
A Rare Mono-Rhamnolipid Congener Efficiently Produced by Recombinant Pseudomonas aeruginosa YM4 via the Expression of Global Transcriptional Regulator irrE

Abstract
1. Introduction
2. Results
2.1. Screening for Suitable Promoters
2.2. Effects of the Global Transcriptional Regulator IrrE on Growth and Rhamnolipds Titer in Batch Fermentation
2.3. IrrE of D. radiodurans Improved the Robustness of P. aeruginosa
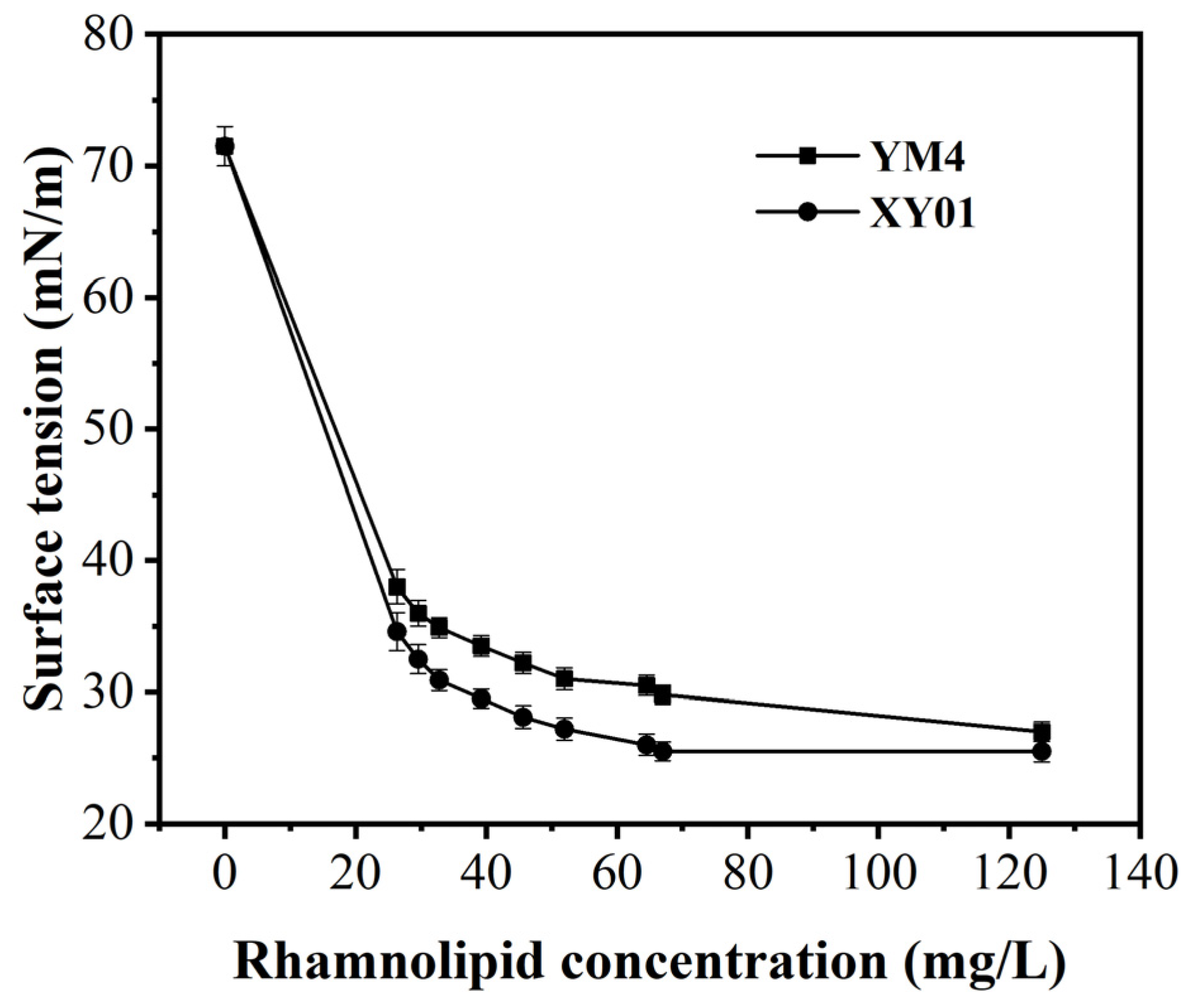
2.4. Characterization of Rhamnolipids
2.4.1. HPLC-ELSD Analysis of Rhamnolipids
2.4.2. HPLC-MS Analysis of Rhamnolipids
2.4.3. Fourier-Transform Infrared (FTIR) Analysis of Rhamnolipids
2.4.4. The Critical Micelle Concentration (CMC) of Rhamnolipids
2.4.5. Emulsifying Activity of Rhamnolipids on Diesel Fuel
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strain, Medium, and Culture Conditions
5.2. Construction of Reporter Plasmid
5.3. Promoter Strength Measurements
5.4. Construction of PrhlA-irrE Recombinant Plasmid
5.5. Extraction of Rhamnolipids and Determination of Strain Growth Rate
5.6. Effect of IrrE on the Stress Tolerance of P. aeruginosa Cells
5.7. Quantitative Analysis of Rhamnolipids
5.8. Characterization of Rhamnolipids
5.8.1. HPLC-MS Analysis of Rhamnolipids
5.8.2. Fourier-Transform Infrared (FTIR) Analysis of Rhamnolipids
5.8.3. Determination of the Critical Micelle Concentration (CMC)
5.8.4. Determination of the Emulsification Index (EI24)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nguyen, T.T.; Sabatini, D.A. Characterization and emulsification properties of rhamnolipid and sophorolipid biosurfactants and their applications. Int. J. Mol. Sci. 2011, 12, 1232–1244. [Google Scholar] [CrossRef]
- Sinumvayo, J.P.; Ishimwe, N. Agriculture and food applications of rhamnolipids and its production by Pseudomonas aeruginosa. J. Chem. Eng. Process Technol. 2015, 6, 223. [Google Scholar] [CrossRef]
- Piljac, A.; Stipčević, T.; Piljac-Žegarac, J.; Piljac, G. Successful treatment of chronic decubitus ulcer with 0.1% dirhamnolipid ointment. J. Cutan. Med. Surg. 2008, 12, 142–146. [Google Scholar] [CrossRef]
- Sharma, R.; Singh, J.; Verma, N. Optimization of rhamnolipid production from Pseudomonas aeruginosa PBS towards application for microbial enhanced oil recovery. 3 Biotech 2018, 8, 20. [Google Scholar] [CrossRef]
- Palos Pacheco, R.; Kegel, L.L.; Pemberton, J.E. Interfacial and Solution Aggregation Behavior of a Series of Bioinspired Rhamnolipid Congeners Rha–C14–C x (x = 6, 8, 10, 12, 14). J. Phys. Chem. B 2021, 125, 13585–13596. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Lai, L.; Lu, Q.; Mei, P.; Wang, Y.; Cheng, L.; Liu, Y. Comparative studies on the surface/interface properties and aggregation behavior of mono-rhamnolipid and di-rhamnolipid. Colloids Surf. B Biointerfaces 2019, 181, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Nitschke, M.; Costa, S.G.; Contiero, J. Structure and applications of a rhamnolipid surfactant produced in soybean oil waste. Appl. Biochem. Biotechnol. 2010, 160, 2066–2074. [Google Scholar] [CrossRef]
- Cabrera-Valladares, N.; Richardson, A.-P.; Olvera, C.; Treviño, L.G.; Déziel, E.; Lépine, F.; Soberón-Chávez, G. Monorhamnolipids and 3-(3-hydroxyalkanoyloxy) alkanoic acids (HAAs) production using Escherichia coli as a heterologous host. Appl. Microbiol. Biotechnol. 2006, 73, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Wittgens, A.; Tiso, T.; Arndt, T.T.; Wenk, P.; Hemmerich, J.; Müller, C.; Wichmann, R.; Küpper, B.; Zwick, M.; Wilhelm, S. Growth independent rhamnolipid production from glucose using the non-pathogenic Pseudomonas putida KT2440. Microb. Cell Factories 2011, 10, 80. [Google Scholar] [CrossRef]
- Invally, K.; Sancheti, A.; Ju, L.-K. A new approach for downstream purification of rhamnolipid biosurfactants. Food Bioprod. Process. 2019, 114, 122–131. [Google Scholar] [CrossRef]
- Bazsefidpar, S.; Mokhtarani, B.; Panahi, R.; Hajfarajollah, H. Overproduction of rhamnolipid by fed-batch cultivation of Pseudomonas aeruginosa in a lab-scale fermenter under tight DO control. Biodegradation 2019, 30, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Herzog, M.; Tiso, T.; Blank, L.M.; Winter, R. Interaction of rhamnolipids with model biomembranes of varying complexity. Biochim. Biophys. Acta (BBA)-Biomembr. 2020, 1862, 183431. [Google Scholar] [CrossRef]
- Angelova, B.; Schmauder, H.-P. Lipophilic compounds in biotechnology—Interactions with cells and technological problems. J. Biotechnol. 1999, 67, 13–32. [Google Scholar] [CrossRef] [PubMed]
- Andrä, J.; Rademann, J.; Howe, J.; Koch, M.H.; Heine, H.; Zähringer, U.; Brandenburg, K. Endotoxin-like properties of a rhamnolipid exotoxin from Burkholderia (Pseudomonas) plantarii: Immune cell stimulation and biophysical characterization. Biol. Chem. 2006, 387, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Gao, G.; Xu, G.; Fan, L.; Yin, L.; Shen, B.; Hua, Y. Deinococcus radiodurans PprI switches on DNA damage response and cellular survival networks after radiation damage. Mol. Cell. Proteom. 2009, 8, 481–494. [Google Scholar] [CrossRef]
- Hua, Y.; Narumi, I.; Gao, G.; Tian, B.; Satoh, K.; Kitayama, S.; Shen, B. PprI: A general switch responsible for extreme radioresistance of Deinococcus radiodurans. Biochem. Biophys. Res. Commun. 2003, 306, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhu, J.; Zhu, X.; Song, M.; Zhang, T.; Xin, F.; Dong, W.; Ma, J.; Jiang, M. Expression of global regulator IrrE for improved succinate production under high salt stress by Escherichia coli. Bioresour. Technol. 2018, 254, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Zhang, Y.; Suo, Y.; Liao, Z.; Ma, Y.; Fu, H.; Wang, J. The global regulator IrrE from Deinococcus radiodurans enhances the furfural tolerance of Saccharomyces cerevisiae. Biochem. Eng. J. 2018, 136, 69–77. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, R.; Zhao, Z.; Zhou, Z.; Lu, W.; Zhang, W.; Chen, M. IrrE, an exogenous gene from Deinococcus radiodurans, improves the growth of and ethanol production by a Zymomonas mobilis strain under ethanol and acid stresses. J. Microbiol. Biotechnol. 2010, 20, 1156–1162. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, M.; Xu, Z.; Xu, H.; Li, S. Construction of a robust Sphingomonas sp. strain for welan gum production via the expression of global transcriptional regulator IrrE. Front. Bioeng. Biotechnol. 2020, 8, 674. [Google Scholar] [CrossRef]
- Song, B.; Zhou, Q.; Xue, H.-J.; Liu, J.-J.; Zheng, Y.-Y.; Shen, Y.-B.; Wang, M.; Luo, J.-M. IrrE improves organic solvent tolerance and Δ1-dehydrogenation productivity of Arthrobacter simplex. J. Agric. Food Chem. 2018, 66, 5210–5220. [Google Scholar] [CrossRef] [PubMed]
- Xu, A.; Zhang, X.; Cao, S.; Zhou, X.; Yu, Z.; Qian, X.; Zhou, J.; Dong, W.; Jiang, M. Transcription-associated fluorescence-activated droplet sorting for Di-rhamnolipid hyperproducers. ACS Synth. Biol. 2022, 11, 1992–2000. [Google Scholar] [CrossRef] [PubMed]
- Wittgens, A.; Rosenau, F. On the road towards tailor-made rhamnolipids: Current state and perspectives. Appl. Microbiol. Biotechnol. 2018, 102, 8175–8185. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Y.; Lin, J.; Wang, W.; Li, S. High-yield di-rhamnolipid production by Pseudomonas aeruginosa YM4 and its potential application in MEOR. Molecules 2019, 24, 1433. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Tao, W.; Yu, D.; Li, S. Emulsifying Properties of Rhamnolipids and Their In Vitro Antifungal Activity against Plant Pathogenic Fungi. Molecules 2022, 27, 7746. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wang, J.; Yang, R.; Li, J.; Lin, M.; Lin, Z. Laboratory-evolved mutants of an exogenous global regulator, IrrE from Deinococcus radiodurans, enhance stress tolerances of Escherichia coli. PLoS ONE 2011, 6, e16228. [Google Scholar] [CrossRef]
- Vujičić-Žagar, A.; Dulermo, R.; Le Gorrec, M.; Vannier, F.; Servant, P.; Sommer, S.; de Groot, A.; Serre, L. Crystal structure of the IrrE protein, a central regulator of DNA damage repair in Deinococcaceae. J. Mol. Biol. 2009, 386, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhou, Z.; Zhang, W.; Chen, Z.; Song, Y.; Lu, W.; Lin, M.; Chen, M. The site-directed A184S mutation in the HTH domain of the global regulator IrrE enhances Deinococcus radiodurans R1 tolerance to UV radiation and MMC shock. J. Microbiol. Biotechnol. 2015, 25, 2125–2134. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Wang, T.; Li, X.; Yang, Y.; Zhou, M.; Li, M.; Yan, Z. Enhancement of bioelectricity generation via heterologous expression of IrrE in Pseudomonas aeruginosa-inoculated MFCs. Biosens. Bioelectron. 2018, 117, 23–31. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, Y.; Zanaroli, G.; Wang, Z.; Xu, P.; Tang, H. Enhancing bioremediation potential of Pseudomonas putida by developing its acid stress tolerance with glutamate decarboxylase dependent system and global regulator of extreme radiation resistance. Front. Microbiol. 2019, 10, 2033. [Google Scholar] [CrossRef]
- Zhao, F.; Han, S.; Zhang, Y. Comparative studies on the structural composition, surface/interface activity and application potential of rhamnolipids produced by Pseudomonas aeruginosa using hydrophobic or hydrophilic substrates. Bioresour. Technol. 2020, 295, 122269. [Google Scholar] [CrossRef] [PubMed]
- Sodagari, M.; Invally, K.; Ju, L.-K. Maximize rhamnolipid production with low foaming and high yield. Enzym. Microb. Technol. 2018, 110, 79–86. [Google Scholar] [CrossRef]
- Zhao, P.; Zhou, Z.; Zhang, W.; Lin, M.; Chen, M.; Wei, G. Global transcriptional analysis of Escherichia coli expressing IrrE, a regulator from Deinococcus radiodurans, in response to NaCl shock. Mol. BioSystems 2015, 11, 1165–1171. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, W.; Chen, M.; Pan, J.; Lu, W.; Ping, S.; Yan, Y.; Hou, X.; Yuan, M.; Zhan, Y. Genome-wide transcriptome and proteome analysis of Escherichia coli expressing IrrE, a global regulator of Deinococcus radiodurans. Mol. Biosyst. 2011, 7, 1613–1620. [Google Scholar] [CrossRef] [PubMed]
- Tiso, T.; Zauter, R.; Tulke, H.; Leuchtle, B.; Li, W.-J.; Behrens, B.; Wittgens, A.; Rosenau, F.; Hayen, H.; Blank, L.M. Designer rhamnolipids by reduction of congener diversity: Production and characterization. Microb. Cell Factories 2017, 16, 225. [Google Scholar] [CrossRef]
- Zhao, F.; Yuan, M.; Lei, L.; Li, C.; Xu, X. Enhanced production of mono-rhamnolipid in Pseudomonas aeruginosa and application potential in agriculture and petroleum industry. Bioresour. Technol. 2021, 323, 124605. [Google Scholar] [CrossRef] [PubMed]
- Shatila, F.; Diallo, M.M.; Şahar, U.; Ozdemir, G.; Yalçın, H.T. The effect of carbon, nitrogen and iron ions on mono-rhamnolipid production and rhamnolipid synthesis gene expression by Pseudomonas aeruginosa ATCC 15442. Arch. Microbiol. 2020, 202, 1407–1417. [Google Scholar] [CrossRef]
- Sood, U.; Singh, D.N.; Hira, P.; Lee, J.-K.; Kalia, V.C.; Lal, R.; Shakarad, M. Rapid and solitary production of mono-rhamnolipid biosurfactant and biofilm inhibiting pyocyanin by a taxonomic outlier Pseudomonas aeruginosa strain CR1. J. Biotechnol. 2020, 307, 98–106. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Retention Time (min) | Structure | Molecular Weight | Relative Abundance (%) | ||||
|---|---|---|---|---|---|---|---|---|
| Standard * | YM4 | XY01 | XY02 | XY03 | ||||
| 1 | 4.87 | NA | NA | 2.99 | - | - | - | - |
| 2 | 5.53 | NA | NA | 3.11 | - | - | - | - |
| 3 | 10.56 | Rha2–C10–C10 | 650 | 46.52 | 58.72 | - | 56.14 | 57.89 |
| 4 | 13.28 | Rha–C10–C10 | 504 | 41.88 | 38.86 | - | 41.22 | 40.29 |
| 5 | 14.87 | NA | NA | 2.22 | 1.33 | - | 1.11 | 0.78 |
| 6 | 16.28 | Rha–C10–C12 | 532 | 2.11 | 0.64 | 94.14 | 0.52 | 0.58 |
| 7 | 19.04 | NA | NA | 1.17 | 0.45 | 1.19 | 1.01 | 0.37 |
| 8 | 11.95 | NA | NA | - | - | 2.89 | - | - |
| 9 | 22.33 | NA | NA | - | - | 0.78 | - | - |
| Strians | Genetic Characteristics | Carbon Source | Titer (g/L) | Mono-Rhamnolipid Congeners (Titer Ratio,%) | Reference |
|---|---|---|---|---|---|
| P. putida KT2440 | Expressing the rhlAB operon in P. putida KT2440 | Glucose | 2.4 | Rha–C8–C10; 2% Rha–C10–C10; 63% Rha–C10–C12:1; 16% Rha–C10–C12; 19% | [35] |
| P. aeruginosa SG ΔrhlC | Knocking out rhlC in P. aeruginosa SG | Glycerol | 14.22 | Rha–C10; 3.4% Rha–C8–C10; 10.9% Rha–C10–C10; 25.9% Rha–C10–C12:1; 43.5% Rha–C10–C12; 16.4% | [36] |
| P. aeruginosa ATCC 15442 | Natural deficiency of rhlC | Glucose | NA * | Rha–C8–C10; NA Rha–C10–C10; NA Rha–C10–C12:1; NA Rha–C10–C12; NA | [37] |
| P. aeruginosa CR1 | Natural deficiency of rhlC | Glycerol | 21.77 | Rha–C8–C10; NA Rha–C10–C10; NA Rha–C12:2–C10:1; NA | [38] |
| P. aeruginosa XY01 | Expressing irrE of D. radiodurans in P. aeruginosa YM4 | Soybean oil | 26 | Rha–C10–C12; 94.1% | This work |
| Strains and Plasmids | Characteristics | Sources |
|---|---|---|
| Strains | ||
| P. aeruginosa YM4 | Rhamnolipid-producing strain (original strain) | This lab |
| P. aeruginosa PAO1 | Source of promoter PrhlA | This lab |
| B. subtilis 168 (Pveg-gfp) | Source of gfp | This lab |
| E. coli TOP10 | Plasmid amplification in vivo | This lab |
| P. aeruginosa XY01 | Strain YM4 derivative; harboring pHERD20T-PrhlA-irrE01 | This work |
| P. aeruginosa XY02 | Strain YM4 derivative; harboring pHERD20T-PrhlA-irrE02 | This work |
| P. aeruginosa XY03 | Strain YM4 derivative; harboring pHERD20T-PrhlA-irrE03 | This work |
| Plasmids | ||
| pHERD20T | Broad host-range vector; pUCP20T Plac replaced with 1.3 kb AflⅡ-EcoRⅠ fragment of araC-PBAD cassette (5087 bp) | Provided by Dr. Xu Anming @ Nanjing Tech University |
| pHERD20T-PBAD-gfp | pHERD20T PBAD fused with gfp gene | This work |
| pHERD20T-PrhlA-gfp | Reporter plasmid; pHERD20T PBAD replaced with PrhlA and fused with gfp gene | This work |
| pHERD20T-PrhlA-irrE01 | pHERD20T PBAD replaced with PrhlA and fused with irrE gene from D. radiodurans | This work |
| pHERD20T-PrhlA-irrE02 | pHERD20T PBAD replaced with PrhlA and fused with irrE gene from D. gobiensis I-O | This work |
| pHERD20T-PrhlA-irrE03 | pHERD20T PBAD replaced with PrhlA and fused with irrE gene from D. deserti | This work |
| Premier ID | Sequence (5′→3′) |
|---|---|
| pHERD20T-F | AAGCTTAGCTTGGCACTGGCCGTCG |
| pHERD20T-R | CCCGGGCTATCGCCACCGTCG |
| PrhlA-F | TGGCGATAGCCCGGGCGCCAGAGCGTTTCGACA |
| PrhlA-R·gfp | CTCCTTTTGACATTTCACACCTCCCAAAAATTTTCGAACAG |
| gfp-F | TGGGAGGTGTGAAATGTCAAAAGGAGAAGAACTTTTTACAGGTG |
| gfp-R | CGGCCAGTGCCAAGCTAAGCTTTTATTTATAAAGTTCGTCCATACCGTG |
| PrhlA-R01 | TCGGCATTTCACACCTCCCAAAAATTTTCGAACAG |
| irrE01-F | TTTGGGAGGTGTGAAATGCCGAGCGCGAATGTTTCA |
| irrE01-R | TGCCAAGCTAAGCTTTCACTGTGCCGCATCTTGCGGTTCAT |
| PrhlA-R02 | TGCCAGTTCTCTCATTTCACACCTCCCAAAAATTTTCGAACAG |
| irrE02-F | AGGTGTGAAATGAGAGAACTGGCAGCGGCGTAT |
| irrE02-R | TGCCAAGCTAAGCTTTCATGTGCCTCCTCTGCCAT |
| PrhlA-R03 | CGGATCTGTCATTTCACACCTCCCAAAAATTTTCGAACAG |
| irrE03-F | TTTGGGAGGTGTGAAATGACAGATCCGGCGCC |
| irrE03-R | TGCCAAGCTAAGCTTTCAGCTCTGATCTCCCGGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Li, D.; Yue, S.; Yuan, Z.; Li, S. A Rare Mono-Rhamnolipid Congener Efficiently Produced by Recombinant Pseudomonas aeruginosa YM4 via the Expression of Global Transcriptional Regulator irrE. Molecules 2024, 29, 1992. https://doi.org/10.3390/molecules29091992
Wang X, Li D, Yue S, Yuan Z, Li S. A Rare Mono-Rhamnolipid Congener Efficiently Produced by Recombinant Pseudomonas aeruginosa YM4 via the Expression of Global Transcriptional Regulator irrE. Molecules. 2024; 29(9):1992. https://doi.org/10.3390/molecules29091992
Chicago/Turabian StyleWang, Xinying, Dongmei Li, Shenghui Yue, Zhangzhong Yuan, and Shuang Li. 2024. "A Rare Mono-Rhamnolipid Congener Efficiently Produced by Recombinant Pseudomonas aeruginosa YM4 via the Expression of Global Transcriptional Regulator irrE" Molecules 29, no. 9: 1992. https://doi.org/10.3390/molecules29091992
APA StyleWang, X., Li, D., Yue, S., Yuan, Z., & Li, S. (2024). A Rare Mono-Rhamnolipid Congener Efficiently Produced by Recombinant Pseudomonas aeruginosa YM4 via the Expression of Global Transcriptional Regulator irrE. Molecules, 29(9), 1992. https://doi.org/10.3390/molecules29091992