Exposure to Sodium Fluoride Produces Signs of Apoptosis in Rat Leukocytes

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Animals
2.2. The Treatment Protocol
- Control group: Rats without any treatment that ingested food (Rat Pellet Chow; Purina, Inc., MO, USA) freely with water from containers of the liquid for human consumption (Electropura-brand drinking bottled water; The Pepsi Bottling Group México, S. de R.L. de C.V.). According to chemical analyses, this water presented a fluoride concentration of <0.5 ppm; thus, it is considered to fall within normal limits and has been reported as not interfering with the experiment.
- Experimental group 1: Rats to whose drinking water we had added a concentration of NaF equivalent to 1 ppm of fluoride and with free access to food [14].
- Experimental group 50: Rats to whose drinking water we added a concentration of NaF equivalent to 50 ppm of fluoride and with free access to food [14].
2.3. Obtaining and Processing the Leukocytes
2.4. Determination of the Metabolic Activity of Leukocytes
2.5. Total Leukocyte Extract
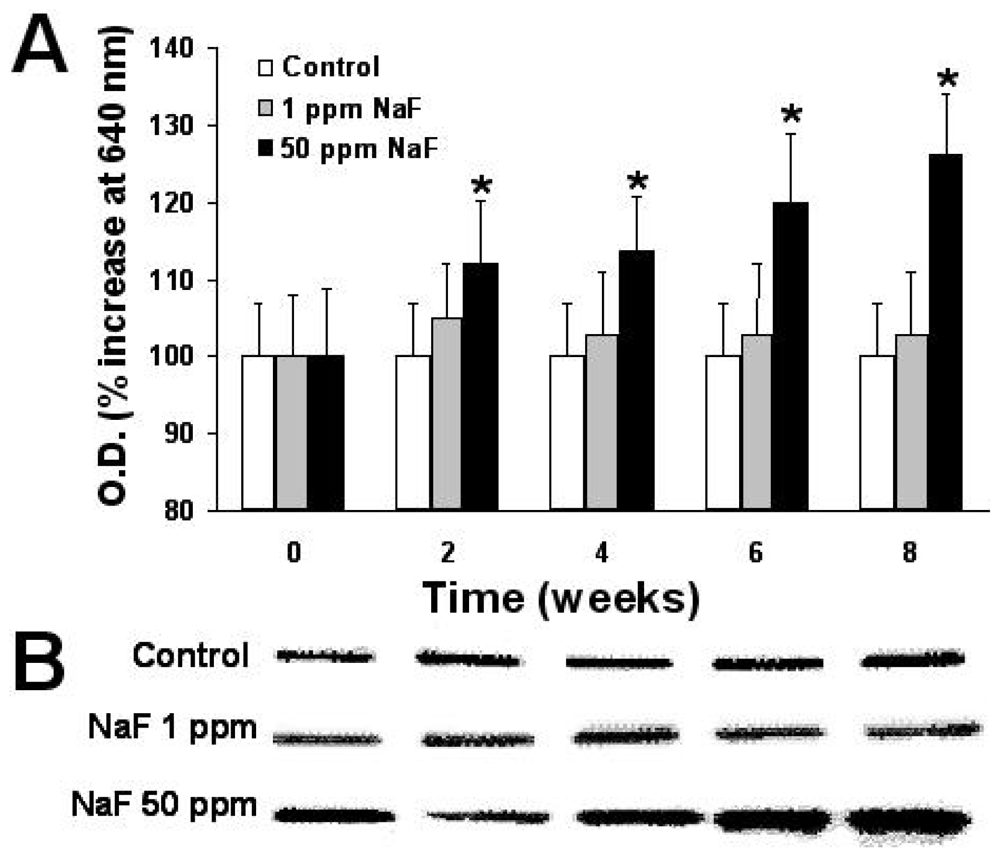
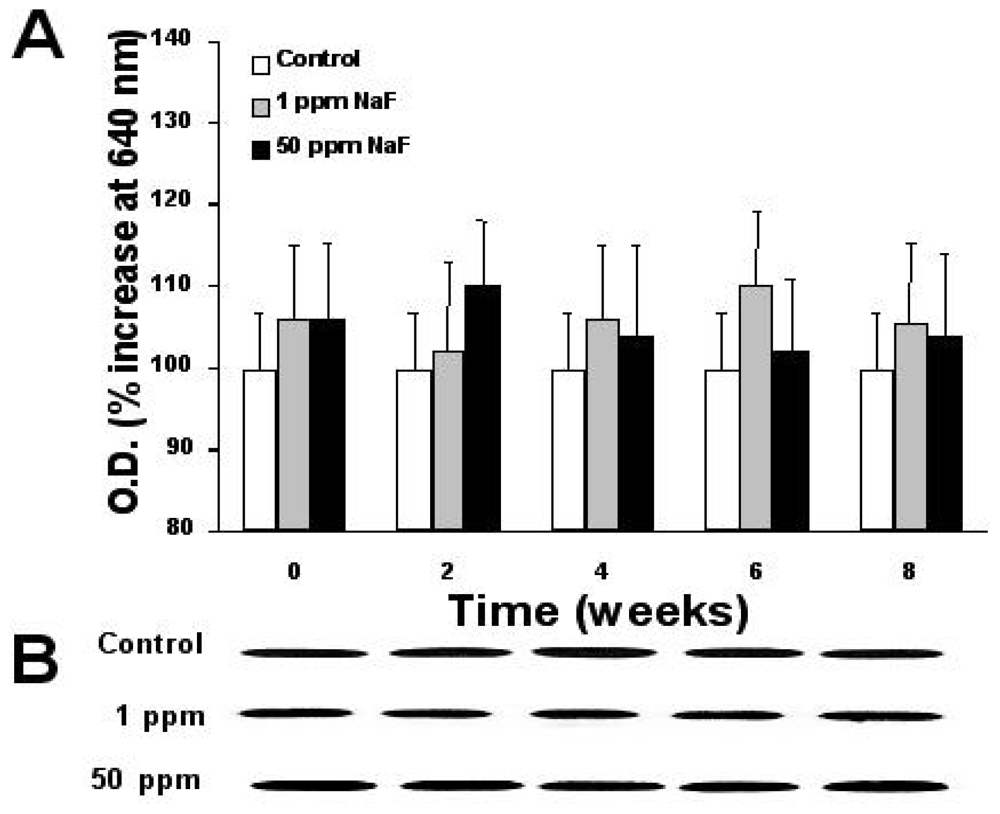
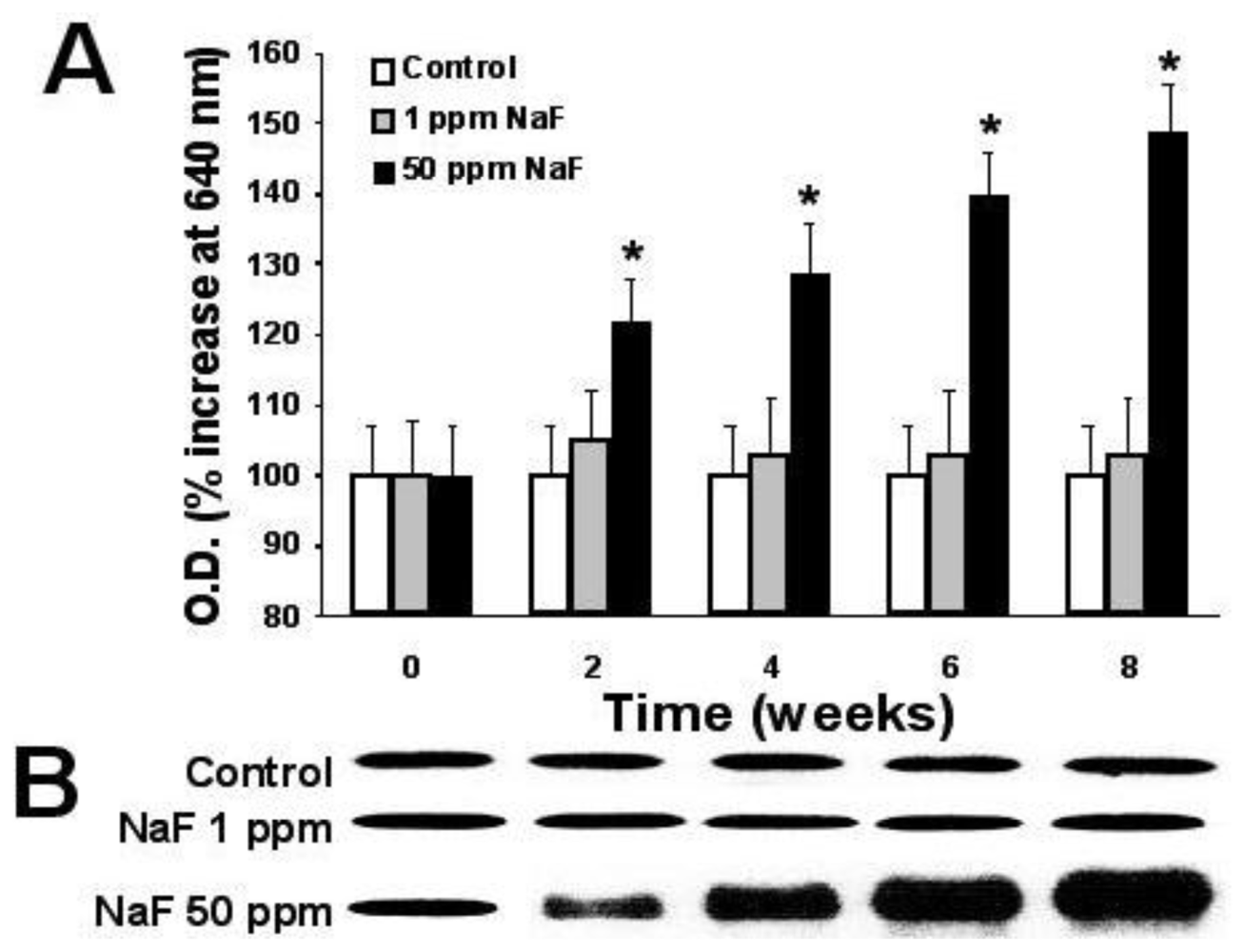
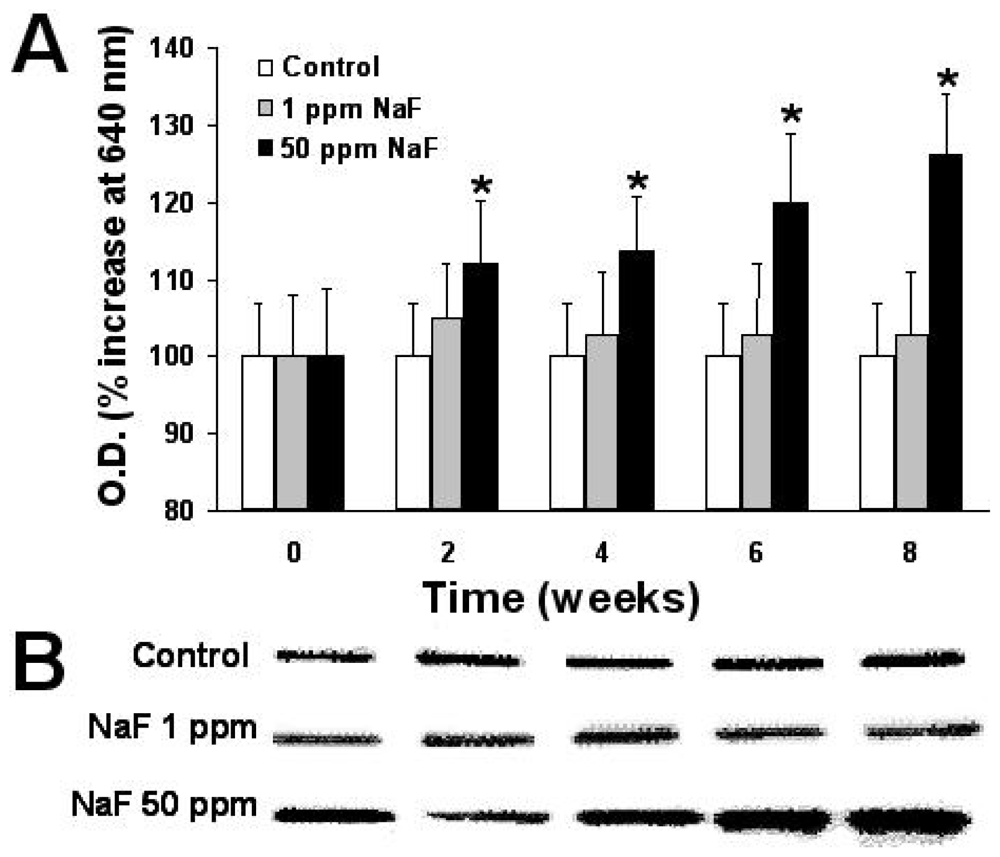
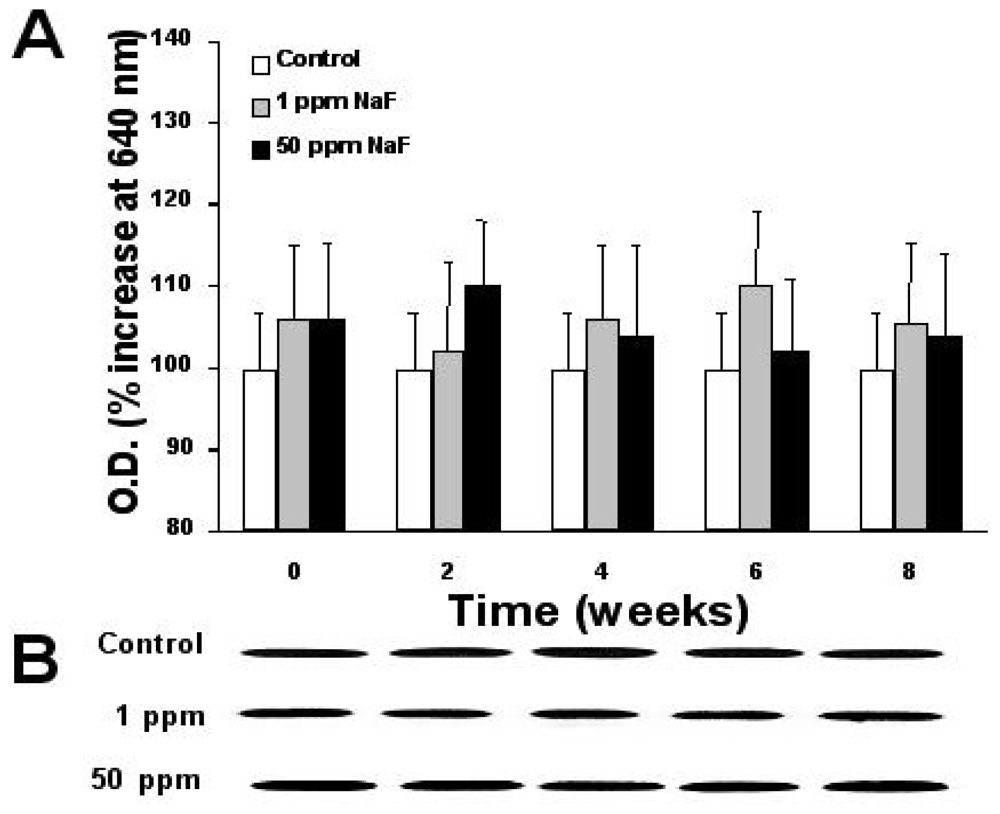
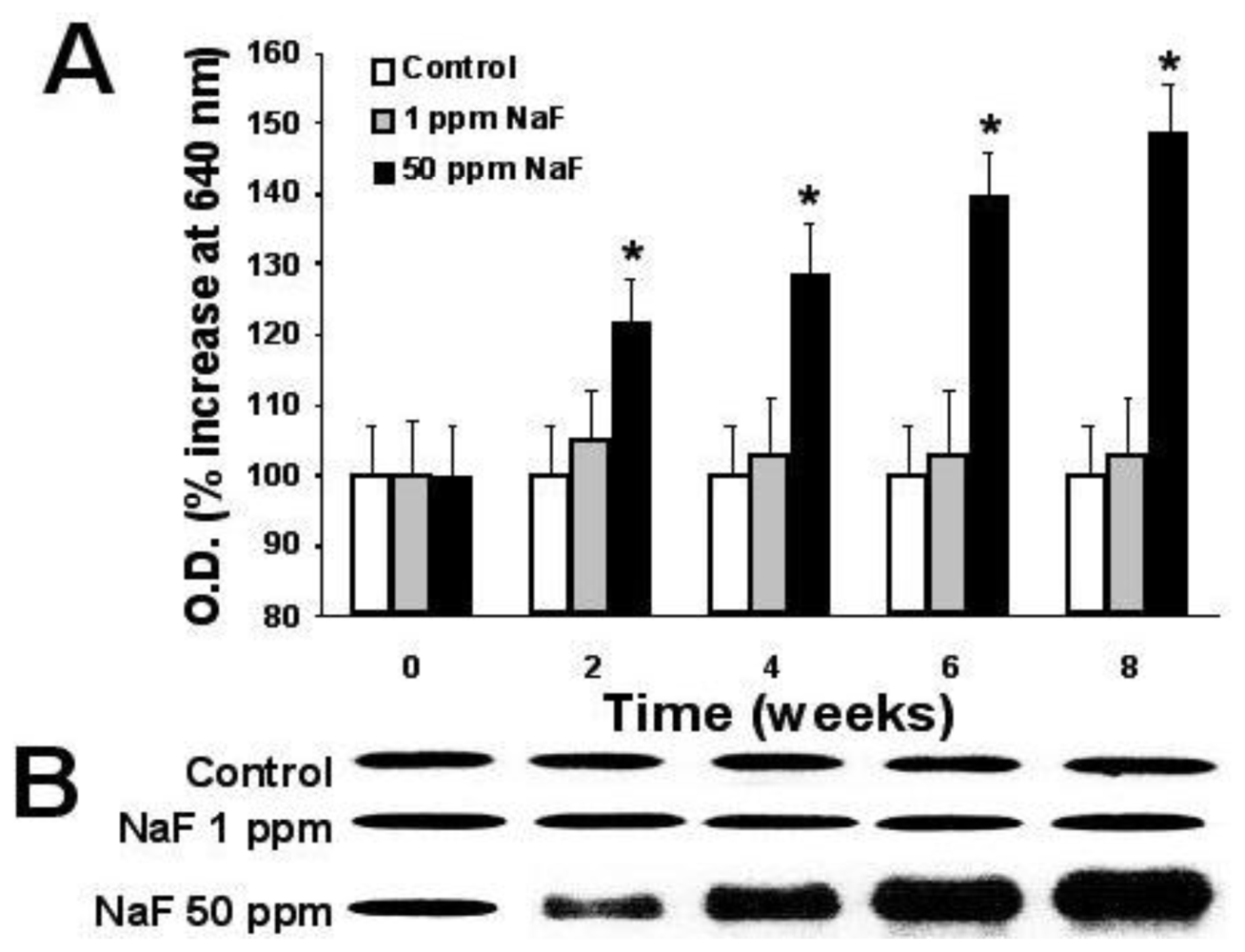
2.6. Expression of Proteins p-53, bcl-2, and Caspase-3
2.7. Statistical Analysis
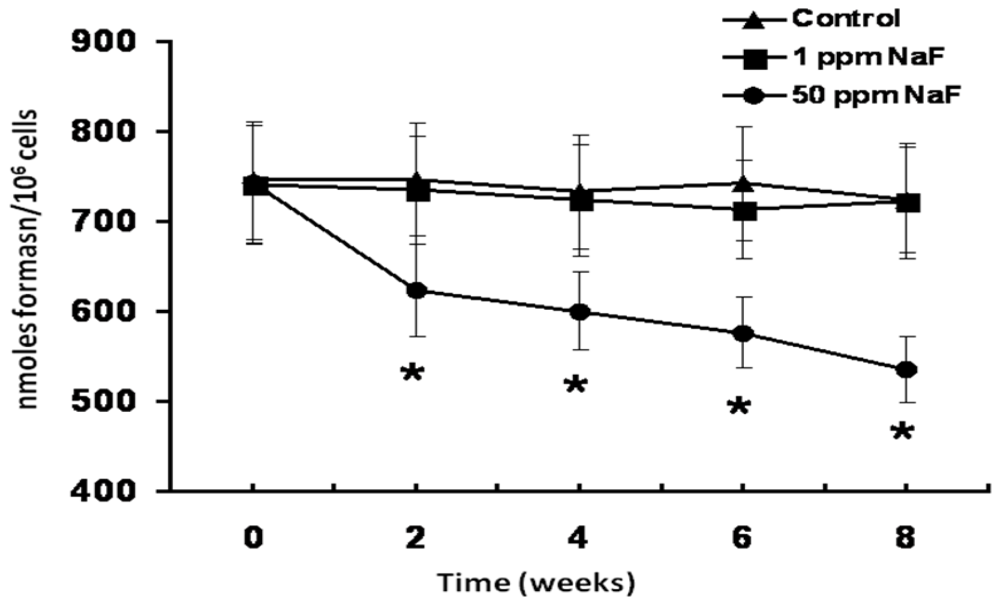
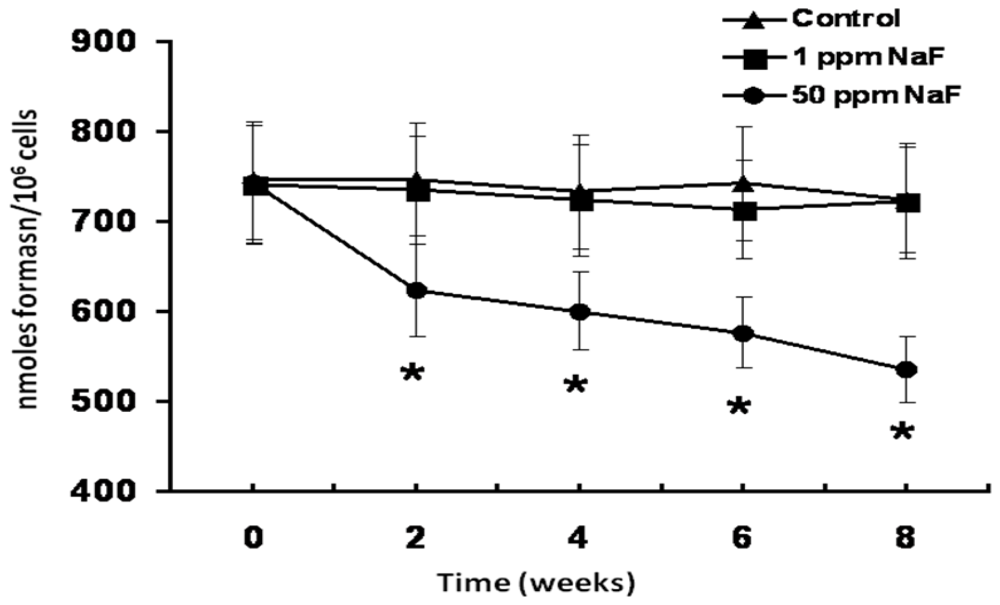
3. Results
4. Discussion
5. Conclusion
Acknowledgments
References
- Foulkes, RG. Thirty-five years of fluoride. Fluoride 2002, 35, 213–227. [Google Scholar]
- Otsuki, S; Morshed, SR; Chowdhury, SA; Takayama, F; Satoh, T; Hashimoto, K; Sugiyama, K; Amano, O; Yasui, T; Yokote, Y; Akahane, K; Sakagami, H. Posible link between glycolysis and apoptosis induced by sodium fluoride. J. Dent. Res 2005, 84, 919–923. [Google Scholar]
- Goh, EH; Neff, AW. Effects of fluoride on Xenopus embryo development. Food. Chem. Toxicol 2003, 41, 1501–1508. [Google Scholar]
- Machalinska, A; Machoy-Mokrzynska, A; Marlicz, W; Stecewicz, I; Machalinski, B. NaF-induced apoptosis in human bone marrow and cord blood CD34 positive cells. Fluoride 2001, 34, 258–263. [Google Scholar]
- Elliott, J; Scarpello, JHB; Morgan, NG. Differential effects of genistein on apoptosis induced by fluoride and pertussis toxin in human and rat pancreatic islets and RINm5F cells. J. Endocrinol 2002, 172, 137–143. [Google Scholar]
- Gutiérrez-Salinas, J; Morales-González, JA. Producción de radicales libres derivados del oxígeno y el daño al hepatocito. Med. Int. Mex 2004, 20, 287–295. [Google Scholar]
- Wang, AG; Chu, QL; He, WH; Xia, T; Liu, JL; Zhang, M; Nussler, AK; Chen, XM; Yang, KD. Effects on protein and mRNA expression levels of p53 induced by fluoride in humans embryonic hepatocytes. Toxicol. Lett 2005, 158, 158–163. [Google Scholar]
- Bogatcheva, NV; Wang, P; Birukava, AA; Verin, AD; Garcia, JGN. Mechanism of fluorideinduced MAP kinase activation in pulmonary artery endothelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol 2006, 290, L1139–L1145. [Google Scholar]
- Hidaro, S; Ando, M. Apoptotic cell death following exposure to fluoride in rat alveolar macrophages. Arch. Toxicol 1996, 70, 249–251. [Google Scholar]
- Zhan, XA; Wang, M; Xu, ZR; Li, WF; Li, JX. Evaluation of caspase-dependent apoptosis during fluoride-induced liver lesion in pigs. Arch. Toxicol 2005, 78, 145–156. [Google Scholar]
- Morales-González, JA; Bueno-Cardoso, A; Marichi-Rodriguez, F; Gutiérrez-Salinas, J. Programmed cell death (apoptosis): The regulating mechanisms of cellular proliferation. Arch. Neurocien. (Mex.) 2004, 9, 85–93. [Google Scholar]
- Darmani, H; Al-Hiyasat, AS; Elvetieha, AM. Effects of sodium fluoride in drinking water on fertility in female mice. Fluoride 2001, 34, 242–249. [Google Scholar]
- Gutiérrez-Salinas, J; Morales-González, JA. La ingesta de fluoruro de sodio produce estrés oxidativo en la mucosa bucal de la rata. Rev. Mex. Cien. Farm 2006, 37, 11–22. [Google Scholar]
- Morales-González, JA; Gutiérrez-Salinas, J; García-Ortiz, L; del Carmen Chima-Galán, M; Madrigal-Santillán, E; Esquivel-Soto, J; Esquivel-Chirino, C; García-Luna y González-Rubio, M. Effect of sodium fluoride ingestion on malondialdehyde concentration and the activity of antioxidant enzymes in rat erythrocytes. Int. J. Mol. Sci 2010, 11, 2443–2452. [Google Scholar]
- Gutiérrez-Salinas, J; Cruz-Tovar, L; García-Méndez, S. Incremento en la concentración de óxido nitrico y meta-hemoglobina en eritrocitos contenidos en bolsas para transfusión sanguínea. Rev. Mex. Patol. Clin 2008, 55, 21–28. [Google Scholar]
- Gutiérrez-Salinas, J; Zentella de Piña, M; Piña, E. Acute ethanol intake produces lipid peroxidation in rat red blood cells membranes. Biochem. Mol. Biol. Int 1993, 29, 263–270. [Google Scholar]
- Russo-Carbolante, EMS; Azzolini, AECS; Polizello, ACM; Lucisano-Valim, YM. Comparative study of four isolation procedures to obtain rat neutrophils. Compar. Clin. Pathol 2002, 11, 71–76. [Google Scholar]
- Lokaj, V; Oburkova, PS. Tetrazolium-reductase activity in leucocytes. Immunol. Zprav 1975, 6, 42–44. [Google Scholar]
- Lowry, OH; Rosebrough, NJ; Farr, AI; Randal, RJ. Protein measurement with the folin phenol reagent. J. Biol. Chem 1951, 1, 265–275. [Google Scholar]
- Laemmli, VK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar]
- Towbin, H; Gordon, J. Immunoblotting and dot immunobindig. Current status and out-look. J. Immunol. Methods 1984, 72, 313–340. [Google Scholar]
- Gutiérrez-Salinas, J; Morales-González, JA; Vargas-Castro, LD; Marichi-Rodríguez, F; Esquivel-Soto, J. Changes in laminin expresion in periodontal ligament from theet subjet to orthodontic pressure in humans. Rev. ADM 2005, 62, 171–176. [Google Scholar]
- Denbesten, PK. Effects of fluoride on protein secretion and removal during enamel development in the rat. J. Dent. Res 1986, 65, 1272–1277. [Google Scholar]
- Denbesten, PK. Mechanism and timing of fluoride effects on developing enamel. J. Pub. Health. Dent 1999, 59, 247–251. [Google Scholar]
- Heindel, JJ; Bates, HK; Price, CJ; Marr, MC; Myers, CB; Schwetz, BA. Developmental toxicity evaluation of sodium fluoride administered to rats and rabbits in drinking water. Fundam. Appl. Toxicol 1996, 30, 162–177. [Google Scholar]
- Reumaux, D; Boer, M; Meijer, AB; Duthilleul, P; Roos, D. Expression of myeloperoxidase (MPO) by neutrophils is necessary for their activation by anti-neutrophil coytoplasm autoantibodies (ANCA) against MPO. J. Leukoc. Biol 2003, 73, 841–849. [Google Scholar]
- Lowe, SW; Ruley, HE; Jacks, T; Housman, DE. p53 dependent apoptosis modulates the cytotoxicity of anticancer agents. Cell 1993, 74, 957–967. [Google Scholar]
- Lane, DP. p53, guardian of the genome. Nature 1984, 77, 1087–1090. [Google Scholar]
- Gutiérrez-Salinas, J. Morales González, JA, Ed.; Daño al hígado por radicales libres derivados del oxígeno. In Alcohol, Alcoholismo y Cirrosis: Un Enfoque Multisdiciplinario, 1a ed; Universidad Autónoma del Estado de Hidalgo: Pachuca Hgo, México, 2007; pp. 97–109. [Google Scholar]
- Reed, JC. Double identity for proteins off the Bcl-2 family. Nature 1997, 387, 773–776. [Google Scholar]
- Siganaki, M; Koutsopoulos, AV; Neofytou, E; Vlachaki, E; Maria Psarrou, M; Soulitzis, N; Pentilas, N; Schiza, S; Siafakas, NM; Tzortzaki, EG. Deregulation of apoptosis mediators’ p53 and bcl2 in lung tissue of COPD patients. Resp. Res 2010, 11, 46. [Google Scholar]
- Parck, JR; Hockenbery, DM. Bcl-2: A novel regulator of apoptosis. J. Cell. Biochem 1996, 60, 12–17. [Google Scholar]
- Machalinski, B; Baskiewicz-Masiuk, M; Sadowska, B; Machal inska, A; Marchlewicz, M; Wiszniewsda, B; Stecewicz, I. The influence of sodium fluoride and sodium hexafluorosilicate on human leukemic cell lines. Fluoride 2003, 36, 231–240. [Google Scholar]
- Guan, ZZ; Yu, PS; Yang, ND; Zhang, ZJ. An experimental study of blood biochemical diagnostic indices for chronic fluorosis. Fluoride 1989, 22, 112–118. [Google Scholar]
- Bratton, DL; Henson, PM. Autoimmunity and apoptosis: Refusing to go quietly. Nat. Med 2005, 11, 26–27. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (n) | AIW (g) | AFW (g) | WCR (mL/day) | D-NaF (mg/kg/week) |
|---|---|---|---|---|
| Control (25) | 252.13 ± 3.14 | 314.13 ± 5.39 | 29.05 ± 0.30 | 0 |
| NaF 1ppm (25) | 252.70 ± 3.32 | 312.23 ± 3.32 | 31.05 ± 0.32 | 0.868 ± 0.01 |
| NaF 50 ppm (25) | 251.65 ± 4.81 | 312.12 ± 5.21 | 30.48 ± 0.27 | 42.67 ± 0.38 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gutiérrez-Salinas, J.; Morales-González, J.A.; Madrigal-Santillán, E.; Esquivel-Soto, J.; Esquivel-Chirino, C.; González-Rubio, M.G.-L.y.; Suástegui-Domínguez, S.; Valadez-Vega, C. Exposure to Sodium Fluoride Produces Signs of Apoptosis in Rat Leukocytes. Int. J. Mol. Sci. 2010, 11, 3610-3622. https://doi.org/10.3390/ijms11093610
Gutiérrez-Salinas J, Morales-González JA, Madrigal-Santillán E, Esquivel-Soto J, Esquivel-Chirino C, González-Rubio MG-Ly, Suástegui-Domínguez S, Valadez-Vega C. Exposure to Sodium Fluoride Produces Signs of Apoptosis in Rat Leukocytes. International Journal of Molecular Sciences. 2010; 11(9):3610-3622. https://doi.org/10.3390/ijms11093610
Chicago/Turabian StyleGutiérrez-Salinas, José, José A. Morales-González, Eduardo Madrigal-Santillán, Jaime Esquivel-Soto, César Esquivel-Chirino, Manuel García-Luna y González-Rubio, Sigrit Suástegui-Domínguez, and Carmen Valadez-Vega. 2010. "Exposure to Sodium Fluoride Produces Signs of Apoptosis in Rat Leukocytes" International Journal of Molecular Sciences 11, no. 9: 3610-3622. https://doi.org/10.3390/ijms11093610
APA StyleGutiérrez-Salinas, J., Morales-González, J. A., Madrigal-Santillán, E., Esquivel-Soto, J., Esquivel-Chirino, C., González-Rubio, M. G.-L. y., Suástegui-Domínguez, S., & Valadez-Vega, C. (2010). Exposure to Sodium Fluoride Produces Signs of Apoptosis in Rat Leukocytes. International Journal of Molecular Sciences, 11(9), 3610-3622. https://doi.org/10.3390/ijms11093610