa, T.; Mori, M. Influence of bile salts on β-glucuronidase activity of intestinal bacteria. Lett. Appl. Microbiol 1996, 22, 271–274. [Google Scholar]
- Noh, D.O.; Gilliland, S.E. Influence of bile on β-galactosidase activity of component species of yogurt starter cultures. J. Dairy Sci 1994, 77, 3532–3537. [Google Scholar]
- Sanchez, B.; de los Reyes-Gavilan, C.G.; Margolles, A. The F1F0-ATPase of Bifidobacterium animalis is involved in bile tolerance. Environ. Microbiol 2006, 8, 1825–1833. [Google Scholar]
- de Man, J.C.; Rogosa, M.; Sharpe, M.E. A medium for the cultivation of lactobacilli. J. Appl. Bacteriol 1960, 23, 130–135. [Google Scholar]
- Chung, S.H.; Kim, I.H.; Park, H.G.; Kang, H.S.; Yoon, C.S.; Jeong, H.Y.; Choi, N.J.; Kwon, E.G.; Kim, Y.J. Synthesis of conjugated linoleic acid by human-derived Bifidobacterium breve LMC 017: Utilization as a functional starter culture for milk fermentation. J. Agric. Food Chem 2008, 56, 3311–3316. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, J.-Y.; Zhang, L.-W.; Du, M.; Han, X.; Yi, H.-X.; Guo, C.-F.; Zhang, Y.-C.; Luo, X.; Zhang, Y.-H.; Shan, Y.-J.; et al. Effect of Tween Series on Growth and cis-9, trans-11 Conjugated Linoleic Acid Production of Lactobacillus acidophilus F0221 in the Presence of Bile Salts. Int. J. Mol. Sci. 2011, 12, 9138-9154. https://doi.org/10.3390/ijms12129138
Li J-Y, Zhang L-W, Du M, Han X, Yi H-X, Guo C-F, Zhang Y-C, Luo X, Zhang Y-H, Shan Y-J, et al. Effect of Tween Series on Growth and cis-9, trans-11 Conjugated Linoleic Acid Production of Lactobacillus acidophilus F0221 in the Presence of Bile Salts. International Journal of Molecular Sciences. 2011; 12(12):9138-9154. https://doi.org/10.3390/ijms12129138
Chicago/Turabian StyleLi, Jing-Yan, Lan-Wei Zhang, Ming Du, Xue Han, Hua-Xi Yi, Chun-Feng Guo, Ying-Chun Zhang, Xue Luo, Yan-He Zhang, Yu-Juan Shan, and et al. 2011. "Effect of Tween Series on Growth and cis-9, trans-11 Conjugated Linoleic Acid Production of Lactobacillus acidophilus F0221 in the Presence of Bile Salts" International Journal of Molecular Sciences 12, no. 12: 9138-9154. https://doi.org/10.3390/ijms12129138
APA StyleLi, J.-Y., Zhang, L.-W., Du, M., Han, X., Yi, H.-X., Guo, C.-F., Zhang, Y.-C., Luo, X., Zhang, Y.-H., Shan, Y.-J., & Hou, A.-J. (2011). Effect of Tween Series on Growth and cis-9, trans-11 Conjugated Linoleic Acid Production of Lactobacillus acidophilus F0221 in the Presence of Bile Salts. International Journal of Molecular Sciences, 12(12), 9138-9154. https://doi.org/10.3390/ijms12129138