
Verification of In Vitro Anticancer Activity and Bioactive Compounds in Cordyceps Militaris-Infused Sweet Potato Shochu Spirits

 , , , ,
, , , , 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
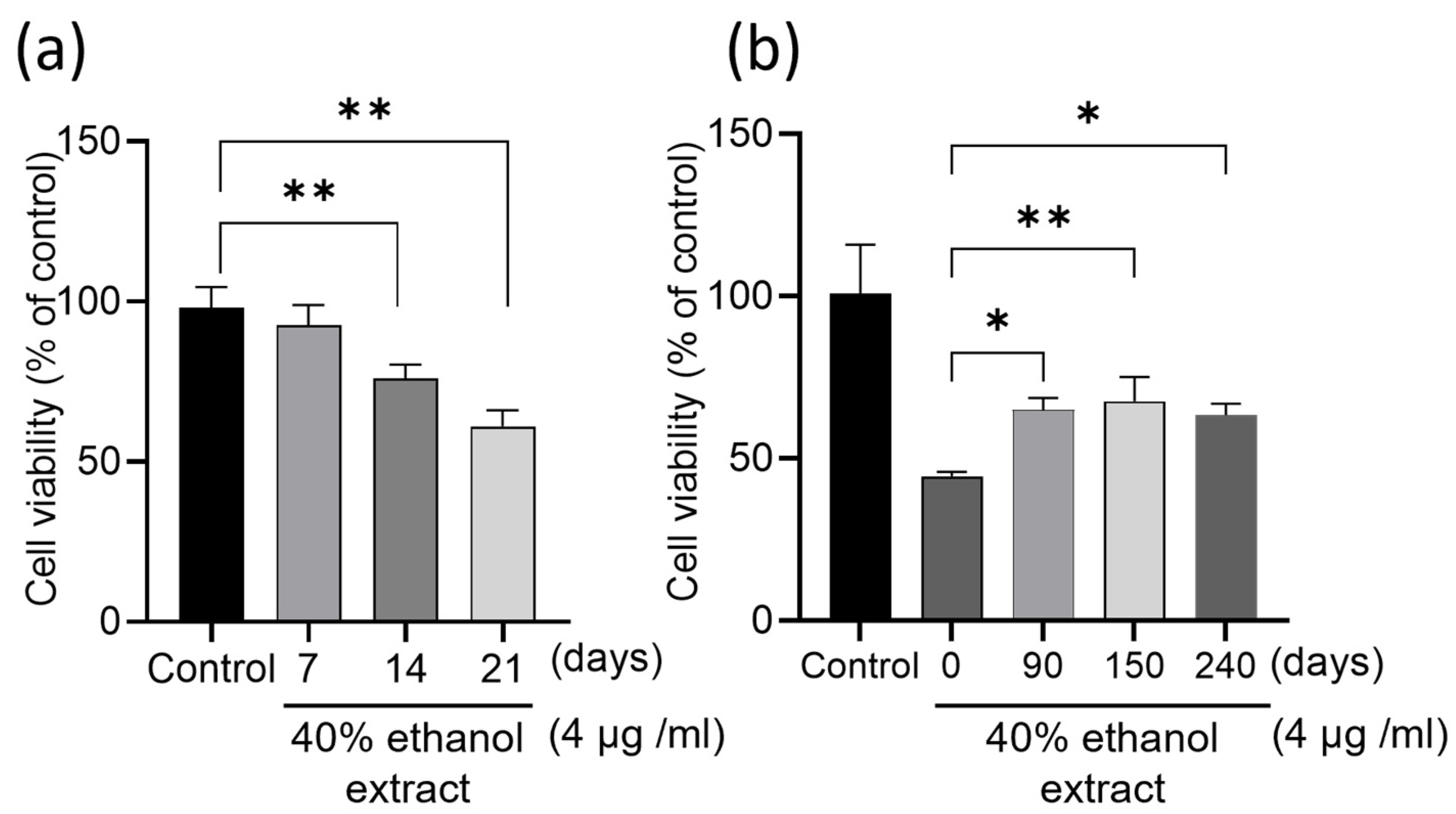
2.1. Influence of Infusion Times and Storage Period on HCT-116 Cell Proliferation
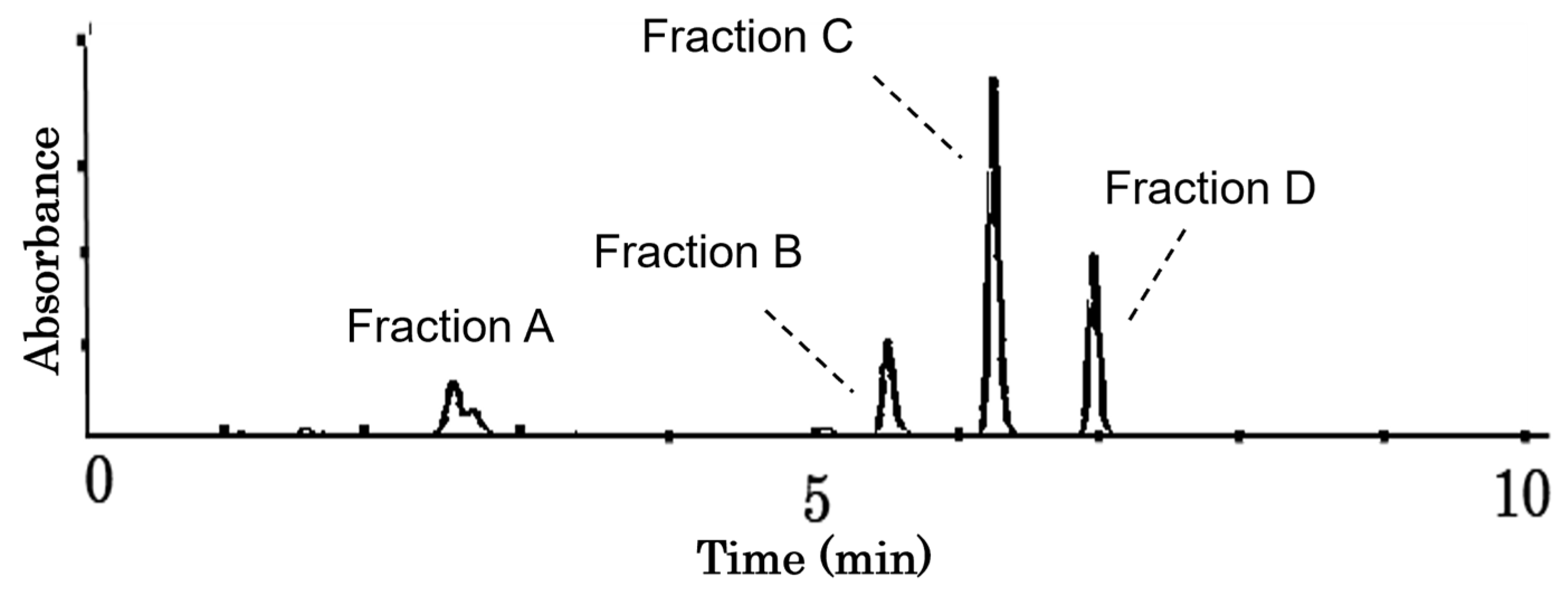
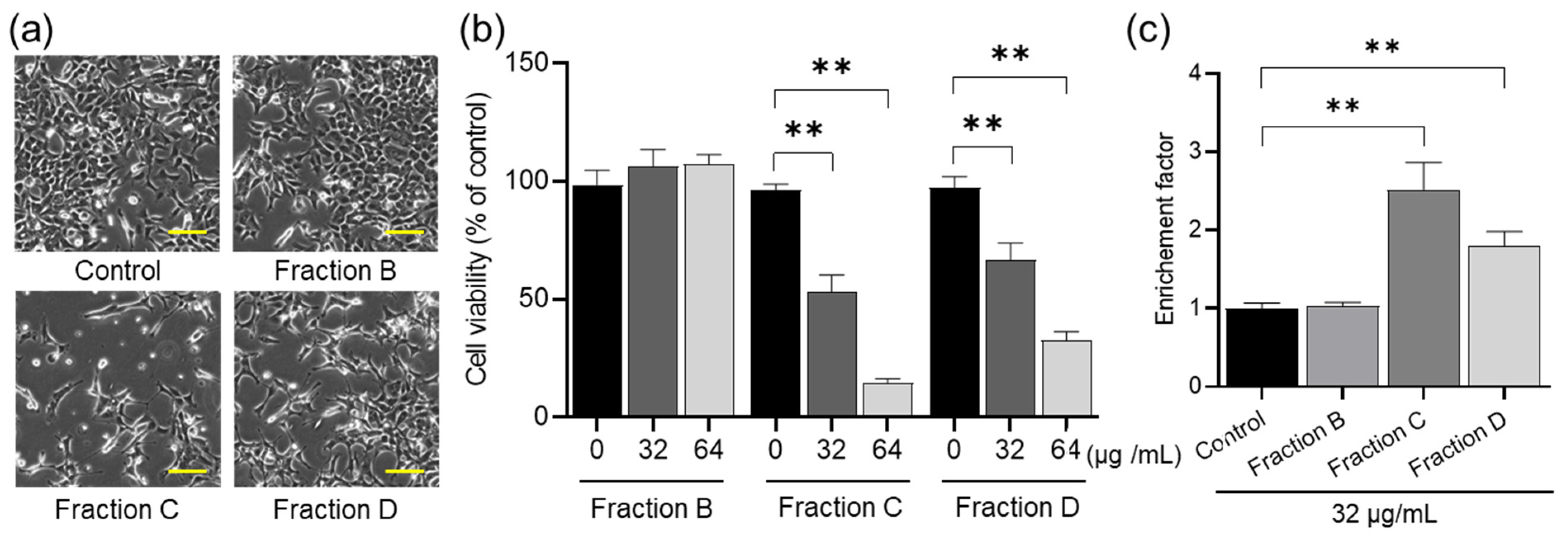
2.2. Antitumor Effects of Fractions B to D on HCT-116 Cells
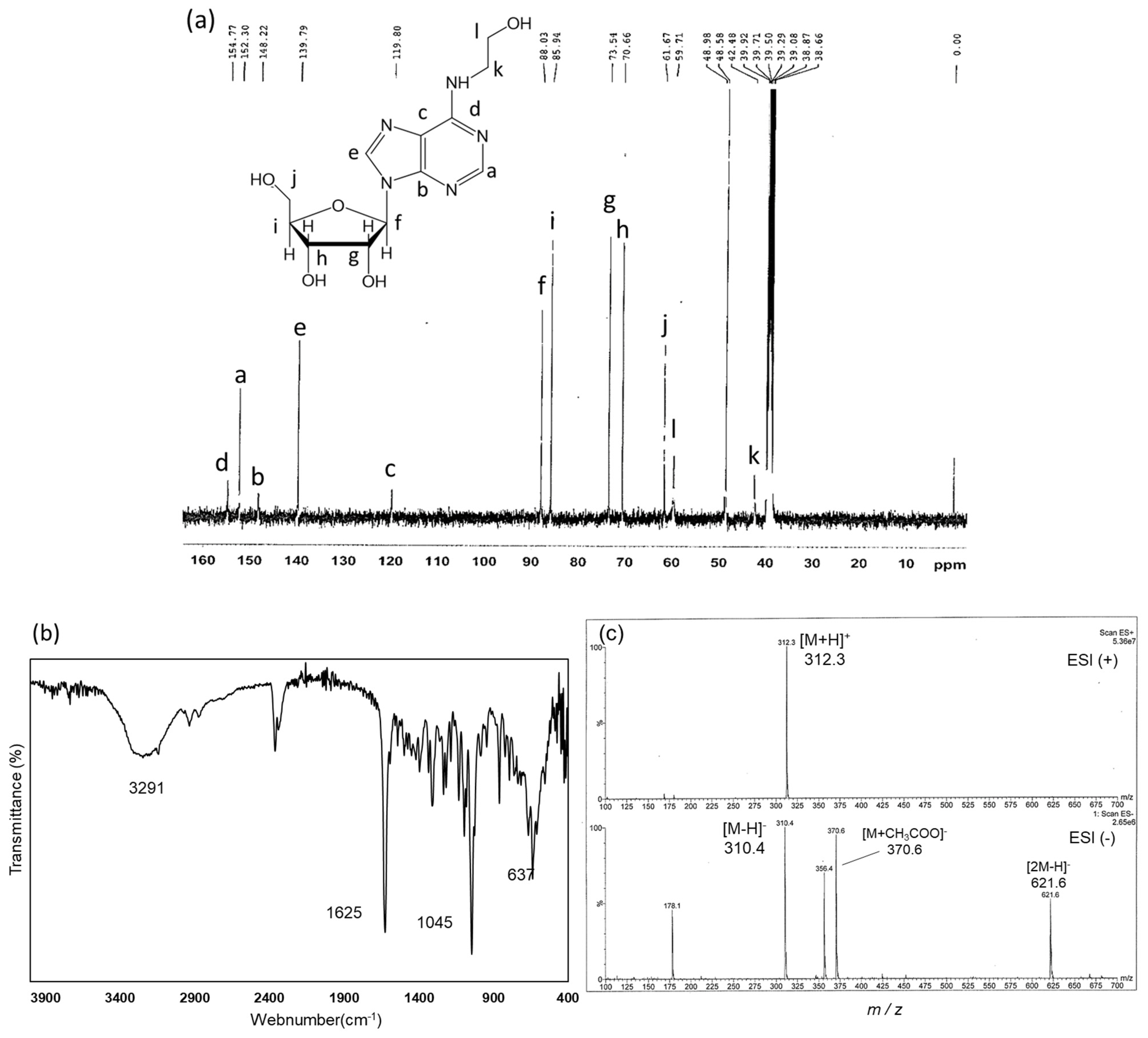
2.3. Structural Identification of Compounds in Fractions B, C, and D
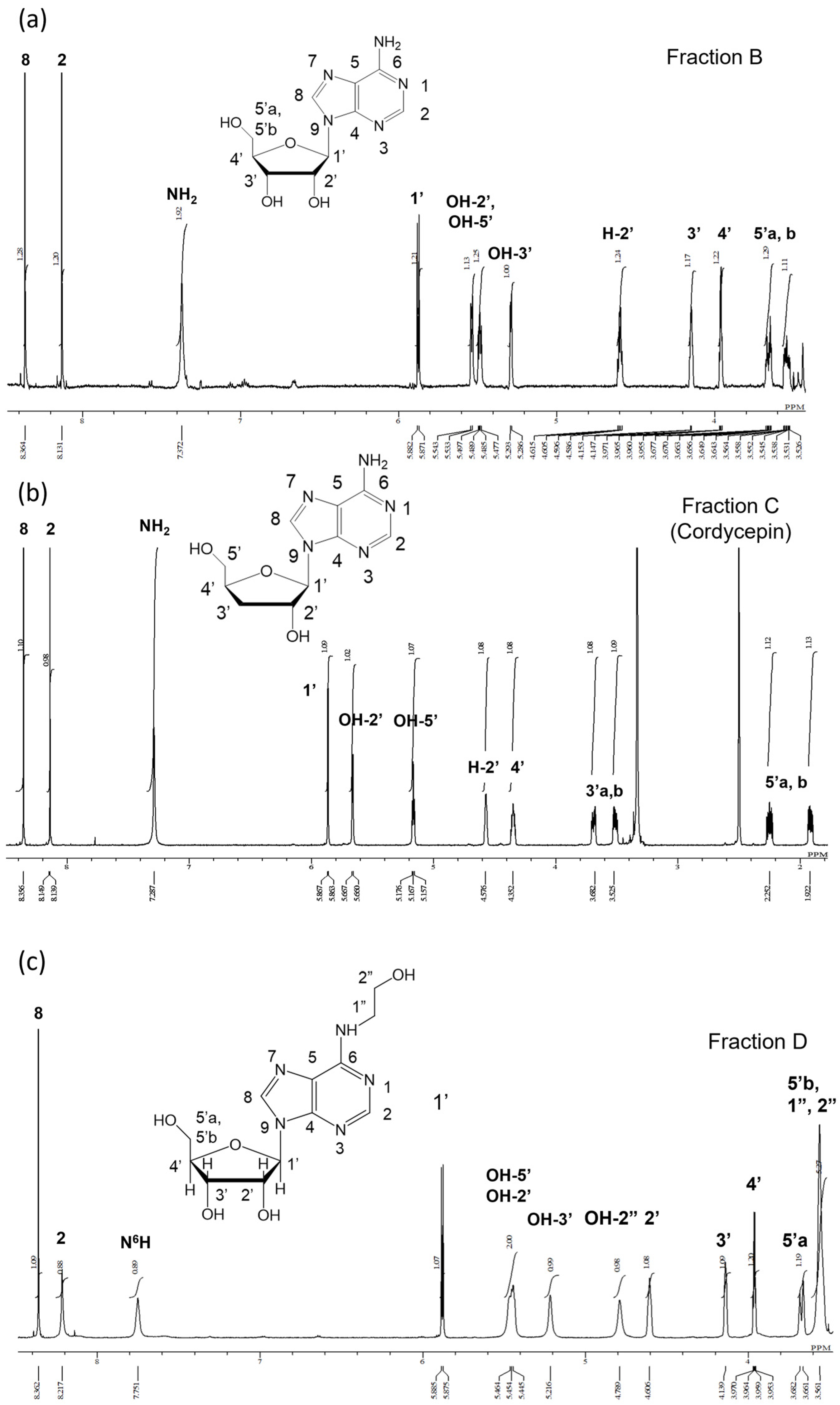
2.3.1. Identification of Compounds in Fractions B, C, and D using 1H-NMR
2.3.2. Structural Analysis of Fraction D Using 13C-NMR, FT-IR, and ESI-MS
2.4. Interaction between Human Adenosine A3 Receptors (A3ARs) and Adenosine, Cordycepin, and HEA
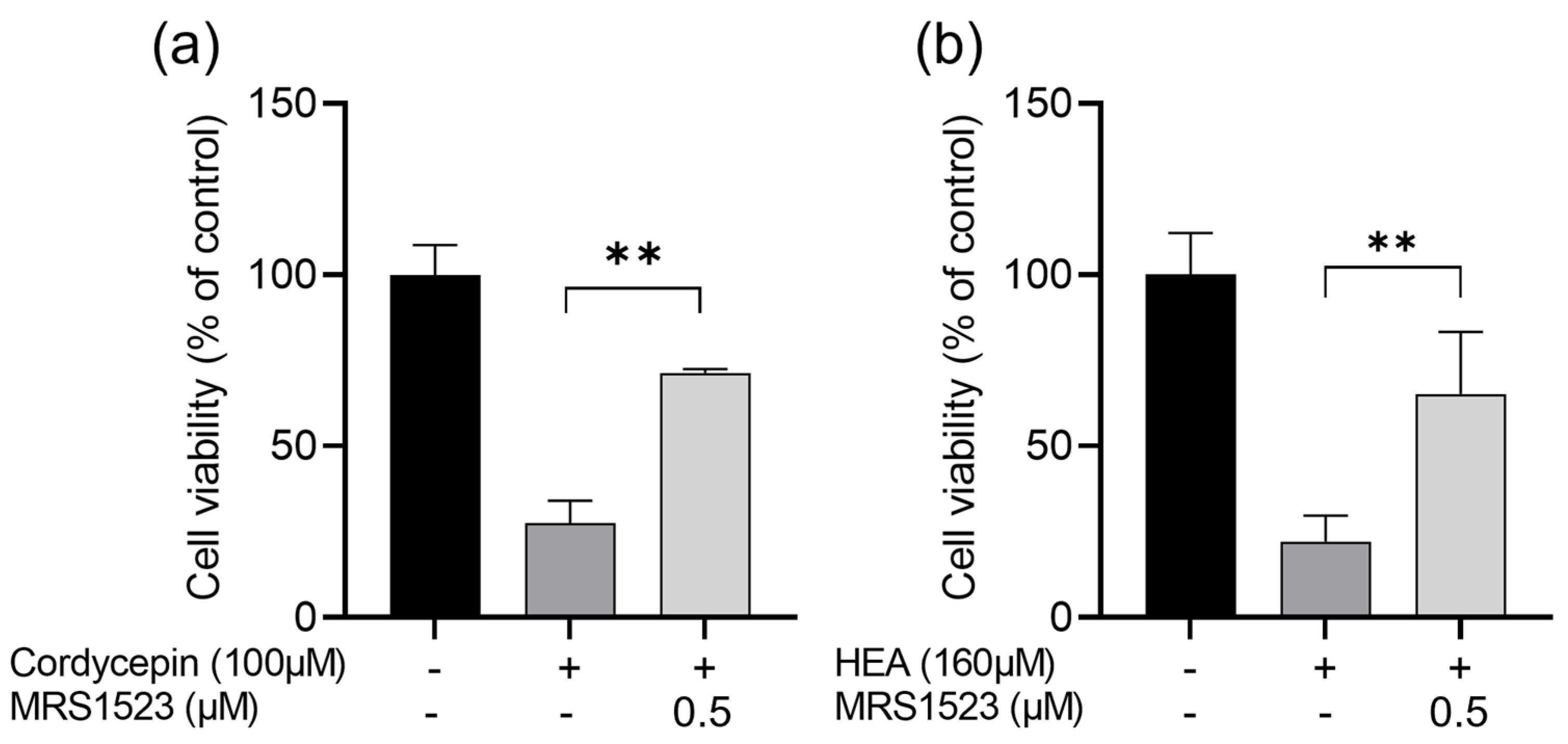
2.4.1. Cordycepin and HEA Inhibit HCT-116 Cell Proliferation as A3AR Agonists
2.4.2. Prediction of Binding Site and Energy on A3AR via MOE-ASEDock
3. Discussion
3.1. Effect of Soaking Time and Storage Period on Bioactivity of Shochu Spirits
3.2. Interaction of Adenosine, Cordycepin, and HEA with A3ARs
3.2.1. The Relationship between Cordycepin or HEA and A3ARs in Inhibiting Cell Proliferation
3.2.2. In Silico Analysis of the Interaction of A3AR with Cordycepin and HEA
3.3. Differences in Proliferation Inhibition by Adenosine, Cordycepin, and HEA in HL-60 and HCT-116 Cells
4. Materials and Methods
4.1. Reagents
4.2. Preparation of Cordyceps Militaris-Infused Spirits in Sweet Potato Shochu and Fractionation of Their Extracts
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. Identification of Chemical Compounds in Spirit Extract Fraction
4.6. ELISA Detection of DNA Fragmentation
4.7. Adenosine Receptor Antagonist Assay
4.8. In Silico Simulation of the Interaction between Adenosine, Cordycepin, and HEA and Human A3ARs
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rodríguez-Solana, R.; Vázquez-Araújo, L.; Salgado, J.M.; Domínguez, J.M.; Cortés-Diéguez, S. Optimization of the process of aromatic and medicinal plant maceration in grape marc distillates to obtain herbal liqueurs and spirits. J. Sci. Food Agric. 2016, 96, 4760–4771. [Google Scholar] [CrossRef] [PubMed]
- Senica, M.; Mikulic-Petkovsek, M. Changes in beneficial bioactive compounds in eight traditional herbal liqueurs during a one-month maceration process. J. Sci. Food Agric. 2020, 100, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Niyomvong, N.; Trakunjae, C.; Boondaeng, A. Fermentation Characteristics and Aromatic Profiles of Plum Wines Produced with Hanseniaspora thailandica Zal1 and Common Wine Yeasts. Molecules 2023, 28, 3009. [Google Scholar] [CrossRef] [PubMed]
- Yahagi, N. Illustrated catalogue of Japanese Cordyceps (entomogenous fungi): The Yahagi collection of Japanese Cordyceps stored in the Tohoku University Museum. Bull. Tohoku Univ. Mus. 2008, 8, 29–89. [Google Scholar]
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-Ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [PubMed]
- Hur, H. Chemical Ingredients of Cordyceps militaris. Mycobiology 2008, 36, 233–235. [Google Scholar] [CrossRef] [PubMed]
- Jędrejko, K.J.; Lazur, J.; Muszyńska, B. Cordyceps militaris: An Overview of Its Chemical Constituents in Relation to Biological Activity. Foods 2021, 10, 2634. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Wang, L.; Zhang, H.; Zhou, C.; Zhao, Y. Isolation, purification and identification of polysaccharides from cultured Cordyceps militaris. Fitoterapia 2004, 75, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.R. Cordyceps: A traditional Chinese medicine and another fungal therapeutic biofactory? Phytochemistry 2008, 69, 1469–1495. [Google Scholar] [CrossRef]
- Choi, S.B.; Park, C.H.; Choi, M.K.; Jun, D.W.; Park, S. Improvement of insulin resistance and insulin secretion by water extracts of Cordyceps militaris, Phellinus linteus, and Paecilomyces tenuipes in 90% pancreatectomized rats. Biosci. Biotechnol. Biochem. 2004, 68, 2257–2264. [Google Scholar] [CrossRef]
- Kim, H.G.; Shrestha, B.; Lim, S.Y.; Yoon, D.H.; Chang, W.C.; Shin, D.J.; Han, S.K.; Park, S.M.; Park, J.H.; Park, H.I.; et al. Cordycepin inhibits lipopolysaccharide-induced inflammation by the suppression of NF-kappaB through Akt and p38 inhibition in RAW 264.7 macrophage cells. Eur. J. Pharmacol. 2006, 545, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.; Ingalhalli, R. Cordyceps militaris (L.: Fr.) Link—An Important Medicinal Mushroom. J. Pharmacogn. Phytochem. 2013, 2, 315–319. [Google Scholar]
- Sho, C.; Sakao, K.; Ono, T.; Takase, Y.; Iwai, K.; Hou, D.-X. Induction of apoptosis of human colon carcinoma cells by an extract of Cordyceps militaris soaked in sweetpotato shochu subtitle_in_Japanese. J. Brew. Soc. Jpn. 2015, 110, 110–116. [Google Scholar] [CrossRef]
- Sho, C.; Sakao, K.; Yano, S.; Hisanaga, A.; Takase, Y.; Iwai, K.; Hou, D.-X. Anticancer activity of an extract of Cordyceps militaris soaked in sweetpotato shochu in human leukemia cells. subtitle_in_Japanese. J. Brew. Soc. Jpn. 2015, 110, 444–452. [Google Scholar] [CrossRef]
- Boison, D.; Yegutkin, G.G. Adenosine Metabolism: Emerging Concepts for Cancer Therapy. Cancer Cell 2019, 36, 582–596. [Google Scholar] [CrossRef] [PubMed]
- Mazziotta, C.; Rotondo, J.C.; Lanzillotti, C.; Campione, G.; Martini, F.; Tognon, M. Cancer biology and molecular genetics of A(3) adenosine receptor. Oncogene 2022, 41, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Madi, L.; Ochaion, A.; Rath-Wolfson, L.; Bar-Yehuda, S.; Erlanger, A.; Ohana, G.; Harish, A.; Merimski, O.; Barer, F.; Fishman, P. The A3 adenosine receptor is highly expressed in tumor versus normal cells: Potential target for tumor growth inhibition. Clin. Cancer Res. 2004, 10, 4472–4479. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Yoshikawa, N.; Yamaguchi, Y.; Kagota, S.; Shinozuka, K.; Kunitomo, M. Antitumor effect of cordycepin (3′-deoxyadenosine) on mouse melanoma and lung carcinoma cells involves adenosine A3 receptor stimulation. Anticancer. Res. 2006, 26, 43–47. [Google Scholar] [PubMed]
- Yoshikawa, N.; Yamada, S.; Takeuchi, C.; Kagota, S.; Shinozuka, K.; Kunitomo, M.; Nakamura, K. Cordycepin (3′-deoxyadenosine) inhibits the growth of B16-BL6 mouse melanoma cells through the stimulation of adenosine A3 receptor followed by glycogen synthase kinase-3beta activation and cyclin D1 suppression. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2008, 377, 591–595. [Google Scholar] [CrossRef]
- Yoshikawa, N.; Nishiuchi, A.; Kubo, E.; Yamaguchi, Y.; Kunitomo, M.; Kagota, S.; Shinozuka, K.; Nakamura, K. Cordyceps sinensis Acts As an Adenosine A3 Receptor Agonist on Mouse Melanoma and Lung Carcinoma Cells, and Human Fibrosarcoma and Colon Carcinoma Cells. Pharmacol. Pharm. 2011, 02, 266–270. [Google Scholar] [CrossRef]
- Cao, H.L.; Liu, Z.J.; Chang, Z. Cordycepin induces apoptosis in human bladder cancer cells via activation of A3 adenosine receptors. Tumour Biol. 2017, 39, 1010428317706915. [Google Scholar] [CrossRef]
- Zhang, Z.; Tudi, T.; Liu, Y.; Zhou, S.; Feng, N.; Yang, Y.; Tang, C.; Tang, Q.; Zhang, J. Preparative isolation of cordycepin, N(6)-(2-hydroxyethyl)-adenosine and adenosine from Cordyceps militaris by macroporous resin and purification by recycling high-speed counter-current chromatography. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2016, 1033–1034, 218–225. [Google Scholar] [CrossRef]
- Ciuffreda, P.; Casati, S.; Manzocchi, A. Complete (1)H and (13)C NMR spectral assignment of alpha- and beta-adenosine, 2′-deoxyadenosine and their acetate derivatives. Magn. Reson. Chem. 2007, 45, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Masuda, M.; Hatashita, M.; Fujihara, S.; Suzuki, Y.; Sakurai, A. Simple and efficient isolation of cordycepin from culture broth of a Cordyceps militaris mutant. J. Biosci. Bioeng. 2015, 120, 732–735. [Google Scholar] [CrossRef]
- Meng, Z.; Kang, J.; Wen, T.; Lei, B.; Hyde, K.D. Cordycepin and N6-(2-hydroxyethyl)-adenosine from Cordyceps pruinosa and their interaction with human serum albumin. PLoS ONE 2015, 10, e0121669. [Google Scholar] [CrossRef] [PubMed]
- Fishman, P.; Bar-Yehuda, S.; Farbstein, T.; Barer, F.; Ohana, G. Adenosine acts as a chemoprotective agent by stimulating G-CSF production: A role for A1 and A3 adenosine receptors. J. Cell. Physiol. 2000, 183, 393–398. [Google Scholar] [CrossRef]
- Fishman, P.; Bar-Yehuda, S.; Ohana, G.; Pathak, S.; Wasserman, L.; Barer, F.; Multani, A.S. Adenosine acts as an inhibitor of lymphoma cell growth: A major role for the A3 adenosine receptor. Eur. J. Cancer 2000, 36, 1452–1458. [Google Scholar] [CrossRef]
- Quan, N.V.; Iuchi, Y.; Anh, L.H.; Hasan, M.; Xuan, T.D. Simple Isolation of Cordycepin from Cordyceps militaris by Dual-Normal Phase Column Chromatography and Its Potential for Making Kombucha Functional Products. Separations 2022, 9, 290. [Google Scholar] [CrossRef]
- Sokol-Letowska, A.; Kucharska, A.Z.; Szumny, A.; Winska, K.; Nawirska-Olszanska, A. Phenolic Composition Stability and Antioxidant Activity of Sour Cherry Liqueurs. Molecules 2018, 23, 2156. [Google Scholar] [CrossRef]
- Kallithraka, S.; Salacha, M.I.; Tzourou, I. Changes in phenolic composition and antioxidant activity of white wine during bottle storage: Accelerated browning test versus bottle storage. Food Chem. 2009, 113, 500–505. [Google Scholar] [CrossRef]
- Sokół-Łętowska, A.; Kucharska, A.Z.; Wińska, K.; Szumny, A.; Nawirska-Olszańska, A.; Mizgier, P.; Wyspiańska, D. Composition and antioxidant activity of red fruit liqueurs. Food Chem. 2014, 157, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Li, X.; Yang, W.; Yu, L.; Jiang, X.; Chen, Y.; Shen, Z.; Li, C.; Gu, M.; Shi, L. N6-(2-hydroxyethyl)-Adenosine Induces Apoptosis via ER Stress and Autophagy of Gastric Carcinoma Cells In Vitro and In Vivo. Int. J. Mol. Sci. 2020, 21, 5815. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, K.A.; Gao, Z.G. Adenosine. In Encyclopedia of Neuroscience; Squire, L.R., Ed.; Academic Press: Oxford, UK, 2009; pp. 83–95. [Google Scholar]
- Ohana, G.; Bar-Yehuda, S.; Arich, A.; Madi, L.; Dreznick, Z.; Rath-Wolfson, L.; Silberman, D.; Slosman, G.; Fishman, P. Inhibition of primary colon carcinoma growth and liver metastasis by the A3 adenosine receptor agonist CF101. Br. J. Cancer 2003, 89, 1552–1558. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, V.P.; Griffith, M.T.; Hanson, M.A.; Cherezov, V.; Chien, E.Y.; Lane, J.R.; Ijzerman, A.P.; Stevens, R.C. The 2.6 angstrom crystal structure of a human A2A adenosine receptor bound to an antagonist. Science 2008, 322, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Glukhova, A.; Thal, D.M.; Nguyen, A.T.; Vecchio, E.A.; Jörg, M.; Scammells, P.J.; May, L.T.; Sexton, P.M.; Christopoulos, A. Structure of the Adenosine A(1) Receptor Reveals the Basis for Subtype Selectivity. Cell 2017, 168, 867–877.e813. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.-G.; Blaustein, J.B.; Gross, A.S.; Melman, N.; Jacobson, K.A. N6-Substituted adenosine derivatives: Selectivity, efficacy, and species differences at A3 adenosine receptors. Biochem. Pharmacol. 2003, 65, 1675–1684. [Google Scholar] [CrossRef] [PubMed]
- Ciancetta, A.; Jacobson, K.A. Structural Probing and Molecular Modeling of the A3 Adenosine Receptor: A Focus on Agonist Binding. Molecules 2017, 22, 449. [Google Scholar] [CrossRef]
- Lu, J.J.; Meng, L.H.; Cai, Y.J.; Chen, Q.; Tong, L.J.; Lin, L.P.; Ding, J. Dihydroartemisinin induces apoptosis in HL-60 leukemia cells dependent of iron and p38 mitogen-activated protein kinase activation but independent of reactive oxygen species. Cancer Biol. Ther. 2008, 7, 1017–1023. [Google Scholar] [CrossRef]
- Katsube, N.; Iwashita, K.; Tsushida, T.; Yamaki, K.; Kobori, M. Induction of apoptosis in cancer cells by Bilberry (Vaccinium myrtillus) and the anthocyanins. J. Agric. Food Chem. 2003, 51, 68–75. [Google Scholar] [CrossRef]
- Hatae, N.; Nakamura, J.; Okujima, T.; Ishikura, M.; Abe, T.; Hibino, S.; Choshi, T.; Okada, C.; Yamada, H.; Uno, H.; et al. Effect of the orthoquinone moiety in 9,10-phenanthrenequinone on its ability to induce apoptosis in HCT-116 and HL-60 cells. Bioorg Med. Chem. Lett. 2013, 23, 4637–4640. [Google Scholar] [CrossRef]
- Aplin, A.E.; Howe, A.K.; Juliano, R.L. Cell adhesion molecules, signal transduction and cell growth. Curr. Opin. Cell Bio 1999, 11, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Hay, B.A. Cell proliferation and apoptosis. Curr. Opin. Cell Bio 1999, 11, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Gessi, S.; Cattabriga, E.; Avitabile, A.; Gafa, R.; Lanza, G.; Cavazzini, L.; Bianchi, N.; Gambari, R.; Feo, C.; Liboni, A.; et al. Elevated expression of A3 adenosine receptors in human colorectal cancer is reflected in peripheral blood cells. Clin. Cancer Res. 2004, 10, 5895–5901. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakao, K.; Sho, C.; Miyata, T.; Takara, K.; Oda, R.; Hou, D.-X. Verification of In Vitro Anticancer Activity and Bioactive Compounds in Cordyceps Militaris-Infused Sweet Potato Shochu Spirits. Molecules 2024, 29, 2119. https://doi.org/10.3390/molecules29092119
Sakao K, Sho C, Miyata T, Takara K, Oda R, Hou D-X. Verification of In Vitro Anticancer Activity and Bioactive Compounds in Cordyceps Militaris-Infused Sweet Potato Shochu Spirits. Molecules. 2024; 29(9):2119. https://doi.org/10.3390/molecules29092119
Chicago/Turabian StyleSakao, Kozue, Cho Sho, Takeshi Miyata, Kensaku Takara, Rio Oda, and De-Xing Hou. 2024. "Verification of In Vitro Anticancer Activity and Bioactive Compounds in Cordyceps Militaris-Infused Sweet Potato Shochu Spirits" Molecules 29, no. 9: 2119. https://doi.org/10.3390/molecules29092119
APA StyleSakao, K., Sho, C., Miyata, T., Takara, K., Oda, R., & Hou, D.-X. (2024). Verification of In Vitro Anticancer Activity and Bioactive Compounds in Cordyceps Militaris-Infused Sweet Potato Shochu Spirits. Molecules, 29(9), 2119. https://doi.org/10.3390/molecules29092119