
Squalane as a Promising Agent Protecting UV-Induced Inhibition of Collagen Biosynthesis and Wound Healing in Human Dermal Fibroblast

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. The Impact of UV Radiation on Human Dermal Fibroblast’s Metabolism
1.2. Squalane and Squalene: Structural and Functional Distinctions in Dermatological Applications
2. Results and Discussion
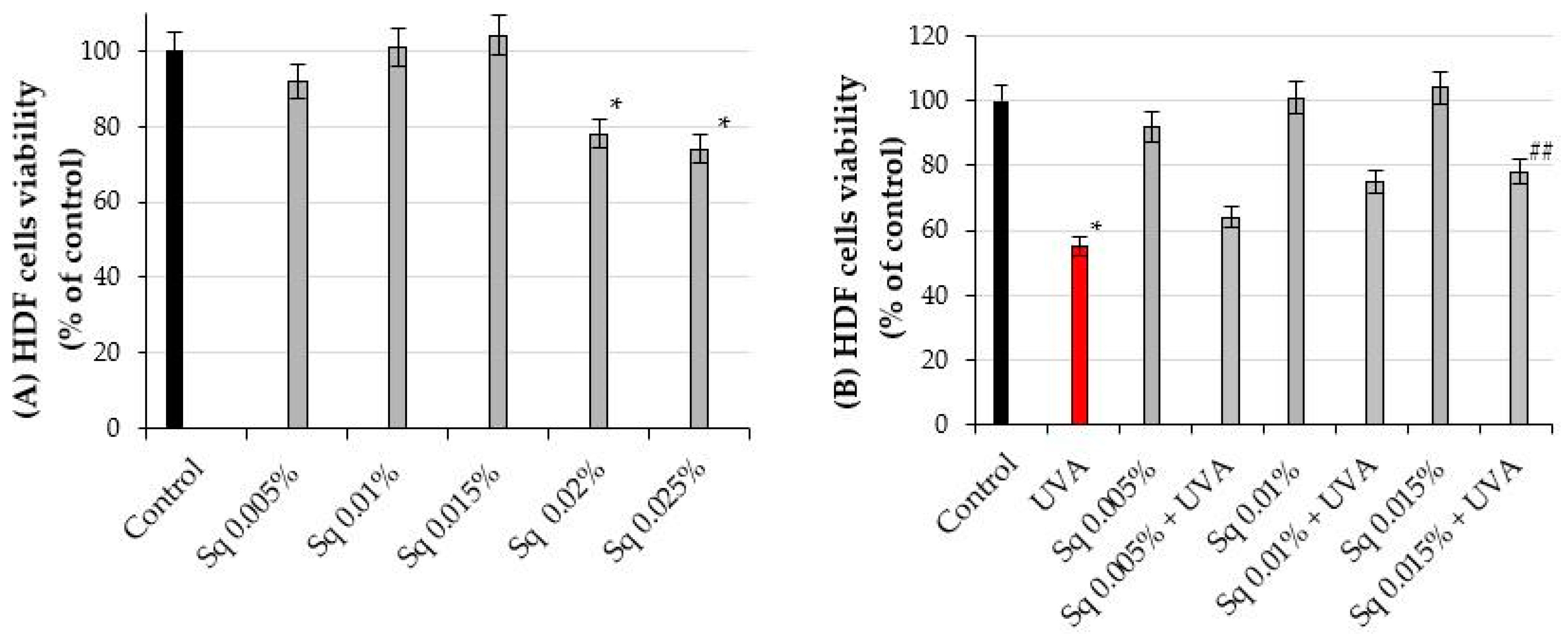
2.1. Effect of Squalane on Viability of UVA-Irradiated Human Dermal Fibroblasts
2.2. The Effect of Sq on UVA-Induced Inhibition of Collagen Biosynthesis and Prolidase Activity in HDsF
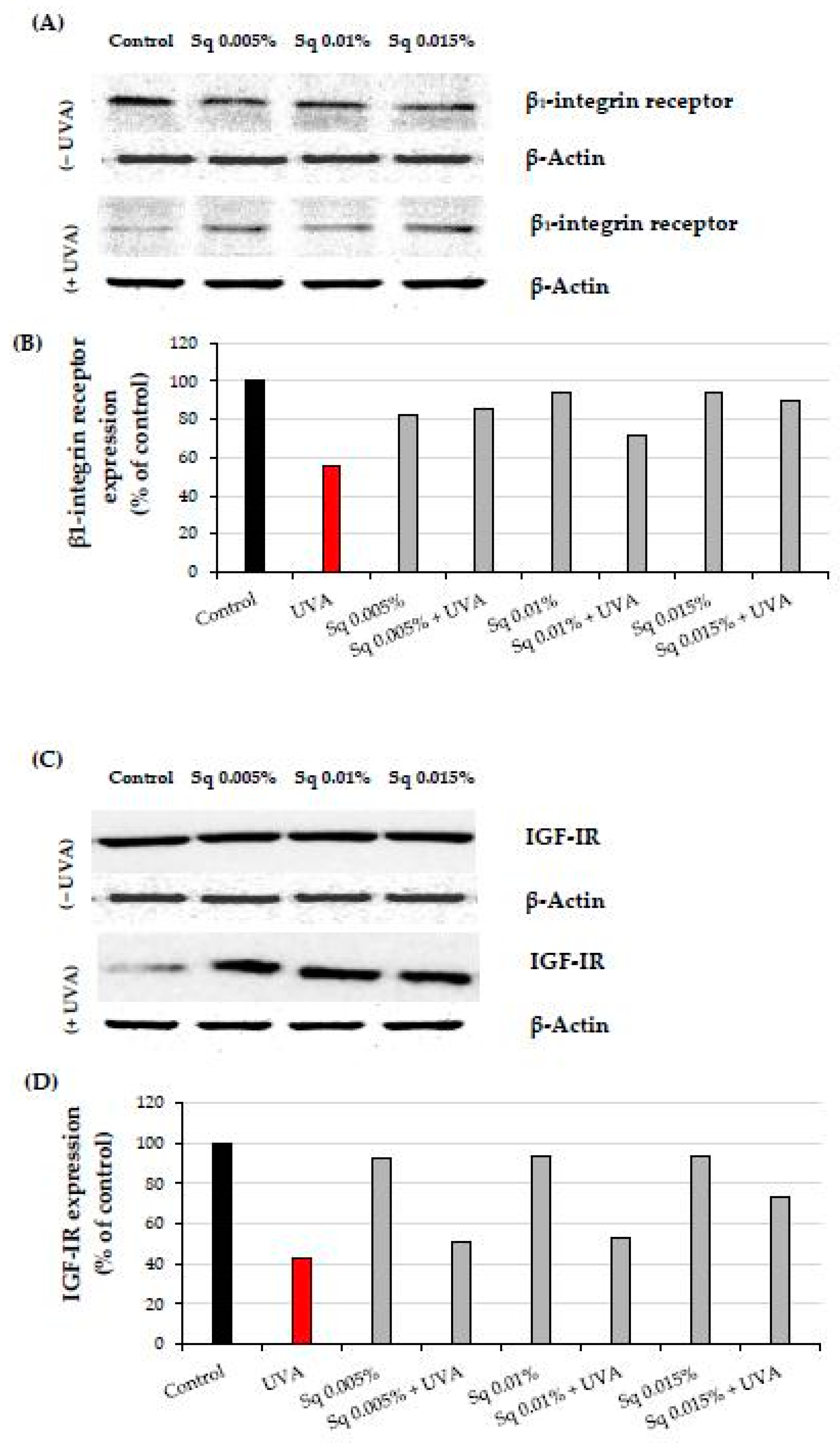
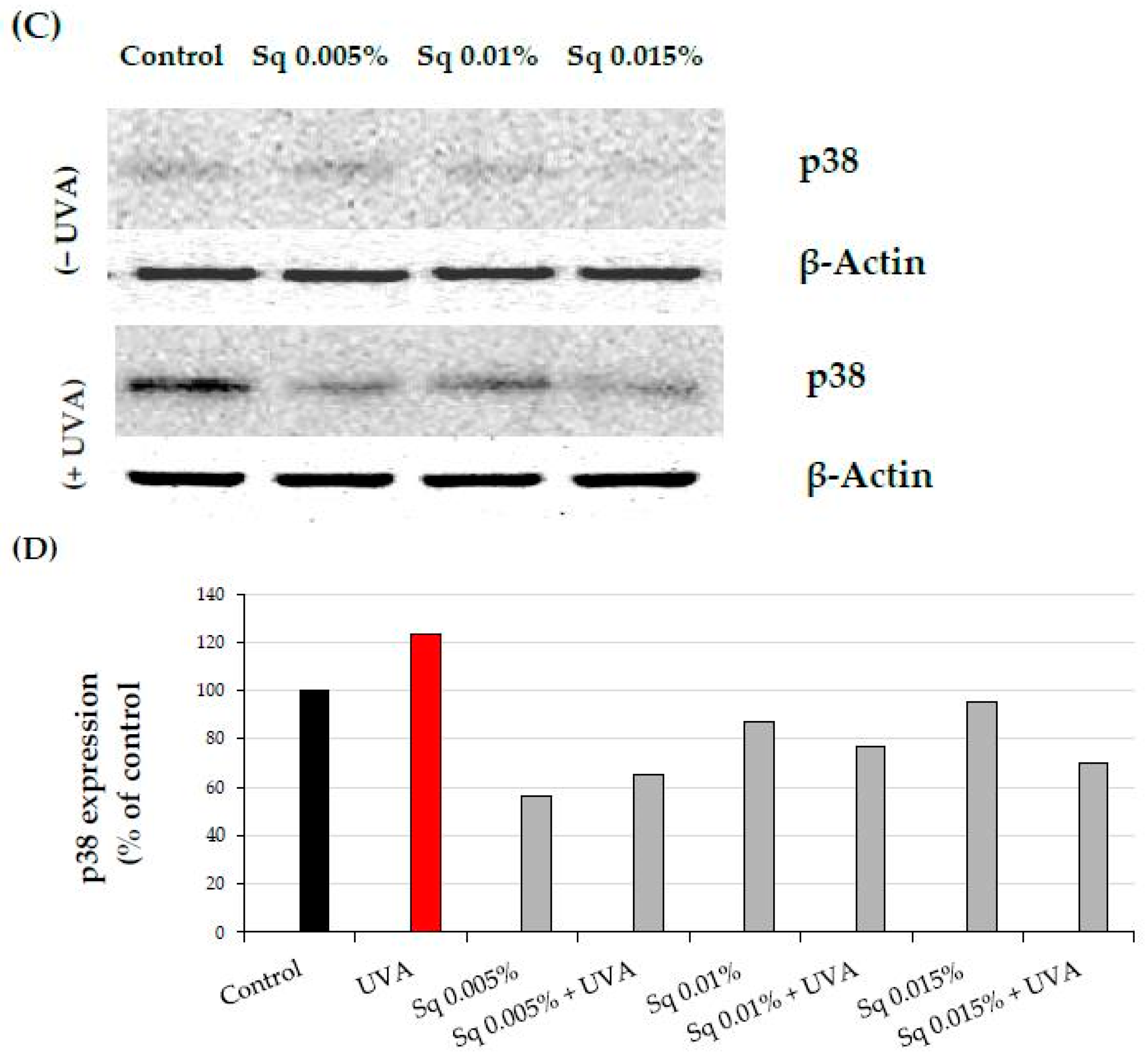
2.3. The Effect of Sq on the UVA-Induced Inhibition of Cell Signaling in HDFs
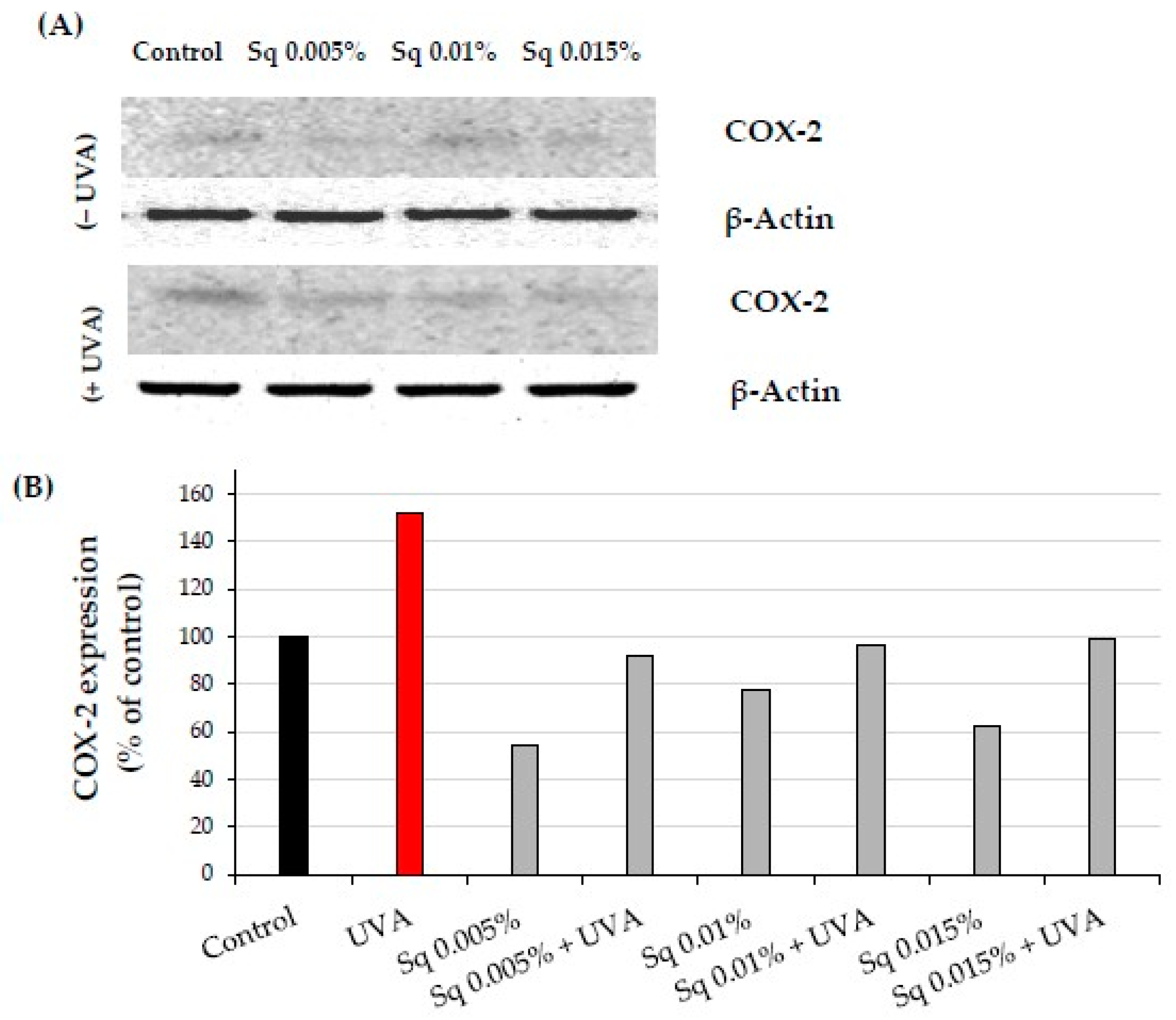
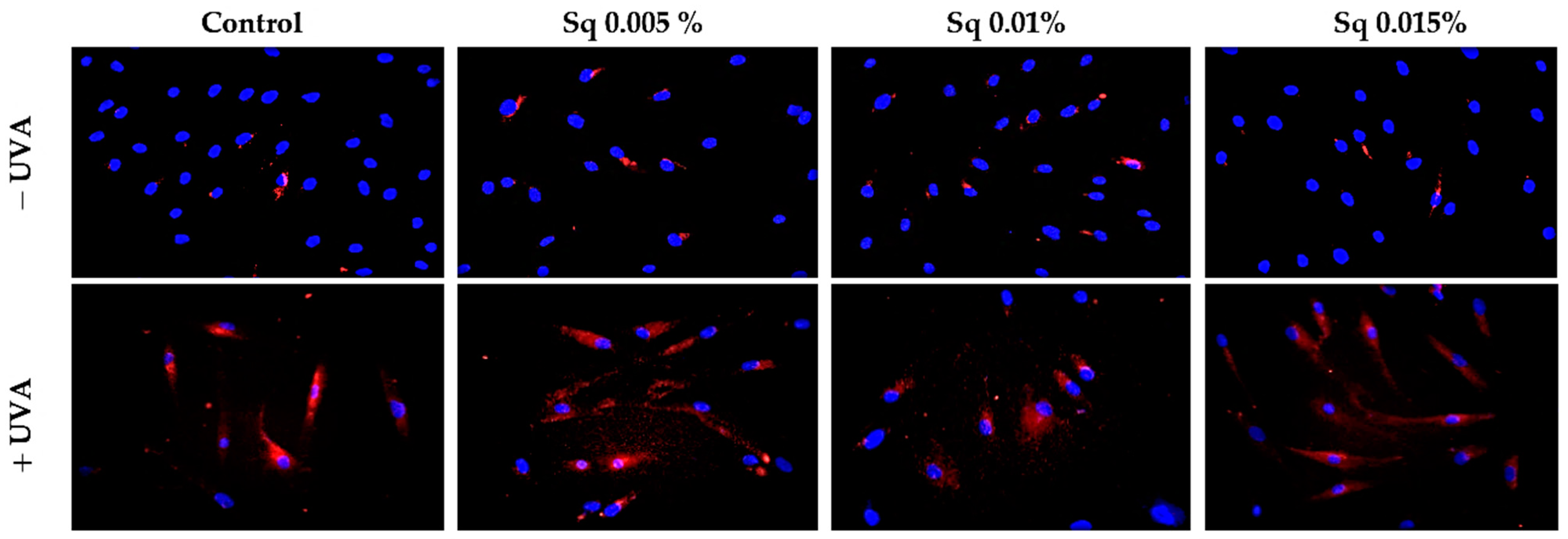
2.4. The Effect of Sq on UVA-Induced Inflammation in HDF
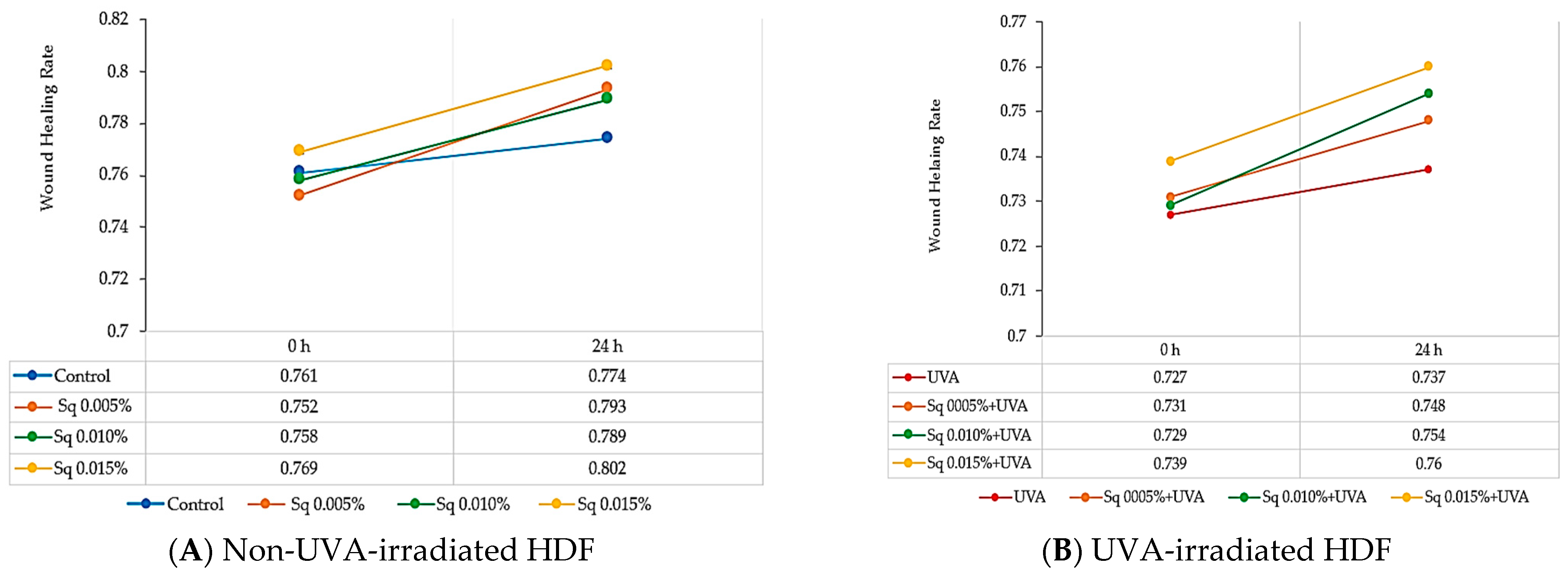
2.5. The Effect of Sq on Wound Healing in UVA-Irradiated HDFs
3. Materials and Methods
3.1. Materials
- Product Specifications:
- Product Name (IUPAC): Neossance Squalane (2,6,10,15,19,23-hexamethyltetracosane)
- Purity (%): Min. 92
- Density (g/cm3): Min. 0.806–Max. 0.811
- Iodine Number (g I2/100 g): Max. 2
- Acid Value (mg KOH/g oil): Max. 0.5
- Odor: Nearly odorless
- Color: Colorless
3.2. Methods
3.2.1. Cell Culture
3.2.2. Assessment of Cytotoxicity
3.2.3. Collagen Biosynthesis Assay
3.2.4. Prolidase Activity Assay
3.2.5. Western Blot Analysis
3.2.6. Protein Expression Visualization by Confocal Microscopy
3.2.7. Antibodies
3.2.8. Fibroblast Migration Using Scratch Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| COX-2 | Cyclooxygenase-2 |
| ERK1/2 | Extracellular Signal-Regulated Kinases 1/2 |
| HDF | Human Dermal Fibroblasts |
| Sq | Squalane |
| UVA | Ultraviolet A Radiation |
| ROS | Reactive Oxygen Species |
| NF-κB | Nuclear Factor Kappa B |
| IGF-IR | Insulin-Like Growth Factor I Receptor |
| β1-integrin | Beta 1 Integrin |
| Nrf2 | Nuclear Factor Erythroid 2–Related Factor 2 |
| TEWL | Transepidermal Water Loss |
| MAPKs | Mitogen-Activated Protein Kinases |
| TGF-β | Transforming Growth Factor Beta |
References
- Yaar, M.; Gilchrest, B.A. Photoageing: Mechanism, Prevention and Therapy. Br. J. Dermatol. 2007, 157, 874–887. [Google Scholar] [CrossRef]
- Lavker, R.M.; Gerberick, G.F.; Veres, D.; Irwin, C.J.; Kaidbey, K.H. Cumulative Effects from Repeated Exposures to Suberythemal Doses of UVB and UVA in Human Skin. J. Am. Acad. Dermatol. 1995, 32, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Hu, J.Y.; Wang, S.Q. The Role of Antioxidants in Photoprotection: A Critical Review. J. Am. Acad. Dermatol. 2012, 67, 1013–1024. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, H.; Yang, Y.; Zhang, S.; Wang, J.; Zhang, D.; Yu, H. Metformin Attenuates UVA-Induced Skin Photoaging by Suppressing Mitophagy and the PI3K/AKT/mTOR Pathway. Int. J. Mol. Sci. 2022, 23, 6960. [Google Scholar] [CrossRef] [PubMed]
- Surazynski, A.; Miltyk, W.; Palka, J.; Phang, J.M. Prolidase-Dependent Regulation of Collagen Biosynthesis. Amino Acids 2008, 35, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Baszanowska, W.; Misiura, M.; Oscilowska, I.; Palka, J.; Miltyk, W. Extracellular Prolidase (PEPD) Induces Anabolic Processes through EGFR, Β1-Integrin, and IGF-1R Signaling Pathways in an Experimental Model of Wounded Fibroblasts. Int. J. Mol. Sci. 2021, 22, 942. [Google Scholar] [CrossRef]
- Xia, Z.; Dickens, M.; Raingeaud, J.; Davis, R.J.; Greenberg, M.E. Opposing Effects of ERK and JNK-P38 MAP Kinases on Apoptosis. Science 1995, 270, 1326–1331. [Google Scholar] [CrossRef]
- Verheij, M.; Bose, R.; Lin, X.H.; Yao, B.; Jarvis, W.D.; Grant, S.; Birrer, M.J.; Szabo, E.; Zon, L.I.; Kyriakis, J.M.; et al. Requirement for Ceramide-Initiated SAPK/JNK Signalling in Stress-Induced Apoptosis. Nature 1996, 380, 75–79. [Google Scholar] [CrossRef]
- Kohen, R. Skin Antioxidants: Their Role in Aging and in Oxidative Stress--New Approaches for Their Evaluation. Biomed. Pharmacother. Biomedecine Pharmacother. 1999, 53, 181–192. [Google Scholar] [CrossRef]
- Dadej, I.; Wołowiec, J. The Role of UVA of the Skin Pathology. Adv. Dermatol. Allergol. Dermatol. Alergol. 2003, 20, 170–175. [Google Scholar]
- Battie, C.; Jitsukawa, S.; Bernerd, F.; Del Bino, S.; Marionnet, C.; Verschoore, M. New Insights in Photoaging, UVA Induced Damage and Skin Types. Exp. Dermatol. 2014, 23, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Vostalova, J.; Rajnochova Svobodova, A.; Galandakova, A.; Sianska, J.; Dolezal, D.; Ulrichova, J. Differential Modulation of Inflammatory Markers in Plasma and Skin after Single Exposures to UVA or UVB Radiation in Vivo. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czechoslov. 2013, 157, 137–145. [Google Scholar] [CrossRef]
- Huang, Z.-R.; Lin, Y.-K.; Fang, J.-Y. Biological and Pharmacological Activities of Squalene and Related Compounds: Potential Uses in Cosmetic Dermatology. Molecules 2009, 14, 540–554. [Google Scholar] [CrossRef]
- Oliveira, A.L.S.; Valente, D.; Moreira, H.R.; Pintado, M.; Costa, P. Effect of Squalane-Based Emulsion on Polyphenols Skin Penetration: Ex Vivo Skin Study. Colloids Surf. B Biointerfaces 2022, 218, 112779. [Google Scholar] [CrossRef]
- Barp, L.; Miklavčič Višnjevec, A.; Moret, S. Analytical Determination of Squalene in Extra Virgin Olive Oil and Olive Processing By-Products, and Its Valorization as an Ingredient in Functional Food—A Critical Review. Molecules 2024, 29, 5201. [Google Scholar] [CrossRef]
- Final Report on the Safety Assessment of Squalane and Squalene. J. Am. Coll. Toxicol. 1990, 1, 37–56. [CrossRef]
- Wolosik, K.; Chalecka, M.; Palka, J.; Surazynski, A. Protective Effect of Amaranthus Cruentus L. Seed Oil on UVA-Radiation-Induced Apoptosis in Human Skin Fibroblasts. Int. J. Mol. Sci. 2023, 24, 10795. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.I.; Naina Mohamed, I. Interdependence of Anti-Inflammatory and Antioxidant Properties of Squalene–Implication for Cardiovascular Health. Life 2021, 11, 103. [Google Scholar] [CrossRef]
- Jariashvili, K.; Madhan, B.; Brodsky, B.; Kuchava, A.; Namicheishvili, L.; Metreveli, N. Uv Damage of Collagen: Insights from Model Collagen Peptides. Biopolymers 2012, 97, 189–198. [Google Scholar] [CrossRef]
- Wolosik, K.; Chalecka, M.; Palka, J.; Mitera, B.; Surazynski, A. Amaranthus Cruentus L. Seed Oil Counteracts UVA-Radiation-Induced Inhibition of Collagen Biosynthesis and Wound Healing in Human Skin Fibroblasts. Int. J. Mol. Sci. 2024, 25, 925. [Google Scholar] [CrossRef]
- Afaq, F.; Malik, A.; Syed, D.; Maes, D.; Matsui, M.S.; Mukhtar, H. Pomegranate Fruit Extract Modulates UV-B-Mediated Phosphorylation of Mitogen-Activated Protein Kinases and Activation of Nuclear Factor Kappa B in Normal Human Epidermal Keratinocytes Paragraph Sign. Photochem. Photobiol. 2005, 81, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K.; Matsui, M.S.; Elmets, C.A.; Mukhtar, H. Polyphenolic Antioxidant (-)-Epigallocatechin-3-Gallate from Green Tea Reduces UVB-Induced Inflammatory Responses and Infiltration of Leukocytes in Human Skin. Photochem. Photobiol. 1999, 69, 148–153. [Google Scholar] [CrossRef]
- Rippe, R.A.; Schrum, L.W.; Stefanovic, B.; Solis-Herruzo, J.A.; Brenner, D.A. NF-kappaB Inhibits Expression of the Alpha1(I) Collagen Gene. DNA Cell Biol. 1999, 18, 751–761. [Google Scholar] [CrossRef]
- O’Dea, E.L.; Kearns, J.D.; Hoffmann, A. UV as an Amplifier Rather than Inducer of NF-κB Activity. Mol. Cell 2008, 30, 632–641. [Google Scholar] [CrossRef]
- Ryšavá, A.; Vostálová, J.; Rajnochová Svobodová, A. Effect of Ultraviolet Radiation on the Nrf2 Signaling Pathway in Skin Cells. Int. J. Radiat. Biol. 2021, 97, 1383–1403. [Google Scholar] [CrossRef] [PubMed]
- Gęgotek, A.; Jastrząb, A.; Jarocka-Karpowicz, I.; Muszyńska, M.; Skrzydlewska, E. The Effect of Sea Buckthorn (Hippophae Rhamnoides L.) Seed Oil on UV-Induced Changes in Lipid Metabolism of Human Skin Cells. Antioxidants 2018, 7, 110. [Google Scholar] [CrossRef]
- Darr, D.; Fridovich, I. Free Radicals in Cutaneous Biology. J. Invest. Dermatol. 1994, 102, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Poon, F.; Kang, S.; Chien, A.L. Mechanisms and Treatments of Photoaging. Photodermatol. Photoimmunol. Photomed. 2015, 31, 65–74. [Google Scholar] [CrossRef]
- Carmichael, J.; DeGraff, W.G.; Gazdar, A.F.; Minna, J.D.; Mitchell, J.B. Evaluation of a Tetrazolium-Based Semiautomated Colorimetric Assay: Assessment of Chemosensitivity Testing. Cancer Res. 1987, 47, 936–942. [Google Scholar]
- Peterkofsky, B.; Diegelmann, R. Use of a Mixture of Proteinase-Free Collagenases for the Specific Assay of Radioactive Collagen in the Presence of Other Proteins. Biochemistry 1971, 10, 988–994. [Google Scholar] [CrossRef]
- Myara, I.; Charpentier, C.; Lemonnier, A. Optimal Conditions for Prolidase Assay by Proline Colorimetric Determination: Application to Iminodipeptiduria. Clin. Chim. Acta 1982, 125, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolosik, K.; Chalecka, M.; Gasiewska, G.; Palka, J.; Surazynski, A. Squalane as a Promising Agent Protecting UV-Induced Inhibition of Collagen Biosynthesis and Wound Healing in Human Dermal Fibroblast. Molecules 2025, 30, 1964. https://doi.org/10.3390/molecules30091964
Wolosik K, Chalecka M, Gasiewska G, Palka J, Surazynski A. Squalane as a Promising Agent Protecting UV-Induced Inhibition of Collagen Biosynthesis and Wound Healing in Human Dermal Fibroblast. Molecules. 2025; 30(9):1964. https://doi.org/10.3390/molecules30091964
Chicago/Turabian StyleWolosik, Katarzyna, Magda Chalecka, Gabriela Gasiewska, Jerzy Palka, and Arkadiusz Surazynski. 2025. "Squalane as a Promising Agent Protecting UV-Induced Inhibition of Collagen Biosynthesis and Wound Healing in Human Dermal Fibroblast" Molecules 30, no. 9: 1964. https://doi.org/10.3390/molecules30091964
APA StyleWolosik, K., Chalecka, M., Gasiewska, G., Palka, J., & Surazynski, A. (2025). Squalane as a Promising Agent Protecting UV-Induced Inhibition of Collagen Biosynthesis and Wound Healing in Human Dermal Fibroblast. Molecules, 30(9), 1964. https://doi.org/10.3390/molecules30091964