
Finite Element Modelling of a Gram-Negative Bacterial Cell and Nanospike Array for Cell Rupture Mechanism Study



Abstract
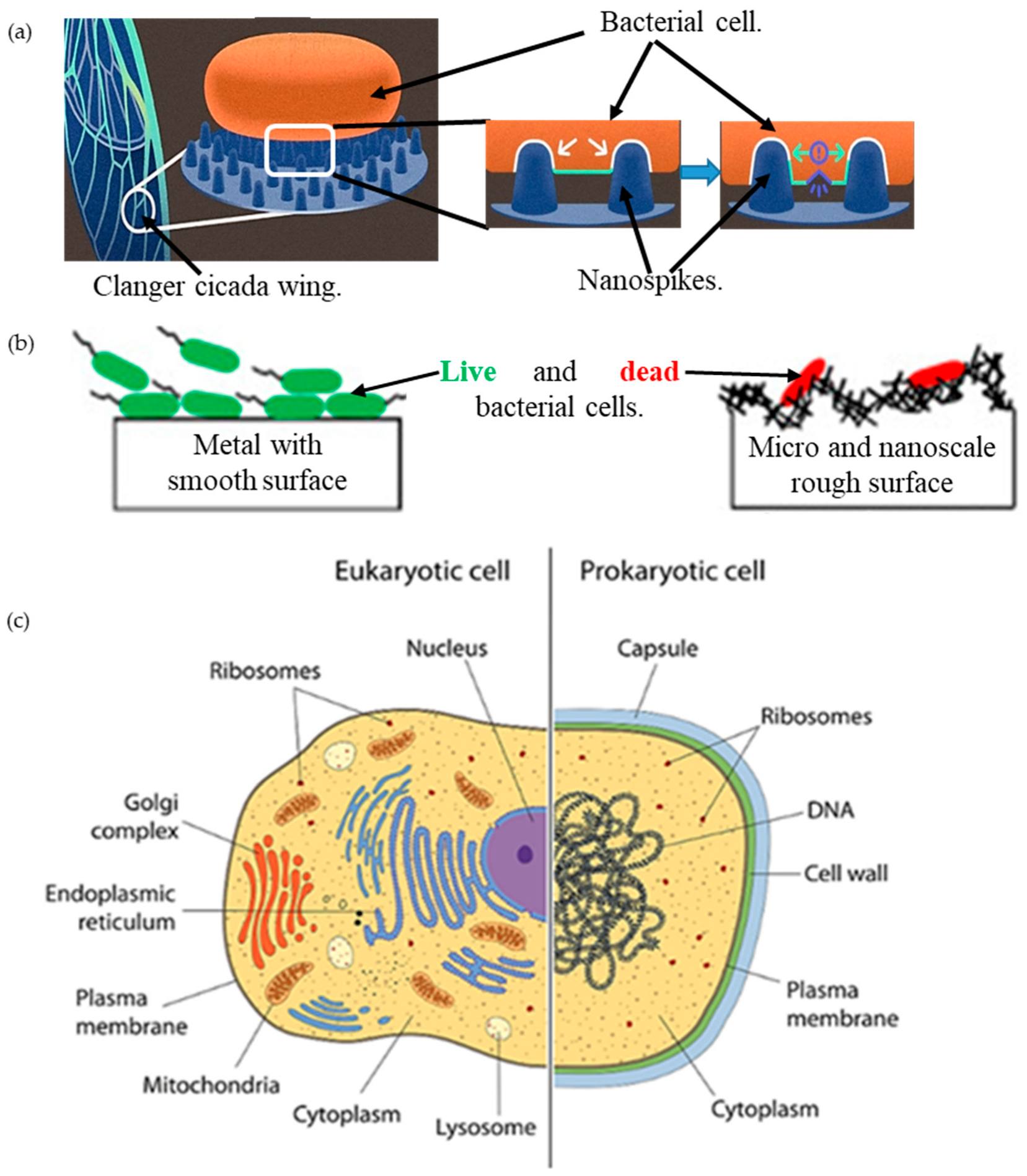
1. Introduction

2. Results and Discussions
3. Computational Modeling
3.1. Modeling the Cell and the Nanospike Geometry
3.2. Setup the Simulation
3.3. Assigning the Boundary Conditions
3.4. Grid Generations and Grid Independence Test
3.5. Validation of the Simulation Model with Literature
4. Conclusions
- The bacterial cell wall is deformed on the location of nanospike tips as full contact is generated under the applied turgor pressure.
- The location of maximum stress and strain values on the cell wall was observed on the three-phase contact point on which a common region between the cell wall, nanospike, and liquid of the bacterial cell exists.
- The generated stress on the cell wall due to the applied load is less than the cell tensile strength, it continues to cause a creep deformation, and gradually a rupture can be expected in the cell at around the contact point. The rupture is similar to that of a paper punching mechanism.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Dunne, W.M., Jr. Bacterial adhesion: Seen any good biofilms lately? Clin. Microbiol. Rev. 2002, 15, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.; Monteiro, F.J.; Ferraz, M.P. Infection of orthopedic implants with emphasis on bacterial adhesion process and techniques used in studying bacterial-material interactions. Biomatter 2012, 2, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Ishantha Senevirathne, S.; Hasan, J.; Mathew, A.; Jaggessar, A.; Yarlagadda, P.K. Trends in Bactericidal Nanostructured Surfaces: An Analytical Perspective. ACS Appl. Bio Mater. 2021, 4, 7626–7642. [Google Scholar] [CrossRef] [PubMed]
- Hasan, J.; Xu, Y.; Yarlagadda, T.; Schuetz, M.; Spann, K.; Yarlagadda, P.K. Antiviral and antibacterial nanostructured surfaces with excellent mechanical properties for hospital applications. ACS Biomater. Sci. Eng. 2020, 6, 3608–3618. [Google Scholar] [CrossRef] [PubMed]
- Senevirathne, S.; Hasan, J.; Mathew, A.; Woodruff, M.; Yarlagadda, P. Bactericidal efficiency of micro-and nanostructured surfaces: A critical perspective. RSC Adv. 2021, 11, 1883–1900. [Google Scholar] [CrossRef] [PubMed]
- Hasan, J.; Crawford, R.J.; Ivanova, E.P. Antibacterial surfaces: The quest for a new generation of biomaterials. Trends Biotechnol. 2013, 31, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Hasan, J.; Webb, H.K.; Truong, V.K.; Pogodin, S.; Baulin, V.A.; Watson, G.S.; Watson, J.A.; Crawford, R.J.; Ivanova, E.P. Selective bactericidal activity of nanopatterned superhydrophobic cicada Psaltoda claripennis wing surfaces. Appl. Microbiol. Biotechnol. 2013, 97, 9257–9262. [Google Scholar] [CrossRef]
- Hazell, G.; Fisher, L.E.; Murray, W.A.; Nobbs, A.H.; Su, B. Bioinspired bactericidal surfaces with polymer nanocone arrays. J. Colloid Interface Sci. 2018, 528, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, X.; Wang, Z.L. Controlled replication of butterfly wings for achieving tunable photonic properties. Nano Lett. 2006, 6, 2325–2331. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Hasan, J.; Webb, H.K.; Truong, V.K.; Watson, G.S.; Watson, J.A.; Baulin, V.A.; Pogodin, S.; Wang, J.Y.; Tobin, M.J. Natural bactericidal surfaces: Mechanical rupture of Pseudomonas aeruginosa cells by cicada wings. Small 2012, 8, 2489–2494. [Google Scholar] [CrossRef]
- Shahali, H.; Hasan, J.; Mathews, A.; Wang, H.; Yan, C.; Tesfamichael, T.; Yarlagadda, P.K. Multi-biofunctional properties of three species of cicada wings and biomimetic fabrication of nanopatterned titanium pillars. J. Mater. Chem. B 2019, 7, 1300–1310. [Google Scholar] [CrossRef]
- Jaggessar, A.; Yarlagadda, P.; Qiu, T.; Li, T.; Tesfamichael, T. Fabrication of nano pyramid texture on Ti-6Al-4V using nanosphere lithography. Mater. Today Proc. 2018, 5, 11593–11600. [Google Scholar] [CrossRef]
- Jaggessar, A.; Mathew, A.; Tesfamichael, T.; Wang, H.; Yan, C.; Yarlagadda, P.K. Bacteria death and osteoblast metabolic activity correlated to hydrothermally synthesised TiO2 surface properties. Molecules 2019, 24, 1201. [Google Scholar] [CrossRef] [PubMed]
- Hasan, J.; Chatterjee, K. Recent advances in engineering topography mediated antibacterial surfaces. Nanoscale 2015, 7, 15568–15575. [Google Scholar] [CrossRef] [PubMed]
- Hasan, J.; Jain, S.; Chatterjee, K. Nanoscale topography on black titanium imparts multi-biofunctional properties for orthopedic applications. Sci. Rep. 2017, 7, 41118. [Google Scholar] [CrossRef] [PubMed]
- Vidyasagar, A.; Pappas, S. What Are Bacteria? 15 October 2021. Available online: https://www.livescience.com/51641-bacteria.html#section-structure-of-bacteria (accessed on 14 October 2022).
- Arnoldi, M.; Fritz, M.; Bäuerlein, E.; Radmacher, M.; Sackmann, E.; Boulbitch, A. Bacterial turgor pressure can be measured by atomic force microscopy. Phys. Rev. E 2000, 62, 1034. [Google Scholar] [CrossRef]
- Pogodin, S.; Hasan, J.; Baulin, V.A.; Webb, H.K.; Truong, V.K.; Nguyen, T.H.P.; Boshkovikj, V.; Fluke, C.J.; Watson, G.S.; Watson, J.A. Biophysical model of bacterial cell interactions with nanopatterned cicada wing surfaces. Biophys. J. 2013, 104, 835–840. [Google Scholar] [CrossRef]
- Velic, A.; Jaggessar, A.; Tesfamichael, T.; Li, Z.; Yarlagadda, P.K. Effects of nanopillar size and spacing on mechanical perturbation and bactericidal killing efficiency. Nanomaterials 2021, 11, 2472. [Google Scholar] [CrossRef]
- Linklater, D.P.; Baulin, V.A.; Juodkazis, S.; Crawford, R.J.; Stoodley, P.; Ivanova, E.P. Mechano-bactericidal actions of nanostructured surfaces. Nat. Rev. Microbiol. 2021, 19, 8–22. [Google Scholar] [CrossRef]
- Modaresifar, K.; Azizian, S.; Ganjian, M.; Fratila-Apachitei, L.E.; Zadpoor, A.A. Bactericidal effects of nanopatterns: A systematic review. Acta Biomater. 2019, 83, 29–36. [Google Scholar] [CrossRef]
- Bhadra, C.M.; Khanh Truong, V.; Pham, V.T.; Al Kobaisi, M.; Seniutinas, G.; Wang, J.Y.; Juodkazis, S.; Crawford, R.J.; Ivanova, E.P. Antibacterial titanium nano-patterned arrays inspired by dragonfly wings. Sci. Rep. 2015, 5, 16817. [Google Scholar] [CrossRef] [PubMed]
- Wandiyanto, J.V.; Cheeseman, S.; Truong, V.K.; Al Kobaisi, M.; Bizet, C.; Juodkazis, S.; Thissen, H.; Crawford, R.J.; Ivanova, E.P. Outsmarting superbugs: Bactericidal activity of nanostructured titanium surfaces against methicillin-and gentamicin-resistant Staphylococcus aureus ATCC 33592. J. Mater. Chem. B 2019, 7, 4424–4431. [Google Scholar] [CrossRef]
- Hasan, J.; Jain, S.; Padmarajan, R.; Purighalla, S.; Sambandamurthy, V.K.; Chatterjee, K. Multi-scale surface topography to minimize adherence and viability of nosocomial drug-resistant bacteria. Mater. Des. 2018, 140, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Hasan, J.; Bright, R.; Hayles, A.; Palms, D.; Zilm, P.; Barker, D.; Vasilev, K. Preventing Peri-implantitis: The Quest for a Next Generation of Titanium Dental Implants. ACS Biomater. Sci. Eng. 2022, 8, 4697–4737. [Google Scholar] [CrossRef]
- Wood, J.; Hayles, A.; Bright, R.; Palms, D.; Vasilev, K.; Hasan, J. Nanomechanical tribological characterisation of nanostructured titanium alloy surfaces using AFM: A friction vs velocity study. Colloids Surf. B Biointerfaces 2022, 217, 112600. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, L.D.; Hasan, J.; Jain, A.; Nagalingam, R.S. A nanopillar array on black titanium prepared by reactive ion etching augments cardiomyogenic commitment of stem cells. Nanoscale 2019, 11, 20766–20776. [Google Scholar] [CrossRef]
- Kasraei, S.; Azarsina, M. Addition of silver nanoparticles reduces the wettability of methacrylate and silorane-based composites. Braz. Oral Res. 2012, 26, 505–510. [Google Scholar] [CrossRef]
- Boinovich, L.B.; Modin, E.B.; Aleshkin, A.V.; Emelyanenko, K.A.; Zulkarneev, E.R.; Kiseleva, I.A.; Vasiliev, A.L.; Emelyanenko, A.M. Effective antibacterial nanotextured surfaces based on extreme wettability and bacteriophage seeding. ACS Appl. Nano Mater. 2018, 1, 1348–1359. [Google Scholar] [CrossRef]
- Bhadra, C.M.; Werner, M.; Baulin, V.A.; Truong, V.K.; Kobaisi, M.A.; Nguyen, S.H.; Balcytis, A.; Juodkazis, S.; Wang, J.Y.; Mainwaring, D.E. Subtle variations in surface properties of black silicon surfaces influence the degree of bactericidal efficiency. Nano-Micro Lett. 2018, 10, 1–8. [Google Scholar] [CrossRef]
- Linklater, D.P.; Nguyen, H.K.D.; Bhadra, C.M.; Juodkazis, S.; Ivanova, E.P. Influence of nanoscale topology on bactericidal efficiency of black silicon surfaces. Nanotechnology 2017, 28, 245301. [Google Scholar] [CrossRef]
- Nakade, K.; Jindai, K.; Sagawa, T.; Kojima, H.; Shimizu, T.; Shingubara, S.; Ito, T. Adhesion and bactericidal properties of a wettability-controlled artificial nanostructure. ACS Appl. Nano Mater. 2018, 1, 5736–5741. [Google Scholar] [CrossRef]
- Valiei, A.; Lin, N.; McKay, G.; Nguyen, D.; Moraes, C.; Hill, R.J.; Tufenkji, N. Surface Wettability Is a Key Feature in the Mechano-Bactericidal Activity of Nanopillars. ACS Appl. Mater. Interfaces 2022, 14, 27564–27574. [Google Scholar] [CrossRef]
- Arias, S.L.; Devorkin, J.; Spear, J.C.; Civantos, A.; Allain, J.P. Bacterial Envelope Damage Inflicted by Bioinspired Nanostructures Grown in a Hydrogel. ACS Appl. Bio Mater. 2020, 3, 7974–7988. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zuber, F.; Maniura-Weber, K.; Brugger, J.; Ren, Q. Nanostructured surface topographies have an effect on bactericidal activity. J. Nanobiotechnology 2018, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-K.; Cho, Y.-S.; Park, H.-H. PEGDMA-Based Pillar-Shape Nanostructured Antibacterial Films Having Mechanical Robustness. ACS Appl. Bio Mater. 2022, 5, 3006–3012. [Google Scholar] [CrossRef]
- Fontelo, R.; da Costa, D.S.; Reis, R.; Novoa-Carballal, R.; Pashkuleva, I. Bactericidal nanopatterns generated by block copolymer self-assembly. Acta Biomater. 2020, 112, 174–181. [Google Scholar] [CrossRef]
- Choi, G.; Song, Y.; Lim, H.; Lee, S.H.; Lee, H.K.; Lee, E.; Choi, B.G.; Lee, J.J.; Im, S.G.; Lee, K.G. Antibacterial nanopillar array for an implantable intraocular lens. Adv. Healthc. Mater. 2020, 9, 2000447. [Google Scholar] [CrossRef]
- Jaggessar, A.; Shahali, H.; Mathew, A.; Yarlagadda, P.K. Bio-mimicking nano and micro-structured surface fabrication for antibacterial properties in medical implants. J. Nanobiotechnology 2017, 15, 1–20. [Google Scholar] [CrossRef]
- Mo, S.; Mehrjou, B.; Tang, K.; Wang, H.; Huo, K.; Qasim, A.M.; Wang, G.; Chu, P.K. Dimensional-dependent antibacterial behavior on bioactive micro/nano polyetheretherketone (PEEK) arrays. Chem. Eng. J. 2020, 392, 123736. [Google Scholar] [CrossRef]
- Hasan, J.; Raj, S.; Yadav, L.; Chatterjee, K. Engineering a nanostructured “super surface” with superhydrophobic and superkilling properties. RSC Adv. 2015, 5, 44953–44959. [Google Scholar] [CrossRef]
- Román-Kustas, J.; Hoffman, J.B.; Reed, J.H.; Gonsalves, A.E.; Oh, J.; Li, L.; Hong, S.; Jo, K.D.; Dana, C.E.; Miljkovic, N. Molecular and topographical organization: Influence on cicada wing wettability and bactericidal properties. Adv. Mater. Interfaces 2020, 7, 2000112. [Google Scholar] [CrossRef]
- Jenkins, J.; Mantell, J.; Neal, C.; Gholinia, A.; Verkade, P.; Nobbs, A.H.; Su, B. Antibacterial effects of nanopillar surfaces are mediated by cell impedance, penetration and induction of oxidative stress. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Jiang, R.; Liu, Z.; Dou, H.; Song, L.; Tian, L.; Ming, W.; Ren, L.; Zhao, J. Bioinspired nanopillar surface for switchable mechano-bactericidal and releasing actions. J. Hazard. Mater. 2022, 432, 128685. [Google Scholar] [CrossRef]
- Xiao, K.; Cao, X.; Chen, X.; Hu, H.; Wu, C. Bactericidal efficacy of nanopatterned surface tuned by topography. J. Appl. Phys. 2020, 128, 064701. [Google Scholar] [CrossRef]
- Luan, Y.; Liu, S.; Pihl, M.; van der Mei, H.C.; Liu, J.; Hizal, F.; Choi, C.-H.; Chen, H.; Ren, Y.; Busscher, H.J. Bacterial interactions with nanostructured surfaces. Curr. Opin. Colloid Interface Sci. 2018, 38, 170–189. [Google Scholar] [CrossRef]
- Cui, Q.; Liu, T.; Li, X.; Song, K.; Ge, D. Nanopillared polycarbonate surfaces having variable feature parameters as bactericidal coatings. ACS Appl. Nano Mater. 2020, 3, 4599–4609. [Google Scholar] [CrossRef]
- Hazell, G.; May, P.W.; Taylor, P.; Nobbs, A.H.; Welch, C.; Su, B. Studies of black silicon and black diamond as materials for antibacterial surfaces. Biomater. Sci. 2018, 6, 1424–1432. [Google Scholar] [CrossRef]
- Linklater, D.P.; De Volder, M.; Baulin, V.A.; Werner, M.; Jessl, S.; Golozar, M.; Maggini, L.; Rubanov, S.; Hanssen, E.; Juodkazis, S. High aspect ratio nanostructures kill bacteria via storage and release of mechanical energy. ACS Nano 2018, 12, 6657–6667. [Google Scholar] [CrossRef]
- Michalska, M.; Gambacorta, F.; Divan, R.; Aranson, I.S.; Sokolov, A.; Noirot, P.; Laible, P.D. Tuning antimicrobial properties of biomimetic nanopatterned surfaces. Nanoscale 2018, 10, 6639–6650. [Google Scholar] [CrossRef]
- Kelleher, S.M.; Habimana, O.; Lawler, J.; O’reilly, B.; Daniels, S.; Casey, E.; Cowley, A. Cicada wing surface topography: An investigation into the bactericidal properties of nanostructural features. ACS Appl. Mater. Interfaces 2016, 8, 14966–14974. [Google Scholar] [CrossRef]
- Yu, Q.; Cho, J.; Shivapooja, P.; Ista, L.K.; López, G.P. Nanopatterned smart polymer surfaces for controlled attachment, killing, and release of bacteria. ACS Appl. Mater. Interfaces 2013, 5, 9295–9304. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.S.; Green, D.W.; Watson, J.A.; Zhou, Z.; Li, X.; Cheung, G.S.; Gellender, M. A simple model for binding and rupture of bacterial cells on nanopillar surfaces. Adv. Mater. Interfaces 2019, 6, 1801646. [Google Scholar] [CrossRef]
- Li, X. Bactericidal mechanism of nanopatterned surfaces. Phys. Chem. Chem. Phys. 2016, 18, 1311–1316. [Google Scholar] [CrossRef]
- Li, X.; Chen, T. Enhancement and suppression effects of a nanopatterned surface on bacterial adhesion. Phys. Rev. E 2016, 93, 052419. [Google Scholar] [CrossRef]
- Xue, F.; Liu, J.; Guo, L.; Zhang, L.; Li, Q. Theoretical study on the bactericidal nature of nanopatterned surfaces. J. Theor. Biol. 2015, 385, 1–7. [Google Scholar] [CrossRef]
- Velic, A.; Tesfamichael, T.; Li, Z.; Yarlagadda, P.K. Parametric study on nanopattern bactericidal activity. Procedia Manuf. 2019, 30, 514–521. [Google Scholar] [CrossRef]
- Velic, A.; Hasan, J.; Li, Z.; Yarlagadda, P.K. Mechanics of bacterial interaction and death on nanopatterned surfaces. Biophys. J. 2021, 120, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Carstens, A. How Cicada Wings Kill Bacteria. Nanopillar Cones Covering Cicada Wings Bond with Bacterial Membranes, Stretching the Portion between the Cones to the Point of Rupturing. Biological Strategy. 8 April 2021. Available online: https://asknature.org/strategy/how-cicada-wings-kill-bacteria/ (accessed on 14 October 2022).
- Bose, P.; Henderson, E. What Are the Differences between Eukaryotes and Prokaryotes? 2022. Available online: https://www.azolifesciences.com/article/What-are-the-Differences-between-Eukaryotes-and-Prokaryotes.aspx (accessed on 10 March 2022).
- Cui, Q.; Liu, T.; Li, X.; Zhao, L.; Wu, Q.; Wang, X.; Song, K.; Ge, D. Validation of the mechano-bactericidal mechanism of nanostructured surfaces with finite element simulation. Colloids Surf. B Biointerfaces 2021, 206, 111929. [Google Scholar] [CrossRef] [PubMed]
- Simulia, D.S. Abaqus 6.11 Theory Manual; DS SIMULIA Corp: Providence, RI, USA, 2011; Volume 524. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Modelled cell details | |
| Cell | Gram-negative, Escherichia coli (E. coli) |
| Cylindrical length | 1 μm |
| Radius of the hemispherical ends | 0.4 μm |
| Wall thickness | 0.006 μm |
| Modelled cell mechanics | |
| Turgor pressure | 0.03 MPa |
| Adhesion strength | 6.5 kPa |
| Critical elastic strength | 5 MPa |
| Tensile strength | 13 MPa |
| Young modulus | 25 MPa |
| Poisson’s ratio | 0.16 |
| Modelled nano-spikes details | |
| Cylindrical length | 0.2 μm |
| Radius of hemispherical end | 0.03 μm |
| Center to center spacing | 0.2 μm |
| Nanospike array | 3 × 6 |
| Grid Systems | Global Size (μm) | No. of Grid Elements | Simulation Time (min.) | Max. von Mises Stress MPa | Stress Increase (%) |
|---|---|---|---|---|---|
| 1 | 0.01 | 18,500 | 11 | 5.48 | - |
| 2 | 0.007 | 37,800 | 20 | 6.37 | 16.24 |
| 3 | 0.005 | 73,800 | 90 | 6.97 | 9.42 |
| 4 | 0.003 | 107,600 | 240 | 7.17 | 2.87 |
| 5 | 0.001 Selected | 143,500 | 380 | 7.22 | 0.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.; Aldawsari, F.S.S.; Yarlagadda, P.K.D.V. Finite Element Modelling of a Gram-Negative Bacterial Cell and Nanospike Array for Cell Rupture Mechanism Study. Molecules 2023, 28, 2184. https://doi.org/10.3390/molecules28052184
Islam M, Aldawsari FSS, Yarlagadda PKDV. Finite Element Modelling of a Gram-Negative Bacterial Cell and Nanospike Array for Cell Rupture Mechanism Study. Molecules. 2023; 28(5):2184. https://doi.org/10.3390/molecules28052184
Chicago/Turabian StyleIslam, Majedul, Falah Sahal S. Aldawsari, and Prasad K. D. V. Yarlagadda. 2023. "Finite Element Modelling of a Gram-Negative Bacterial Cell and Nanospike Array for Cell Rupture Mechanism Study" Molecules 28, no. 5: 2184. https://doi.org/10.3390/molecules28052184
APA StyleIslam, M., Aldawsari, F. S. S., & Yarlagadda, P. K. D. V. (2023). Finite Element Modelling of a Gram-Negative Bacterial Cell and Nanospike Array for Cell Rupture Mechanism Study. Molecules, 28(5), 2184. https://doi.org/10.3390/molecules28052184