Anti-Allergic Activity of a Platycodon Root Ethanol Extract

Abstract
:1. Introduction
2. Results and Discussion
2.1. The Effects of PE on BMMC Viability
2.2. The Effects of PE on PMA and A23187-Induced IL-6 Secretion
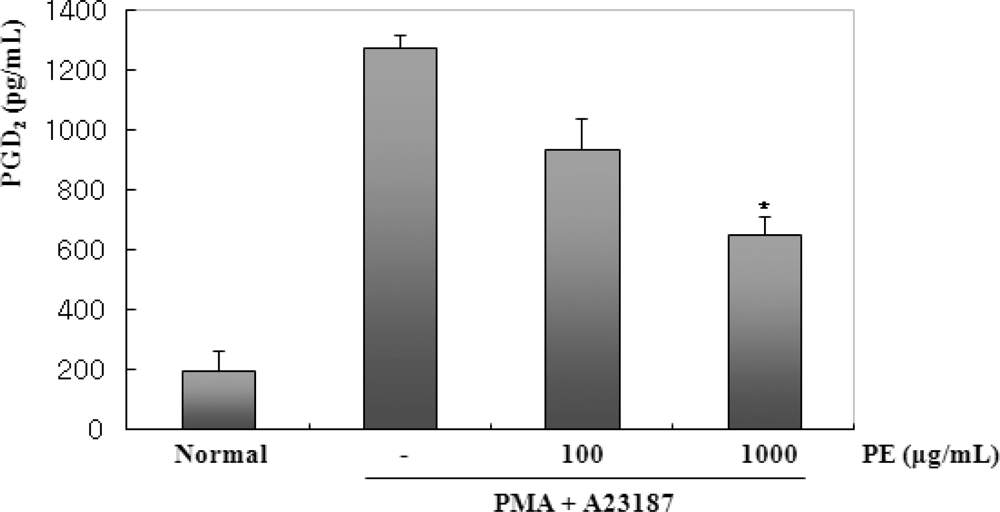
2.3. The Effects of PE on PMA and A23187-Induced PGD2 Production
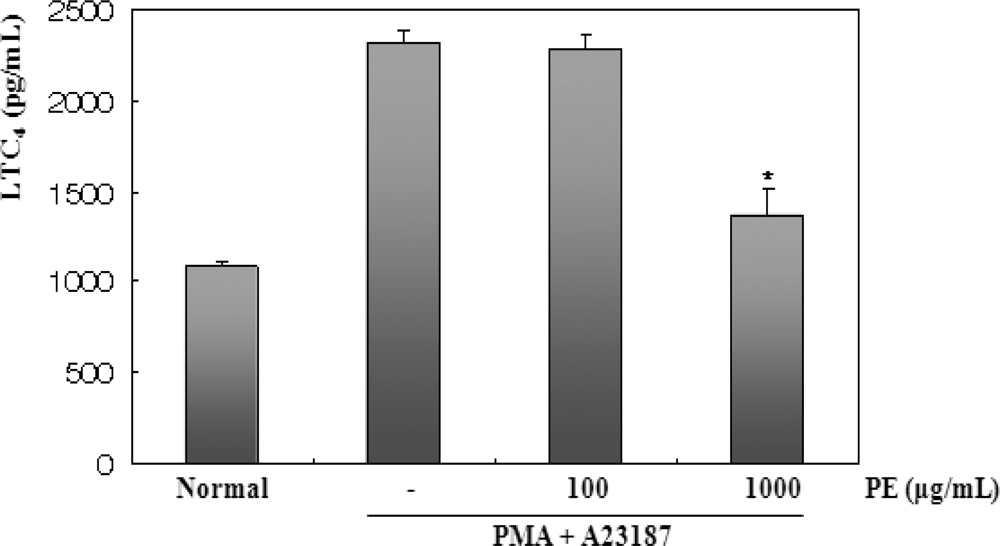
2.4. The effects of PE on PMA and A23187-Induced LTC4 Production
2.5. The Effects of PE on PMA and A23187-Induced COX-2 Protein Expression
2.6. The Inhibitory Effects of PE on PMA and A23187-Induced Degranulation
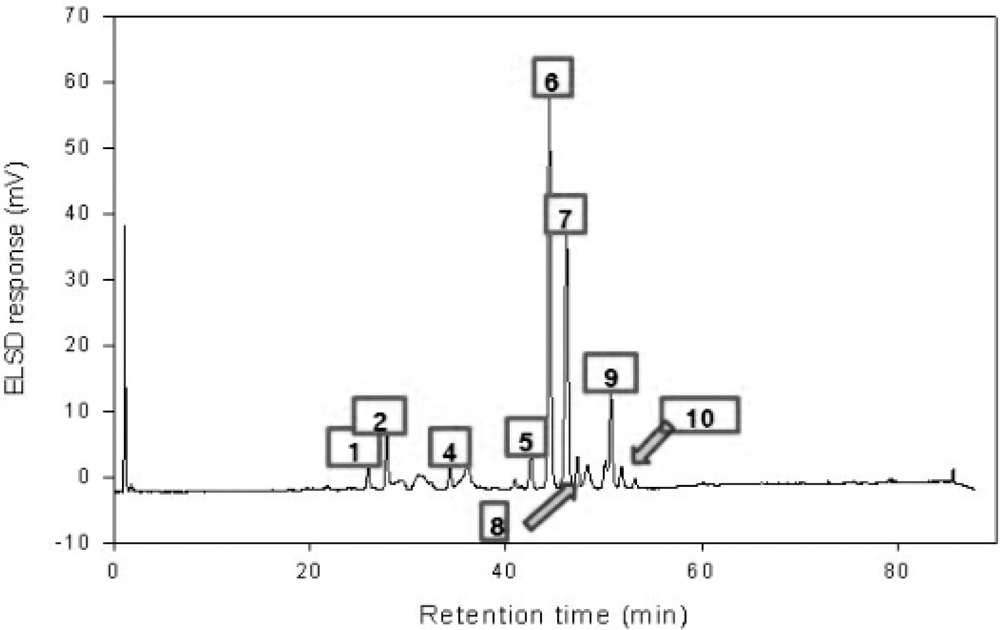
2.7. High Performance Liquid Chromatography (HPLC)
3. Experimental Section
3.1. Plant Materials
3.2. Animals
3.3. Drugs and Chemicals
3.4. Preparation and Activation of Bone Marrow-Derived Mast Cells (BMMC)
3.5. MTS Assay for Cell Viability
3.6. Enzyme-Linked Immunosorbent Assay (ELISA)
3.7. Determination of Prostaglandin D2 (PGD2) Levels
3.8. Determination of Leukotriene C4 (LTC4) Levels
3.9. Western Blot Analysis
3.10. Assay of β-Hexosaminidase Release
3.11. Chromatography
3.12. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Choi, JH; Hwang, YP; Lee, HS; Jeong, HG. Inhibitory effect of Platycodi Radix on ovalbumin-induced airway inflammation in a murine model of asthma. Food Chem. Toxicol 2009, 47, 1272–1279. [Google Scholar]
- Ahn, KS; Noh, EJ; Zhao, HL; Jung, SH; Kang, SS; Kim, YS. Inhibition of inducible nitric oxide synthase and cyclooxygenase II by Platycodon grandiflorum saponins via suppression of nuclear factor-kappaB activation in RAW 264.7 cells. Life Sci 2005, 76, 2315–2328. [Google Scholar]
- Khanal, T; Choi, JH; Hwang, YP; Chung, YC; Jeong, HG. Protective effects of saponins from the root of Platycodon grandiflorum against fatty liver in chronic ethanol feeding via the activation of AMP-dependent protein kinase. Food Chem. Toxicol 2009, 47, 2749–2754. [Google Scholar]
- Stevens, RL; Austen, KF. Recent advances in the cellular and molecular biology of mast cells. Immunol. Today 1989, 10, 381–386. [Google Scholar]
- Yamaguchi, M; Sayama, K; Yano, K; Lantz, CS; Noben-Trauth, N; Ra, C; Costa, JJ; Galli, SJ. IgE enhances Fc epsilon receptor I expression and IgE-dependent release of histamine and lipid mediators from human umbilical cord blood-derived mast cells: Synergistic effect of IL-4 and IgE on human mast cell Fc epsilon receptor I expression and mediator release. J Immunol 1999, 162, 5455–5465. [Google Scholar]
- Murakami, M; Kudo, I. Diversity and regulatory functions of mammalian secretory phospholipase A2s. Adv. Immunol 2001, 77, 163–194. [Google Scholar]
- Kitamura, Y. Recent progress of mast cell research. Arerugi 2005, 54, 45–47. [Google Scholar]
- Muraguchi, A; Kishimoto, T; Miki, Y; Kuritani, T; Kaieda, T; Yoshizaki, K; Yamamura, Y. T cell-replacing factor (TRF)-induced IgG secretion in a human B blastoid cell line and demonstration of acceptors for TRF. J. Immunol 1981, 127, 412–416. [Google Scholar]
- Naka, T; Nishimoto, N; Kishimoto, T. The paradigm of IL-6: From basic science to medicine. Arthritis Res 2002, 3, S233–242. [Google Scholar]
- Kamimura, D; Ishihara, K; Hirano, T. IL-6 signal transduction and its physiological roles: The signal orchestration model. Rev. Physiol. Biochem. Pharmacol 2003, 149, 1–38. [Google Scholar]
- Smith, WL; Song, I. The enzymology of prostaglandin endoperoxide H synthases-1 and -2. Prostaglandins Other Lipid Mediat 2002, 68–69, 115–128. [Google Scholar]
- Crofford, LJ. COX-1 and COX-2 tissue expression: Implications and predictions. J. Rheumatol. Suppl 1997, 49, 15–19. [Google Scholar]
- Masferrer, JL; Seibert, K. Regulation of prostaglandin synthesis by glucocorticoids. Receptor 1994, 4, 25–30. [Google Scholar]
- Mitchell, JA; Akarasereenont, P; Thiemermann, C; Flower, RJ; Vane, JR. Selectivity of nonsteroidal antiinflammatory drugs as inhibitors of constitutive and inducible cyclooxygenase. Proc. Natl. Acad. Sci USA 1993, 90, 11693–11697. [Google Scholar]
- Seibert, K; Masferrer, JL. Role of inducible cyclooxygenase (COX-2) in inflammation. Receptor 1994, 4, 17–23. [Google Scholar]
- Kay, AB. Overview of ‘allergy and allergic diseases: With a view to the future’. Br. Med. Bull 2000, 56, 843–864. [Google Scholar]
- Barnes, PJ; Adcock, I. Anti-inflammatory actions of steroids: Molecular mechanisms. Trends Pharmacol. Sci 1993, 14, 436–441. [Google Scholar]
- Murakami, M; Matsumoto, R; Austen, KF; Arm, JP. Prostaglandin endoperoxide synthase-1 and -2 couple to different transmembrane stimuli to generate prostaglandin D2 in mouse bone marrow-derived mast cells. J. Biol. Chem 1994, 269, 22269–22275. [Google Scholar]
- Murakami, M; Austen, KF; Arm, JP. The immediate phase of c-kit ligand stimulation of mouse bone marrow-derived mast cells elicits rapid leukotriene C4 generation through posttranslational activation of cytosolic phospholipase A2 and 5-lipoxygenase. J. Exp. Med 1995, 182, 197–206. [Google Scholar]
- Moon, TC; Murakami, M; Kudo, I; Son, KH; Kim, HP; Kang, SS; Chang, HW. A new class of COX-2 inhibitor, rutaecarpine from Evodia rutaecarpa. Inflam. Res 1999, 48, 621–625. [Google Scholar]
- Lee, SH; Son, MJ; Ju, HK; Lin, CX; Moon, TC; Choi, HG; Son, JK; Chang, HW. Dual inhibition of cyclooxygenases-2 and 5-lipoxygenase by deoxypodophyllotoxin in mouse bone marrow-derived mast cells. Biol. Pharm. Bull 2004, 27, 786–788. [Google Scholar]
- Fiorucci, S; Meli, R; Bucci, M; Cirino, G. Dual inhibitors of cyclooxygenase and 5-lipoxygenase. A new avenue in anti-inflammatory therapy? Biochem. Pharmacol 2001, 62, 1433–1438. [Google Scholar]
- Bochenek, G; Nizankowska, E; Gielicz, A; Swierczynska, M; Szczeklik, A. Plasma 9alpha, 11beta-PGF2, a PGD2 metabolite, as a sensitive marker of mast cell activation by allergen in bronchial asthma. Thorax 2004, 59, 459–464. [Google Scholar]
- Levi-Schaffer, F; Riesel, N; Soffer, D; Abramsky, O; Brenner, T. Mast cell activity in experimental allergic encephalomyelitis. Mol. Chem. Neuropathol 1991, 15, 173–184. [Google Scholar]
- Ha, YW; Na, YC; Seo, JJ; Kim, SN; Linhardt, RJ; Kim, YS. Qualitative and quantitative determination of ten major saponins in Platycodi Radix by high performance liquid chromatography with evaporative light scattering detection and mass spectrometry. J. Chromatogr. A 2006, 1135, 27–35. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Name | μg/mg |
|---|---|---|
| 1 | Deapi-Platycoside E | 1.4694 |
| 2 | Platycoside E | 1.6027 |
| 3 | Deapi-Platycodin D3 | ND |
| 4 | Platycodin D3 | 1.3450 |
| 5 | Deapi-Platycodin D | 1.6375 |
| 6 | Platycodin D | 5.6279 |
| 7 | Polygalacin D | 6.3629 |
| 8 | 3″-O-acetyl polygalacin D | 2.3310 |
| 9 | Platycodin A | 2.7392 |
| 10 | 2″-O-acetyl polygalacin D | 2.3579 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Oh, Y.-C.; Kang, O.-H.; Choi, J.-G.; Lee, Y.-S.; Brice, O.-O.; Jung, H.J.; Hong, S.-H.; Lee, Y.-M.; Shin, D.-W.; Kim, Y.-S.; et al. Anti-Allergic Activity of a Platycodon Root Ethanol Extract. Int. J. Mol. Sci. 2010, 11, 2746-2758. https://doi.org/10.3390/ijms11072746
Oh Y-C, Kang O-H, Choi J-G, Lee Y-S, Brice O-O, Jung HJ, Hong S-H, Lee Y-M, Shin D-W, Kim Y-S, et al. Anti-Allergic Activity of a Platycodon Root Ethanol Extract. International Journal of Molecular Sciences. 2010; 11(7):2746-2758. https://doi.org/10.3390/ijms11072746
Chicago/Turabian StyleOh, You-Chang, Ok-Hwa Kang, Jang-Gi Choi, Young-Seob Lee, Obiang-Obounou Brice, Hyun Ju Jung, Seung-Heon Hong, Young-Mi Lee, Dong-Won Shin, Yeong-Shik Kim, and et al. 2010. "Anti-Allergic Activity of a Platycodon Root Ethanol Extract" International Journal of Molecular Sciences 11, no. 7: 2746-2758. https://doi.org/10.3390/ijms11072746
APA StyleOh, Y.-C., Kang, O.-H., Choi, J.-G., Lee, Y.-S., Brice, O.-O., Jung, H. J., Hong, S.-H., Lee, Y.-M., Shin, D.-W., Kim, Y.-S., & Kwon, D.-Y. (2010). Anti-Allergic Activity of a Platycodon Root Ethanol Extract. International Journal of Molecular Sciences, 11(7), 2746-2758. https://doi.org/10.3390/ijms11072746