D-Limonene Inhibits Pichia kluyveri Y-11519 in Sichuan Pickles by Disrupting Metabolism

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
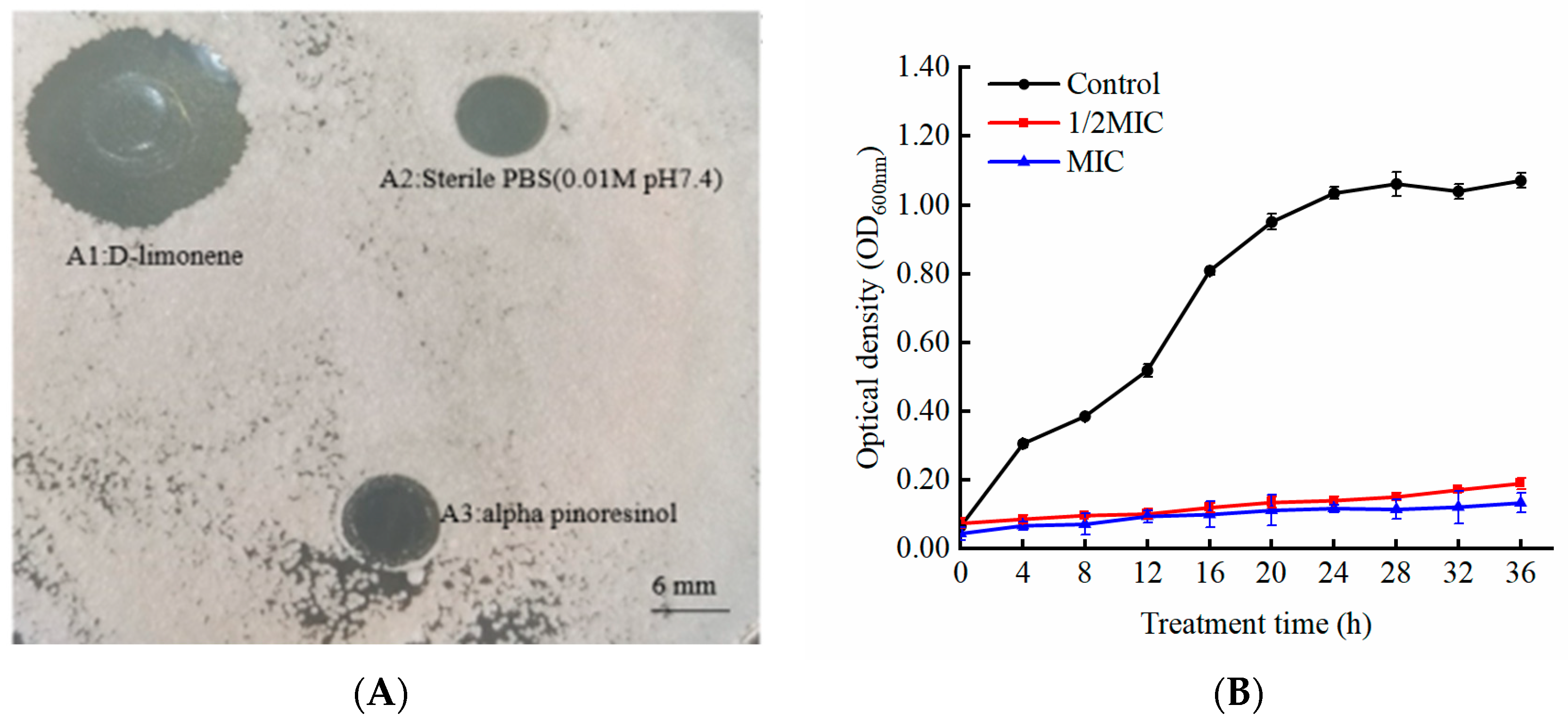
2.1. The Antifungal Activity of D-Limonene
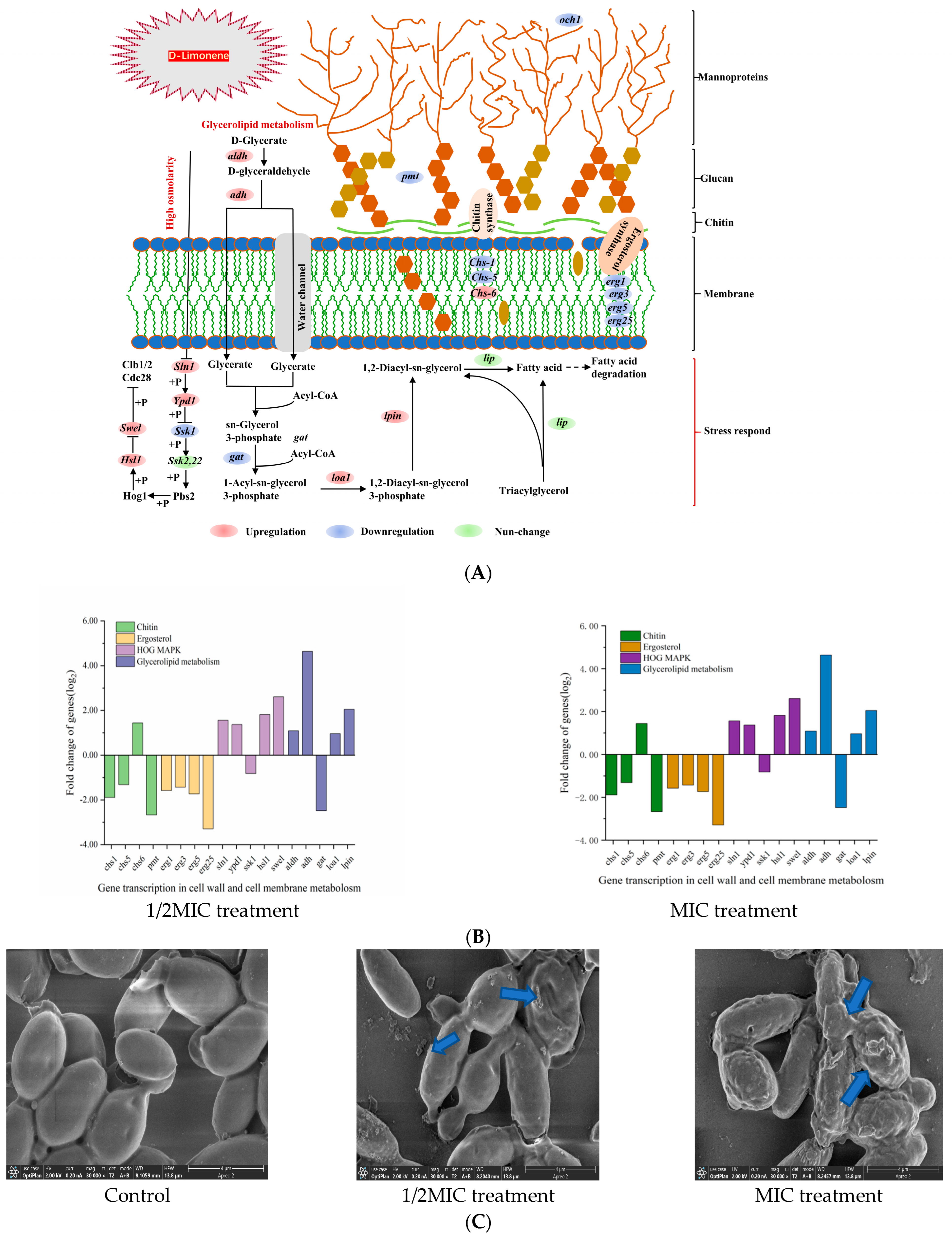
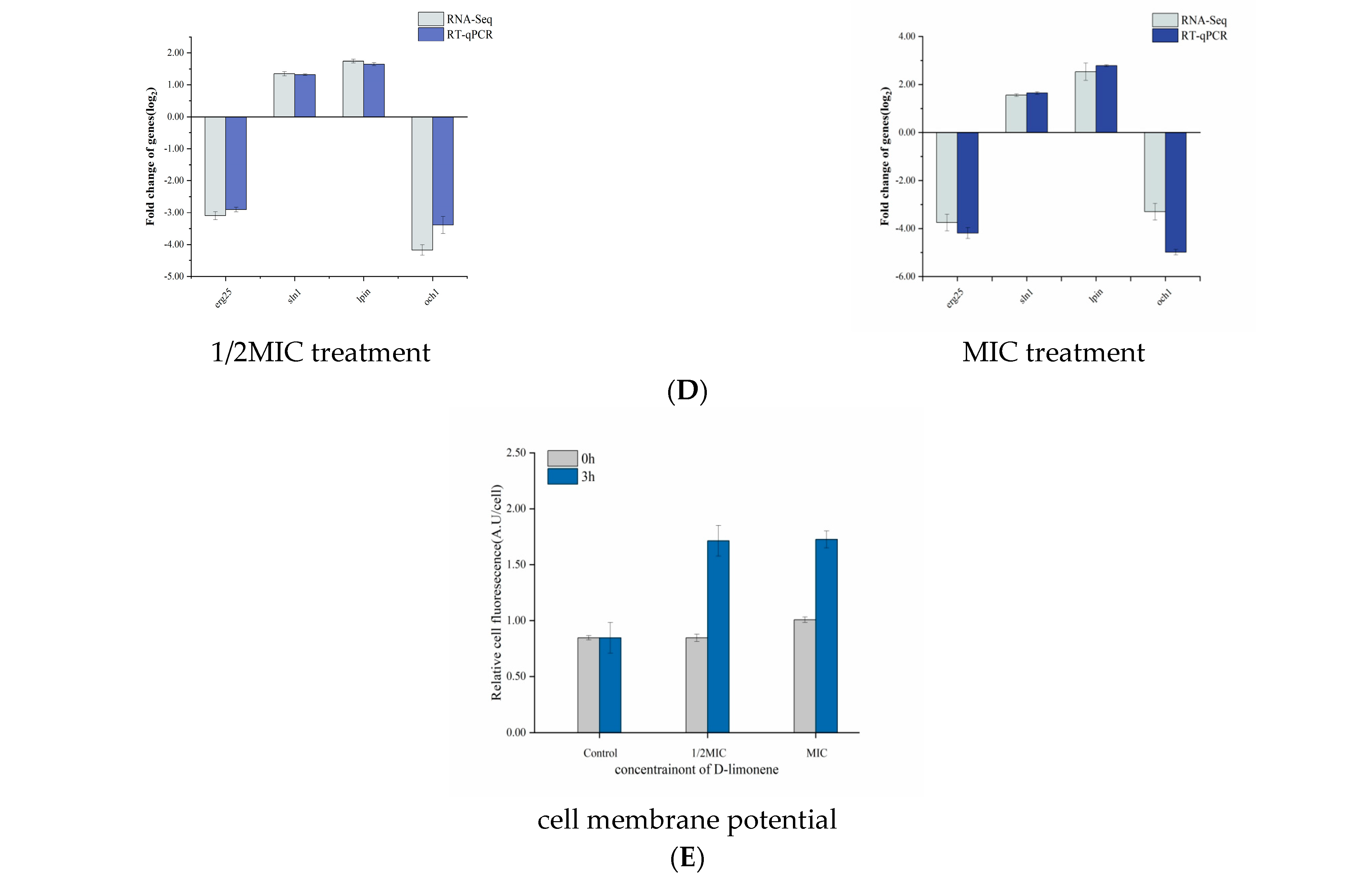
2.2. The Effect of D-Limonene on the Cell Wall and Membrane
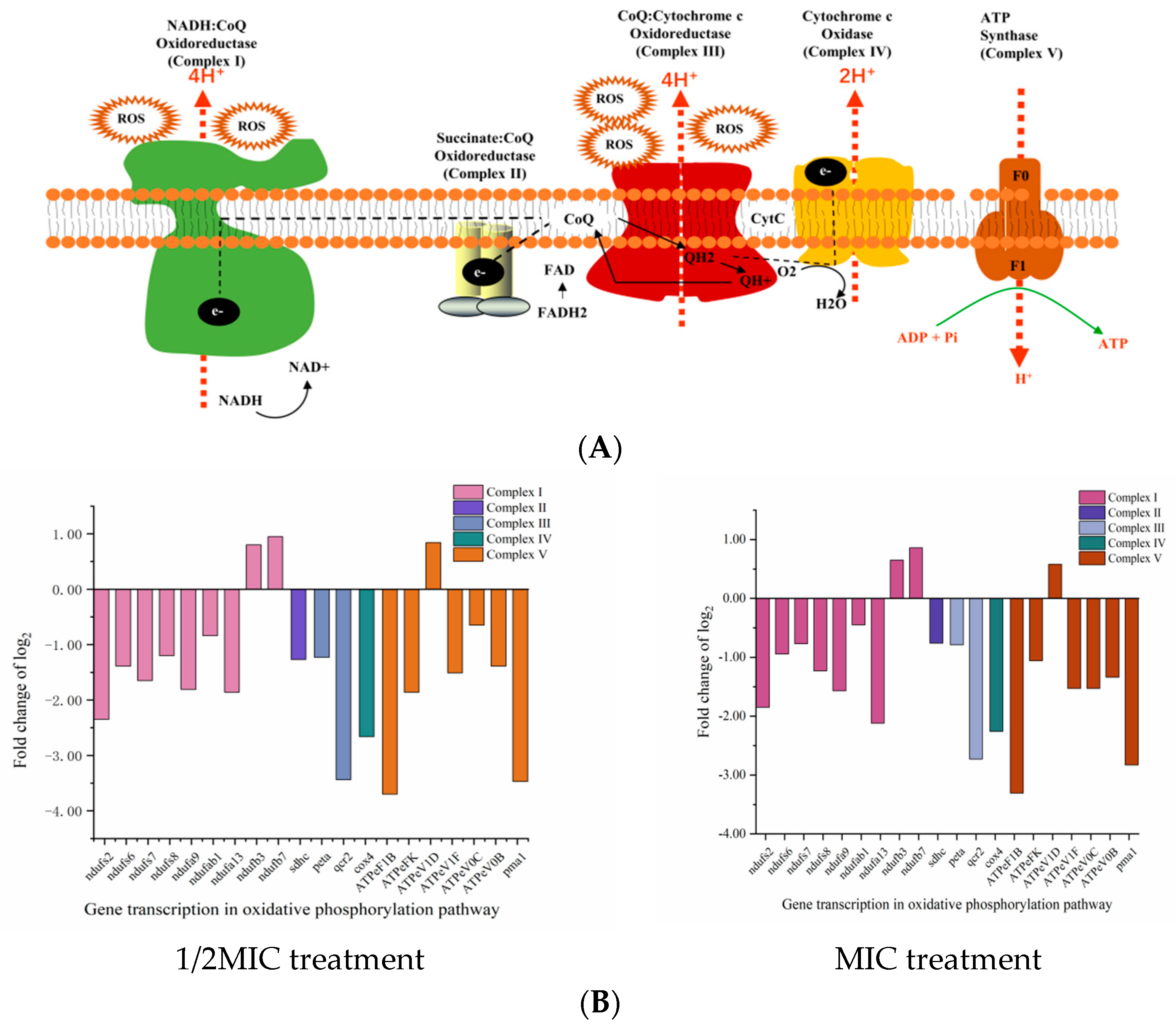
2.3. The Effect of D-Limonene on the Oxidative Phosphorylation Pathway
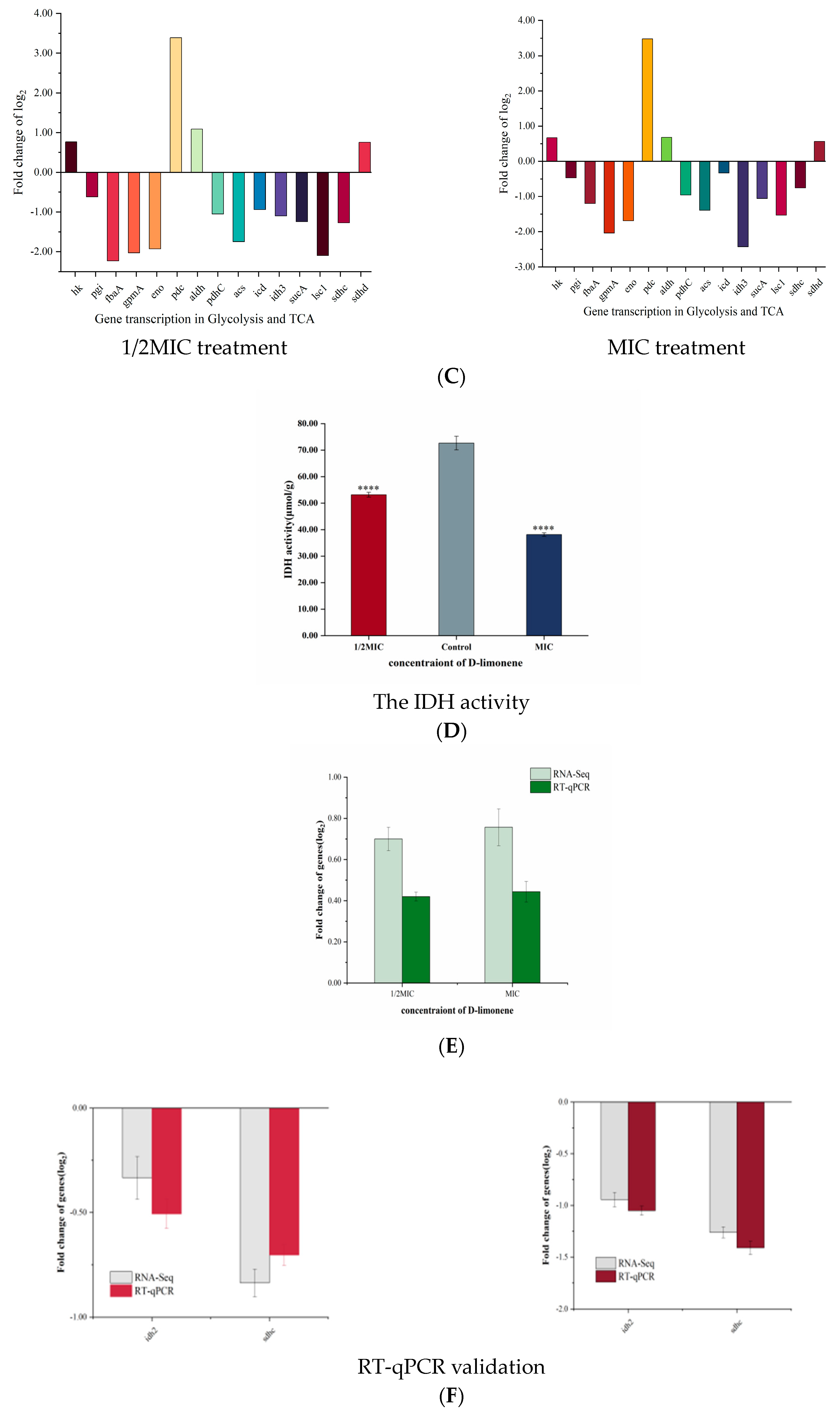
2.4. The Effect of D-Limonene on the Glycolytic Pathway
2.5. The Effect of D-Limonene on the TCA Cycle
3. Discussion
4. Materials and Methods
4.1. Strains and Reagents
4.2. Antifungal Activity
4.3. Cell Morphology
4.4. Cell Membrane Potential
4.5. Mitochondrial Adenosine Triphosphate (ATP) Content
4.6. Endogenous ROS
4.7. The HK Activity
4.8. The IDH Activity
4.9. RNA Sequencing and RT-PCR Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rao, Y.; Qian, Y.; She, X.; Yang, J.; He, P.; Jiang, Y.; Wang, M.; Xiang, W. Pellicle formation, microbial succession and lactic acid utilisation during the aerobic deteriorating process of Sichuan pickle. Int. J. Food Sci. Technol. 2017, 53, 767–775. [Google Scholar] [CrossRef]
- Rao, Y.; Tao, Y.; Li, Y.; She, X.; Yang, J.; Qian, Y.; Du, H.; Liu, L.; Xiao, H. Characterization of a probiotic starter culture with anti-Candida activity for Chinese pickle fermentation. Food Funct. 2019, 10, 6936–6944. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.-L.; Zhang, N.-D.; Lu, Y.; Zhao, Q.-H.; Xu, Q.; Rao, Y.; Liu, L.; Zhang, Q. Effect of Weissella cibaria co-inoculation on the quality of Sichuan Pickle fermented by Lactobacillus plantarum. LWT 2020, 121, 108975. [Google Scholar] [CrossRef]
- Ao, X.; Yan, J.; Chen, C.; Zhao, J.; Liu, S.; Zhao, K.; Chen, S.; He, L. Isolation and identification of the spoilage microorganisms in Sichuan homemade Paocai and their impact on quality and safety. Food Sci. Nutr. 2019, 7, 2939–2947. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, S.H.; Jeon, C.O. Kimchi microflora: History, current status, and perspectives for industrial kimchi production. Appl. Microbiol. Biotechnol. 2014, 98, 2385–2393. [Google Scholar] [CrossRef]
- Labbani, F.-Z.K.; Turchetti, B.; Bennamoun, L.; Dakhmouche, S.; Roberti, R.; Corazzi, L.; Meraihi, Z.; Buzzini, P. A novel killer protein from Pichia kluyveri isolated from an Algerian soil: Purification and characterization of its in vitro activity against food and beverage spoilage yeasts. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2015, 107, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Kesmen, Z.; Özbekar, E.; Büyükkiraz, M. Multifragment melting analysis of yeast species isolated from spoiled fruits. J. Appl. Microbiol. 2018, 124, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Vadkertiova, R.; Molnarova, J.; Vranova, D.; Slavikova, E. Yeasts and yeast-like organisms associated with fruits and blossoms of different fruit trees. Can. J. Microbiol. 2012, 58, 1344–1352. [Google Scholar] [CrossRef]
- Hernández, A.; Pérez-Nevado, F.; Ruiz-Moyano, S.; Serradilla, M.J.; Villalobos, M.C.; Martín, A.; Córdoba, M.G. Spoilage yeasts: What are the sources of contamination of foods and beverages? Int. J. Food Microbiol. 2018, 286, 98–110. [Google Scholar] [CrossRef]
- Vicente, J.; Calderón, F.; Santos, A.; Marquina, D.; Benito, S. High potential of Pichia kluyveri and other Pichia species in wine technology. Int. J. Mol. Sci. 2021, 22, 1196. [Google Scholar] [CrossRef]
- Arroyo-López, F.; Romero-Gil, V.; Bautista-Gallego, J.; Rodríguez-Gómez, F.; Jiménez-Díaz, R.; García-García, P.; Querol, A.; Garrido-Fernández, A. Yeasts in table olive processing: Desirable or spoilage microorganisms? Int. J. Food Microbiol. 2012, 160, 42–49. [Google Scholar] [CrossRef]
- Cai, T.; Shi, P.; Zhang, S.; Xiang, W.; Liu, J.; Lin, Z.; Tang, J. Inhibition of Perilla frutescens Essential Oil on Pellicle Formation of Candida tropicalis and Pichia kluyveri and Its Effect on Volatile Compounds in Sichuan Pickles. Foods 2023, 12, 1593. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, J.; Cha, I.-T.; Jung, M.Y.; Song, H.S.; Kim, Y.B.; Lee, C.; Kang, S.-Y.; Bae, J.-W.; Choi, Y.-E.; et al. Community structures and genomic features of undesirable white colony-forming yeasts on fermented vegetables. J. Microbiol. 2019, 57, 30–37. [Google Scholar] [CrossRef]
- Zhang, F.; Tang, Y.; Ren, Y.; Yao, K.; He, Q.; Wan, Y.; Chi, Y. Microbial composition of spoiled industrial-scale Sichuan paocai and characteristics of the microorganisms responsible for paocai spoilage. Int. J. Food Microbiol. 2018, 275, 32–38. [Google Scholar] [CrossRef]
- Ribeiro-Santos, R.; Andrade, M.; Sanches-Silva, A.; de Melo, N.R. Essential oils for food application: Natural substances with established biological activities. Food Bioprocess Technol. 2018, 11, 43–71. [Google Scholar] [CrossRef]
- Angane, M.; Swift, S.; Huang, K.; Butts, C.A.; Quek, S.Y. Essential oils and their major components: An updated review on antimicrobial activities, mechanism of action and their potential application in the food industry. Foods 2022, 11, 464. [Google Scholar] [CrossRef]
- Liu, Q.; Meng, X.; Li, Y.; Zhao, C.-N.; Tang, G.-Y.; Li, H.-B. Antibacterial and Antifungal Activities of Spices. Int. J. Mol. Sci. 2017, 18, 1283. [Google Scholar] [CrossRef]
- Redondo-Blanco, S.; Fernández, J.; López-Ibáñez, S.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Plant Phytochemicals in Food Preservation: Antifungal Bioactivity: A Review. J. Food Prot. 2020, 83, 163–171. [Google Scholar] [CrossRef]
- Yin, W.; Shang, M.; Li, X.; Sang, S.; Chen, L.; Long, J.; Jiao, A.; Wang, J.; Jin, Z.; Qiu, C. Recent developments in sources, chemical constituents, health benefits and food applications of essential oils extracted from medicine food homology plants. Food Biosci. 2023, 55, 102997. [Google Scholar] [CrossRef]
- Yoon, D.; Lee, M.-H.; Cha, D. Measurement of Intracellular ROS in Caenorhabditis elegans Using 2′,7′-Dichlorodihydrofluorescein Diacetate. Bio-Protocol 2018, 8, e2774. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Pahmeyer, M.J.; Assadpour, E.; Jafari, S.M. Extraction and purification of d-limonene from orange peel wastes: Recent advances. Ind. Crops Prod. 2022, 177, 114484. [Google Scholar] [CrossRef]
- Azghar, A.; Dalli, M.; Azizi, S.; Benaissa, E.M.; Ben Lahlou, Y.; Elouennass, M.; Maleb, A. Chemical Composition and Antibacterial Activity of Citrus Peels Essential Oils Against Multidrug-Resistant Bacteria: A Comparative Study. J. Herb. Med. 2023, 42, 100799. [Google Scholar] [CrossRef]
- Justino de Araujo, A.C.; Freitas, P.R.; Rodrigues Dos Santos Barbosa, C.; Muniz, D.F.; Rocha, J.E.; Albuquerque da Silva, A.C.; Datiane de Morais Oliveira-Tintino, C.; Ribeiro-Filho, J.; Everson da Silva, L.; Confortin, C.; et al. GC-MS-FID characterization and antibacterial activity of the Mikania cordifolia essential oil and limonene against MDR strains. Food Chem Toxicol 2020, 136, 111023. [Google Scholar] [CrossRef]
- Yu, H.; Lin, Z.-X.; Xiang, W.-L.; Huang, M.; Tang, J.; Lu, Y.; Zhao, Q.-H.; Zhang, Q.; Rao, Y.; Liu, L. Antifungal activity and mechanism of D-limonene against foodborne opportunistic pathogen Candida tropicalis. LWT 2022, 159, 113144. [Google Scholar] [CrossRef]
- Xiong, H.-B.; Zhou, X.-H.; Xiang, W.-L.; Huang, M.; Lin, Z.-X.; Tang, J.; Cai, T.; Zhang, Q. Integrated transcriptome reveals that D-limonene inhibits Candida tropicalis by disrupting metabolism. LWT 2023, 176, 114535. [Google Scholar] [CrossRef]
- Miron, D.; Battisti, F.; Silva, F.K.; Lana, A.D.; Pippi, B.; Casanova, B.; Gnoatto, S.; Fuentefria, A.; Mayorga, P.; Schapoval, E.E. Antifungal activity and mechanism of action of monoterpenes against dermatophytes and yeasts. Rev. Bras. Farmacogn. 2014, 24, 660–667. [Google Scholar] [CrossRef]
- Han, Y.; Sun, Z.; Chen, W. Antimicrobial Susceptibility and Antibacterial Mechanism of Limonene against Listeria monocytogenes. Molecules 2019, 25, 33. [Google Scholar] [CrossRef]
- Gow, N.A.R.; Lenardon, M.D. Architecture of the dynamic fungal cell wall. Nat. Rev. Microbiol. 2023, 21, 248–259. [Google Scholar] [CrossRef]
- Clavijo-Giraldo, D.M.; Pérez-García, L.A.; Hernández-Chávez, M.J.; Martínez-Duncker, I.; Mora-Montes, H.M. Contribution of N-Linked Mannosylation Pathway to Candida parapsilosis and Candida tropicalis Biofilm Formation. Infect. Drug Resist. 2023, 16, 6843–6857. [Google Scholar] [CrossRef]
- Lambou, K.; Perkhofer, S.; Fontaine, T.; Latge, J.-P. Comparative functional analysis of the OCH1 mannosyltransferase families in Aspergillus fumigatus and Saccharomyces cerevisiae. Yeast 2010, 27, 625–636. [Google Scholar] [CrossRef]
- Román, E.; Correia, I.; Prieto, D.; Alonso, R.; Pla, J. The HOG MAPK pathway in Candida albicans: More than an osmosensing pathway. Int. Microbiol. 2020, 23, 23–29. [Google Scholar] [CrossRef]
- Konte, T.; Terpitz, U.; Plemenitas, A. Reconstruction of the High-Osmolarity Glycerol (HOG) Signaling Pathway from the Halophilic Fungus Wallemia ichthyophaga in Saccharomyces cerevisiae. Front. Microbiol. 2016, 7, 901. [Google Scholar] [CrossRef]
- Jordá, T.; Puig, S. Regulation of Ergosterol Biosynthesis in Saccharomyces cerevisiae. Genes 2020, 11, 795. [Google Scholar] [CrossRef]
- Pan, J.; Hu, C.; Yu, J.-H. Lipid Biosynthesis as an Antifungal Target. J. Fungi 2018, 4, 50. [Google Scholar] [CrossRef]
- Athenstaedt, K.; Daum, G. Phosphatidic acid, a key intermediate in lipid metabolism. Eur. J. Biochem. 2001, 266, 1–16. [Google Scholar] [CrossRef]
- Hassaninasab, A.; Hsieh, L.-S.; Su, W.-M.; Han, G.-S.; Carman, G.M. Yck1 casein kinase I regulates the activity and phosphorylation of Pah1 phosphatidate phosphatase from Saccharomyces cerevisiae. J. Biol. Chem. 2019, 294, 18256–18268. [Google Scholar] [CrossRef]
- Baldanzi, G. Inhibition of diacylglycerol kinases as a physiological way to promote diacylglycerol signaling. Adv. Biol. Regul. 2014, 55, 39–49. [Google Scholar] [CrossRef]
- Rocha, P.R.; Medeiros, M.C.; Kintzel, U.; Vogt, J.; Araújo, I.M.; Mestre, A.L.; Mailänder, V.; Schlett, P.; Dröge, M.; Schneider, L. Extracellular electrical recording of pH-triggered bursts in C6 glioma cell populations. Sci. Adv. 2016, 2, e1600516. [Google Scholar] [CrossRef]
- Cai, Y.; Zou, G.; Xi, M.; Hou, Y.; Shen, H.; Ao, J.; Li, M.; Wang, J.; Luo, A. Juglone inhibits Listeria monocytogenes ATCC 19115 by targeting cell membrane and protein. Foods 2022, 11, 2558. [Google Scholar] [CrossRef]
- Bai, J.; Wu, Y.; Liu, X.; Zhong, K.; Huang, Y.; Gao, H. Antibacterial activity of shikimic acid from pine needles of Cedrus deodara against Staphylococcus aureus through damage to cell membrane. Int. J. Mol. Sci. 2015, 16, 27145–27155. [Google Scholar] [CrossRef]
- Ouyang, Q.; Tao, N.; Zhang, M. A Damaged Oxidative Phosphorylation Mechanism Is Involved in the Antifungal Activity of Citral against Penicillium digitatum. Front. Microbiol. 2018, 9, 239. [Google Scholar] [CrossRef]
- Ju, J.; Xie, Y.; Yu, H.; Guo, Y.; Cheng, Y.; Qian, H.; Yao, W. Analysis of the synergistic antifungal mechanism of eugenol and citral. Lwt 2020, 123, 109128. [Google Scholar] [CrossRef]
- Berry, B.J.; Trewin, A.J.; Amitrano, A.M.; Kim, M.; Wojtovich, A.P. Use the Protonmotive Force: Mitochondrial Uncoupling and Reactive Oxygen Species. J. Mol. Biol. 2018, 430, 3873–3891. [Google Scholar] [CrossRef]
- Chen, J.; Wang, H.; Chen, Y.; Zhu, Q.; Wan, J. Inhibitive effect and mechanism of cinnamaldehyde on growth and OTA production of Aspergillus niger in vitro and in dried red chilies. Food Res. Int. 2023, 168, 112794. [Google Scholar] [CrossRef]
- Hu, Z.; Yuan, K.; Zhou, Q.; Lu, C.; Du, L.; Liu, F. Mechanism of antifungal activity of Perilla frutescens essential oil against Aspergillus flavus by transcriptomic analysis. Food Control 2021, 123, 107703. [Google Scholar] [CrossRef]
- Li, S.-F.; Zhang, S.-B.; Lv, Y.-Y.; Zhai, H.-C.; Hu, Y.-S.; Cai, J.-P. Heptanal inhibits the growth of Aspergillus flavus through disturbance of plasma membrane integrity, mitochondrial function and antioxidant enzyme activity. LWT 2022, 154, 112655. [Google Scholar] [CrossRef]
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial electron transport chain: Oxidative phosphorylation, oxidant production, and methods of measurement. Redox Biol. 2020, 37, 101674. [Google Scholar] [CrossRef]
- Kaim, G. ATP synthesis by F-type ATP synthase is obligatorily dependent on the transmembrane voltage. EMBO J. 1999, 18, 4118–4127. [Google Scholar] [CrossRef]
- Li, X.-N.; Zhang, L.-M.; Wang, Y.-Y.; Zhang, Y.; Jin, Z.-H.; Li, J.; Wang, R.-R.; Xiao, W.-L. SWL-1 Reverses Fluconazole Resistance in Candida albicans by Regulating the Glycolytic Pathway. Front. Microbiol. 2020, 11, 572608. [Google Scholar] [CrossRef]
- Zhou, Y.; Yan, K.; Qin, Q.; Raimi, O.G.; Du, C.; Wang, B.; Ahamefule, C.S.; Kowalski, B.; Jin, C.; van Aalten, D.M.F.; et al. Phosphoglucose Isomerase Is Important for Aspergillus fumigatus Cell Wall Biogenesis. mBio 2022, 13, e01426-22. [Google Scholar] [CrossRef]
- Krivoruchko, A.; Zhang, Y.; Siewers, V.; Chen, Y.; Nielsen, J. Microbial acetyl-CoA metabolism and metabolic engineering. Metab. Eng. 2015, 28, 28–42. [Google Scholar] [CrossRef]
- Akram, M. Citric Acid Cycle and Role of its Intermediates in Metabolism. Cell Biochem. Biophys. 2013, 68, 475–478. [Google Scholar] [CrossRef]
- Li, Y.; Shao, X.; Xu, J.; Wei, Y.; Xu, F.; Wang, H. Tea tree oil exhibits antifungal activity against Botrytis cinerea by affecting mitochondria. Food Chem. 2017, 234, 62–67. [Google Scholar] [CrossRef]
- Ju, J.; Guo, Y.; Cheng, Y.; Yaoc, W. Analysis of the synergistic antifungal mechanism of small molecular combinations of essential oils at the molecular level. Ind. Crops Prod. 2022, 188, 115612. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, Y.; Xie, Y. Paeonol Disrupts the Integrity of Aspergillus flavus Cell Walls via Releasing Surface Proteins, Inhibiting the Biosynthesis of β-1,3-Glucan and Promoting the Degradation of Chitin, and an Identification of Cell Surface Proteins. Foods 2021, 10, 2951. [Google Scholar] [CrossRef]
- Ouyang, Q.; Duan, X.; Li, L.; Tao, N. Cinnamaldehyde Exerts Its Antifungal Activity by Disrupting the Cell Wall Integrity of Geotrichum citri-aurantii. Front. Microbiol. 2019, 10, 55. [Google Scholar] [CrossRef]
- Bie, X.; Zhang, S.; Luo, X.; Qi, R.-Q. Candida albicans cell wall mannoprotein synergizes with lipopolysaccharide to affect RAW264.7 proliferation, phagocytosis and apoptosis. Microb. Pathog. 2019, 131, 98–105. [Google Scholar] [CrossRef]
- Immervoll, T.; Gentzsch, M.; Tanner, W. PMT3 and PMT4, two new members of the protein-O-mannosyltransferase gene family of Saccharomyces cerevisiae. Yeast 2004, 11, 1345–1351. [Google Scholar] [CrossRef]
- Lin, X.; Willger, S.D.; Ernst, J.F.; Alspaugh, J.A.; Lengeler, K.B. Characterization of the PMT Gene Family in Cryptococcus neoformans. PLoS ONE 2009, 4, e6321. [Google Scholar]
- Mizunuma, M. GSK-3 kinase Mck1 and calcineurin coordinately mediate Hsl1 down-regulation by Ca2+ in budding yeast. EMBO J. 2001, 20, 1074–1085. [Google Scholar] [CrossRef] [PubMed]
- Tatebayashi, K.; Tanaka, K.; Yang, H.-Y.; Yamamoto, K.; Matsushita, Y.; Tomida, T.; Imai, M.; Saito, H. Transmembrane mucins Hkr1 and Msb2 are putative osmosensors in the SHO1 branch of yeast HOG pathway. EMBO J. 2007, 26, 3521–3533. [Google Scholar] [CrossRef]
- Hameed, S. Protein kinases as potential anticandidal drug targets. Front. Biosci. 2020, 25, 1412–1432. [Google Scholar] [CrossRef]
- Clotet, J.; Escoté, X.; Adrover, M.À.; Yaakov, G.; Garí, E.; Aldea, M.; de Nadal, E.; Posas, F. Phosphorylation of Hsl1 by Hog1 leads to a G2 arrest essential for cell survival at high osmolarity. EMBO J. 2006, 25, 2338–2346. [Google Scholar] [CrossRef]
- Zhang, X.-Z.; Wang, L.; Liu, D.-W.; Tang, G.-Y.; Zhang, H.-Y. Synergistic Inhibitory Effect of Berberine and d-Limonene on Human Gastric Carcinoma Cell Line MGC803. J. Med. Food 2014, 17, 955–962. [Google Scholar] [CrossRef]
- Zore, G.B.; Thakre, A.D.; Jadhav, S.; Karuppayil, S.M. Terpenoids inhibit Candida albicans growth by affecting membrane integrity and arrest of cell cycle. Phytomedicine 2011, 18, 1181–1190. [Google Scholar] [CrossRef]
- Sun, F.J.; Li, M.; Gu, L.; Wang, M.L.; Yang, M.H. Recent progress on anti-Candida natural products. Chin. J. Nat. Med. 2021, 19, 561–579. [Google Scholar] [CrossRef]
- Hu, Z.; He, B.; Ma, L.; Sun, Y.; Niu, Y.; Zeng, B. Recent Advances in Ergosterol Biosynthesis and Regulation Mechanisms in Saccharomyces cerevisiae. Indian J. Microbiol. 2017, 57, 270–277. [Google Scholar] [CrossRef]
- Ouyang, Q.; Tao, N.; Jing, G. Transcriptional profiling analysis of Penicillium digitatum, the causal agent of citrus green mold, unravels an inhibited ergosterol biosynthesis pathway in response to citral. BMC Genom. 2016, 17, 599. [Google Scholar] [CrossRef]
- Bahn, Y.-S.; Sun, L.; Liao, K.; Wang, D. Honokiol induces superoxide production by targeting mitochondrial respiratory chain complex I in Candida albicans. PLoS ONE 2017, 12, e0184003. [Google Scholar]
- Kim, H.; Lee, D.G. Naringin-generated ROS promotes mitochondria-mediated apoptosis in Candida albicans. IUBMB Life 2021, 73, 953–967. [Google Scholar] [CrossRef] [PubMed]
- Lightowlers, B.; Tian, J.; Ban, X.; Zeng, H.; He, J.; Chen, Y.; Wang, Y. The Mechanism of Antifungal Action of Essential Oil from Dill (Anethum graveolens L.) on Aspergillus flavus. PLoS ONE 2012, 7, e30147. [Google Scholar]
- Xu, Y.-D.; Guo, Y.-J.; Mao, H.-R.; Xiong, Z.-X.; Luo, M.-Y.; Luo, R.-Q.; Lu, S.; Huang, L.; Hong, Y. Integration of transcriptomics and proteomics to elucidate inhibitory effect and mechanism of rosmarinic acid from Perilla frutescens (L.) Britt. in treating Trichophyton mentagrophytes. Chin. Med. 2023, 18, 67. [Google Scholar] [CrossRef] [PubMed]
- Jainkittivong, A.; Butsarakamruha, T.; Langlais, R.P. Antifungal activity of Morinda citrifolia fruit extract against Candida albicans. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodontol. 2009, 108, 394–398. [Google Scholar] [CrossRef]
- Mandras, N.; Roana, J.; Scalas, D.; Del Re, S.; Cavallo, L.; Ghisetti, V.; Tullio, V. The Inhibition of Non-albicans Candida Species and Uncommon Yeast Pathogens by Selected Essential Oils and Their Major Compounds. Molecules 2021, 26, 4937. [Google Scholar] [CrossRef] [PubMed]
- Costa, W.K.; de Oliveira, A.M.; da Silva Santos, I.B.; Guimaraes Silva, V.B.; Carvalho da Silva, E.K.; de Oliveira Alves, J.V.; Sant’Anna da Silva, A.P.; de Menezes Lima, V.L.; dos Santos Correia, M.T.; da Silva, M.V. Antibacterial mechanism of Eugenia stipitata McVaugh essential oil and synergistic effect against Staphylococcus aureus. S. Afr. J. Bot. 2022, 147, 724–730. [Google Scholar] [CrossRef]
- Wu, Y.X.; Zhang, Y.D.; Li, N.; Wu, D.D.; Li, Q.M.; Chen, Y.Z.; Zhang, G.C.; Yang, J. Inhibitory effect and mechanism of action of juniper essential oil on gray mold in cherry tomatoes. Front. Microbiol. 2022, 13, 1000526. [Google Scholar] [CrossRef]
- Wang, P.; Yu, A.; Ji, X.; Mu, Q.; Salman Haider, M.; Wei, R.; Leng, X.; Fang, J. Transcriptome and metabolite integrated analysis reveals that exogenous ethylene controls berry ripening processes in grapevine. Food Res. Int. 2022, 155, 111084. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, C.; Sun, Y.; Lin, H.; Li, Z.; Zhang, Q.; Cai, T.; Xiang, W.; Tang, J.; Yasurin, P. D-Limonene Inhibits Pichia kluyveri Y-11519 in Sichuan Pickles by Disrupting Metabolism. Molecules 2024, 29, 3561. https://doi.org/10.3390/molecules29153561
Zeng C, Sun Y, Lin H, Li Z, Zhang Q, Cai T, Xiang W, Tang J, Yasurin P. D-Limonene Inhibits Pichia kluyveri Y-11519 in Sichuan Pickles by Disrupting Metabolism. Molecules. 2024; 29(15):3561. https://doi.org/10.3390/molecules29153561
Chicago/Turabian StyleZeng, Chaoyi, Yue Sun, Haoran Lin, Ziyu Li, Qing Zhang, Ting Cai, Wenliang Xiang, Jie Tang, and Patchanee Yasurin. 2024. "D-Limonene Inhibits Pichia kluyveri Y-11519 in Sichuan Pickles by Disrupting Metabolism" Molecules 29, no. 15: 3561. https://doi.org/10.3390/molecules29153561
APA StyleZeng, C., Sun, Y., Lin, H., Li, Z., Zhang, Q., Cai, T., Xiang, W., Tang, J., & Yasurin, P. (2024). D-Limonene Inhibits Pichia kluyveri Y-11519 in Sichuan Pickles by Disrupting Metabolism. Molecules, 29(15), 3561. https://doi.org/10.3390/molecules29153561