
Organic Sunscreens—Is Their Placenta Permeability the Only Issue Associated with Exposure During Pregnancy? In Silico Studies of Sunscreens’ Placenta Permeability and Interactions with Selected Placental Enzymes


Abstract
1. Introduction
2. Results and Discussion
2.1. Human Placenta Permeability—Qualitative Studies
2.2. Sunscreens’ Affinity for Selected Placenta Enzymes
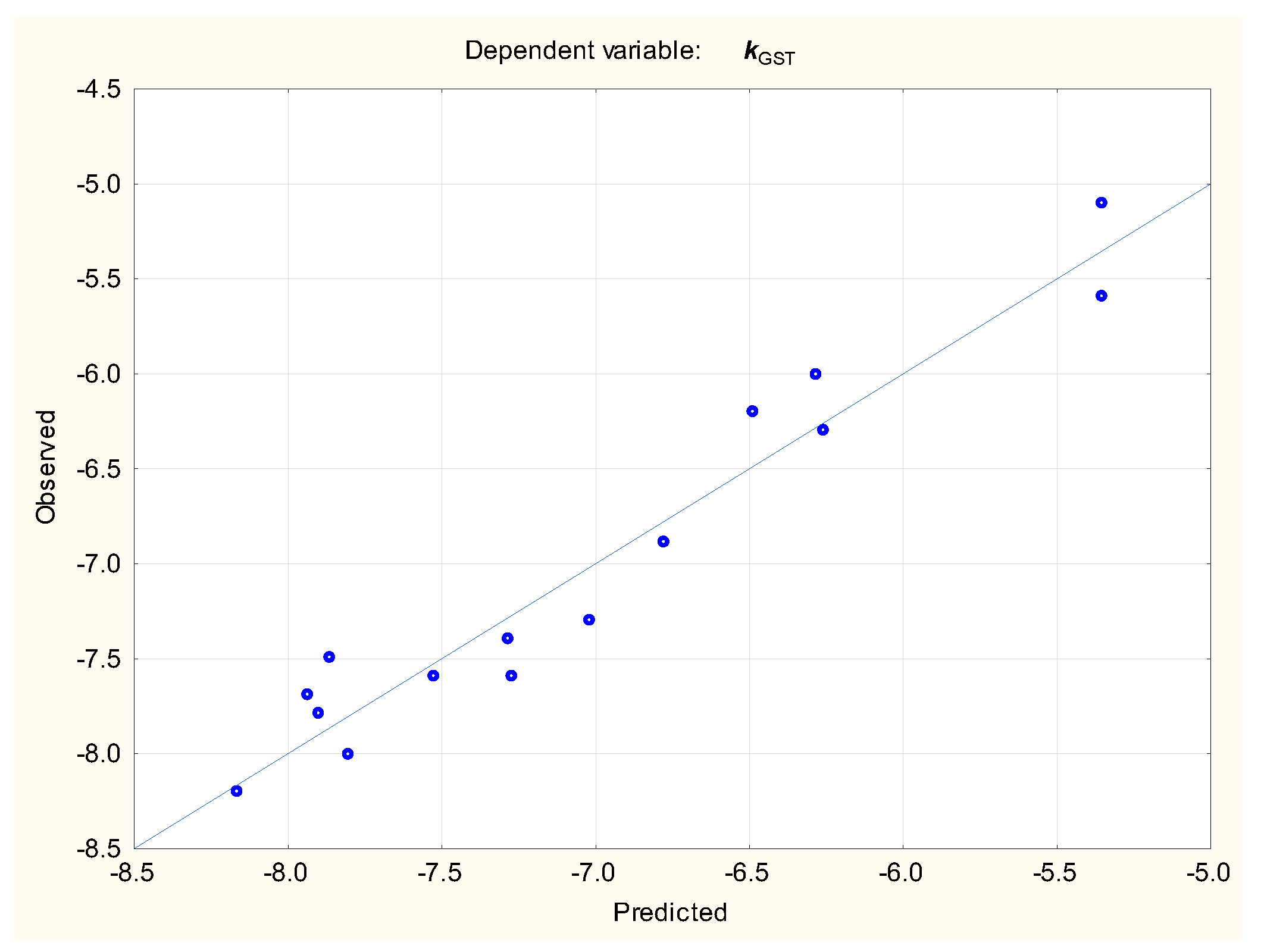
(n = 16, R2 = 0.943, R2adj. = 0.922, F = 45.12, p < 0.01, se = 0.260, RMSECV = 0.335)
(n = 16, R2 = 0.880, R2adj. = 0.851, F = 29.45, p < 0.01, se = 0.374, RMSECV = 0.388)
2.3. Clinical Relevance of Molecular Docking Ligand–Enzyme Affinity Studies
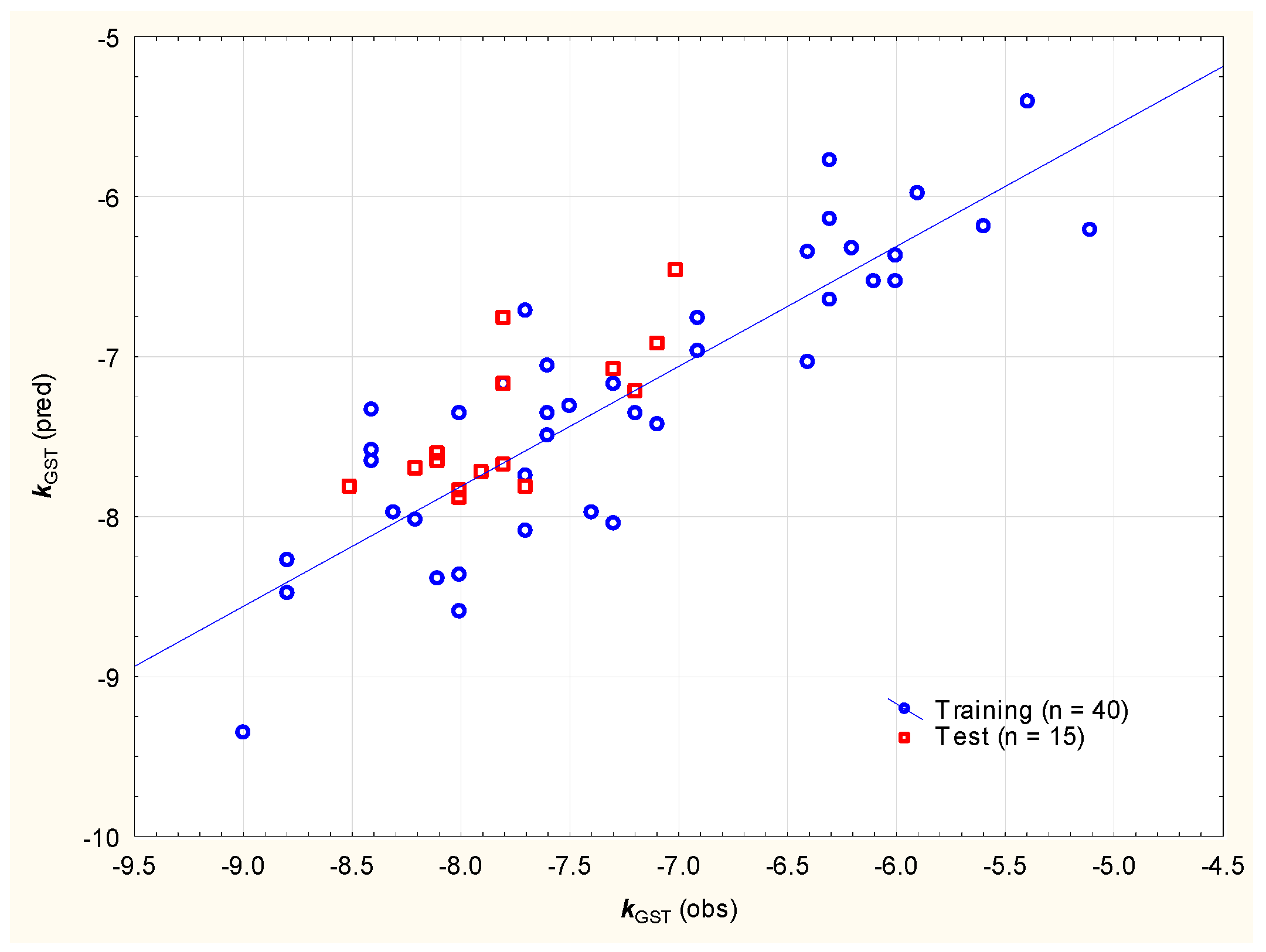
(n = 40, R2 = 0.751, R2adj. = 0.730, F = 36.1, p < 0.01, RMSEP = 0.453, RMSECV = 0.629)
(n = 40, R2 = 0.773, R2adj. = 0.754, F = 40.8, p < 0.01, RMSEP = 0.450, RMSECV = 0.804)
3. Materials and Methods
3.1. Compounds
3.2. Calculated Descriptors
3.3. Placenta Permeability Classification Models
3.4. Molecular Docking
3.5. Multiple Linear Regression Models (MLRs) of Ligands’ Affinity for GST and NAT2 Enzymes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsui, M.M.P.; Leung, H.W.; Wai, T.C.; Yamashita, N.; Taniyasu, S.; Liu, W.; Lam, P.K.S.; Murphy, M.B. Occurrence, distribution and ecological risk assessment of multiple classes of UV filters in surface waters from different countries. Water Res. 2014, 67, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Cruz, M.S.; Barceló, D. Chemical analysis and ecotoxicological effects of organic UV-absorbing compounds in aquatic ecosystems. TrAC—Trends Anal. Chem. 2009, 28, 708–717. [Google Scholar] [CrossRef]
- Paredes, E.; Perez, S.; Rodil, R.; Quintana, J.B.; Beiras, R. Ecotoxicological evaluation of four UV filters using marine organisms from different trophic levels Isochrysis galbana, Mytilus galloprovincialis, Paracentrotus lividus, and Siriella armata. Chemosphere 2014, 104, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Gago-Ferrero, P.; Alonso, M.B.; Bertozzi, C.P.; Marigo, J.; Barbosa, L.; Cremer, M.; Secchi, E.R.; Azevedo, A.; Lailson-Brito, J.; Torres, J.P.M.; et al. First determination of UV filters in marine mammals. Octocrylene levels in Franciscana dolphins. Environ. Sci. Technol. 2013, 47, 5619–5625. [Google Scholar] [CrossRef]
- Sobańska, A.W. Evaluation of drug-likeness and ADME properties of sunscreens and preservatives using reversed-phase thin layer chromatographic retention data and calculated descriptors. J. Pharm. Biomed. Anal. 2021, 201, 114126. [Google Scholar] [CrossRef]
- Huang, Y.; Law, J.C.F.; Lam, T.K.; Leung, K.S.Y. Risks of organic UV filters: A review of environmental and human health concern studies. Sci. Total Environ. 2021, 755, 142486. [Google Scholar] [CrossRef]
- León, Z.; Chisvert, A.; Tarazona, I.; Salvador, A. Solid-phase extraction liquid chromatography-tandem mass spectrometry analytical method for the determination of 2-hydroxy-4-methoxybenzophenone and its metabolites in both human urine and semen. Anal. Bioanal. Chem. 2010, 398, 831–843. [Google Scholar] [CrossRef]
- Schlumpf, M.; Kypke, K.; Wittassek, M.; Angerer, J.; Mascher, H.; Mascher, D.; Vökt, C.; Birchler, M.; Lichtensteiger, W. Exposure patterns of UV filters, fragrances, parabens, phthalates, organochlor pesticides, PBDEs, and PCBs in human milk: Correlation of UV filters with use of cosmetics. Chemosphere 2010, 81, 1171–1183. [Google Scholar] [CrossRef]
- Sunyer-Caldu, A.; Peiro, A.; Diaz, M.; Ibanez, L.; Gago-Ferrero, P.; Diaz-Cruz, M.S. Development of a sensitive analytical method for the simultaneous analysis of Benzophenone-type UV filters and paraben preservatives in umbilical cord blood. MethodsX 2021, 8, 101307. [Google Scholar] [CrossRef]
- Sunyer-Caldú, A.; Peiró, A.; Díaz, M.; Ibáñez, L.; Gil-Solsona, R.; Gago-Ferrero, P.; Diaz-Cruz, M.S. Target analysis and suspect screening of UV filters, parabens and other chemicals used in personal care products in human cord blood: Prenatal exposure by mother-fetus transfer. Environ. Int. 2023, 173, 107834. [Google Scholar] [CrossRef]
- Jiménez-Díaz, I.; Molina-Molina, J.M.; Zafra-Gómez, A.; Ballesteros, O.; Navalón, A.; Real, M.; Sáenz, J.M.; Fernández, M.F.; Olea, N. Simultaneous determination of the UV-filters benzyl salicylate, phenyl salicylate, octyl salicylate, homosalate, 3-(4-methylbenzylidene) camphor and 3-benzylidene camphor in human placental tissue by LC-MS/MS. Assessment of their in vitro endocrine activity. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2013, 936, 80–87. [Google Scholar] [CrossRef]
- Collaris, E.J.H.; Frank, J. Photoallergic contact dermatitis caused by ultraviolet filters in different sunscreens. Int. J. Dermatol. 2008, 47, 35–37. [Google Scholar] [CrossRef] [PubMed]
- Oral, D.; Yirun, A.; Erkekoglu, P. Safety Concerns of Organic Ultraviolet Filters: Special Focus on Endocrine-Disrupting Properties. J. Environ. Pathol. Toxicol. Oncol. 2020, 39, 201–212. [Google Scholar] [CrossRef]
- Kinnberg, K.L.; Petersen, G.I.; Albrektsen, M.; Minghlani, M.; Awad, S.M.; Holbech, B.F.; Green, J.W.; Bjerregaard, P.; Holbech, H. Endocrine-disrupting effect of the ultraviolet filter benzophenone-3 in zebrafish, Danio rerio. Environ. Toxicol. Chem. 2015, 34, 2833–2840. [Google Scholar] [CrossRef]
- Carstensen, L.; Beil, S.; Boernick, H.; Stolte, S. Structure-related endocrine-disrupting potential of environmental transformation products of benzophenone-type UV filters: A review. J. Hazard. Mater. 2022, 430, 128495. [Google Scholar] [CrossRef]
- Watanabe, Y.; Kojima, H.; Takeuchi, S.; Uramaru, N.; Sanoh, S.; Sugihara, K.; Kitamura, S.; Ohta, S. Metabolism of UV-filter benzophenone-3 by rat and human liver microsomes and its effect on endocrine-disrupting activity. Toxicol. Appl. Pharmacol. 2015, 282, 119–128. [Google Scholar] [CrossRef]
- Birkhauser, M. UV filters and the endocrine system. Gynakol. Endokrinol. 2016, 14, 165–173. [Google Scholar] [CrossRef]
- Nicolopoulou-Stamati, P.; Hens, L.; Sasco, A.J. Cosmetics as endocrine disruptors: Are they a health risk? Rev. Endocr. Metab. Disord. 2015, 16, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Maipas, S.; Nicolopoulou-Stamati, P. Sun lotion chemicals as endocrine disruptors. Horm.-Int. J. Endocrinol. Metab. 2015, 14, 32–46. [Google Scholar] [CrossRef]
- Wang, J.; Pan, L.; Wu, S.; Lu, L.; Xu, Y.; Zhu, Y.; Guo, M.; Zhuang, S. Recent Advances on Endocrine Disrupting Effects of UV Filters. Int. J. Environ. Res. Public Health 2016, 13, 782. [Google Scholar] [CrossRef]
- Schlumpf, M.; Kypke, K.; Vökt, C.C.; Birchler, M.; Durrer, S.; Faass, O.; Ehnes, C.; Fuetsch, M.; Gaille, C.; Henseler, M.; et al. Endocrine active UV filters: Developmental toxicity and exposure through breast milk. Chimia 2008, 62, 345–351. [Google Scholar] [CrossRef]
- Schlumpf, M.; Schmid, P.; Durrer, S.; Conscience, M.; Maerkel, K.; Henseler, M.; Gruetter, M.; Herzog, I.; Reolon, S.; Ceccatelli, R.; et al. Endocrine activity and developmental toxicity of cosmetic UV filters—An update. Toxicology 2004, 205, 113–122. [Google Scholar] [CrossRef]
- Lorigo, M.; Mariana, M.; Cairrao, E. Photoprotection of ultraviolet-B filters: Updated review of endocrine disrupting properties. Steroids 2018, 131, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Passamani, F.; Santos, B.A.M.C.; Gonçalves, A.S. Endocrine disruption study of active sunscreens’s compounds by docking and molecular dynamics simulations. Preprint 2023. [Google Scholar] [CrossRef]
- Krause, M.; Klit, A.; Jensen, M.B.; Søeborg, T.; Frederiksen, H.; Schlumpf, M.; Lichtensteiger, W.; Skakkebaek, N.E.; Drzewiecki, K.T. Sunscreens: Are they beneficial for health? An overview of endocrine disrupting properties of UV-filters. Int. J. Androl. 2012, 35, 424–436. [Google Scholar] [CrossRef]
- Kwon, B.; Choi, K. Occurrence of major organic UV filters in aquatic environments and their endocrine disruption potentials: A mini-review. Integr. Environ. Assess. Manag. 2021, 17, 940–950. [Google Scholar] [CrossRef]
- Ozáez, I.; Morcillo, G.; Martínez-Guitarte, J.L. Ultraviolet filters differentially impact the expression of key endocrine and stress genes in embryos and larvae of Chironomus riparius. Sci. Total Environ. 2016, 557, 240–247. [Google Scholar] [CrossRef]
- Kim, S.; Jung, D.; Kho, Y.; Choi, K. Effects of benzophenone-3 exposure on endocrine disruption and reproduction of Japanese medaka (Oryzias latipes)—A two generation exposure study. Aquat. Toxicol. 2014, 155, 244–252. [Google Scholar] [CrossRef]
- Ka, Y.; Ji, K. Waterborne exposure to avobenzone and octinoxate induces thyroid endocrine disruption in wild-type and thr alphaa−/− zebrafish larvae. Ecotoxicology 2022, 31, 948–955. [Google Scholar] [CrossRef]
- Faass, O.; Schlumpf, M.; Reolon, S.; Henseler, M.; Maerkel, K.; Durrer, S.; Lichtensteiger, W. Female sexual behavior, estrous cycle and gene expression in sexually dimorphic brain regions after pre- and postnatal exposure to endocrine active UV filters. Neurotoxicology 2009, 30, 249–260. [Google Scholar] [CrossRef]
- Nashev, L.G.; Schuster, D.; Laggner, C.; Sodha, S.; Langer, T.; Wolber, G.; Odermatt, A. The UV-filter benzophenone-1 inhibits 17β-hydroxysteroid dehydrogenase type 3: Virtual screening as a strategy to identify potential endocrine disrupting chemicals. Biochem. Pharmacol. 2010, 79, 1189–1199. [Google Scholar] [CrossRef] [PubMed]
- Ozáez, I.; Martínez-Guitarte, J.L.; Morcillo, G. Effects of in vivo exposure to UV filters (4-MBC, OMC, BP-3, 4-HB, OC, OD-PABA) on endocrine signaling genes in the insect Chironomus riparius. Sci. Total Environ. 2013, 456, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Krause, M.; Frederiksen, H.; Sundberg, K.; Jørgensen, F.S.; Jensen, L.N.; Nørgaard, P.; Jørgensen, C.; Ertberg, P.; Petersen, J.H.; Feldt-Rasmussen, U.; et al. Maternal exposure to UV filters: Associations with maternal thyroid hormones, IGF-I/IGFBP3 and birth outcomes. Endocr. Connect. 2018, 7, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Rehfeld, A.; Egeberg, D.L.; Almstrup, K.; Petersen, J.H.; Dissing, S.; Skakkebaek, N.E. EDC IMPACT: Chemical UV filters can affect human sperm function in a progesterone-like manner. Endocr. Connect. 2018, 7, 16–25. [Google Scholar] [CrossRef] [PubMed]
- LaPlante, C.D.; Bansal, R.; Dunphy, K.A.; Jerry, D.J.; Vandenberg, L.N. Oxybenzone Alters Mammary Gland Morphology in Mice Exposed During Pregnancy and Lactation. J. Endocr. Soc. 2018, 2, 903–921. [Google Scholar] [CrossRef]
- Ruszkiewicz, J.A.; Pinkas, A.; Ferrer, B.; Peres, T.V.; Tsatsakis, A.; Aschner, M. Neurotoxic effect of active ingredients in sunscreen products, a contemporary review. Toxicol. Rep. 2017, 4, 245. [Google Scholar] [CrossRef]
- Kunz, P.Y.; Galicia, H.F.; Fent, K. Assessment of hormonal activity of UV filters in tadpoles of frog Xenopus laevis at environmental concentrations. Mar. Environ. Res. 2004, 58, 431–435. [Google Scholar] [CrossRef]
- Kunz, P.Y.; Galicia, H.F.; Fent, K. Comparison of in vitro and in vivo estrogenic activity of UV filters in fish. Toxicol. Sci. 2006, 90, 349–361. [Google Scholar] [CrossRef]
- Weisbrod, C.J.; Kunz, P.Y.; Zenker, A.K.; Fent, K. Effects of the UV filter benzophenone-2 on reproduction in fish. Toxicol. Appl. Pharmacol. 2007, 225, 255–266. [Google Scholar] [CrossRef]
- Fent, K.; Kunz, P.; Wolf, M.; Schmutz, H.; Gygax, D.; Schanne, C. Chemical analysis and ecotoxicological effects of UV absorbing organic chemicals in aquatic environments. Chimia 2004, 58, 325–327. [Google Scholar] [CrossRef]
- Blanco-Castañeda, R.; Galaviz-Hernández, C.; Souto, P.C.S.; Lima, V.V.; Giachini, F.R.; Escudero, C.; Damiano, A.E.; Barragán-Zúñiga, L.J.; Martínez-Aguilar, G.; Sosa-Macías, M. The role of xenobiotic-metabolizing enzymes in the placenta: A growing research field. Expert. Rev. Clin. Pharmacol. 2020, 13, 247–263. [Google Scholar] [CrossRef] [PubMed]
- Duru, C.E. Forever chemicals could expose the human fetus to xenobiotics by binding to placental enzymes: Prescience from molecular docking, DFT, and machine learning. Comput. Toxicol. 2023, 26, 100274. [Google Scholar] [CrossRef]
- Enyoh, C.E.; Duru, C.E.; Ovuoraye, P.E.; Wang, Q. Evaluation of nanoplastics toxicity to the human placenta in systems. J. Hazard. Mater. 2023, 446, 130600. [Google Scholar] [CrossRef]
- Sobańska, A.W.; Banerjee, A.; Roy, K. Organic Sunscreens and Their Products of Degradation in Biotic and Abiotic Conditions—In Silico Studies of Drug-Likeness and Human Placental Transport. Int. J. Mol. Sci. 2024, 25, 12373. [Google Scholar] [CrossRef]
- Pirotta, G. Sunscreen Regulation in the World; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar] [CrossRef]
- Mazari, A.M.A.; Zhang, L.; Ye, Z.-W.; Zhang, J.; Tew, K.D.; Townsend, D.M. The Multifaceted Role of Glutathione S-Transferases in Health and Disease. Biomolecules 2023, 13, 688. [Google Scholar] [CrossRef] [PubMed]
- Sim, E.; Abuhammad, A.; Ryan, A. Arylamine N-acetyltransferases: From drug metabolism and pharmacogenetics to drug discovery. Br. J. Pharmacol. 2014, 171, 2705–2725. [Google Scholar] [CrossRef] [PubMed]
- Neural Network Architecture. In Encyclopedia of Machine Learning; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2011; 716p. [CrossRef]
- Miikkulainen, R. Topology of a Neural Network. In Encyclopedia of Machine Learning; Springer: Boston, MA, USA, 2011; pp. 988–989. [Google Scholar] [CrossRef]
- Markowska-Kaczmar, U.; Czeczot, K. Creativity of Neural Networks. In Lecture Notes in Computer Science (Including Subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics); Springer: Berlin/Heidelberg, Germany, 2006; Volume 4029, pp. 86–93. [Google Scholar] [CrossRef]
- Hall, L.H.; Kier, L.B. Electrotopological State Indices for Atom Types: A Novel Combination of Electronic, Topological, and Valence State Information. J. Chem. Inf. Comput. Sci. 1995, 35, 1039–1045. [Google Scholar] [CrossRef]
- Hall, L.H.; Kier, L.B.; Brown, B.B. Molecular Similarity Based on Novel Atom-Type Electrotopological State Indices. J. Chem. Inf. Comput. Sci. 1995, 35, 1074–1080. [Google Scholar] [CrossRef]
- Mathew, N.; Kalyanasundaram, M.; Balaraman, K. Glutathione S-transferase (GST) inhibitors. Expert Opin. Ther. Pat. 2006, 16, 431–444. [Google Scholar] [CrossRef]
- Schultz, M.; Dutta, S.; Tew, K.D. Inhibitors of glutathione S-transferases as therapeutic agents. Adv. Drug Deliv. Rev. 1997, 26, 91–104. [Google Scholar] [CrossRef]
- Gökçe, B. Some anticancer agents as effective glutathione S-transferase (GST) inhibitors. Open Chem. 2023, 21, 20230159. [Google Scholar] [CrossRef]
- Ayna, A.; Khosnaw, L.; Temel, Y.; Ciftci, M. Antibiotics as Inhibitor of Glutathione S-transferase: Biological Evaluation and Molecular Structure Studies. Curr. Drug Metab. 2021, 22, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Alqarni, M.H.; Foudah, A.I.; Muharram, M.M.; Alam, A.; Labrou, N.E. Myricetin as a Potential Adjuvant in Chemotherapy: Studies on the Inhibition of Human Glutathione Transferase A1–1. Biomolecules 2022, 12, 1364. [Google Scholar] [CrossRef]
- Shokeer, A.; Zaki, E.R.; Saleh, N.S.E.-D.; Guneidy, R.A. Dual Action of Resveratrol: Inhibition of Human Glutathione Transferase and its Antagonism with Tamoxifen on Breast Cancer Cell Viability. Egypt. J. Chem. 2024, 67, 601–611. [Google Scholar] [CrossRef]
- Kobzar, O.; Shulha, Y.; Buldenko, V.; Cherenok, S.; Silenko, O. Inhibition of glutathione S-transferases by photoactive calix[4]arene α-ketophosphonic acids. Bioorganic Med. Chem. Lett. 2022, 77, 129019. [Google Scholar] [CrossRef]
- Irvine, J.D.; Takahashi, L.; Lockhart, K.; Cheong, J.; Tolan, J.W.; Selick, H.E.; Grove, J.R. MDCK (Madin-Darby canine kidney) cells: A tool for membrane permeability screening. J. Pharm. Sci. 1999, 88, 28–33. [Google Scholar] [CrossRef]
- Ballester, P.J.; Westwood, I.; Laurieri, N.; Sim, E.; Richards, W.G. Prospective virtual screening with ultrafast shape recognition: The identification of novel inhibitors of arylamine N-acetyltransferases. J. R. Soc. Interface 2010, 7, 335–342. [Google Scholar] [CrossRef]
- Rothen, J.P.; Haefeli, W.E.; Meyer, U.A.; Todesco, L.; Wenk, M. Acetaminophen is an inhibitor of hepatic N-acetyltransferase 2 in vitro and in vivo. Pharmacogenetics 1998, 8, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Abuhammad, A.; Fullam, E.; Lowe, E.D.; Staunton, D.; Kawamura, A.; Westwood, I.M.; Bhakta, S.; Garner, A.C.; Wilson, D.L.; Seden, P.T.; et al. Piperidinols That Show Anti-Tubercular Activity as Inhibitors of Arylamine N-Acetyltransferase: An Essential Enzyme for Mycobacterial Survival Inside Macrophages. PLoS ONE 2012, 7, e52790. [Google Scholar] [CrossRef]
- Kukongviriyapan, V.; Phromsopha, N.; Tassaneeyakul, W.; Kukongviriyapan, U.; Sripa, B.; Hahnvajanawong, V.; Bhudhisawasdi, V. Inhibitory effects of polyphenolic compounds on human arylamine N-acetyltransferase 1 and 2. Xenobiotica 2006, 36, 15–28. [Google Scholar] [CrossRef]
- Shadrack, D.M.; Ndesendo, V.M.K. Molecular Docking and ADMET Study of Emodin Derivatives as Anticancer Inhibitors of NAT2, COX2 and TOP1 Enzymes. Comput. Mol. Biosci. 2017, 7, 1–18. [Google Scholar] [CrossRef]
- Wang, C.C.; Lin, P.; Chou, C.Y.; Wang, S.S.; Tung, C.W. Prediction of human fetal-maternal blood concentration ratio of chemicals. PeerJ 2020, 8, e9562. [Google Scholar] [CrossRef] [PubMed]
- Sobańska, A.W. In silico assessment of risks associated with pesticides exposure during pregnancy. Chemosphere 2023, 329, 138649. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed]
- Moriwaki, H.; Tian, Y.-S.; Kawashita, N.; Takagi, T. Mordred: A molecular descriptor calculator. J. Cheminform. 2018, 10, 4. [Google Scholar] [CrossRef]
- Sushko, I.; Novotarskyi, S.; Körner, R.; Pandey, A.K.; Rupp, M.; Teetz, W.; Brandmaier, S.; Abdelaziz, A.; Prokopenko, V.V.; Tanchuk, V.Y.; et al. Online chemical modeling environment (OCHEM): Web platform for data storage, model development and publishing of chemical information. J. Comput. Aided Mol. Des. 2011, 25, 533–554. [Google Scholar] [CrossRef]
- Mehmood, T.; Sæbø, S.; Liland, K.H. Comparison of variable selection methods in partial least squares regression. J. Chemom. 2020, 34, e3226. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A.J. Small-Molecule Library Screening by Docking with PyRx. Methods Mol. Biol. 2015, 1263, 243–250. [Google Scholar] [CrossRef]
- Gramatica, P. On the development and validation of QSAR models. Methods Mol. Biol. 2013, 930, 499–526. [Google Scholar] [CrossRef]
- Marie, C.; Cabut, S.; Vendittelli, F.; Sauvant-Rochat, M.P. Changes in cosmetics use during pregnancy and risk perception by women. Int. J. Environ. Res. Public Health 2016, 13, 383. [Google Scholar] [CrossRef]
- Shareef, S.; Shoemaker, W.B.; Abdelnour, A.; Albazi, E.; Ashack, K. Assessing public interest in sunscreen safety during pregnancy and lactation. Int. J. Womens Dermatol. 2022, 8, e018. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Ka, Y.; Lee, B.; Lee, I.; Seo, Y.E.; Shin, H.; Kho, Y.; Ji, K. Single and mixture toxicity evaluation of avobenzone and homosalate to male zebrafish and H295R cells. Chemosphere 2023, 343, 140271. [Google Scholar] [CrossRef] [PubMed]
- Wnuk, W.; Michalska, K.; Krupa, A.; Pawlak, K. Benzophenone-3, a chemical UV-filter in cosmetics: Is it really safe for children and pregnant women? Postępy Dermatol. Alergol. 2022, 39, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Carou, M.E.; Szwarcfarb, B.; Deguiz, M.L.; Reynoso, R.; Carbone, S.; Moguilevsky, J.A.; Scacchi, P.; Ponzo, O.J. Impact of 4-methylbenzylidene-camphor (4-MBC) during embryonic and fetal development in the neuroendocrine regulation of testicular axis in prepubertal and peripubertal male rats. Exp. Clin. Endocrinol. Diabetes 2009, 117, 449–454. [Google Scholar] [CrossRef]
- DiNardo, J.C.; Downs, C.A. Can oxybenzone cause Hirschsprung’s disease? Reprod. Toxicol. 2019, 86, 98–100. [Google Scholar] [CrossRef]
- Huo, W.; Cai, P.; Chen, M.; Li, H.; Tang, J.; Xu, C.; Zhu, D.; Tang, W.; Xia, Y. The relationship between prenatal exposure to BP-3 and Hirschsprung’s disease. Chemosphere 2016, 144, 1091–1097. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | PL0 (n = 16) | PL1 (n = 54) |
|---|---|---|
| Mw | 493.72 | 292.63 |
| #HvAt | 32.75 | 20.28 |
| #ArHvAt | 11.06 | 7.35 |
| FCsp3 | 0.47 | 0.43 |
| #HRB | 9.94 | 4.96 |
| #HA | 3.69 | 3.83 |
| #HD | 1.69 | 1.44 |
| MR | 134.97 | 80.67 |
| TPSA | 70.41 | 70.65 |
| XLOGP3 | 6.42 | 2.40 |
| nAcid | 0.00 | 0.20 |
| nBase | 0.25 | 0.43 |
| nSpiro | 0.00 | 0.09 |
| nBridgehead | 0.38 | 0.26 |
| nHetero | 7.31 | 5.52 |
| Lipinski | 0.06 | 0.94 |
| GhoseFilter | 0.25 | 0.80 |
| kGST (kcal/mol) | kNAT2 (kcal/mol) | |
|---|---|---|
| BMDM | −7.50 | −9.60 |
| BP-3 | −7.30 | −8.70 |
| DHHB | −6.90 | −8.20 |
| PABA | −5.10 | −6.30 |
| EHDP | −6.00 | −7.50 |
| Et-PABA | −5.60 | −6.70 |
| PBSA | −7.70 | −9.50 |
| MBC | −7.60 | −8.80 |
| EHMC | −6.30 | −7.80 |
| IMC | −6.30 | −7.70 |
| OCR | −7.80 | −8.70 |
| ET | −7.40 | −7.30 |
| OS | −6.20 | −8.20 |
| HMS | −7.60 | −8.70 |
| DOBT | −8.20 | −7.50 |
| BP-4 | −8.00 | −9.30 |
| Reference | −6.00 | −8.70 |
| GST | Leu A114 | Leu A119 | Val A36 | Leu A35 | Val A10 | Gln A12 | Met A236 | Trp A115 | Total |
|---|---|---|---|---|---|---|---|---|---|
| van der Waals | 13 | 7 | 9 | 1 | 8 | 7 | 6 | 2 | 53 |
| H-bond | 1 | 1 | 2 | 6 | 10 | ||||
| π-σ | 4 | 7 | 11 | ||||||
| alkyl | 2 | 5 | 5 | 7 | 5 | 3 | 3 | 30 | |
| π-alkyl | 0 | ||||||||
| unfavorable donor–donor | 1 | 1 | |||||||
| π-π | 3 | 3 |
| NAT2 | Phe A217 | Phe A93 | Phe A37 | Gly A126 | Ser A125 | Leu A288 | Ser A129 | Cys A68 | Total |
|---|---|---|---|---|---|---|---|---|---|
| van der Waals | 4 | 2 | 10 | 10 | 13 | 9 | 10 | 58 | |
| H-bond | 1 | 2 | 3 | ||||||
| π-sulfur | 6 | 6 | |||||||
| π-π | 9 | 11 | 1 | 21 | |||||
| alkyl | 2 | 2 | 4 | 1 | 6 | 15 | |||
| π-σ | 1 | 1 | 2 |
| kGST | kNAT2 | Mw | #HvAt | #ArHvAt | FCsp3 | #FRB | #HA | MR | #HD | TPSA | XLOGP3 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| kGST | 1.00 | 0.72 | −0.51 | −0.50 | −0.62 | −0.02 | −0.27 | −0.51 | −0.28 | −0.48 | −0.43 | −0.40 |
| kNAT2 | 0.72 | 1.00 | 0.10 | 0.11 | −0.08 | 0.13 | 0.29 | 0.03 | 0.24 | 0.13 | 0.12 | 0.15 |
| Mw | −0.51 | 0.10 | 1.00 | 1.00 | 0.90 | 0.35 | 0.95 | 0.90 | 0.65 | 1.00 | 0.82 | 0.94 |
| #HvAt | −0.50 | 0.11 | 1.00 | 1.00 | 0.89 | 0.36 | 0.95 | 0.88 | 0.64 | 1.00 | 0.80 | 0.94 |
| #ArHvAt | −0.62 | −0.08 | 0.90 | 0.89 | 1.00 | −0.07 | 0.78 | 0.90 | 0.76 | 0.87 | 0.89 | 0.73 |
| FCsp3 | −0.02 | 0.13 | 0.35 | 0.36 | −0.07 | 1.00 | 0.45 | 0.05 | −0.24 | 0.40 | −0.15 | 0.58 |
| #FRB | −0.27 | 0.29 | 0.95 | 0.95 | 0.78 | 0.45 | 1.00 | 0.83 | 0.55 | 0.96 | 0.72 | 0.95 |
| #HA | −0.51 | 0.03 | 0.90 | 0.88 | 0.90 | 0.05 | 0.83 | 1.00 | 0.80 | 0.86 | 0.95 | 0.74 |
| MR | −0.28 | 0.24 | 0.65 | 0.64 | 0.76 | −0.24 | 0.55 | 0.80 | 1.00 | 0.61 | 0.93 | 0.43 |
| #HD | −0.48 | 0.13 | 1.00 | 1.00 | 0.87 | 0.40 | 0.96 | 0.86 | 0.61 | 1.00 | 0.77 | 0.96 |
| TPSA | −0.43 | 0.12 | 0.82 | 0.80 | 0.89 | −0.15 | 0.72 | 0.95 | 0.93 | 0.77 | 1.00 | 0.60 |
| XLOGP3 | −0.40 | 0.15 | 0.94 | 0.94 | 0.73 | 0.58 | 0.95 | 0.74 | 0.43 | 0.96 | 0.60 | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobańska, A.W.; Sobański, A.M. Organic Sunscreens—Is Their Placenta Permeability the Only Issue Associated with Exposure During Pregnancy? In Silico Studies of Sunscreens’ Placenta Permeability and Interactions with Selected Placental Enzymes. Molecules 2024, 29, 5836. https://doi.org/10.3390/molecules29245836
Sobańska AW, Sobański AM. Organic Sunscreens—Is Their Placenta Permeability the Only Issue Associated with Exposure During Pregnancy? In Silico Studies of Sunscreens’ Placenta Permeability and Interactions with Selected Placental Enzymes. Molecules. 2024; 29(24):5836. https://doi.org/10.3390/molecules29245836
Chicago/Turabian StyleSobańska, Anna W., and Andrzej M. Sobański. 2024. "Organic Sunscreens—Is Their Placenta Permeability the Only Issue Associated with Exposure During Pregnancy? In Silico Studies of Sunscreens’ Placenta Permeability and Interactions with Selected Placental Enzymes" Molecules 29, no. 24: 5836. https://doi.org/10.3390/molecules29245836
APA StyleSobańska, A. W., & Sobański, A. M. (2024). Organic Sunscreens—Is Their Placenta Permeability the Only Issue Associated with Exposure During Pregnancy? In Silico Studies of Sunscreens’ Placenta Permeability and Interactions with Selected Placental Enzymes. Molecules, 29(24), 5836. https://doi.org/10.3390/molecules29245836