
Carbohydrate-Binding Properties and Antimicrobial and Anticancer Potential of a New Lectin from the Phloem Sap of Cucurbita pepo
 , , , , ,
, , , , ,  ,
,  and
and 
Abstract

1. Introduction
2. Results
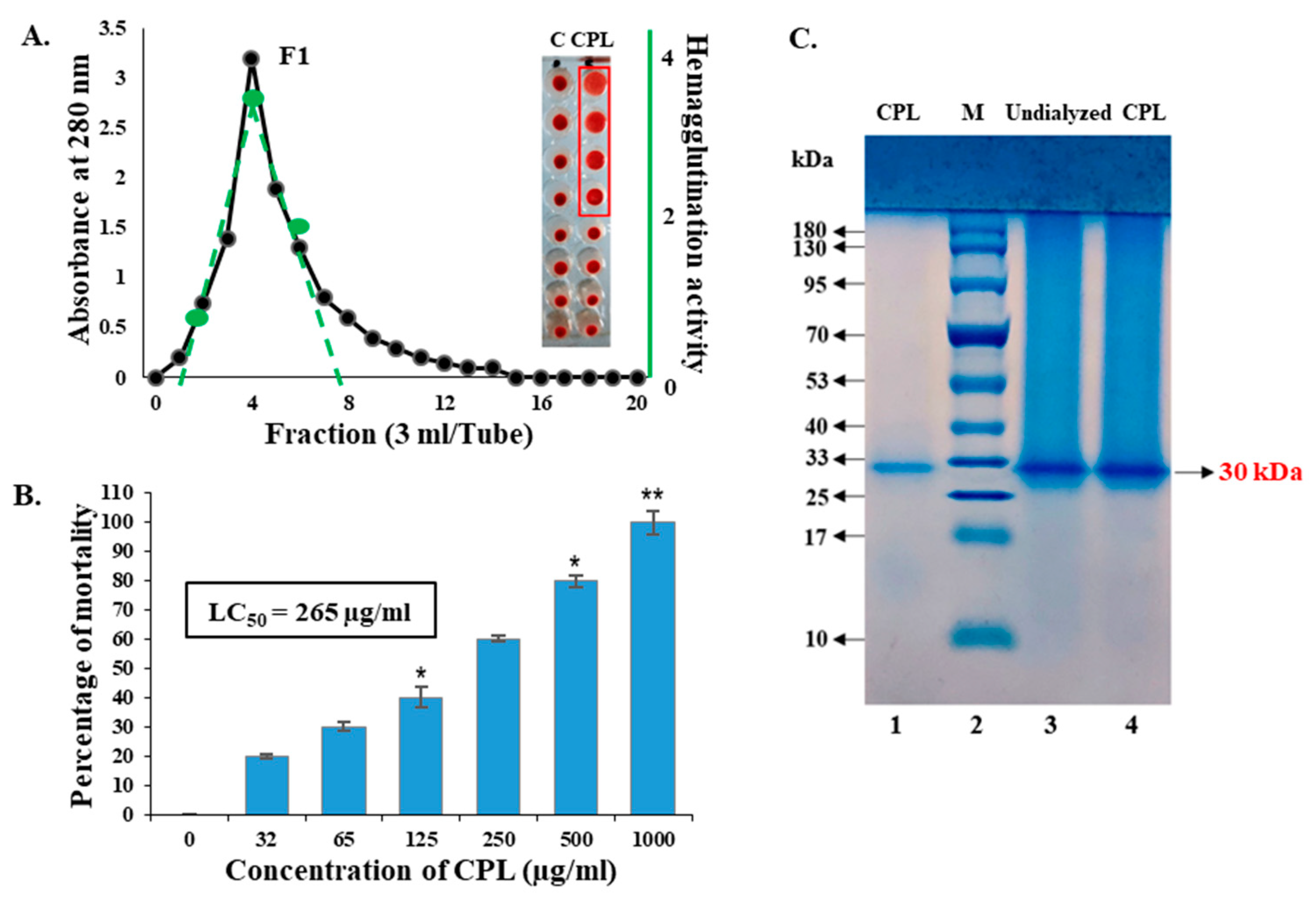
2.1. Purification of the Lectin and Its Minimum Hemagglutination Activity for Mice Erythrocytes
2.2. Toxicity of CPL against Brine Shrimp Nauplii
2.3. Effect of Temperature and pH on the Hemagglutination of CPL
2.4. Effect of Denaturants and Divalent Metal Ions on the Activity of CPL
2.5. Hemagglutination Inhibition Study of CPL by Various Sugars
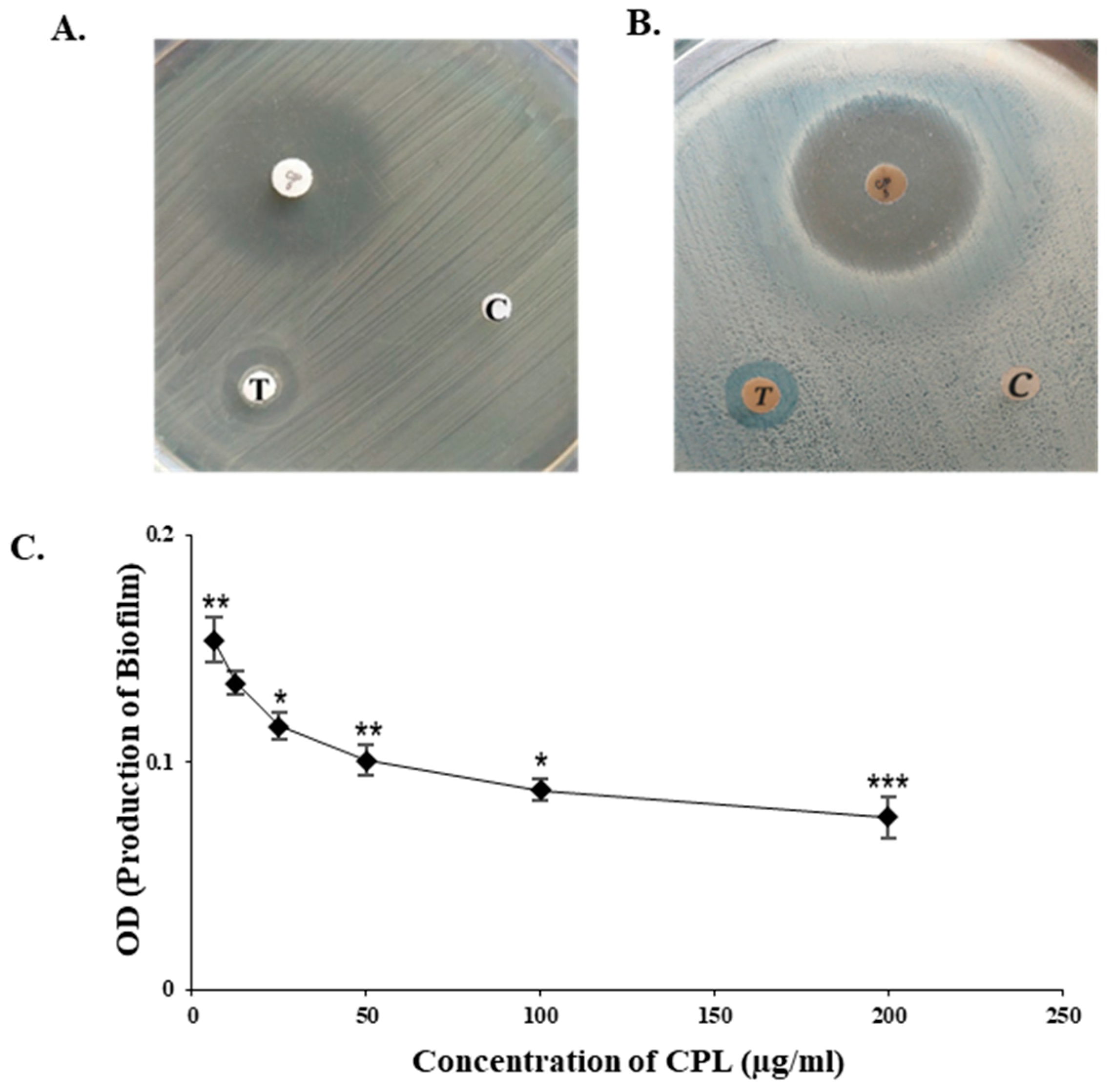
2.6. Bactericidal and Antibiofilm Activity of CPL
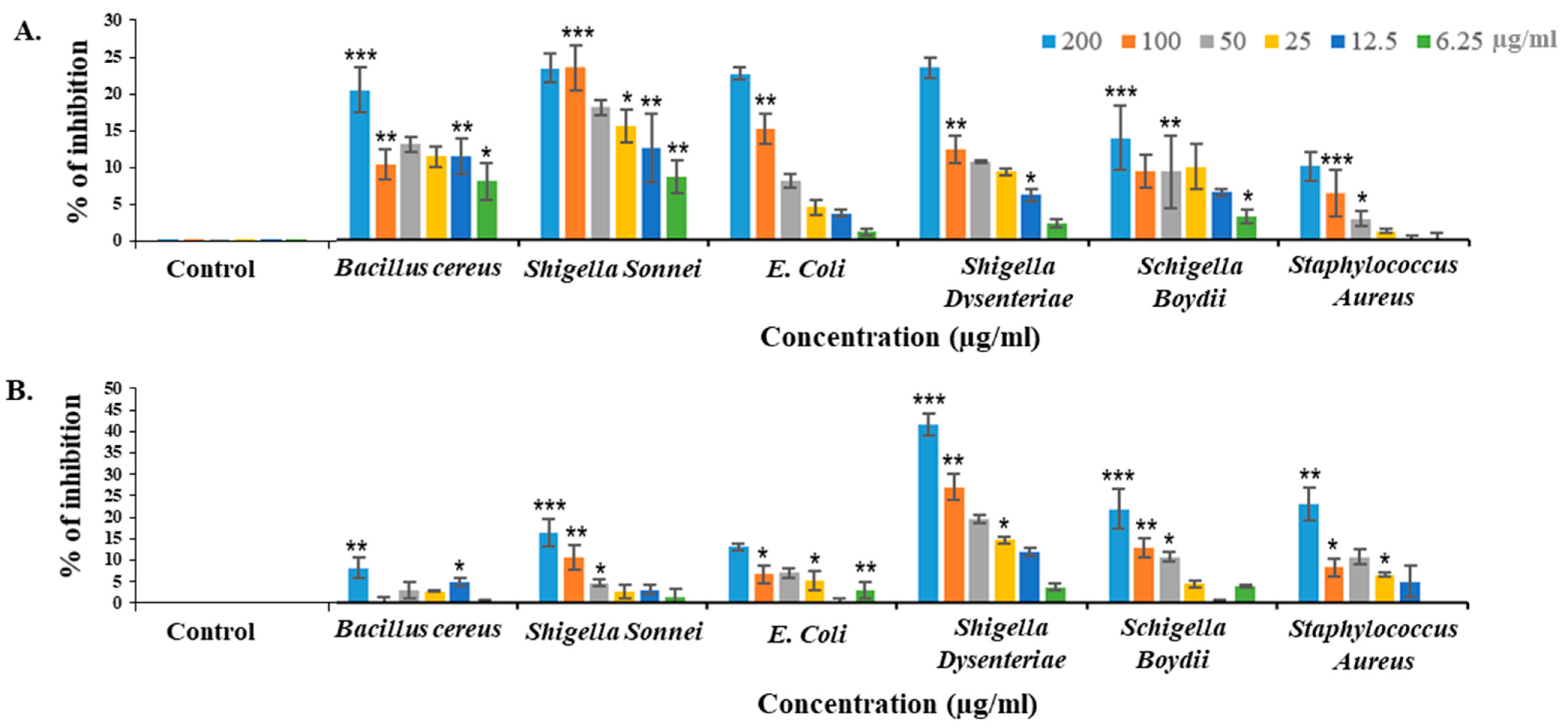
2.7. Bacteriostatic Activity of CPL against Different Pathogenic Bacteria
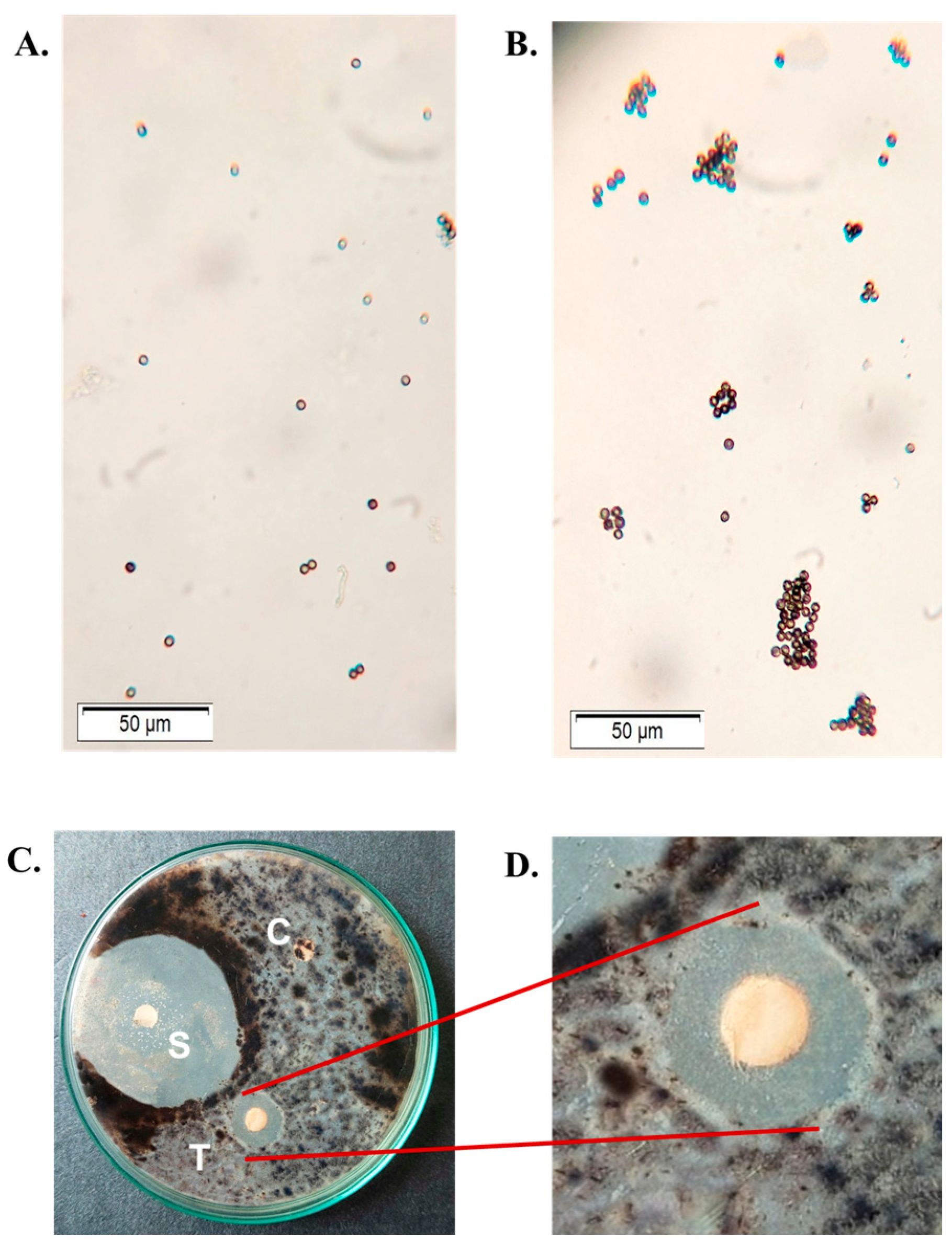
2.8. Antifungal Activity of CPL
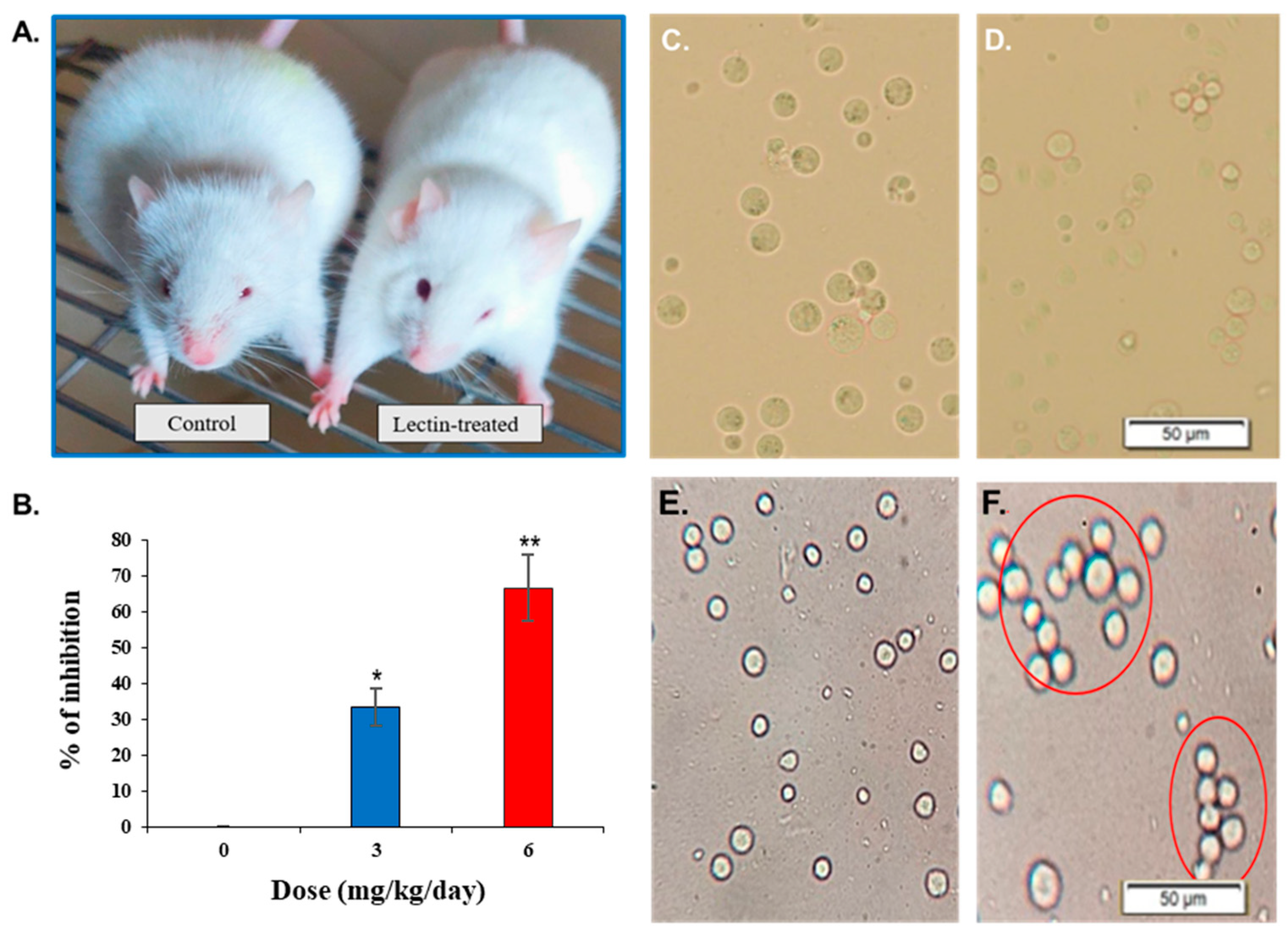
2.9. Agglutination of Ehrlich Ascites Carcinoma (EAC) Cells by CPL and Its In Vivo Anticancer Activity
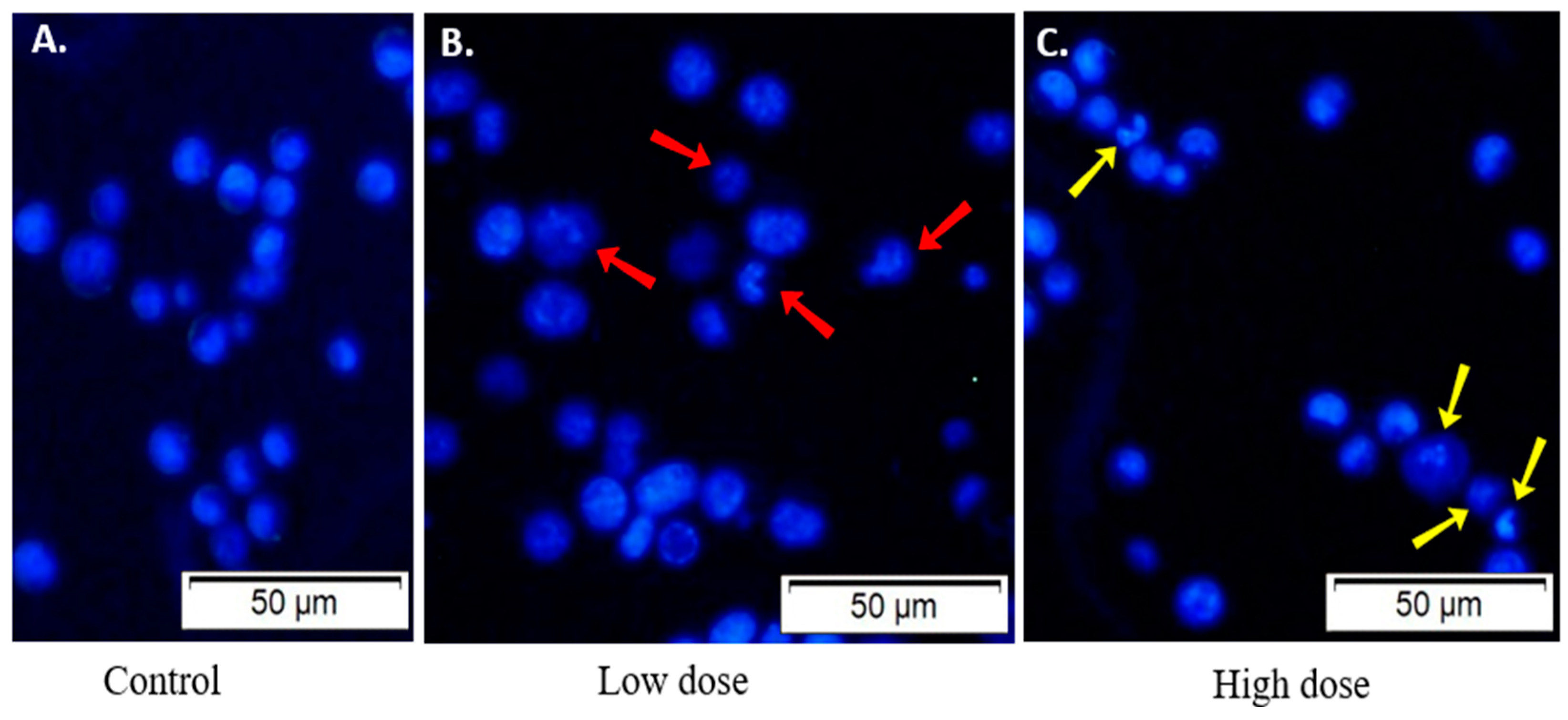
2.10. Examination of the Morphology of Lectin-Treated and Untreated EAC Cells
2.11. Effect of CPL on Blood Parameters of EAC-Bearing and Normal Mice
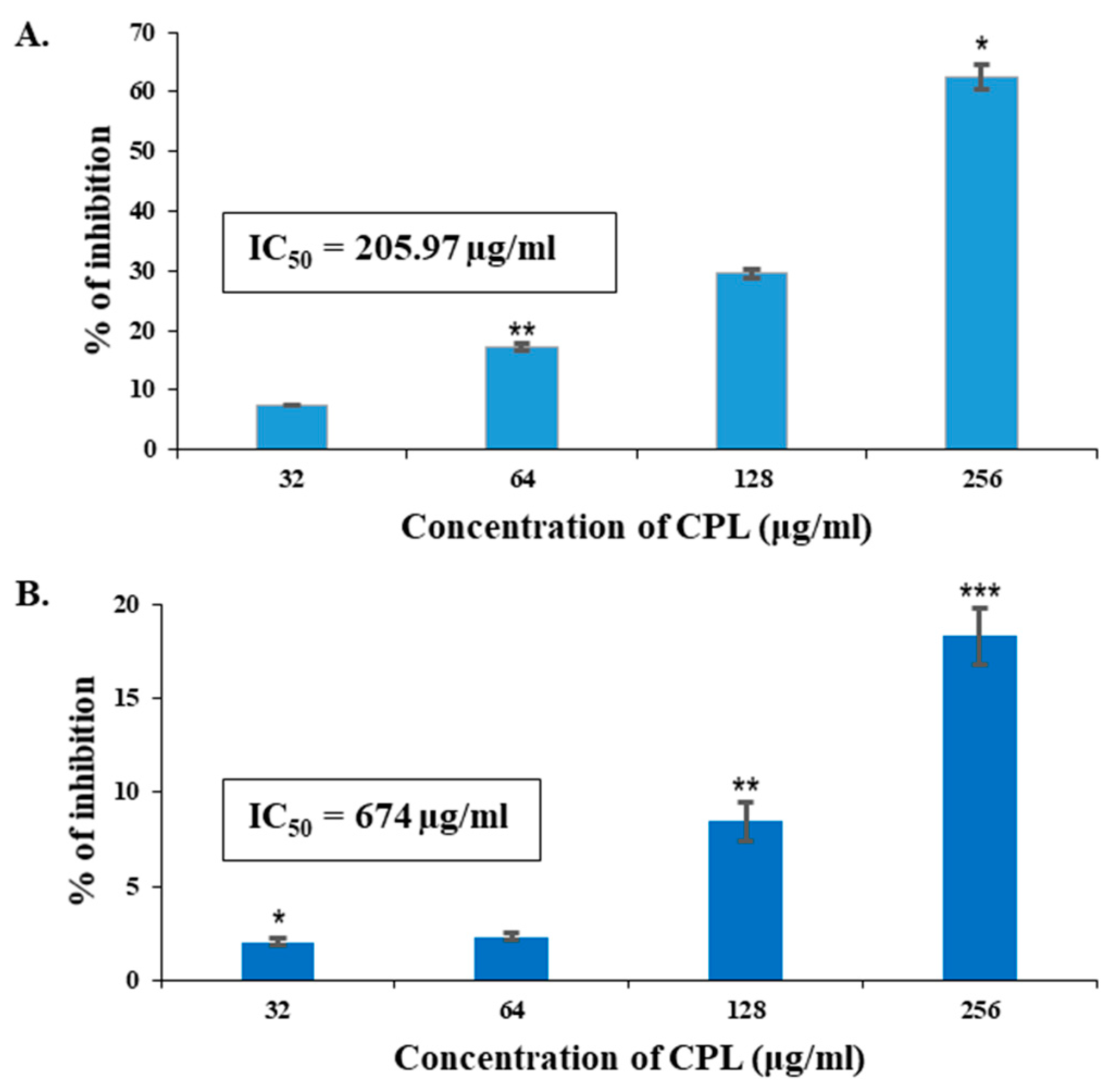
2.12. In Vitro Antiproliferative Activity of CPL against Human Cancer Cell Lines (MCF-7 and A-549) by MTT Assay
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Purification of Cucurbita Pepo Lectin (CPL)
4.3. Hemagglutination Activity of CPL
4.4. Brine Shrimp Lethality Bioassay
4.5. Effect of Temperature and pH on Hemagglutination Activity of CPL
4.6. Treatment of CPL with Urea and Guanidine HCl
4.7. Treatment of CPL with EDTA and Divalent Cations
4.8. Hemagglutination Inhibition Study of CPL by Various Sugars
4.9. Antimicrobial Activity of CPL
4.9.1. Determination of Antibacterial Activity by Disc Diffusion Assay
4.9.2. Determination of Antibiofilm Activity
4.9.3. Determination of Bacteriostatic Activity against Different Pathogenic Bacteria
4.9.4. Fungistatic Activity of CPL against Aspergillus niger
4.9.5. Agglutination of Fungal Spores by CPL
4.10. Anticancer Activity of CPL
4.10.1. Culture of Ehrlich Ascites Carcinoma Cells In Vivo in Swiss Albino Mice and Study of the Anticancer Activity of CPL
4.10.2. Determination of Hematological Parameters of Normal, EAC-Bearing and Lectin-Treated Mice
4.10.3. Observation of Morphologic Changes and Nuclear Damages of EAC Cells through Fluorescence Microscope
4.10.4. Determination of Antiproliferative Activity Assay (In Vitro) of CPL against Different Human Cancer Lines by MTT Assay
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rudiger, H.; Gabius, H.J. Plant Lectins: Occurrence, Biochemistry, Functions and Applications. Glycoconj. J. 2001, 18, 589–613. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Behura, A.; Mawatwal, S.; Kumar, A.; Naik, L.; Mohanty, S.S.; Manna, D.; Dokania, P.; Mishra, A.; Patra, S.K.; et al. Structure-Function and Application of Plant Lectins in Disease Biology and Immunity. Food Chem. Toxicol. 2019, 134, 110827. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N.; Lis, H. History of Lectins: From Hemagglutinins to Biological Recognition Molecules. Glycobiology 2004, 14, 53R–62R. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, T.; Unno, H. Functional Diversity of Novel Lectins with Unique Structural Features in Marine Animals. Cells 2023, 12, 1814. [Google Scholar] [CrossRef] [PubMed]
- Brudner, M.; Karpel, M.; Lear, C.; Chen, L.; Yantosca, L.M.; Scully, C.; Sarraju, A.; Sokolovska, A.; Zariffard, M.R.; Eisen, D.P.; et al. Lectin-Dependent Enhancement of Ebola Virus Infection via Soluble and Transmembrane C-Type Lectin Receptors. PLoS ONE 2013, 8, e60838. [Google Scholar] [CrossRef] [PubMed]
- Valbuena, G.; Madrid, J.F.; Hernandez, F.; Sáez, F.J. Identification of Fucosylated Glycoconjugates in Xenopus laevis Testis by Lectin Histochemistry. Histochem. Cell Biol. 2010, 134, 215–225. [Google Scholar] [CrossRef]
- Konozy, E.H.E.; Osman, M.E.M.; Dirar, A.I.; Ghartey-Kwansah, G. Plant Lectins: A New Antimicrobial Frontier. Biomed. Pharmacother. 2022, 155, 113735. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.F.; Da Silva, M.; Napoleao, T.; Paiva, P.; dos Correia, M.S.; Coelho, L. Lectins: Function, Structure, Biological Properties and Potential Applications. Curr. Top. Pept. Protein Res. 2014, 15, 41–62. Available online: https://hdl.handle.net/1822/43440 (accessed on 22 May 2024).
- Mazalovska, M.; Kouokam, J.C. Plant-Derived Lectins as Potential Cancer Therapeutics and Diagnostic Tools. BioMed Res. Int. 2020, 2020, 1631394. [Google Scholar] [CrossRef]
- Killiny, N. Collection of the Phloem Sap, Pros and Cons. Plant Signal. Behav. 2019, 14, 1618181. [Google Scholar] [CrossRef]
- Swamy, M.J.; Bobbili, K.B.; Mondal, S.; Narahari, A.; Datta, D. Cucurbitaceae Phloem Exudate Lectins: Purification, Molecular Characterization and Carbohydrate Binding Characteristics. Phytochemistry 2022, 201, 113251. [Google Scholar] [CrossRef]
- Ratnam, N.V.; Najibullah, M.; Ibrahim, M. A Review on Cucurbita pepo. Int. J. Pharmacogn. Phytochem. Res. 2017, 9, 1190–1194. [Google Scholar] [CrossRef]
- Ravichandran, P.; Gowri, S.; Prasath, S.S. Isolation and Partial Purification of Lectin from Seeds of Cucurbita pepo. World J. Pharm. Res. 2016, 5, 977–987. [Google Scholar] [CrossRef]
- Narahari, A.; Swamy, M.J. Rapid Affinity-Purification and Physicochemical Characterization of Pumpkin (Cucurbita maxima) Phloem Exudate Lectin. Biosci. Rep. 2010, 30, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.K. A lectin from the exudate of the fruit of the vegetable marrow (Cucurbita pepo) that has a specificity for beta-1,4-linked N-acetylglucosamine oligosaccharides. Biochem. J. 1979, 183, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Swamy, M.J.; Mondal, S. Subunit Association, and Thermal and Chemical Unfolding of Cucurbitaceae Phloem Exudate Lectins. A Review. Int. J. Biol. Macromol. 2023, 233, 123434. [Google Scholar] [CrossRef] [PubMed]
- Bobbili, K.B.; Bandari, S.; Grobe, K.; Swamy, M.J. Mutational Analysis of the Pumpkin (Cucurbita maxima) Phloem Exudate Lectin, PP2 Reveals Ser-104 Is Crucial for Carbohydrate Binding. Biochem. Biophys. Res. Commun. 2014, 450, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Bobbili, K.B.; Datta, D.; Mondal, S.; Polepalli, S.; Pohlentz, G.; Mormann, M.; Swamy, M.J. Purification, Chitooligosaccharide Binding Properties and Thermal Stability of CIA24, a New PP2-like Phloem Exudate Lectin from Ivy Gourd (Coccinia indica). Int. J. Biol. Macromol. 2018, 110, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Nareddy, P.K.; Bobbili, K.B.; Swamy, M.J. Purification, physico-chemical characterization and thermodynamics of chitooligosaccharide binding to cucumber (Cucumis sativus) phloem lectin. Int. J. Biol. Macromol. 2017, 95, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Swamy, M.J. Purification, Biochemical/Biophysical Characterization and Chitooligosaccharide Binding to BGL24, a New PP2-Type Phloem Exudate Lectin from Bottle Gourd (Lagenaria siceraria). Int. J. Biol. Macromol. 2020, 164, 3656–3666. [Google Scholar] [CrossRef]
- Sabnis, D.D.; Hart, J.W. A comparative analysis of phloem exudate proteins from Cucumis melo, Cucumis sativus and Cucurbita maxima by polyacrylamide gel electrophoresis and isoelectric focusing. Planta 1976, 130, 211–218. [Google Scholar] [CrossRef]
- Shivamadhu, M.C.; Srinivas, B.K.; Jayarama, S.; Chandrashekaraiah, S.A. Anti-cancer and anti-angiogenic effects of partially purified lectin from Praecitrullus fistulosus fruit on in vitro and in vivo model. Biomed. Pharmacother. 2017, 96, 1299–1309. [Google Scholar] [CrossRef]
- Sultan, N.A.; Swamy, M.J. Energetics of carbohydrate-binding to Momordica charantia (bitter gourd) lectin: An isothermal titration calorimetric study. Arch. Biochem Biophys. 2005, 437, 115–125. [Google Scholar] [CrossRef]
- Islam, S.S.; Karim, M.R.; Asaduzzaman, A.K.M.; Alam, A.H.M.K.; Mahmud, Z.H.; Kabir, S.R. Trichosanthes dioica seed lectin inhibits Ehrlich ascites carcinoma cells growth in vivo in mice by inducing G0/G1 cell cycle arrest. J. Food Biochem. 2021, 45, e13714. [Google Scholar] [CrossRef]
- Kabir, S.R.; Islam, F.; Alom, M.J.; Zubair, M.A.; Absar, N. Purification, characterizations of a snake guard seeds lectin with antitumor activity against Ehrlich ascites carcinoma cells in vivo in mice. Protein Peptide Lett. 2012, 19, 360–368. [Google Scholar] [CrossRef]
- Sa, R.A.; Santos, N.D.; da Silva, C.S.; Napoleao, T.H.; Gomes, F.S.; Cavada, B.S.; Coelho, L.C.; Navarro, D.M.; Bieber, L.W.; Paiva, P.M. Larvicidal activity of lectins from Myracrodruon urundeuva on Aedes aegypti. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 149, 300–306. [Google Scholar] [CrossRef]
- Silva, A.; Leite, T.; Teixeira, E.; Ponte, L.; Pinto, L. BUL: A novel lectin from Bauhinia ungulata L. seeds with fungistatic and antiproliferative activities. BMC Proc. 2014, 8, P87. [Google Scholar] [CrossRef]
- Kehr, J. Phloem sap proteins: Their identities and potential roles in the interaction between plants and phloem-feeding insects. J. Exp. Bot. 2006, 57, 767–774. [Google Scholar] [CrossRef]
- Bobbili, K.B.; Sivaji, N.; Priya, B.; Suguna, K.; Surolia, A. Structure and interactions of the phloem lectin (phloem protein 2) Cus17 from Cucumis sativus. Structure 2023, 31, 464–479.e5. [Google Scholar] [CrossRef]
- Kabir, S.R.; Nabi, M.M.; Nurujjaman, M.; Reza, M.A.; Alam, A.H.; Uz Zaman, R.; Khalid-Bin-Ferdaus, K.M.; Amin, R.; Khan, M.M.; Hossain, M.A.; et al. Momordica charantia seed lectin: Toxicity, bacterial agglutination and antitumor properties. Appl. Biochem. Biotechnol. 2015, 175, 2616–2628. [Google Scholar] [CrossRef]
- Silva, N.R.G.; de Araujo, F.N. Antibacterial Activity of Plant Lectins: A Review. Braz. Arch. Biol. Technol. 2021, 64, e21200631. [Google Scholar] [CrossRef]
- Marteyn, B.; Gazi, A.; Sansonetti, P. Shigella: A model of virulence regulation in vivo. Gut Microbes. 2012, 3, 104–120. [Google Scholar] [CrossRef]
- Kibria, M.G.; Karim, M.R.; Hasan, I.; Asaduzzaman, A.K.M.; Uddin, M.B.; Kabir, S.R. Antibacterial activity of Tricosanthes cucumerina seed lectin and study of its structural stability by fluorescence spectroscopy. Int. J. Biosci. 2018, 9, 187–192. [Google Scholar] [CrossRef]
- Mitra, A.; Guha, A.K.; Chatterjee, B.P. Typing of Shigella dysenteriae strains of different serogroups by lectins. Zentralblatt Für Bakteriol. 1994, 281, 55–60. [Google Scholar] [CrossRef]
- da Silva, I.V.; Sa, R.D.; de Moura, M.C.; Coelho, L.C.B.B.; Randau, K.P. Antimicrobial and antibiofilm activities of aqueous extracts of Cucurbita pepo L. Rev. Cienc. Med. Biol. 2020, 19, 270–275. [Google Scholar] [CrossRef]
- Yadav, S.; Tomar, A.K.; Yadav, R.N.; Yadav, S. Screening of Antifungal Proteins from Plants of Cucurbitaceae family against Fusarium oxysporum: Potential as Biofungicides. Int. Res. J. Environ. Sci. 2013, 2, 91–96. [Google Scholar]
- Choudhury, S.M.; Gupta, M.; Majumder, U.K. Antineoplastic activities of MT81 and its structural analogue in Ehrlich ascites carcinoma-bearing Swiss Albino mice. Oxid. Med. Cell Longev. 2010, 3, 61–70. [Google Scholar] [CrossRef]
- Gupta, M.; Mazumder, U.K.; Kumar, R.S.; Kumar, T.S. Antitumor activity and antioxidant role of Bauhinia racemosa against Ehrlich ascites carcinoma in Swiss albino mice [corrected]. Acta Pharmacol. Sin. 2004, 25, 1070–1076. [Google Scholar] [PubMed]
- Champattanachai, V.; Netsirisawan, P.; Chaiyawat, P.; Phueaouan, T.; Charoenwattanasatien, R.; Chokchaichamnankit, D.; Punyarit, P.; Srisomsap, C.; Svasti, J. Proteomic analysis and abrogated expression of O-GlcNAcylated proteins associated with primary breast cancer. Proteomics 2013, 13, 2088–2099. [Google Scholar] [CrossRef]
- Phueaouan, T.; Chaiyawat, P.; Netsirisawan, P.; Chokchaichamnankit, D.; Punyarit, P.; Srisomsap, C.; Svasti, J.; Champattanachai, V. Aberrant O-GlcNAc-modified proteins expressed in primary colorectal cancer. Oncol. Rep. 2013, 30, 2929–2936. [Google Scholar] [CrossRef]
- Netsirisawan, P.; Chaiyawat, P.; Chokchaichamnankit, D.; Lirdprapamongkol, K.; Srisomsap, C.; Svasti, J.; Champattanachai, V. Decreasing O-GlcNAcylation affects the malignant transformation of MCF-7 cells via Hsp27 expression and its O-GlcNAc modification. Oncol. Rep. 2018, 40, 2193–2205. [Google Scholar] [CrossRef]
- Kolar, Z.; Negrini, R.; Lisato, L. Effect of some sugars on the growth and differentiation of MCF-7 cells: I. Detection of glycosylative changes using lectin histochemistry. Acta Univ. Palacki. Olomuc. Fac. Med. 1989, 122, 113–120. [Google Scholar] [PubMed]
- Kang, J.G.; Park, S.Y.; Ji, S.; Jang, I.; Park, S.; Kim, H.S.; Kim, S.M.; Yook, J.I.; Park, Y.I.; Roth, J.; et al. O-GlcNAc protein modification in cancer cells increases in response to glucose deprivation through glycogen degradation. J. Biol. Chem. 2009, 284, 34777–34784. [Google Scholar] [CrossRef]
- Nishi, N.; Noguchi, J.; Tokura, S.; Shiota, H. Studies on chitin. 1. Acetylation of chitin. Polym. J. 1979, 11, 27–32. [Google Scholar] [CrossRef]
- Arfin, N.; Podder, M.K.; Kabir, S.R.; Asaduzzaman, A.K.M.; Hasan, I. Antibacterial, antifungal and in vivo anticancer activities of chitin-binding lectins from Tomato (Solanum lycopersicum) fruits. Arab. J. Chem. 2022, 15, 104001. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: London, UK, 1971; p. 333. [Google Scholar]
- Atkinson, H.M.; Trust, T.J. Hemagglutination properties and adherence ability of Aeromonas hydrophila. Infect. Immun. 1980, 27, 938–946. [Google Scholar] [CrossRef]
- Hasan, I.; Ozeki, Y. Histochemical localization of N-acetyl hexosamine-binding lectin HOL-18 in Halichondria okadai (Japanese black sponge), and its antimicrobial and cytotoxic anticancer effects. Int. J. Biol. Macromol. 2019, 124, 819–827. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Protein (mg) | Total Activity (HU) | Specific Activity (HU/mg) | Purification Fold | % of Yield (Per Step) |
|---|---|---|---|---|---|
| Crude extract | 1420 | 30,000 | 21.13 | 1 | 100 |
| Affinity chromatography | 38 | 7300 | 192.10 | 9.09 | 2.67 |
| Native Lectin | Demetallized Lectin | Demetallized Lectins after Reconstitution with Metal Ions | |||
|---|---|---|---|---|---|
| Ca2+ | Mg2+ | Mn2+ | Zn2+ | ||
| 256 ± 0 | 0 | 256 ± 0 | 128 ± 0 | 128 ± 0 | 128 ± 0 |
 | |||||||
|---|---|---|---|---|---|---|---|
| Sl. no | Name of Sugar | Concentration (mM) | Degree of Inhibition | Sl. no | Name of Sugar | Concentration (mM) | Degree of Inhibition |
| 1 | Glucose | 200 | − | 10 | N-acetyl-D-galactosamine | 200 | − |
| 2 | Galactose | 200 | + | 11 | N-acetyl-D-glucosamine | 200 | ++ |
| 3 | Mannose | 200 | − | 12 | Methyl-β-D-galactopyranoside | 200 | − |
| 4 | D-Melibiose | 100 | − | 13 | Methyl-α-D-galactopyranoside | 200 | − |
| 5 | Lactose | 100 | +++ | 14 | 4-Nitrophenyl-β-D-galactopyranoside | 12.5 | − |
| 6 | D (+) Xylose | 200 | − | 15 | 4-Nitrophenyl-α-D-glucopyranoside | 6.25 | ++ |
| 7 | Rhamnose | 200 | +++ | 16 | 2-Nitrophenyl-β-D-galactopyranoside | 12.5 | − |
| 8 | Fucose | 200 | + | 17 | 4-Nitrophenyl-α-D-mannopyranoside | 5 | − |
| 9 | Raffinose | 100 | + | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.A.; Hossain, M.M.; Khanam, A.; Asaduzzaman, A.K.M.; Kabir, S.R.; Ozeki, Y.; Fujii, Y.; Hasan, I. Carbohydrate-Binding Properties and Antimicrobial and Anticancer Potential of a New Lectin from the Phloem Sap of Cucurbita pepo. Molecules 2024, 29, 2531. https://doi.org/10.3390/molecules29112531
Islam MA, Hossain MM, Khanam A, Asaduzzaman AKM, Kabir SR, Ozeki Y, Fujii Y, Hasan I. Carbohydrate-Binding Properties and Antimicrobial and Anticancer Potential of a New Lectin from the Phloem Sap of Cucurbita pepo. Molecules. 2024; 29(11):2531. https://doi.org/10.3390/molecules29112531
Chicago/Turabian StyleIslam, Md. Aminul, Md. Mikail Hossain, Alima Khanam, A. K. M. Asaduzzaman, Syed Rashel Kabir, Yasuhiro Ozeki, Yuki Fujii, and Imtiaj Hasan. 2024. "Carbohydrate-Binding Properties and Antimicrobial and Anticancer Potential of a New Lectin from the Phloem Sap of Cucurbita pepo" Molecules 29, no. 11: 2531. https://doi.org/10.3390/molecules29112531
APA StyleIslam, M. A., Hossain, M. M., Khanam, A., Asaduzzaman, A. K. M., Kabir, S. R., Ozeki, Y., Fujii, Y., & Hasan, I. (2024). Carbohydrate-Binding Properties and Antimicrobial and Anticancer Potential of a New Lectin from the Phloem Sap of Cucurbita pepo. Molecules, 29(11), 2531. https://doi.org/10.3390/molecules29112531