


Effect of N-acetyl-l-cysteine on Cell Phenotype and Autophagy in Pichia pastoris Expressing Human Serum Albumin and Porcine Follicle-Stimulating Hormone Fusion Protein


Abstract
1. Introduction
2. Results
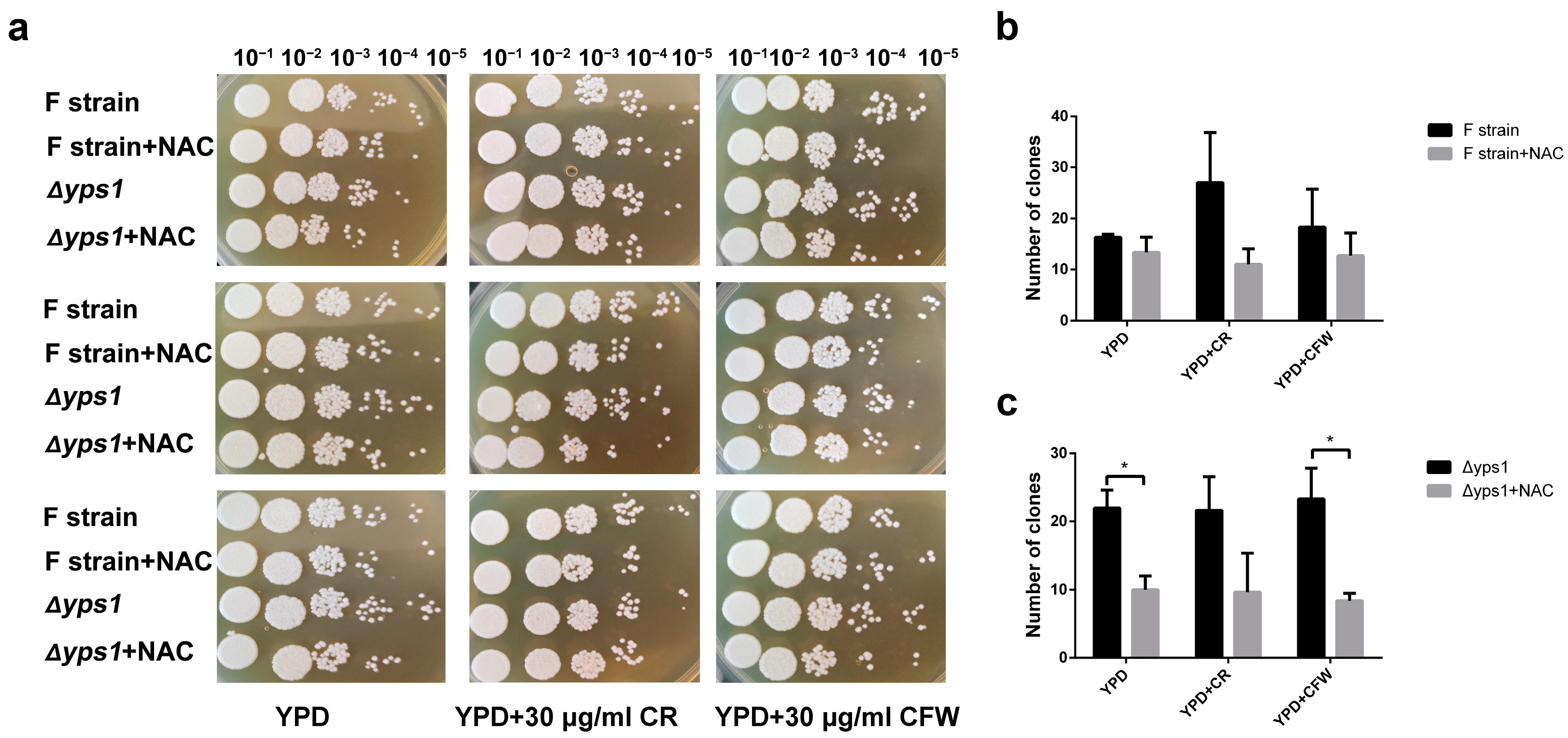
2.1. Effect of NAC on Cell Growth and Phenotype
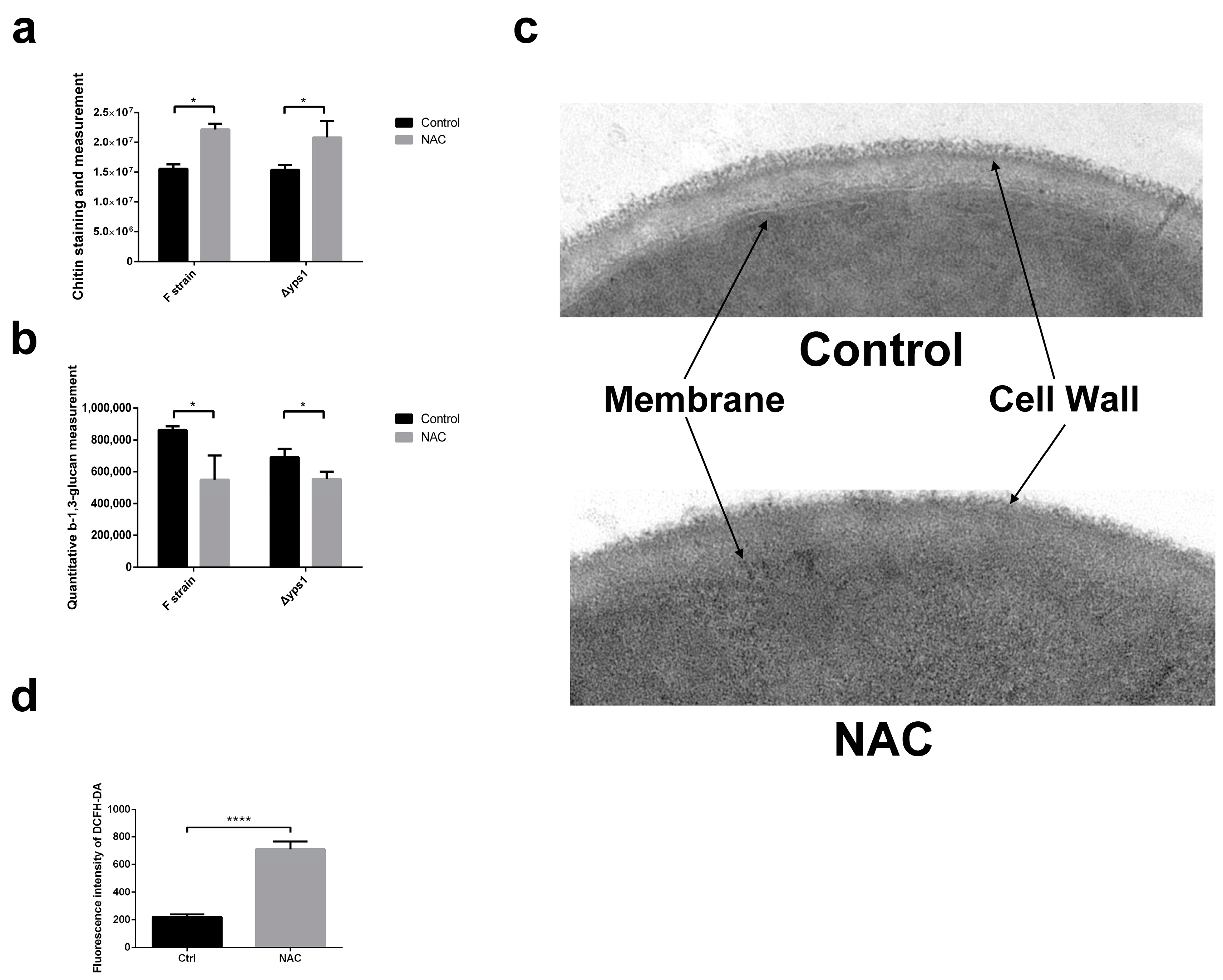
2.2. NAC Affects the Composition of the Cell Wall
2.3. Transcript Expression Analysis of Genes Involved in Vesicular Trafficking
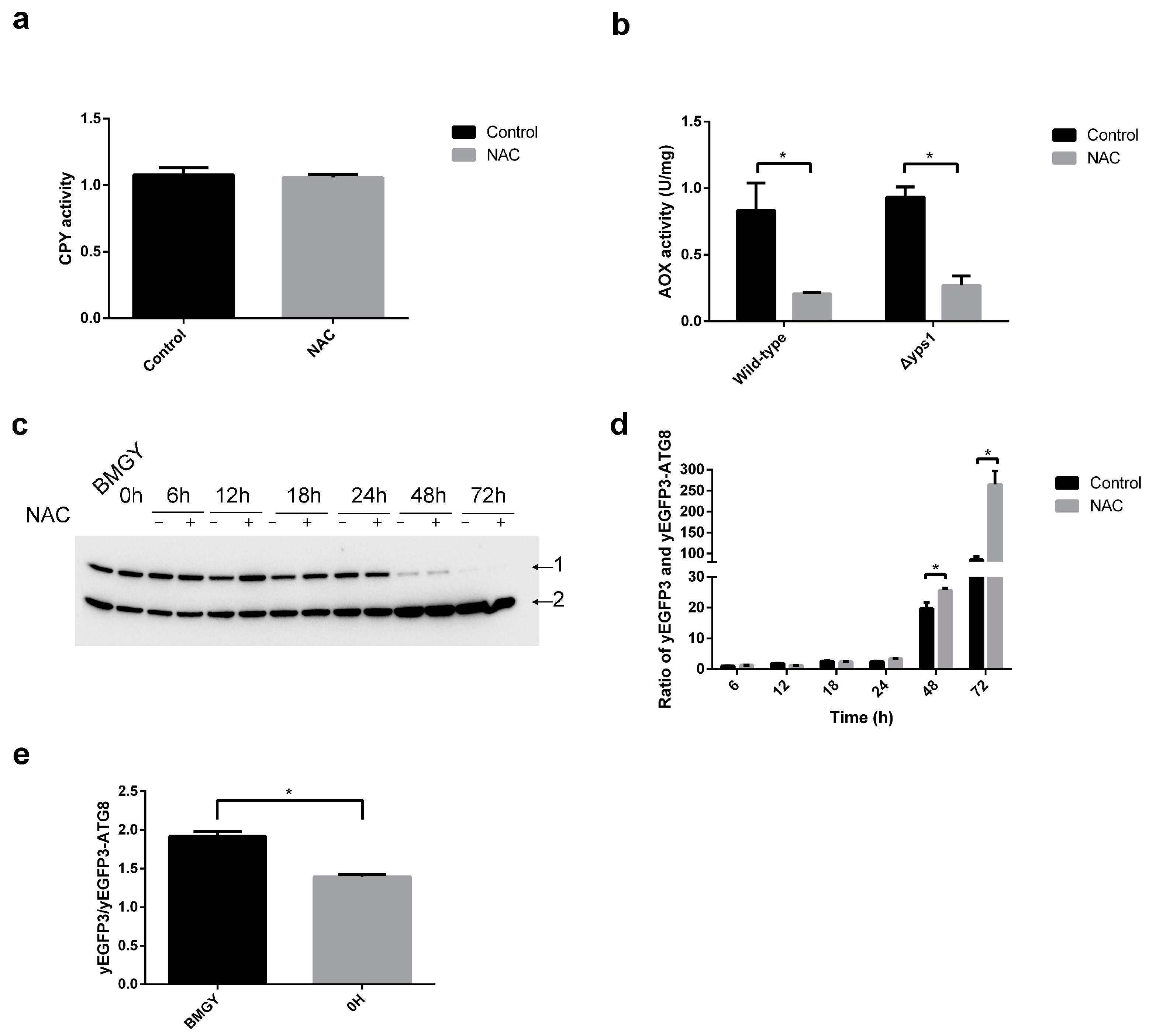
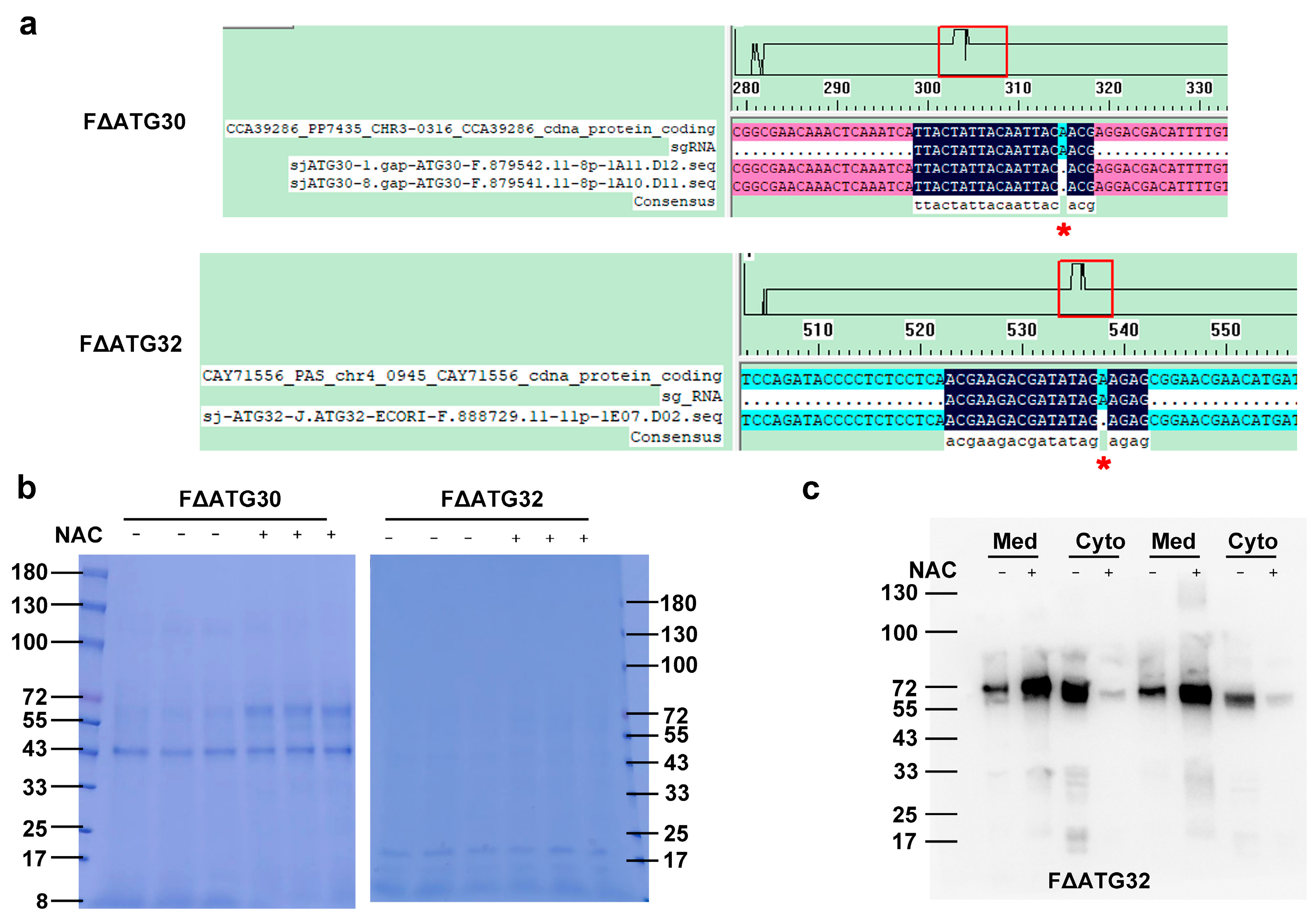
2.4. Autophagy Is Involved in NAC-Promoted HSA-pFSHβ Secretion
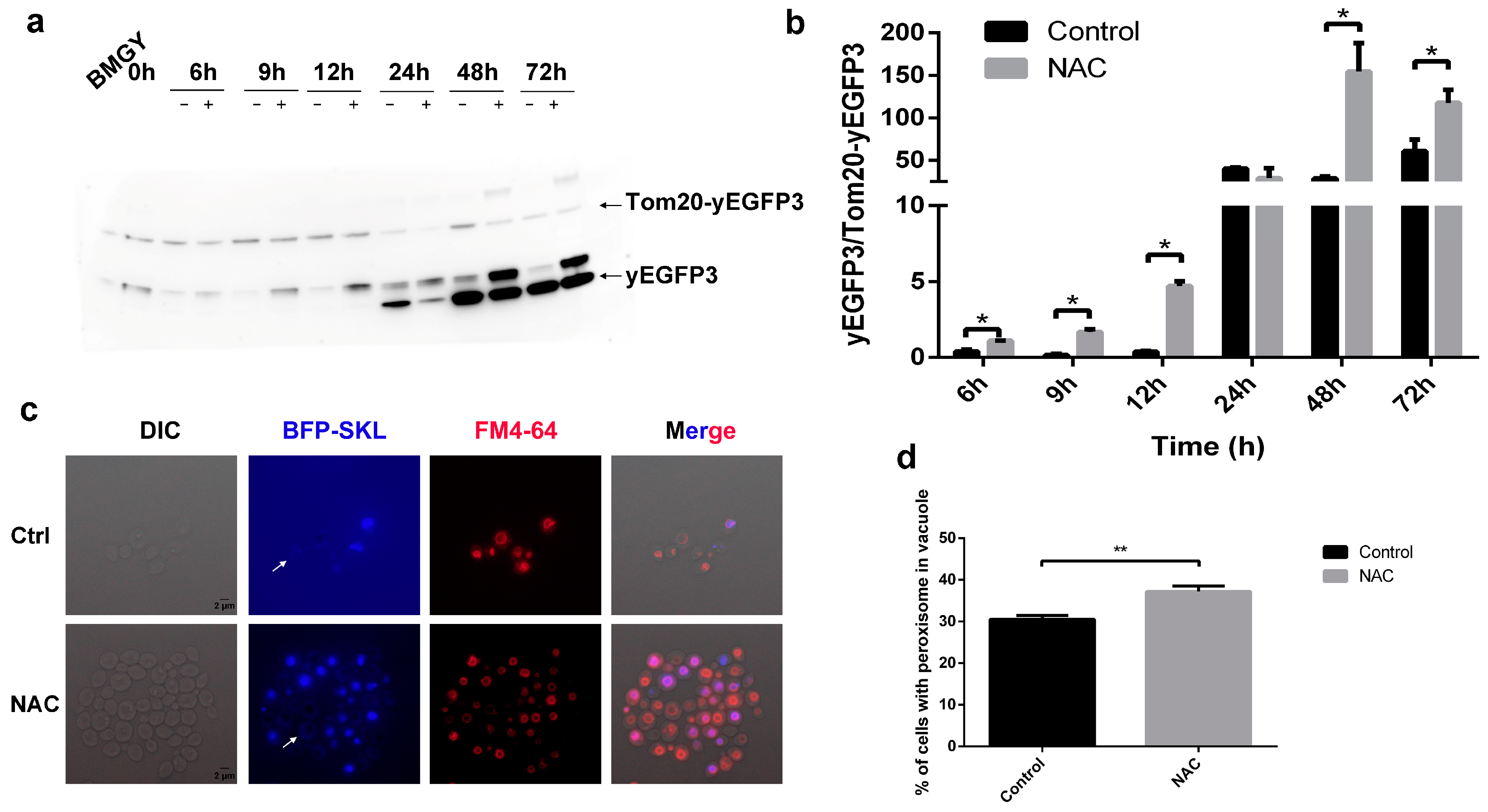
2.5. The Types of Autophagy Affected by NAC
2.6. Role of Autophagy in the Promotion of HSA-pFSHβ Secretion by NAC
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strains, Plasmids, and Reagents
5.2. Construction of Expression Plasmids of pGAPZA-yEGFP3-ATG8, pGAPZA-BFP-SKL, pACTZ-Ape1 and pTom20-yEGFP3
5.3. Transformation and Screening Recombinant Strains
5.4. Construction of ΔAtg30 and ΔAtg32 Disrupted Strains
5.5. Proteins Expression and Analysis
5.6. Phenotypic Analysis
5.7. Quantitative Chitin and β-1,3-glucan Measurement
5.8. Quantitative Real-Time PCR
5.9. Determination of Alcohol Oxidase (AOX)
5.10. Intracellular ROS Measurement
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Matsuzaki, Y.; Kajiwara, K.; Aoki, W.; Ueda, M. Production of Single-Domain Antibodies in Pichia pastoris. Methods Mol. Biol. 2022, 2446, 181–203. [Google Scholar] [CrossRef] [PubMed]
- Montoliu-Gaya, L.; Villegas, S. Production of Therapeutic Single-Chain Variable Fragments (ScFv) in Pichia pastoris. Methods Mol. Biol. 2022, 2313, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Gassler, T.; Sauer, M.; Gasser, B.; Egermeier, M.; Troyer, C.; Causon, T.; Hann, S.; Mattanovich, D.; Steiger, M.G. The industrial yeast Pichia pastoris is converted from a heterotroph into an autotroph capable of growth on CO2. Nat. Biotechnol. 2020, 38, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Shay, L.; Hunt, H.; Wegner, G. High-productivity fermentation process for cultivating industrial microorganisms. J. Ind. Microbiol. 1987, 2, 79–85. [Google Scholar] [CrossRef]
- Karbalaei, M.; Rezaee, S.A.; Farsiani, H. Pichia pastoris: A highly successful expression system for optimal synthesis of heterologous proteins. J. Cell. Physiol. 2020, 235, 5867–5881. [Google Scholar] [CrossRef]
- Yu, P.; Zhu, Q.; Chen, K.; Lv, X. Improving the secretory production of the heterologous protein in Pichia pastoris by focusing on protein folding. Appl. Biochem. Biotechnol. 2015, 175, 535–548. [Google Scholar] [CrossRef]
- Idiris, A.; Tohda, H.; Kumagai, H.; Takegawa, K. Engineering of protein secretion in yeast: Strategies and impact on protein production. Appl. Microbiol. Biotechnol. 2010, 86, 403–417. [Google Scholar] [CrossRef]
- Cregg, J.M.; Madden, K.R.; Barringer, K.J.; Thill, G.P.; Stillman, C.A. Functional characterization of the two alcohol oxidase genes from the yeast Pichia pastoris. Mol. Cell. Biol. 1989, 9, 1316–1323. [Google Scholar] [CrossRef]
- Mattanovich, D.; Graf, A.; Stadlmann, J.; Dragosits, M.; Redl, A.; Maurer, M.; Kleinheinz, M.; Sauer, M.; Altmann, F.; Gasser, B. Genome, secretome and glucose transport highlight unique features of the protein production host Pichia pastoris. Microb. Cell Fact. 2009, 8, 29. [Google Scholar] [CrossRef]
- Puxbaum, V.; Mattanovich, D.; Gasser, B. Quo vadis? The challenges of recombinant protein folding and secretion in Pichia pastoris. Appl. Microbiol. Biotechnol. 2015, 99, 2925–2938. [Google Scholar] [CrossRef]
- Delic, M.; Valli, M.; Graf, A.B.; Pfeffer, M.; Mattanovich, D.; Gasser, B. The secretory pathway: Exploring yeast diversity. FEMS Microbiol. Rev. 2013, 37, 872–914. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, R.; Eyrisch, S.; Ahmad, M.; Helms, V. Protein translocation across the ER membrane. Biochim. Biophys. Acta 2011, 1808, 912–924. [Google Scholar] [CrossRef] [PubMed]
- Thibault, G.; Ng, D.T. The endoplasmic reticulum-associated degradation pathways of budding yeast. Cold Spring Harb. Perspect Biol. 2012, 4, a013193. [Google Scholar] [CrossRef] [PubMed]
- Hampton, R.Y. ER-associated degradation in protein quality control and cellular regulation. Curr. Opin. Cell Biol. 2002, 14, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Navone, L.; Vogl, T.; Luangthongkam, P.; Blinco, J.A.; Luna-Flores, C.H.; Chen, X.; von Hellens, J.; Mahler, S.; Speight, R. Disulfide bond engineering of AppA phytase for increased thermostability requires co-expression of protein disulfide isomerase in Pichia pastoris. Biotechnol. Biofuels 2021, 14, 80. [Google Scholar] [CrossRef] [PubMed]
- Delic, M.; Göngrich, R.; Mattanovich, D. Engineering of protein folding and secretion-strategies to overcome bottlenecks for efficient production of recombinant proteins. Antioxid. Redox Sign. 2014, 21, 414–437. [Google Scholar] [CrossRef]
- Marsalek, L.; Puxbaum, V.; Buchetics, M.; Mattanovich, D.; Gasser, B. Disruption of vacuolar protein sorting components of the HOPS complex leads to enhanced secretion of recombinant proteins in Pichia pastoris. Microb. Cell Fact. 2019, 18, 119. [Google Scholar] [CrossRef]
- Marsalek, L.; Gruber, C.; Altmann, F.; Aleschko, M.; Mattanovich, D.; Gasser, B.; Puxbaum, V. Disruption of genes involved in CORVET complex leads to enhanced secretion of heterologous carboxylesterase only in protease deficient Pichia pastoris. Biotechnol. J. 2017, 12, 1600584. [Google Scholar] [CrossRef]
- Besada-Lombana, P.B.; Da, S.N. Engineering the early secretory pathway for increased protein secretion in Saccharomyces cerevisiae. Metab. Eng. 2019, 55, 142–151. [Google Scholar] [CrossRef]
- Luo, G.; Liu, J.; Bian, T.; Zhang, Z.; Li, M. The mechanism of N-acetyl-l-cysteine in improving the secretion of porcine follicle-stimulating hormone in Pichia pastoris. Yeast 2021, 38, 601–611. [Google Scholar] [CrossRef]
- Deffieu, M.; Bhatia-Kissova, I.; Salin, B.; Galinier, A.; Manon, S.; Camougrand, N. Glutathione participates in the regulation of mitophagy in yeast. J. Biol. Chem. 2009, 284, 14828–14837. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Geng, Z.; Kuerban, B.; Yang, J.; Liu, J.; Li, M. Enhancement of HSA-pFSHβ Production by Disrupting YPS1 and Supplementing N-acetyl-Lcysteine in Pichia Pastoris. Front. Microbiol. 2022, 13, 998647. [Google Scholar] [CrossRef] [PubMed]
- Sazonova, E.A.; Zobnina, A.E.; Padkina, M.V. Effect of disruption of Pichia pastoris YPS1 gene on viability and production of recombinant proteins. Genetika 2013, 49, 696–702. [Google Scholar] [CrossRef]
- Aguilar-Uscanga, B.; Francois, J.M. A study of the yeast cell wall composition and structure in response to growth conditions and mode of cultivation. Lett. Appl. Microbiol. 2003, 37, 268–274. [Google Scholar] [CrossRef]
- Fuchs, B.B.; Mylonakis, E. Our paths might cross: The role of the fungal cell wall integrity pathway in stress response and cross talk with other stress response pathways. Eukaryot Cell 2009, 8, 1616–1625. [Google Scholar] [CrossRef]
- Luo, G. Expression of Porcine Follicle Stimulating Hormone in Pichia pastoris. Ph.D. Thesis, China Agricultural University, Beijing, China, 2019. [Google Scholar]
- Farre, J.C.; Manjithaya, R.; Mathewson, R.D.; Subramani, S. PpAtg30 tags peroxisomes for turnover by selective autophagy. Dev. Cell 2008, 14, 365–376. [Google Scholar] [CrossRef]
- Kanki, T.; Wang, K.; Cao, Y.; Baba, M.; Klionsky, D.J. Atg32 is a mitochondrial protein that confers selectivity during mitophagy. Dev. Cell 2009, 17, 98–109. [Google Scholar] [CrossRef]
- Yorimitsu, T.; Sato, K.; Takeuchi, M. Molecular mechanisms of Sar/Arf GTPases in vesicular trafficking in yeast and plants. Front. Plant Sci. 2014, 5, 411. [Google Scholar] [CrossRef] [PubMed]
- Makaraci, P.; Delgado, C.M.; McDermott, H.; Nguyen, V.; Highfill, C.; Kim, K. Yeast dynamin and Ypt6 function in parallel for the endosome-to-Golgi retrieval of Snc1. Cell Biol. Int. 2019, 43, 1137–1151. [Google Scholar] [CrossRef]
- Brocker, C.; Engelbrecht-Vandre, S.; Ungermann, C. Multisubunit tethering complexes and their role in membrane fusion. Curr. Biol. 2010, 20, R943–R952. [Google Scholar] [CrossRef]
- Zahrl, R.J.; Gasser, B.; Mattanovich, D.; Ferrer, P. Detection and elimination of cellular bottlenecks in protein-producing yeasts. Methods Mol. Biol. 2019, 1923, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Zahrl, R.J.; Prielhofer, R.; Burgard, J.; Mattanovich, D.; Gasser, B. Synthetic activation of yeast stress response improves secretion of recombinant proteins. New Biotechnol. 2023, 73, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Tian, J.; Huang, H.; Lei, A. Improving heterologous expression of porcine follicle-stimulating hormone in Pichia pastoris by integrating molecular strategies and culture condition optimization. Appl. Microbiol. Biotechnol. 2018, 102, 8867–8882. [Google Scholar] [CrossRef] [PubMed]
- Looser, V.; Bruhlmann, B.; Bumbak, F.; Stenger, C.; Costa, M.; Camattari, A.; Fotiadis, D.; Kovar, K. Cultivation strategies to enhance productivity of Pichia pastoris: A review. Biotechnol. Adv. 2015, 33, 1177–1193. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.; Brito, N.; Hernandez-Bolanos, E.; Gonzalez, C. New tools for high-throughput expression of fungal secretory proteins in Saccharomyces cerevisiae and Pichia pastoris. Microb. Biotechnol. 2019, 12, 1139–1153. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, C.A.; Jivan, A.D.; Vo, M.N.; de Sa, C.K.; Deyarmin, J.S.; Hekman, R.M.; Uribe, C.; Hang, A.; Her, K.; Fong, M.M.; et al. Role of BGS13 in the Secretory Mechanism of Pichia pastoris. Appl. Environ. Microbiol. 2019, 85, e01615-19. [Google Scholar] [CrossRef]
- Ast, T.; Cohen, G.; Schuldiner, M. A network of cytosolic factors targets SRP-independent proteins to the endoplasmic reticulum. Cell 2013, 152, 1134–1145. [Google Scholar] [CrossRef]
- Takeshige, K.; Baba, M.; Tsuboi, S.; Noda, T.; Ohsumi, Y. Autophagy in yeast demonstrated with proteinase-deficient mutants and conditions for its induction. J. Cell Biol. 1992, 119, 301–311. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, F.; Zou, S.; Yu, S.; Li, S.; Li, D.; Song, J.; Li, H.; He, Z.; Hu, B.; et al. A Vps21 endocytic module regulates autophagy. Mol. Biol. Cell 2014, 25, 3166–3177. [Google Scholar] [CrossRef]
- Reggiori, F.; Shintani, T.; Nair, U.; Klionsky, D.J. Atg9 cycles between mitochondria and the pre-autophagosomal structure in yeasts. Autophagy 2005, 1, 101–109. [Google Scholar] [CrossRef]
- Okamoto, K.; Kondo-Okamoto, N.; Ohsumi, Y. Mitochondria-anchored receptor Atg32 mediates degradation of mitochondria via selective autophagy. Dev. Cell 2009, 17, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Anding, A.L.; Baehrecke, E.H. Cleaning House: Selective Autophagy of Organelles. Dev. Cell 2017, 41, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.N.; Chowdhury, R.; Trudel, L.J.; Tee, A.R.; Slack, R.S.; Walker, C.L.; Wogan, G.N. Reactive nitrogen species regulate autophagy through ATM-AMPK-TSC2-mediated suppression of mTORC1. Proc. Natl. Acad. Sci. USA 2013, 110, E2950–E2957. [Google Scholar] [CrossRef] [PubMed]
- Das, E.; Sahu, K.K.; Roy, I. The functional role of Ire1 in regulating autophagy and proteasomal degradation under prolonged proteotoxic stress. FEBS J. 2023; Early View. [Google Scholar] [CrossRef]
- Cheong, H.; Klionsky, D.J. Biochemical methods to monitor autophagy-related processes in yeast. Method Enzym. 2008, 451, 1–26. [Google Scholar] [CrossRef]
- Yamashita, S.I.; Yurimoto, H.; Murakami, D.; Yoshikawa, M.; Oku, M.; Sakai, Y. Lag-phase autophagy in the methylotrophic yeast Pichia pastoris. Genes Cells 2009, 14, 861–870. [Google Scholar] [CrossRef]
- Titorenko, V.I.; Terlecky, S.R. Peroxisome metabolism and cellular aging. Traffic 2011, 12, 252–259. [Google Scholar] [CrossRef]
- Wang, J.J.; Xu, W.N.; Li, X.; Li, J.; Li, Q. Absence of fks1p in lager brewing yeast results in aberrant cell wall composition and improved beer flavor stability. World J. Microbiol. Biotechnol. 2014, 30, 1901–1908. [Google Scholar] [CrossRef]
- Yin, Z.; Feng, W.; Chen, C.; Xu, J.; Li, Y.; Yang, L.; Wang, J.; Liu, X.; Wang, W.; Gao, C.; et al. Shedding light on autophagy coordinating with cell wall integrity signaling to govern pathogenicity of Magnaporthe oryzae. Autophagy 2020, 16, 900–916. [Google Scholar] [CrossRef]
- Cormack, B.P.; Bertram, G.; Egerton, M.; Gow, N.; Falkow, S.; Brown, A. Yeast-enhanced green fluorescent protein (yEGFP): A reporter of gene expression in Candida albicans. Microbiology 1997, 143, 303–311. [Google Scholar] [CrossRef]
- Gassler, T.; Heistinger, L.; Mattanovich, D.; Gasser, B.; Prielhofer, R. CRISPR/Cas9-Mediated Homology-Directed Genome Editing in Pichia pastoris. Methods Mol. Biol. 2019, 1923, 211–225. [Google Scholar] [CrossRef]
- Schroder, L.A.; Glick, B.S.; Dunn, W.A. Identification of pexophagy genes by restriction enzyme-mediated integration. Pichia Protoc. 2007, 389, 203–218. [Google Scholar] [CrossRef]
- Tam, Y.J.; Allaudin, Z.N.; Lila, M.A.; Bahaman, A.R.; Tan, J.S.; Rezaei, M.A. Enhanced cell disruption strategy in the release of recombinant hepatitis B surface antigen from Pichia pastoris using response surface methodology. BMC Biotechnol. 2012, 12, 70. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains Name | Description | Genotype | Source |
|---|---|---|---|
| F strain | WT | GS115 HSA-pFSHβ::prAOX(Zeocinr) his4 | |
| FATG8 | WT/prGAP-yEGFP3-ATG8 | F yEGFP3-ATG8::GAP his4 | This study |
| FApe1 | WT/prACT-Ape1-yEGFP3 | F Ape1-yEGFP3::ACT his4 | This study |
| FTom20 | WT/prTOM20-Tom20-yEGFP3 | F Tom20-yEGFP3::TOM20 his4 | This study |
| FBFP-SKL | WT/prGAP-BFP-SKL | F BFP-SKL::GAP his4 | This study |
| FΔATG30 | atg30Δ | F atg30Δ::Hygromycin Br his4 | This study |
| FΔATG32 | atg32Δ | F atg32Δ::Hygromycin Br his4 | This study |
| FTom20-ΔATG32 | FTom20/ΔATG32 | FTom20 atg32Δ::Hygromycin Br his4 | This study |
| FApe1-ΔATG30 | FApe1/ΔATG30 | FApe1 atg30Δ::Hygromycin Br his4 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Geng, Z.; Yang, C.; Zhou, H.; Wang, Y.; Kuerban, B.; Luo, G. Effect of N-acetyl-l-cysteine on Cell Phenotype and Autophagy in Pichia pastoris Expressing Human Serum Albumin and Porcine Follicle-Stimulating Hormone Fusion Protein. Molecules 2023, 28, 3041. https://doi.org/10.3390/molecules28073041
Xu Y, Geng Z, Yang C, Zhou H, Wang Y, Kuerban B, Luo G. Effect of N-acetyl-l-cysteine on Cell Phenotype and Autophagy in Pichia pastoris Expressing Human Serum Albumin and Porcine Follicle-Stimulating Hormone Fusion Protein. Molecules. 2023; 28(7):3041. https://doi.org/10.3390/molecules28073041
Chicago/Turabian StyleXu, Yingqing, Zijian Geng, Chengxi Yang, Hongwei Zhou, Yixing Wang, Buayisham Kuerban, and Gang Luo. 2023. "Effect of N-acetyl-l-cysteine on Cell Phenotype and Autophagy in Pichia pastoris Expressing Human Serum Albumin and Porcine Follicle-Stimulating Hormone Fusion Protein" Molecules 28, no. 7: 3041. https://doi.org/10.3390/molecules28073041
APA StyleXu, Y., Geng, Z., Yang, C., Zhou, H., Wang, Y., Kuerban, B., & Luo, G. (2023). Effect of N-acetyl-l-cysteine on Cell Phenotype and Autophagy in Pichia pastoris Expressing Human Serum Albumin and Porcine Follicle-Stimulating Hormone Fusion Protein. Molecules, 28(7), 3041. https://doi.org/10.3390/molecules28073041