
A Mitochondria-Targeting Fluorescent Probe for the Dual Sensing of Hypochlorite and Viscosity without Signal Crosstalk in Living Cells and Zebrafish


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
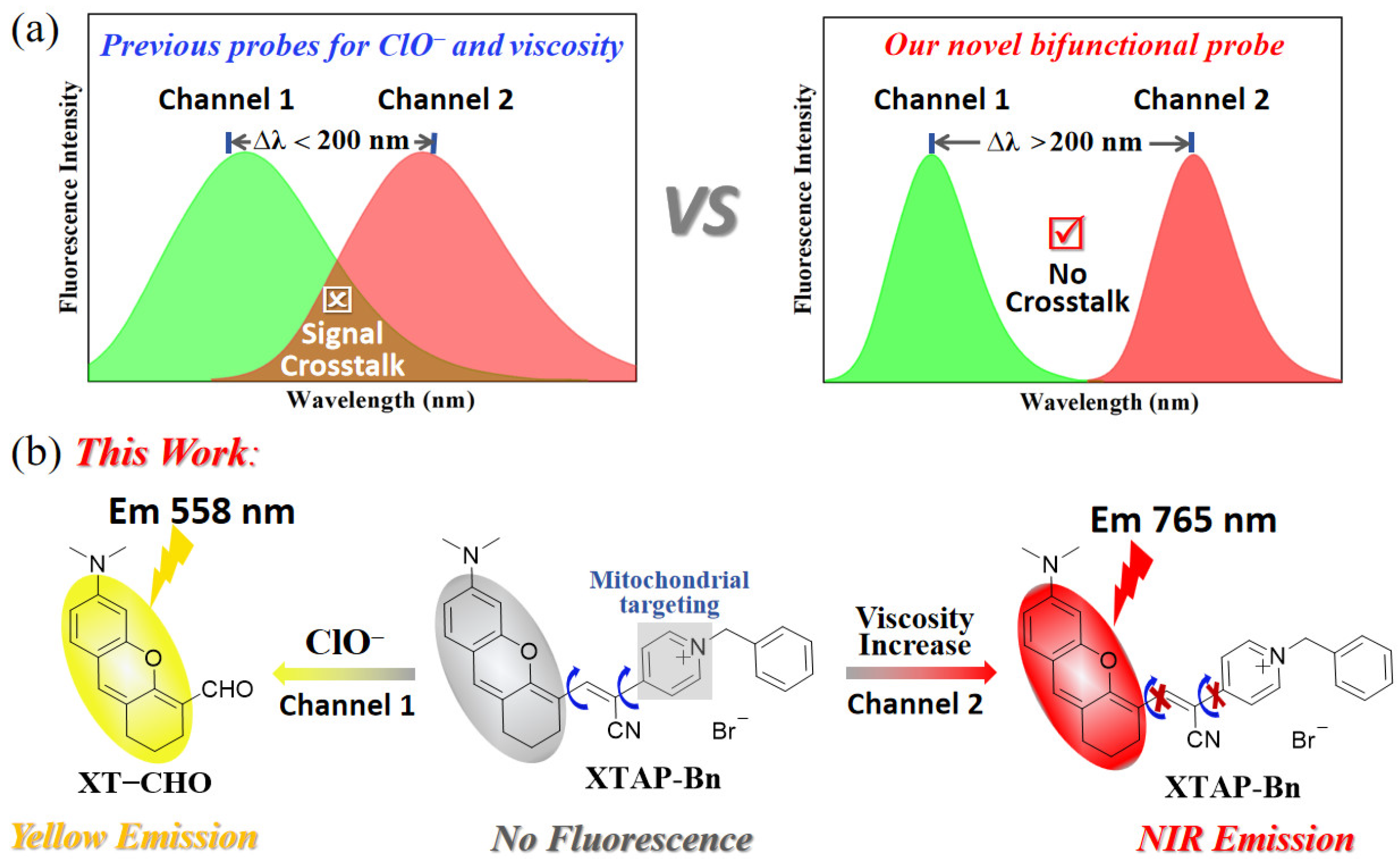
2. Results and Discussion
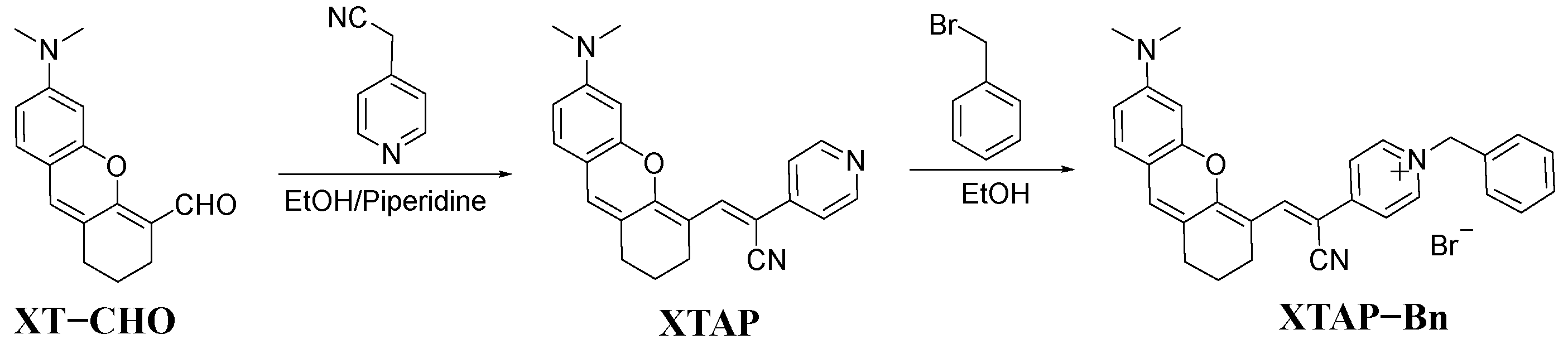
2.1. Synthesis and Characterization of Probe XTAP–Bn
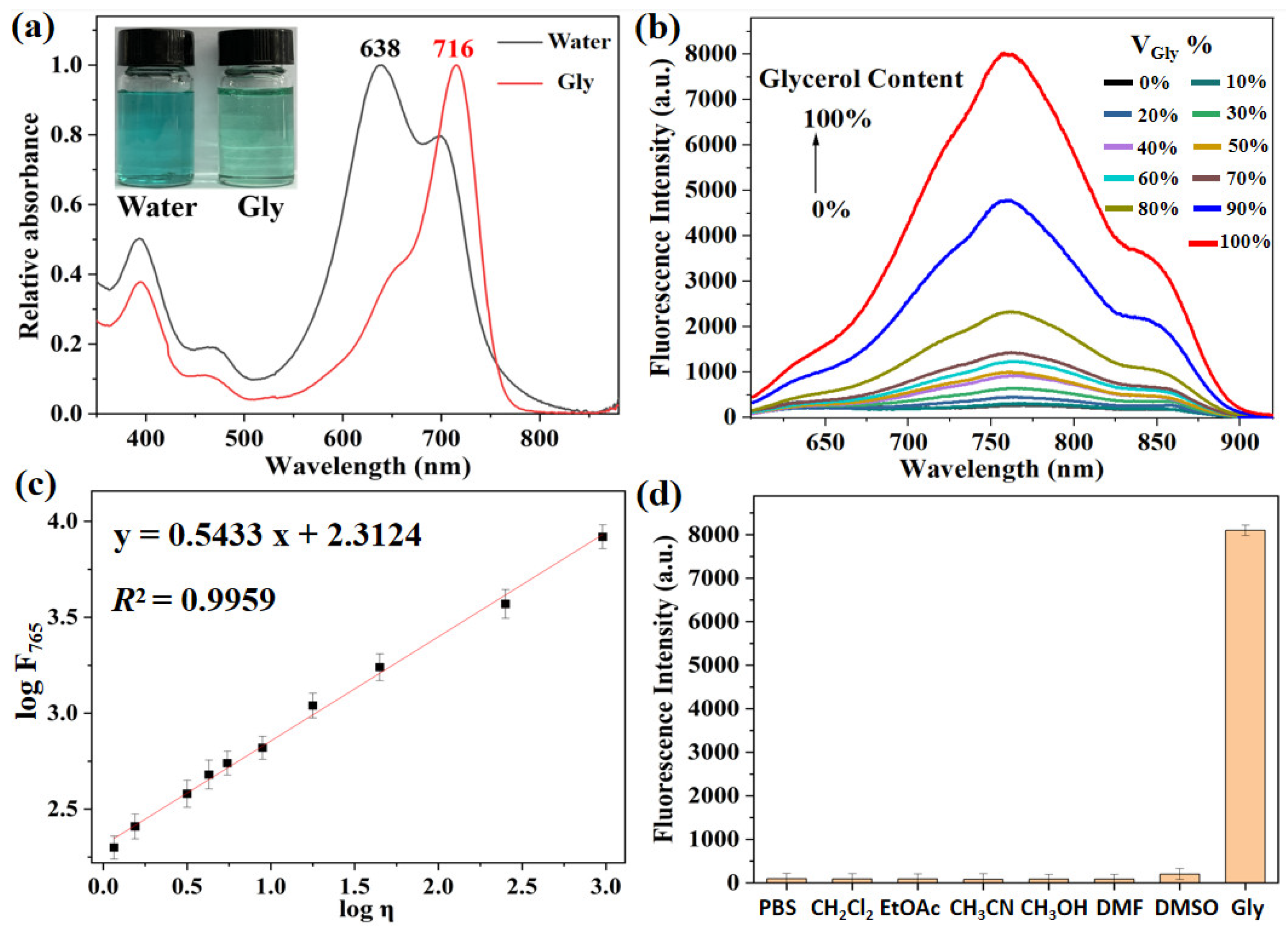
2.2. Spectroscopic Response to Viscosity
2.3. Spectroscopic Response to ClO−
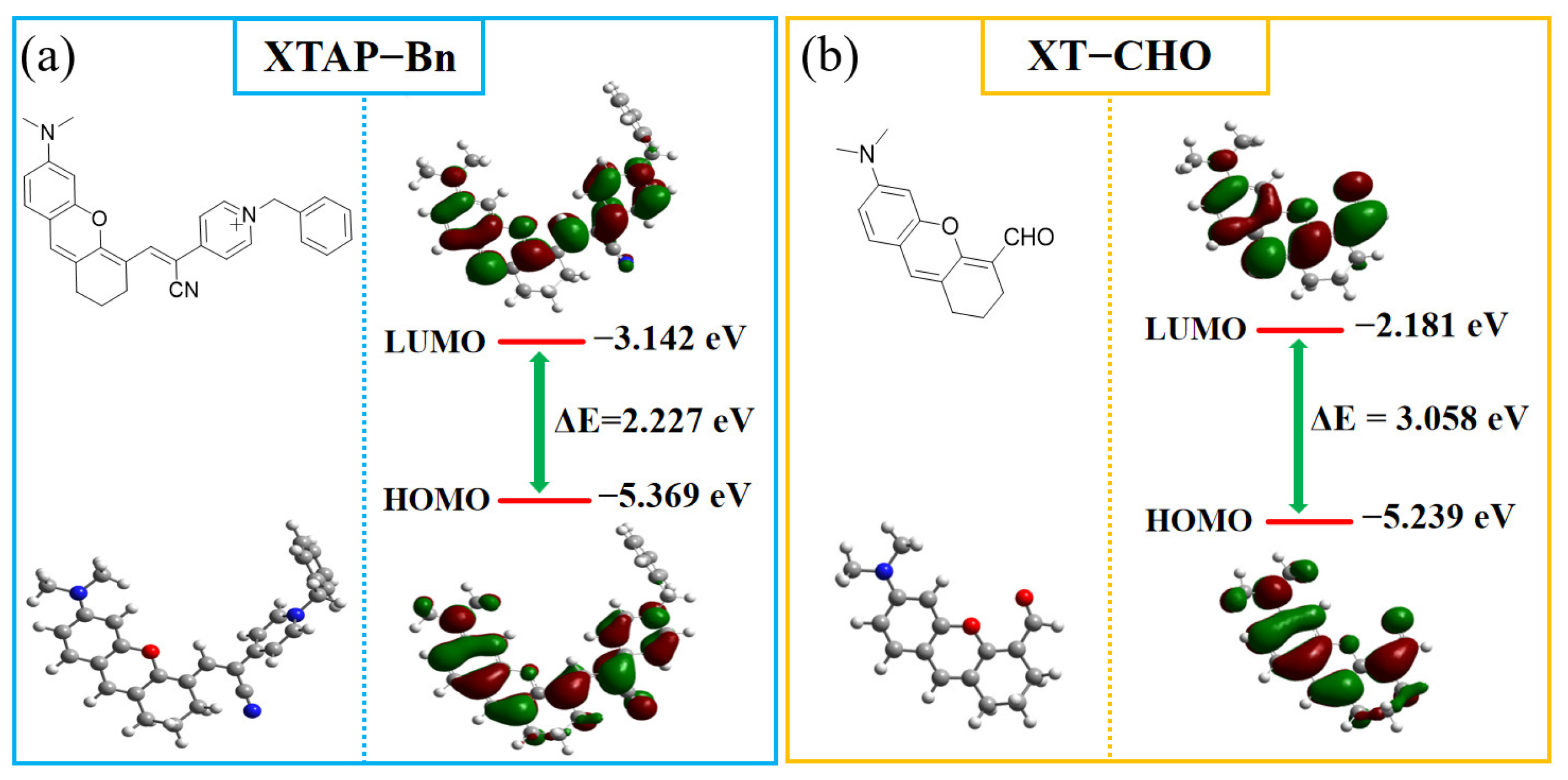
2.4. Sensing Mechanism
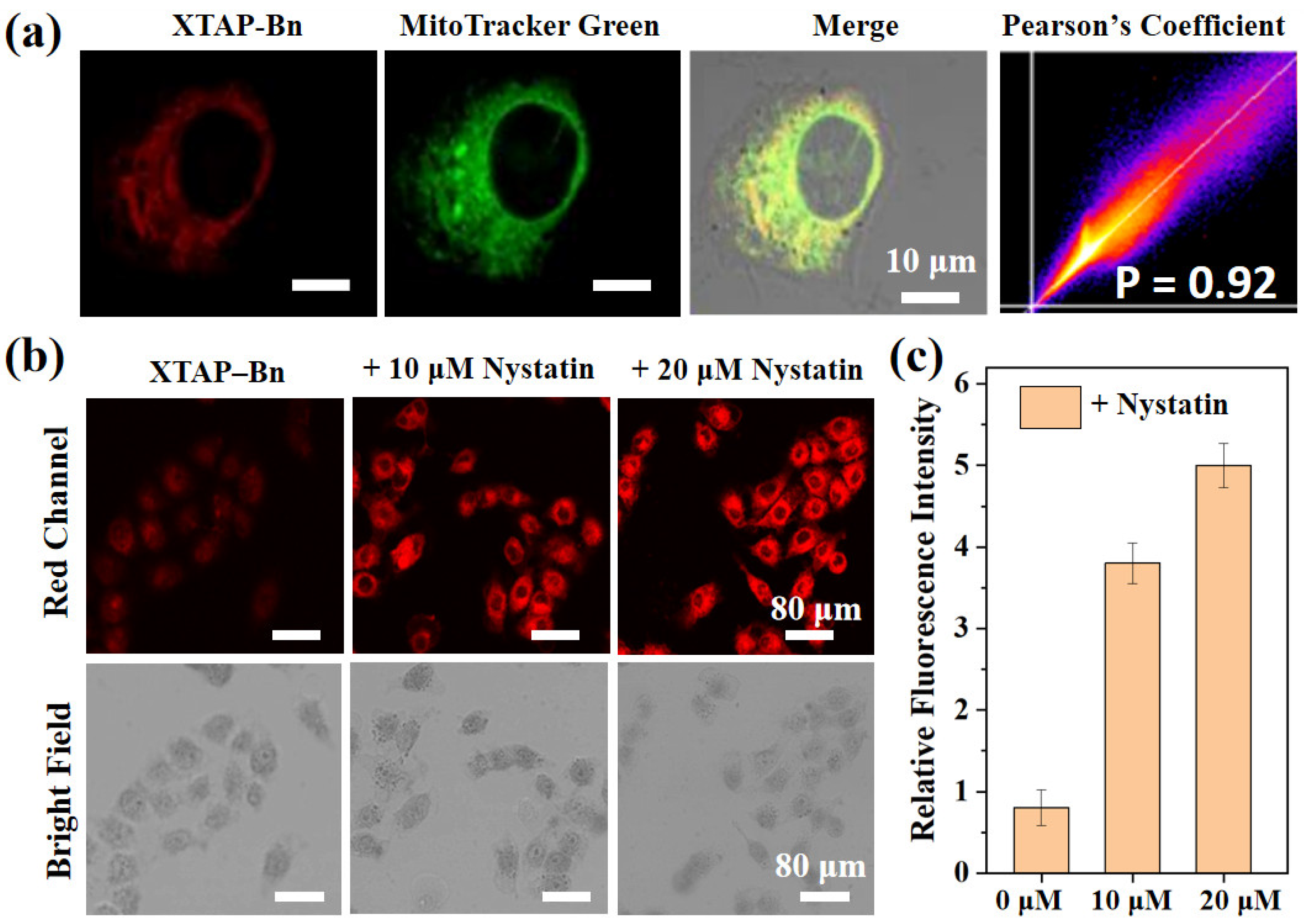
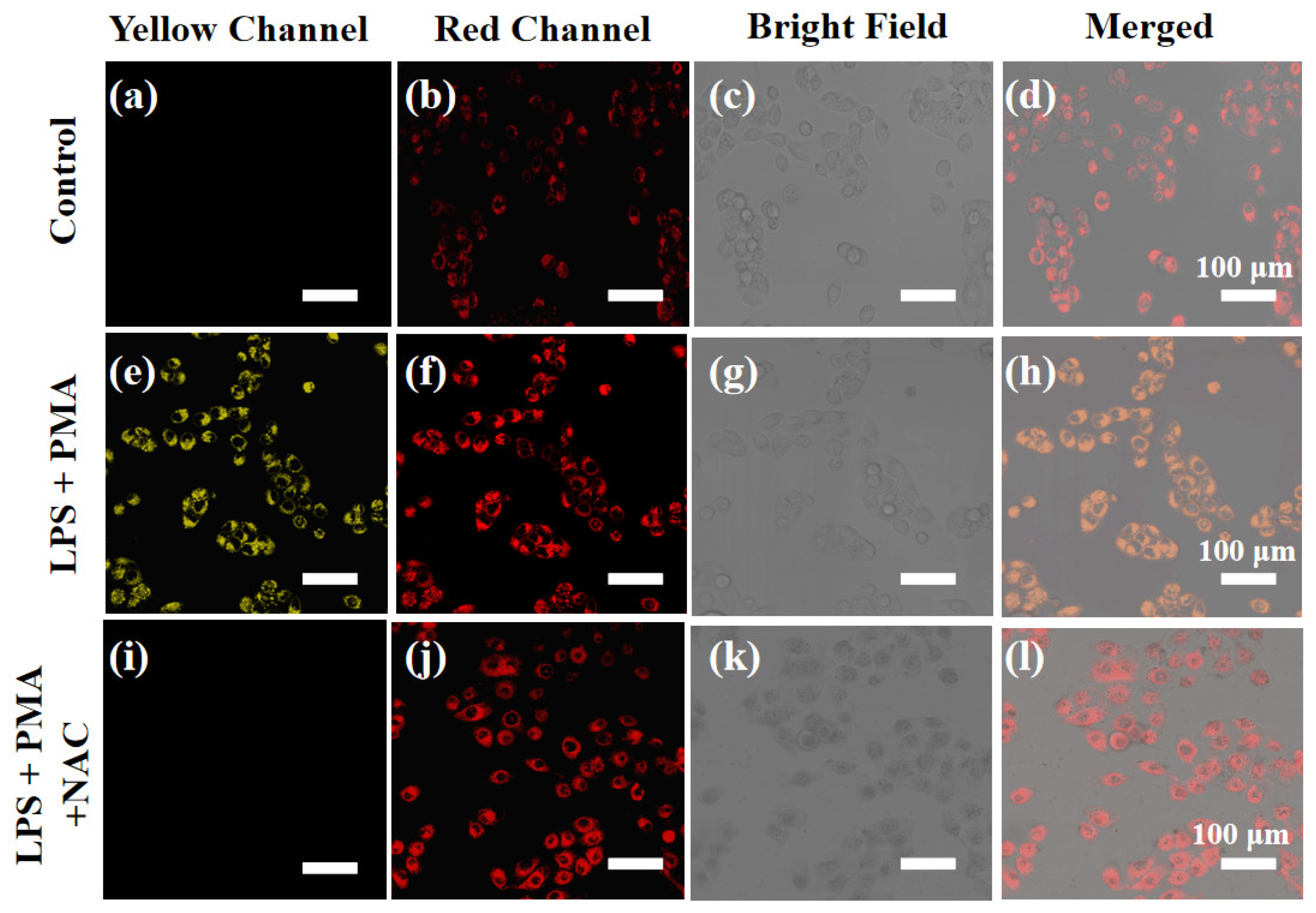
2.5. Cell Imaging
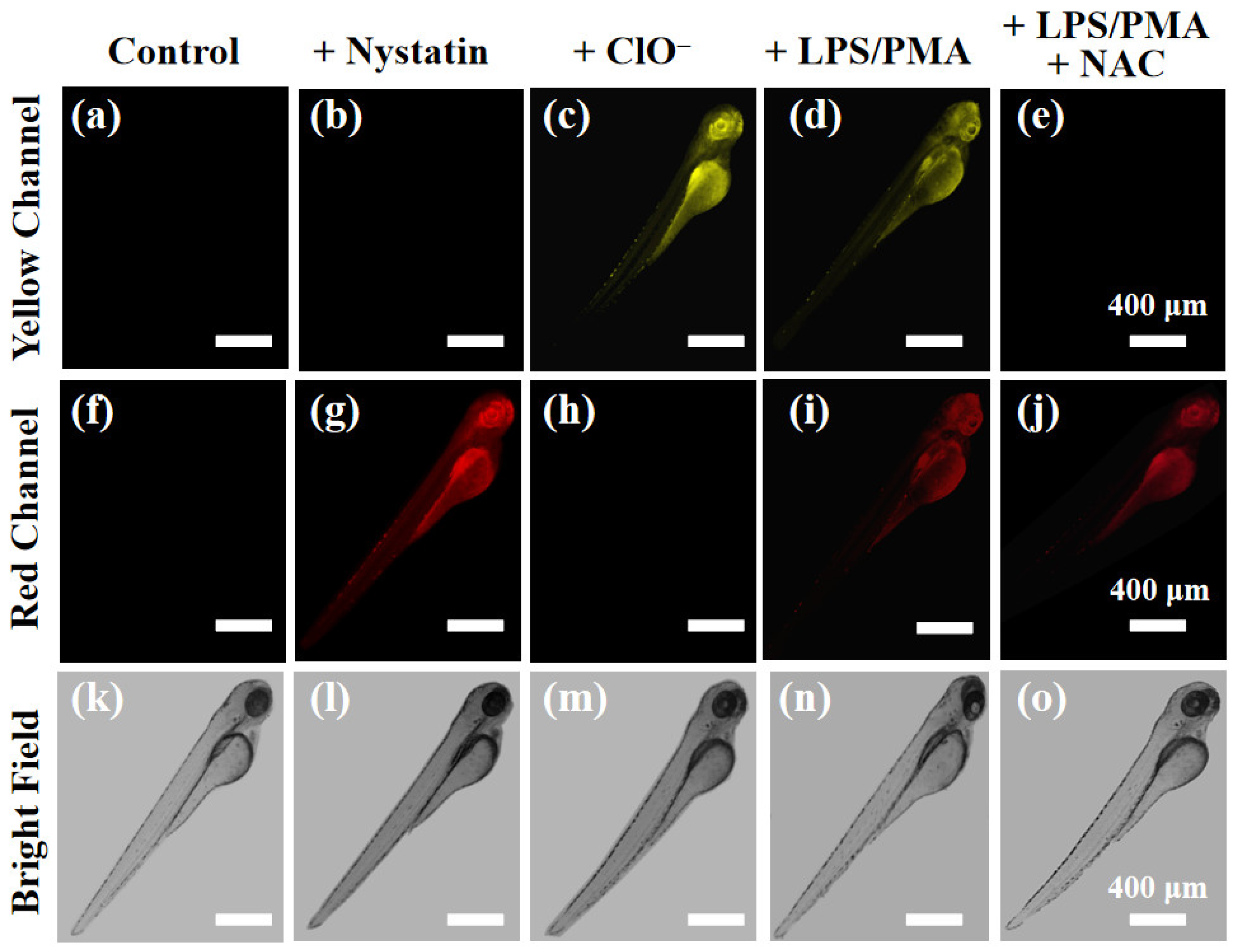
2.6. Zebrafish Imaging
3. Materials and Methods
3.1. Materials and Instruments
3.2. Synthesis of Compound XTAP
3.3. Synthesis of Probe XTAP–Bn
3.4. Optical Study
3.5. Calculation of the Detection Limit
3.6. Theoretical Calculations
3.7. Acytotoxicity Assay
3.8. Living Cell Imaging
3.9. Zebrafish Imaging
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bazhin, A.V. Mitochondria and cancer. Cancers 2020, 12, 2641. [Google Scholar] [CrossRef] [PubMed]
- Katherine, L.P. Cytoarchitecture and physical properties of cytoplasm: Volume, viscosity, diffusion, intracellular surface area. Int. Rev. Cytol. 1999, 192, 189–221. [Google Scholar]
- Meszaros, A.V.; Weidinger, A.; Dorighello, G.; Boros, M.; Redl, H.; Kozlov, A.V. The impact of inflammatory cytokines on liver damage caused by elevated generation of mitochondrial reactive oxygen species. Free Radic. Biol. Med. 2016, 100, S57. [Google Scholar] [CrossRef]
- Ramanathan, R.; Ali, A.H.; Ibdah, J.A. Mitochondrial dysfunction plays central role in nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2022, 23, 7280. [Google Scholar] [CrossRef]
- Gyasi, Y.I.; Pang, Y.P.; Li, X.R.; Gu, J.X.; Cheng, X.J.; Liu, J.; Xu, T.; Liu, Y. Biological applications of near infrared fluorescence dye probes in monitoring Alzheimer’s disease. Eur. J. Med. Chem. 2020, 187, 111982. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Cao, Z.; Moore, D.R.; Jackson, P.L.; Barnes, S.; Lambeth, J.D.; Thannickal, V.J.; Cheng, G. Microbicidal activity of vascular peroxidase 1 in human plasma via generation of hypochlorous acid. Infect. Immun. 2012, 80, 2528–2537. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. Reconciling the chemistry and biology of reactive oxygen species. Nat. Chem. Biol. 2008, 4, 278–286. [Google Scholar] [CrossRef]
- Andersen, J.K. Oxidative stress in neurodegeneration: Cause or consequence. Nat. Med. 2004, 10, 18–25. [Google Scholar] [CrossRef]
- Ramsey, M.R.; Sharpless, N.E. ROS as a tumour suppressor. Nat. Cell. Biol. 2006, 8, 1213–1215. [Google Scholar] [CrossRef]
- Yu, X.; Huang, Y.; Tao, Y.; Fan, L.; Zhang, Y. Mitochondria-targetable small molecule fluorescent probes for the detection of cancer-associated biomarkers: A review. Anal. Chim. Acta 2024, 1289, 342060. [Google Scholar] [CrossRef]
- Fan, G.; Zhang, B.; Wang, J.; Wang, N.; Qin, S.; Zhao, W.; Zhang, J. Accurate construction of NIR probe for visualizing HClO fluctuations in type I, type II diabetes and diabetic liver disease assisted by theoretical calculation. Talanta 2024, 268, 125298. [Google Scholar] [CrossRef]
- Yuan, F.; Wang, B.; Hou, J.T.; Li, J.; Shen, J.; Duan, Y.; Ren, W.X.; Wang, S. Demonstrating HOCl as a potential biomarker for liver fibrosis using a highly sensitive fluorescent probe. Sens. Actuators B Chem. 2023, 378, 133219. [Google Scholar] [CrossRef]
- Shao, S.; Yang, T.; Han, Y. A TICT-based fluorescent probe for hypochlorous acid and its application to cellular and zebrafish imaging. Sens. Actuators B Chem. 2023, 392, 134041. [Google Scholar] [CrossRef]
- Liang, F.; Jiang, J.; Yang, X.; Zhang, G.; Zhou, J.; Han, J.; Geng, Y.; Wang, Z. Si-rhodamine fluorescent probe for monitoring of hypochlorous acid in the brains of mice afflicted with neuroinflammation. Chem. Commun. 2023, 59, 1357–1360. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Chen, Y.; Geng, S.; Yao, S.; Guo, Z.; He, W. Super-resolution imaging of mitochondrial HClO during cell ferroptosis using a near-infrared fluorescent probe. Anal. Chem. 2022, 94, 17904–17912. [Google Scholar] [CrossRef] [PubMed]
- Shangguan, L.; Wang, J.; Qian, X.; Wu, Y.; Liu, Y. Mitochondria-targeted ratiometric chemdosimeter to detect hypochlorite acid for monitoring the drug-damaged liver and kidney. Anal. Chem. 2022, 94, 11881–11888. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Kim, S.J.; Singha, S.; Yang, Y.J.; Park, S.K.; Ahn, K.H. Ratiometric detection of hypochlorous acid in brain tissues of neuroinflammation and maternal immune activation models with a deep-red/near-infrared emitting probe. ACS Sens. 2021, 6, 3253–3261. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhou, Y.; Li, Z.; Zhou, Y.; Liu, X.; Peng, X. Rational design of AIE-based fluorescent probes for hypochlorite detection in real water samples and live cell imaging. J. Hazard. Mater. 2021, 418, 126243. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yuan, S.; Qiao, M.; Jin, X.; Chen, J.; Guo, L.; Su, J.; Qu, D.H.; Zhang, Z. Exploring the depth-dependent microviscosity inside a micelle using butterfly-motion-based fluorescent probes. J. Am. Chem. Soc. 2023, 145, 26494–26503. [Google Scholar] [CrossRef]
- Ge, J.; Cai, W.; Niu, N.; Wen, Y.; Wu, Q.; Wang, L.; Wang, D.; Tang, B.Z.; Zhang, R. Viscosity-responsive NIR-II fluorescent probe with aggregation-induced emission features for early diagnosis of liver injury. Biomaterials 2023, 300, 122190. [Google Scholar] [CrossRef]
- Hong, J.; Guan, X.; Chen, Y.; Tan, X.; Zhang, S.; Feng, G. Mitochondrial membrane potential independent near-infrared mitochondrial viscosity probes for real-time tracking mitophagy. Anal. Chem. 2023, 95, 5687–5694. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Leng, H.; Chen, Q.; Su, J.; Shi, W.; Xia, C.; Zhang, L.; Yan, J. Development of novel near-infrared GFP chromophore-based fluorescent probes for imaging of amyloid-β plaque and viscosity. Sens. Actuators B Chem. 2022, 372, 132648. [Google Scholar] [CrossRef]
- Wu, Y.; Yin, C.; Zhang, W.; Zhang, Y.; Huo, F. Mitochondrial-targeting near-infrared fluorescent probe for visualizing viscosity in drug-induced cells and a fatty liver mouse model. Anal. Chem. 2022, 94, 5069–5074. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Liu, S.; Chen, Z.; Wu, F.; Cao, W.; Tian, Y.; Xiong, H. Bichromatic imaging with hemicyanine fluorophores enables simultaneous visualization of non-alcoholic fatty liver disease and metastatic intestinal cancer. Anal. Chem. 2022, 94, 13556–13565. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhang, W.; Zhang, Y.; Yin, C.; Huo, F. Viscosity activated NIR fluorescent probe for visualizing mitochondrial viscosity dynamic and fatty liver mice. Chem. Eng. J. 2022, 445, 136448. [Google Scholar] [CrossRef]
- Xu, S.L.; Guo, F.F.; Xu, Z.H.; Wang, Y.; James, T.D. A hemicyanine-based fluorescent probe for ratiometric detection of ClO– and turn-on detection of viscosity and its imaging application in mitochondria of living cells and zebrafish. Sens. Actuators B Chem. 2023, 383, 133510. [Google Scholar] [CrossRef]
- Chai, L.; Li, Y.; Yang, H.; Wang, Y.; Huang, R.; Wei, Z.; Zhan, Z. pH-triggered fluorescent probe for sensing of hypochlorite and viscosity in live cells and chronic wound diabetic mice. Sens. Actuators B Chem. 2023, 393, 134345. [Google Scholar] [CrossRef]
- Huang, X.; Luo, T.; Zhang, C.; Li, J.; Jia, Z.; Chen, X.; Hu, Y.; Huang, H. Dual-ratiometric fluorescence probe for viscosity and hypochlorite based on AIEgen with mitochondria-targeting ability. Talanta 2022, 241, 123235. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Sun, Y.; Liu, C.; Jiao, X.; Shang, Y.; Zeng, X.; Zhao, L.; Zhao, J. Highly selective turn-on fluorescent probe for hypochlorite and viscosity detection. J. Mol. Struct. 2021, 1227, 129523. [Google Scholar] [CrossRef]
- Liu, T.Z.; Wang, S.; Xu, J.R.; Miao, J.Y.; Zhao, B.X.; Lin, Z.M. FRET-based fluorescent probe with favorable water solubility for simultaneous detection of SO2 derivatives and viscosity. Talanta 2023, 256, 124302. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, S.; Gong, S.; Feng, G. Dual-channel fluorescent probe for detecting viscosity and ONOO– without signal crosstalk in nonalcoholic fatty liver. Anal. Chem. 2022, 94, 17439–17447. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Xing, S.; Hu, T.; Chen, J.; Niu, Q.; Li, T. Bithiophene-benzothiazole fluorescent sensor for detecting hypochlorite in water, bio-fluids and bioimaging in living cells, plants and zebrafish. Sens. Actuators B Chem. 2023, 375, 132856. [Google Scholar] [CrossRef]
- Chao, J.J.; Zhang, H.; Wang, Z.Q.; Liu, Q.R.; Mao, G.J.; Chen, D.H.; Li, C.Y. A near-infrared fluorescent probe for monitoring abnormal mitochondrial viscosity in cancer and fatty-liver mice model. Anal. Chim. Acta 2023, 1242, 340813. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, C.; Chen, D.-D.; Zhang, L.; Ma, M.-L.; Liu, H.-W.; Cui, H.-R. A Mitochondria-Targeting Fluorescent Probe for the Dual Sensing of Hypochlorite and Viscosity without Signal Crosstalk in Living Cells and Zebrafish. Molecules 2024, 29, 3059. https://doi.org/10.3390/molecules29133059
Gao C, Chen D-D, Zhang L, Ma M-L, Liu H-W, Cui H-R. A Mitochondria-Targeting Fluorescent Probe for the Dual Sensing of Hypochlorite and Viscosity without Signal Crosstalk in Living Cells and Zebrafish. Molecules. 2024; 29(13):3059. https://doi.org/10.3390/molecules29133059
Chicago/Turabian StyleGao, Chao, Dan-Dan Chen, Lin Zhang, Ming-Lan Ma, Hu-Wei Liu, and Hai-Rong Cui. 2024. "A Mitochondria-Targeting Fluorescent Probe for the Dual Sensing of Hypochlorite and Viscosity without Signal Crosstalk in Living Cells and Zebrafish" Molecules 29, no. 13: 3059. https://doi.org/10.3390/molecules29133059
APA StyleGao, C., Chen, D.-D., Zhang, L., Ma, M.-L., Liu, H.-W., & Cui, H.-R. (2024). A Mitochondria-Targeting Fluorescent Probe for the Dual Sensing of Hypochlorite and Viscosity without Signal Crosstalk in Living Cells and Zebrafish. Molecules, 29(13), 3059. https://doi.org/10.3390/molecules29133059