2.1. The effect of EF on unvaccinated chickens
In our study, we have followed the effect of EF from the edible mushroom
P. ostreatus on broiler chickens which have not been vaccinated with BIAVAC and BIAROMVAC vaccines (
Figure 1). In this case the maternal titers of infectious bursal disease virus antibodies (IBD–AB) (control) are seen to decrease, being at the minimum level (96) in four weeks post hatching (
Figure 1a). This antibody titer (96) was below the estimated cut-off level in this enzyme–linked immunosorbent assay (ELISA) system.
By analyzing the variation (
Table 1) we can observe significant differences (
p<0.001) in the change of IBD–AB maternal titer. The variation coefficient is low (5.97%) in the first week of chicken life and high (20.37%) in week four.
The
F-ratio, which in this case equals 3152.98, is a ratio of the between-group estimate to the within-group estimate. Since the
P-value of the
F-test is less than 0.05, there is a statistically significant difference between the means of the five weeks at the 95.0% confidence level. If we compare the distribution of maternal antibodies within each group during one week, but also between weeks, we can notice a significant difference (
Table 2).
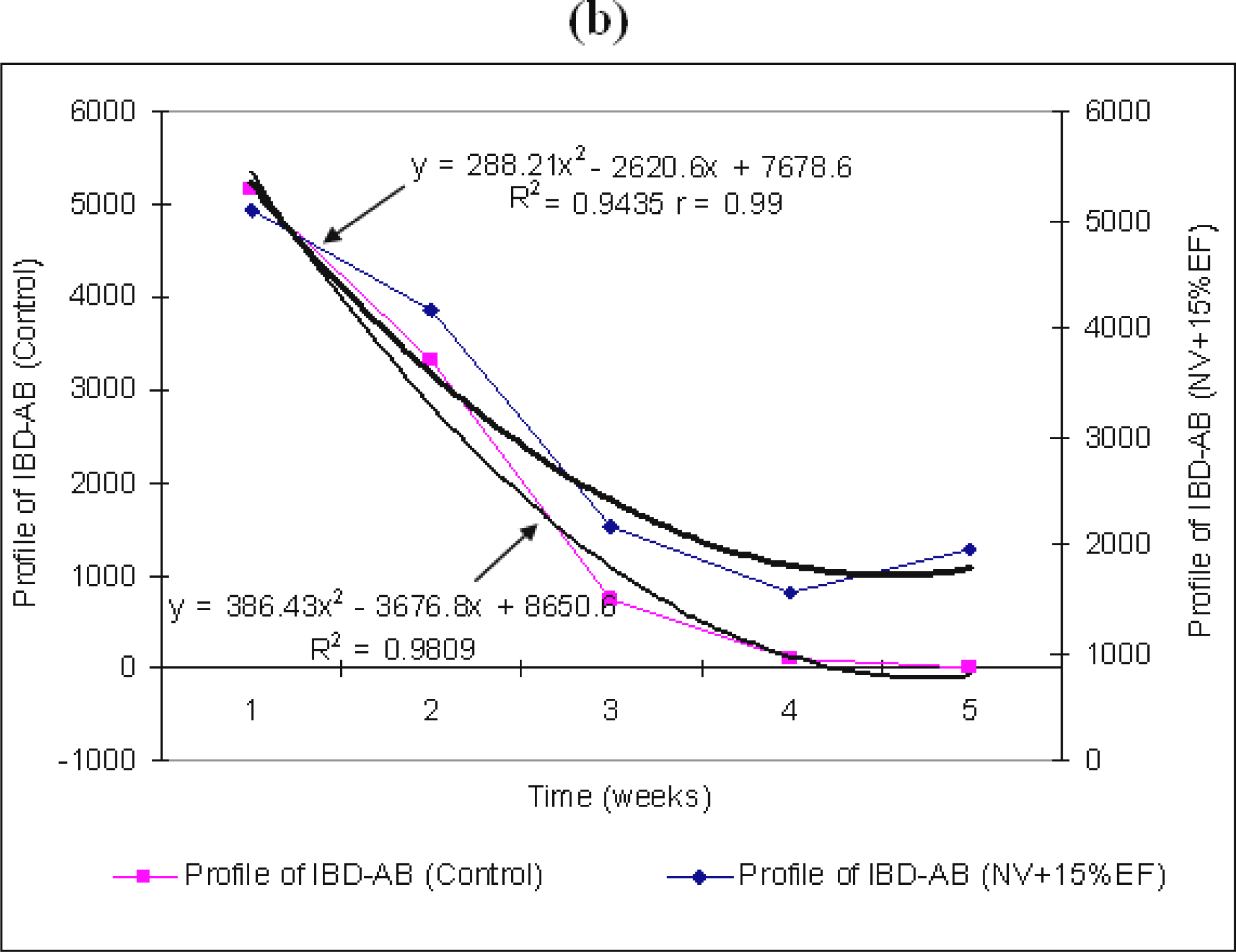
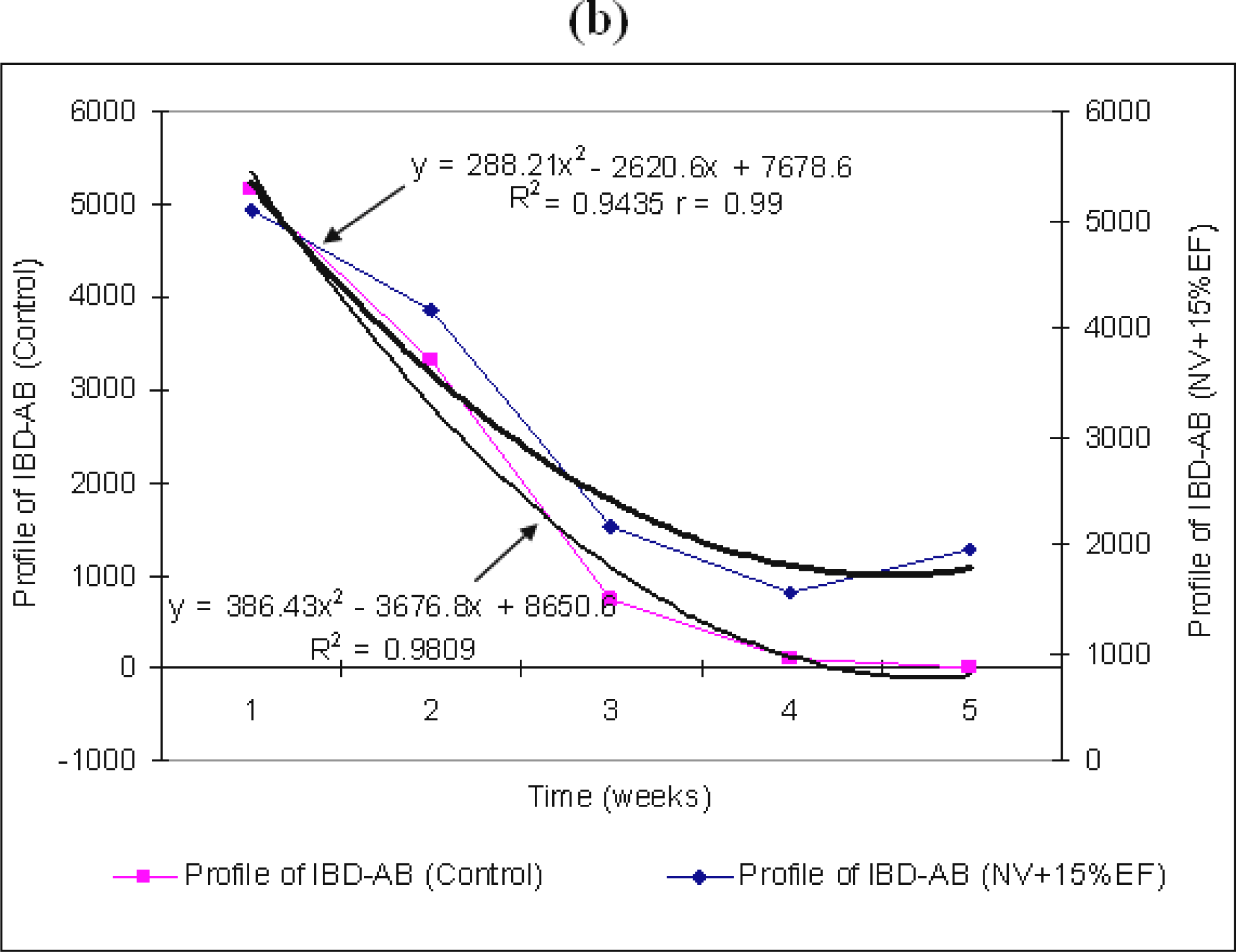
The difference between the antibodies titer averages in weeks four and five is not significant. Yet, by adding 5% EF in the water intake, we can notice a stabilization of the titer level for the stimulation of production of maternal IBD–AB, starting with the first days of week five (
p<0.0001). The standard deviation from the average of the maternal IBD–AB titer value is high in the mid-period of our experiment (21.26% in week three) and low (0.67%) in the last week (
Table 1).
The differences between the antibody titer averages between groups are significant, but those between the values of weeks three and four are insignificant. They are significant, but negative, between the average values of weeks three and five (−741.95) and between weeks four and five (−944.0) (
Table 2). This last aspect has been shown by stimulation of antibodies production (
Figure 1a) and is statistically explained by the chicken positive reaction (r=0.94) to EF action for IBD–AB stimulation and a homogeneity of broilers answers (biological activity) in IBD–AB production.
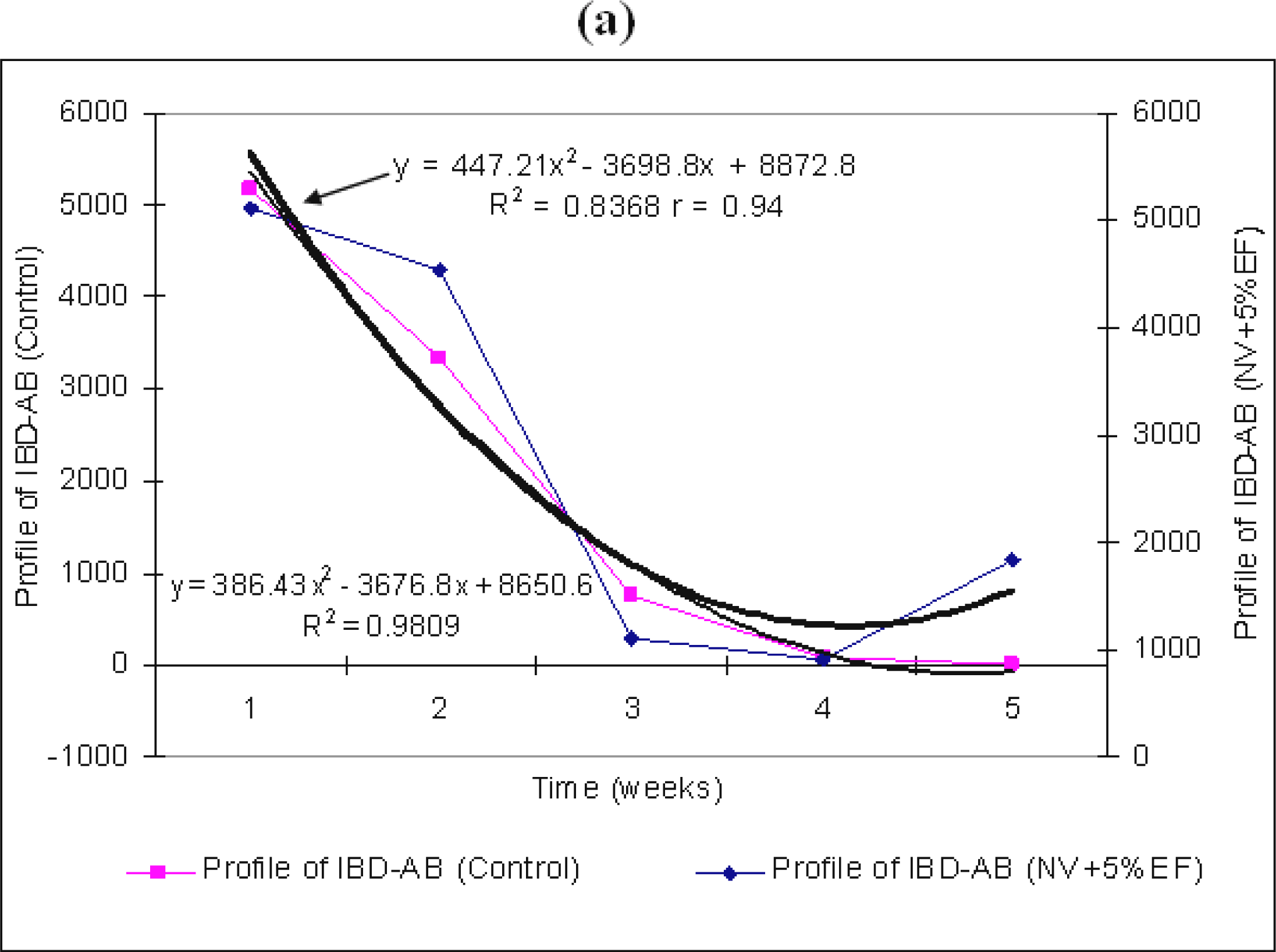
By supplementing the water intake with 15% EF from
P. ostreatus (
Figure 1b), standards deviations are low at the beginning of the experiment (3.68%) and towards the end of it (7.19% and 7.63%). The differences between antibodies titre averages between all averages antibodies groups are significant. A positive ratio (r=0.99) has been established between the two variables, EF and the capacity of chicken bodies to produce antibodies (
Figure 1b). The unvaccinated broilers + 5% EF have shown a better global behaviour, whose average of variation coefficient has been 9.56%, respect to both the unvaccinated broilers + 15% EF with an average of 8.65% and those unvaccinated without EF, which show an average of about 8.106%.
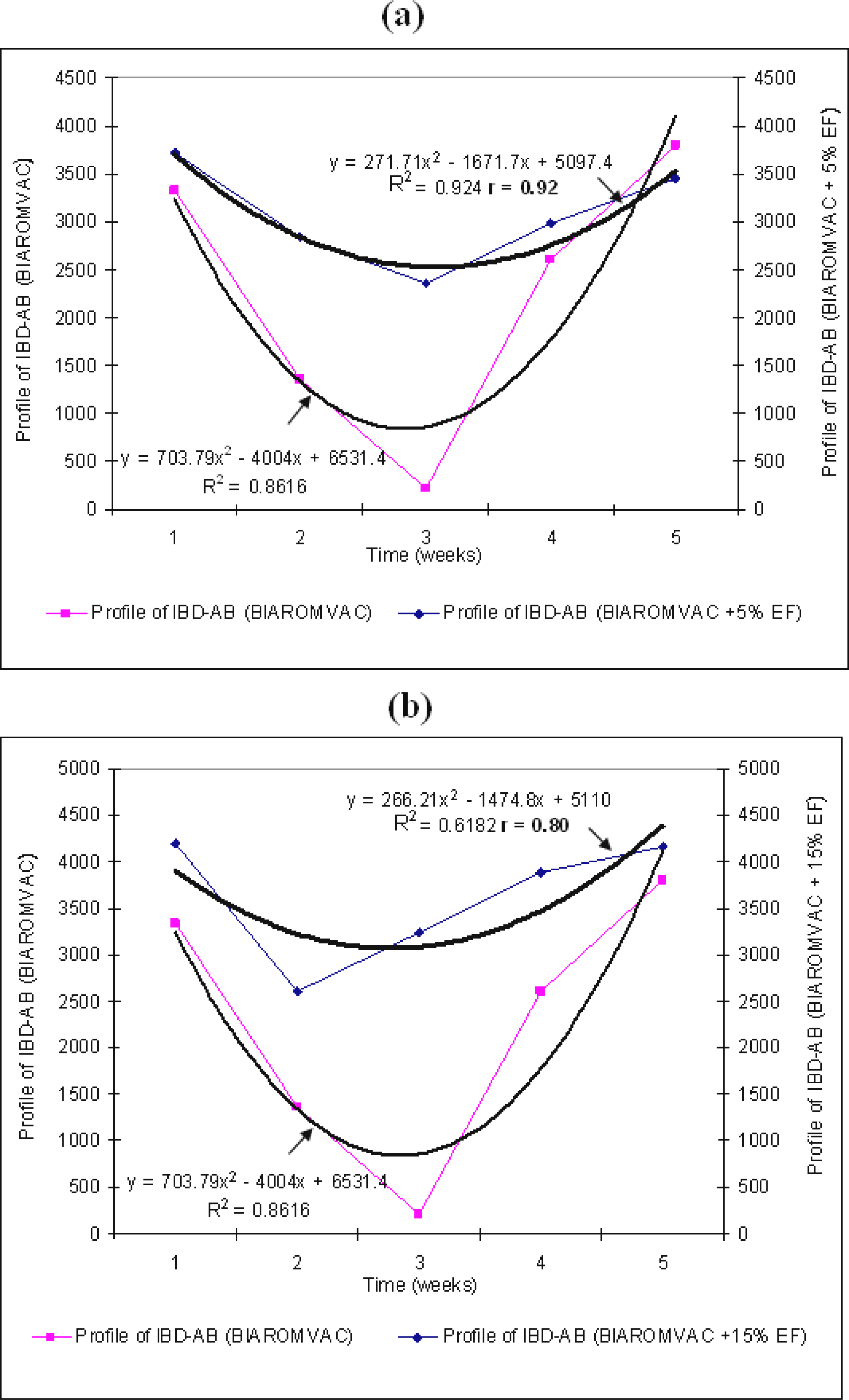
2.2. The effect of EF on broiler vaccination with BIAVAC vaccine
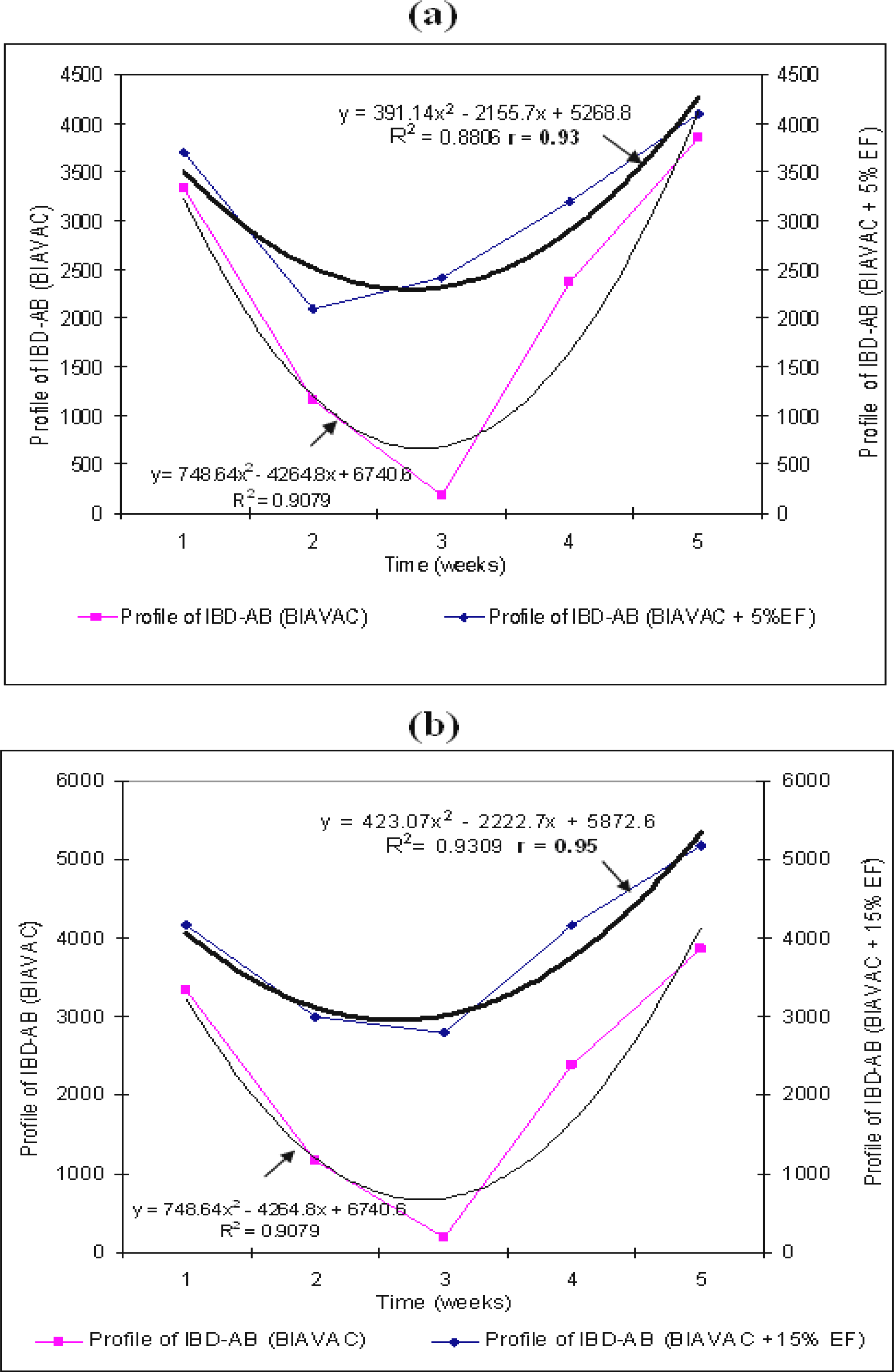
In this experimental group, we have followed the change of IBD–AB antibodies in broilers vaccinated with BIAVAC and treated with EF. Thus, all broilers, although vaccinated with BIAVAC, without EF treatment, have shown a decrease of the level of maternal IBD–AB antibodies. The lowest antibody titre (185) was below the estimated cut-off level in this ELISA system. The production of antibodies increased starting with week 4 (
p<0.0001;
Figure 2a;
Table 3).
The final level of this biosynthesis of antibodies is above the value of the titre on the 1
st day. By treating the intake water with 5% EF, the values of IBD–AB titre are maintained at a relatively high level (
p<0.0001), with slightly high indicators of variation coefficient, at the beginning and at the end of the treatment. We can notice (
Figure 2a;
Table 3) the action of EF in synergism with BIAVAC vaccine upon the production of antibodies (r=0.93). The standard deviations are significant when 15% EF was administered (
p<0.0001;
Figure 2b,
Table 3).
By a multiple comparison between the pairs means of IBD–AB: 1–2, 1–3 and 1–4 which correspond to the immunosuppressive period (weeks 1 and 3), we can notice significant positive differences with the BIAVAC experiment. Starting with week 4, between the pairs means of IBD–AB, we can notice significant negative differences (
Table 4). In this 2
nd half of the experimental period, the production of IBD–AB antibodies can be noticed, with a high level (3859.00), which is above the level of the 1
st day of post hatching. In the next experimental variants of the broilers (BIAVAC + 5% EF and BIAVAC + 15% EF), between the pairs means of IBD–AB we can notice significant differences of value, as compared to BIAVAC variants at the 95.0% confidence level (
Table 4).
The averages of variations are relatively homogenous, 11.63% (BIAVAC non-EF), 14.76% (BIAVAC + 5% EF) and 13.87% (BIAVAC + 15% EF), while the correlation coefficients between the variables of the system are positive, r=0.93 (BIAVAC + 5% EF) and r=0.95 (BIAVAC + 15% EF) (
Figures 2a,b).
2.3. The effect of EF on broilers vaccinated with BIAROMVAC vaccine
In this experiment, we have followed the production of IBD–AB antibodies in non-vaccinated chickens, vaccinated with BIAROMVAC, as compared to the ones under BIAROMVAC + 5% EF treatment and BIAROMVAC + 15% EF treatment, respectively. Thus, in chickens only vaccinated with BIAROMVAC vaccine, (like BIAVAC vaccine for the previous experiment), we can notice the temporary suppressive action of maternal IBD–AB antibodies production, in the first two weeks post-vaccinated inoculation (
Figures 3a,b). All broilers had a maternal IBD–AB titer of 217 in the ELISA. These antibodies were below the estimated cut-off level in this ELISA. In the experimental groups, to which we have added 5% and 15% EF, respectively, all along the duration of the experiment, the antibodies level remained high, with a slow decrease at the beginning of the treatment (
Figure 3,
Table 5;
p<0.0001).
We have noticed a better response in broilers treated with BIAROMVAC + 5%EF, whose average variation was 12.85%, as compared to the treatments BIAROMVAC without EF and BIAROMVAC + 15% EF, with averages of 14.39 and 14.02%, respectively. By comparison, the all pairs means of maternal IBD–AB have shown significant differences at 95.0% confidence level (
Table 6), except for one case (2–4) of the treatment with BIAROMVAC + 5% EF and two cases (1–4 and 1–5) of the treatment with BIAROMVAC + 15% EF. The correlation coefficient between the system variables are positive (r=0.92 and respectively, r=0.80) (
Figures 3a,b).
The real guideline basis of this experiment relies on the immunosuppressive effects in non-vaccinated chickens by BIAVAC, BIAROMVAC vaccines, and other IBDV vaccines in the first critical days of broilers life [
9,
10]. The field and clinical observations have shown differences in response of different lines genetic backgrounds upon the influence of some IBDV pathogenesis upon maternal antibodies. Also, from economical reasons, the studies are worth initiated and continued since the production of broilers decreases because of maternal antibodies decrease in progenies (
Tables 1,
3,
5). Another reason of these studies reflects the fact that not much is known as far the immunosuppressive abilities of IBD vaccines in commercial broilers is concerned [
11–
13]. In the present work, we compared immuno-modulatory effects of different concentrations of EF with polysaccharides extracted from
P. ostreatus with, but also without BIAVAC and BIAROMVAC vaccines, on commercial broilers. Our study proves the immunomodulatory ability of EF with polysaccharides extracted from
P. ostreatus together with BIAVAC and BIAROMVAC vaccines, towards the stimulation of IBD–AB antibodies production.
There are studies which demonstrate, to a certain degree of certitude, the existence of some connections between IBDV–induction of immunosuppression and the severe injuries which appear in bursa of Fabricius. Even more, there have been situations, even as hypothesis, of association of IBDV with B cells, macrophages and follicular dendritic cells in bursa and spleen [
8]. Other fundamental studies mention that these IBD–AB vaccines may induce suppressive secondary effects in the immune system, such as systemic T cell activity. Thus, the macrophages may induce the release of cytokines and nitric oxide which can suppress T cell activity. Another hypothesis is that the suppression of T helper cell activity might favour to reduce IBD–AB antibodies response in broilers [
14]. Also in the cases of our research, we have noticed such a temporary suppression of IBD–AB antibodies in all experiments in broilers (
Figure 1–
3), when we have used BIAVAC and BIAROMVAC vaccines. By supplementing the water intake with EF which has polysaccharides extracted from
P. ostreatus we have noticed a stimulation of IBD–AB antibodies production in all experimental groups. Scientific data show that substances from edible mushrooms are responsible for imunodulating effects. These substances are β-
d-glucan polysaccharides. The β-
d-glucans are a heterogeneous group of glucose polymers. These polysaccharides are the major cells wall structural components found in fungi and also present in plant and some bacteria. Studies with β-
d-glucan from
P. ostratus or from
L. edodes, have shown several immuno-modulatory properties which include increase of T-cells activity, restoration of T-helper cells suppression, induction of cytotoxic activity of peritoneal macrophages. The β-glucans bind to receptors on cells membranes of macrophages, neutrophils, natural killer cells, T-cells dendritic cells, fibroblast and vascular endothelial cells. The molecular structure of these substances from fungi influences the affinity to receptors and the stimulation of the immunomdulator system [
15,
16].
2.4. Reactivity analysis of the parabolic biological activity relationship
The results of previous sections opens the possibility for further systematic treatment of IBD–AB activity (
y) through employing the fitted response curves of
Figures 1–
3 beyond the statistical correlation factor and fitted parameters.
Actually, since the reported ANOVA analysis in
Figures 1–
3 unfolds under the parabolic form:
it readily reminding of the chemical reactivity law modeling the (molecular) systems’ energy change with the number of donated/accepted electrons Δ
N :
through the electronegativity
χ and chemical hardness
η indices:
in terms of ionization potential (
IP) and electron affinity (
EA), respectively [
17].
Now, the idea is to rearrange
Equation (1) under the equivalent form given by
Equation (2) that will reveal us how the fitting coefficients of
Equation (1) will correspond to the reactivity indices in
Equation (2). Yet, such transformation is not direct, but upon considering appropriate operations. As such, the first step regards the translation of the form given by
Equation (1) into a parabolic form centred on its optimum values, either for dependent
Yopt and independent
Xopt variables:
while the optimum expression are found by the variation of
Equation (1):
or, analytically as:
Then, in order for coefficients
α and
β to play the “reactive” role in
Equation (4) as electronegativity and chemical hardness do in
Equation (2), respectively, they have to be considered with a similar expression as that of
Equation (3), namely:
however, linked with the input
equation (1) by means of assignments:
thus providing the results:
Now, paralleling the chemical reactivity principles of electronegativity and chemical hardness [
17] to the coefficients
α and
β of
Equation (4), respectively, one can infer the hierarchies among two biological-chemical tested systems for which the two pairs of coefficients, say (
αI,
βI) for the system “I” and (
αII,
βII) for the system “II”, have been computed.
The electronegativity principles would lead with the rule that as the α index of a system is higher as the system will display larger reactivity/activity, here translated as growing of the antibody metabolic action; in short: “as α increases as grows the biological activity”, i.e., the stimulation of antibody synthesis; this may be eventually called as The First Principle of Reactive Biological Activity (ReBiAc1).
Instead, the chemical hardness principles, especially the maximum hardness one [
17], would lead with the idea that the
β factor tendency is detrimental to the biological activity, or, analytically: “as
β increases as slows the biological activity”, here - the immunological action; this may be eventually called as
The Second Principle of Reactive Biological Activity (ReBiAc2).
Table 7 collects all the fitted and
reactive biological activity (ReBiAc) data from the
Figures 1–
3 and the
equations (1)–
(9), respectively; it allows a global view of the experiments either between groups with various EF belonging to the same vaccine or not-vaccinated chickens. Furthermore, the present ReBiAc analysis finely clarifies upon the hierarchy of experiments in providing viable biological activity (here viewed as the potency of antibody synthesis). For instance, if one looks to the ordering correlation factors from the experimental systems I–IX (of
Table 7) there finds the puzzling hierarchy:
while this is rearranged in terms of increasing beneficial activity by the ReBiAc1 principle:
or as the decrease of antagonist activity in the light of ReBiAc2 principle:
From both these chains of bio-activity orderings appears the decisive influence EF has on anti-bursal vaccines since placed on the extreme of ReBiAc principles’ records (maximums for α, and minimums for β values).
This way, the analytical and conceptual tools for cross-judging both the favourable and detrimental (natural or induced) biological actions were formulated and tested. Moreover, is phenomenologically proven that each biological system described by a parabolic dependency contains the intrinsic positive (beneficial) and negative (detrimental) effects at the metabolic level, however in different degree. Nevertheless, the actual study shows how the beneficial influence may be adjusted to prevail on the harmful one by controlling the induced biological activity, here through administrating various EF with polysaccharides from Pleurotus.


 : the titer of maternal IBD–AB in un-vaccinated chicken NV + 5% (a) and 15% EF (b). Weeks mean weeks post hatch.
: the titer of maternal IBD–AB in un-vaccinated chicken NV + 5% (a) and 15% EF (b). Weeks mean weeks post hatch.




{kind=link}
{kind=link}
{kind=link}
{kind=link}