
Imidazolyl Ethanamide Pentandioic Acid (IEPA) as Potential Radical Scavenger during Tumor Therapy in Human Hematopoietic Stem Cells


Abstract
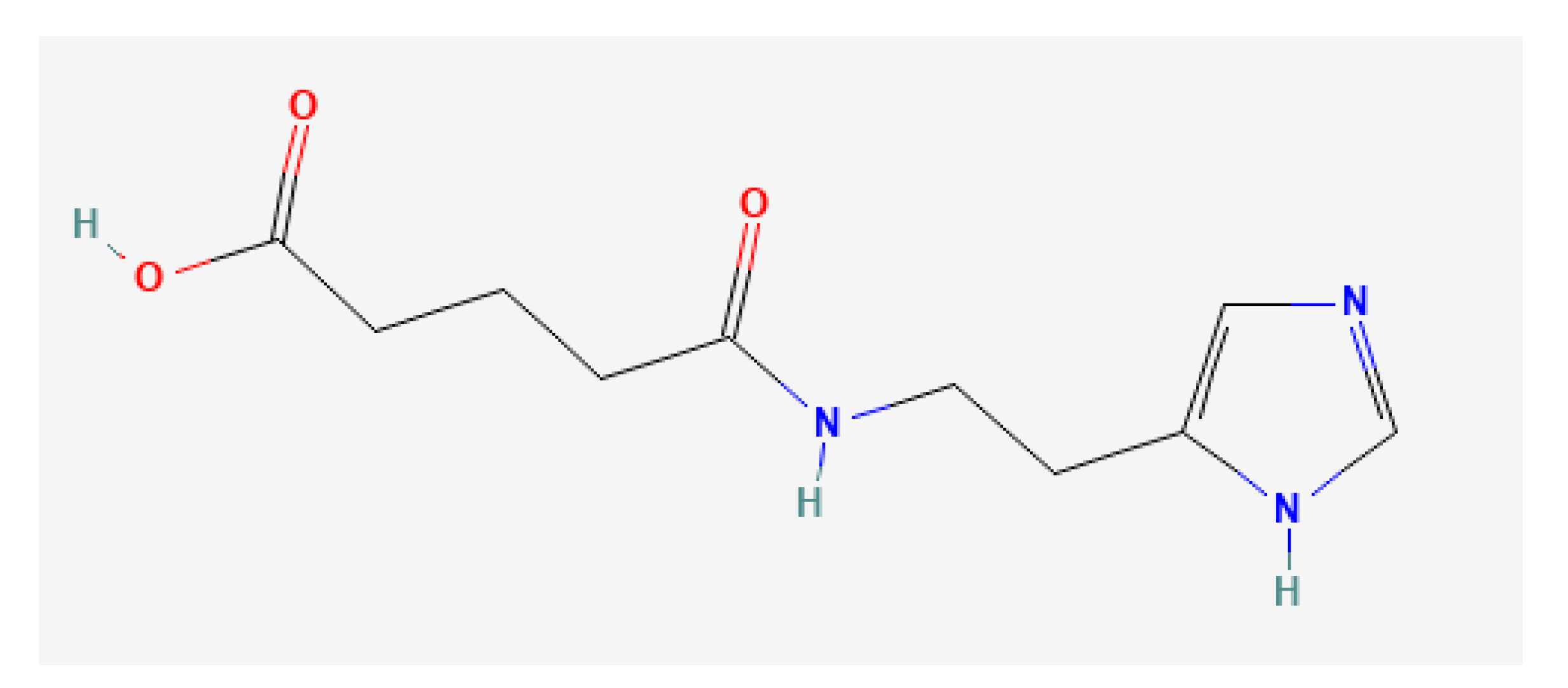
1. Introduction
2. Results
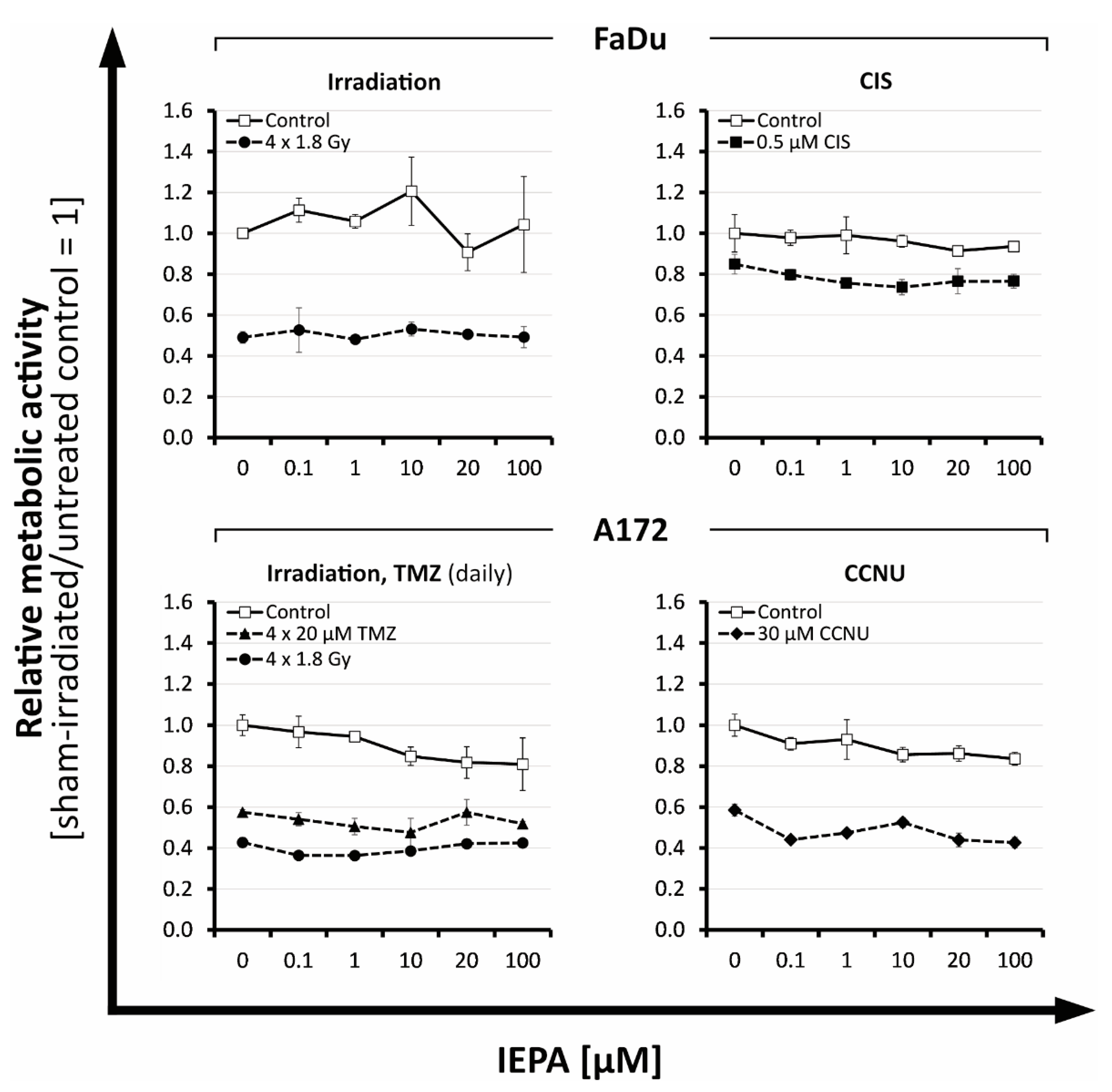
2.1. Metabolic Activity: Dose-Finding Experiments with IEPA, IR, and ChT in Tumor Cells
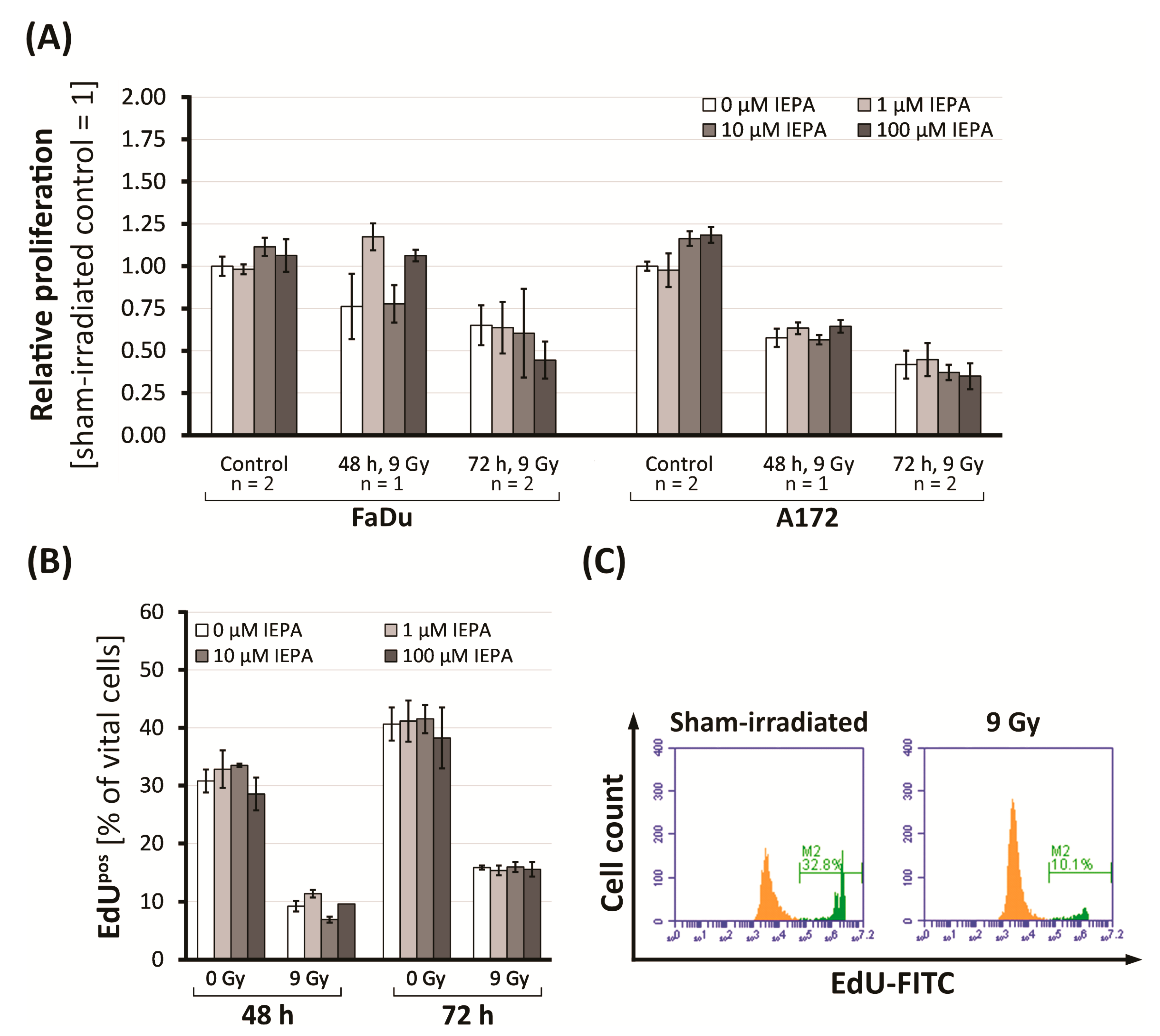
2.2. Effects of IEPA and IR on Proliferation in Tumor Cells and CD34+ HSPCs
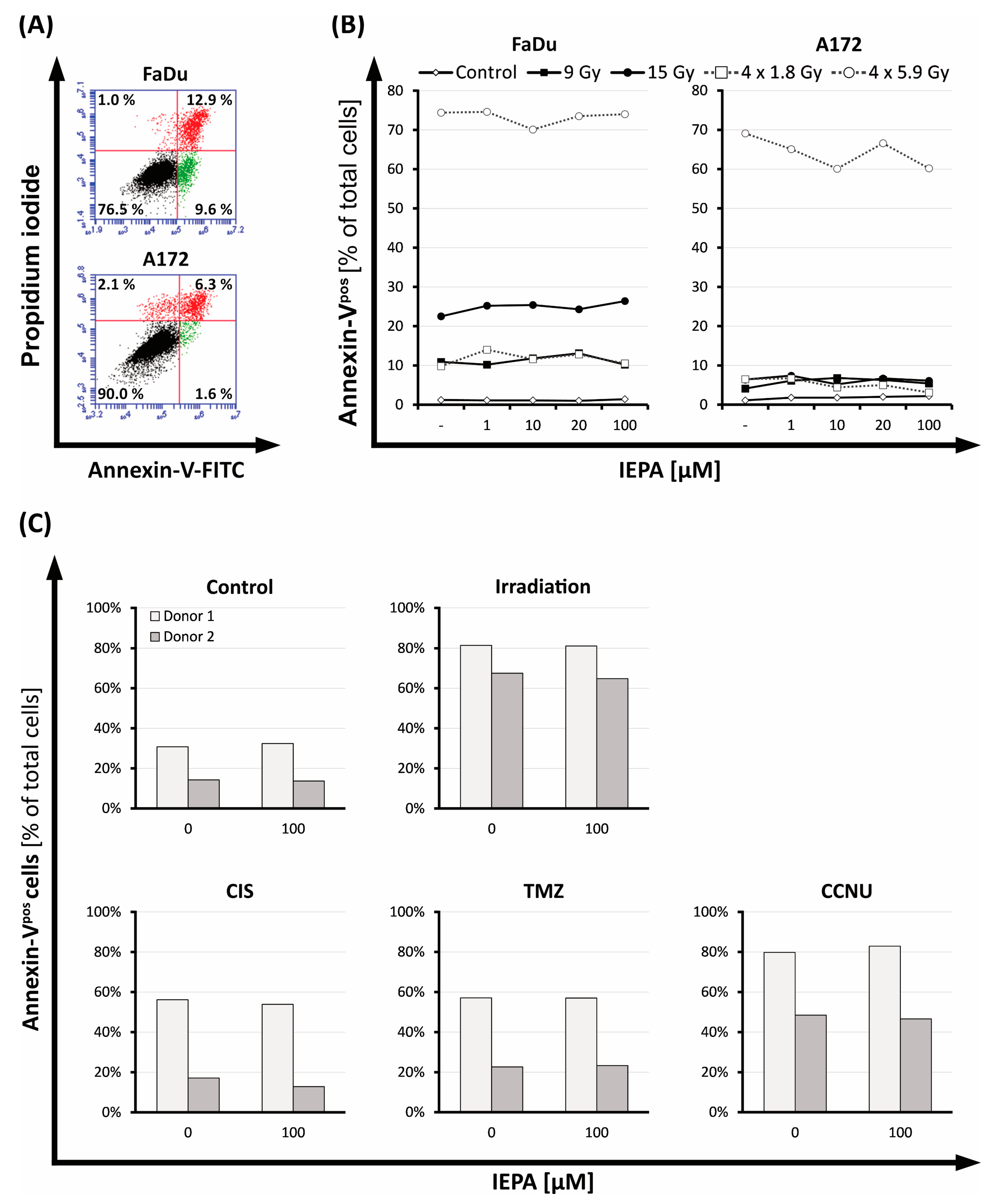
2.3. Evaluation of Cell Death after IEPA and IR or ChT Treatment in Tumor Cells and CD34+ HSPCs
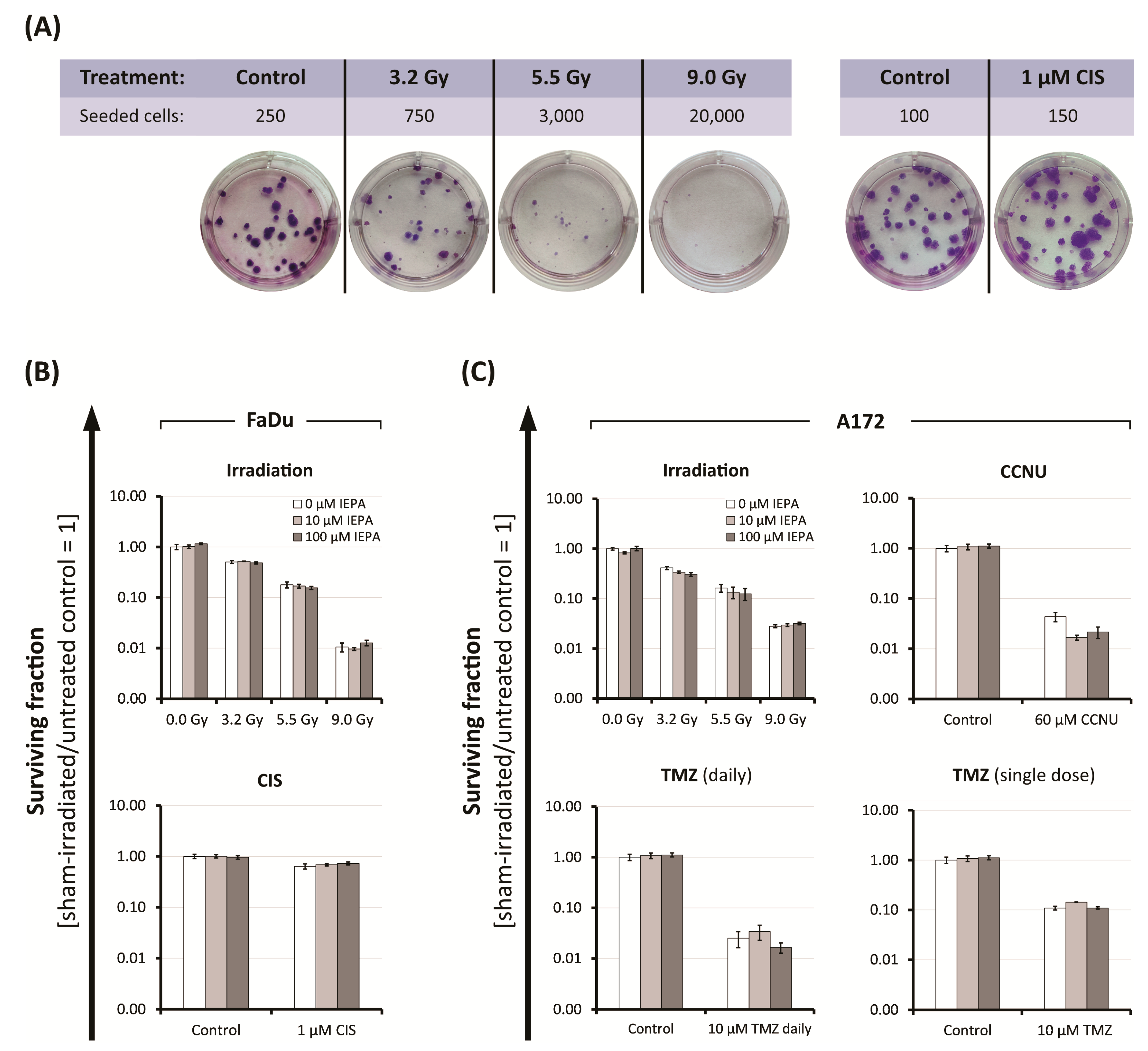
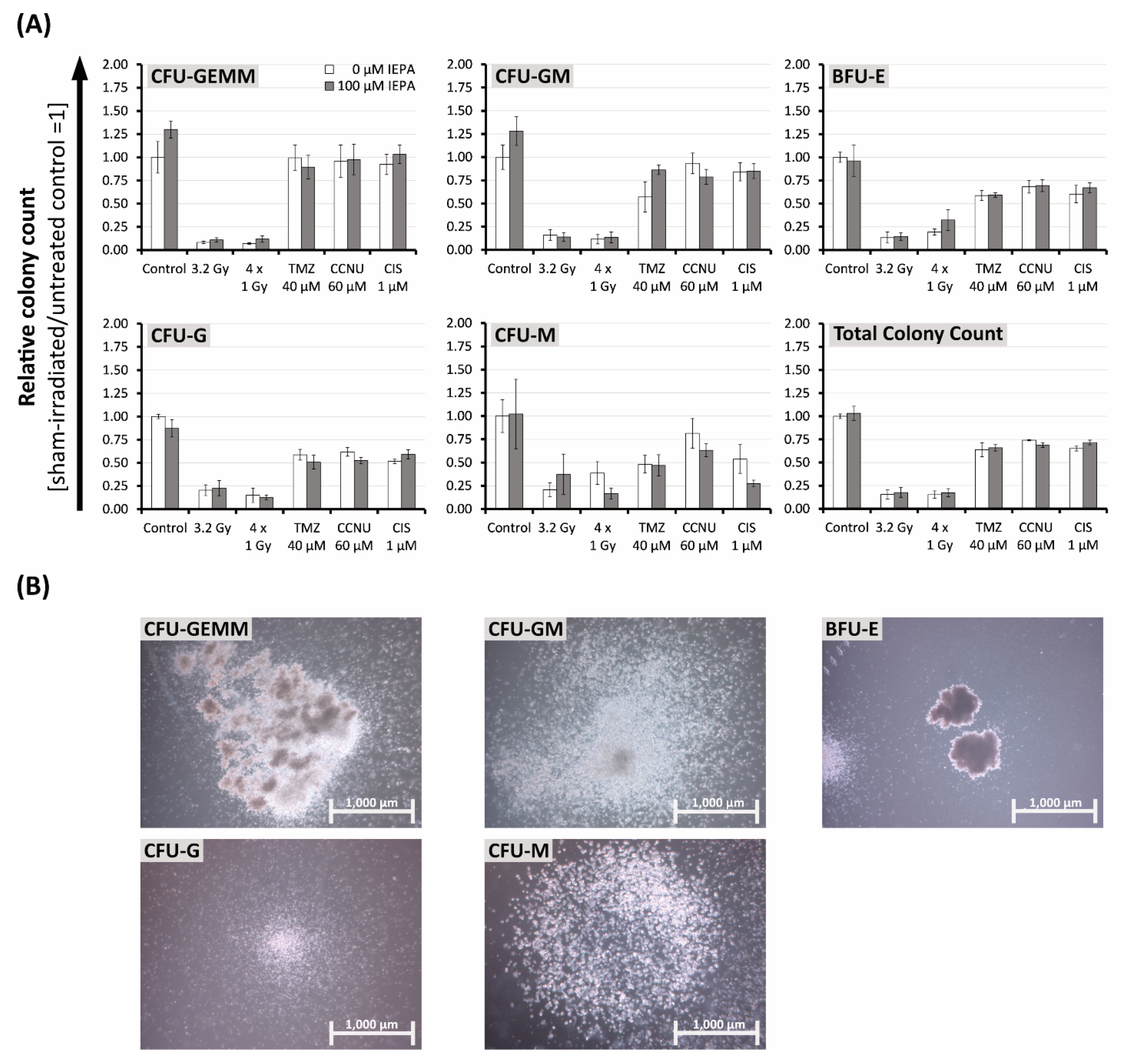
2.4. Effects of IEPA and IR or ChT Treatment on Long-Term Clonogenic Survival of Tumor Cells
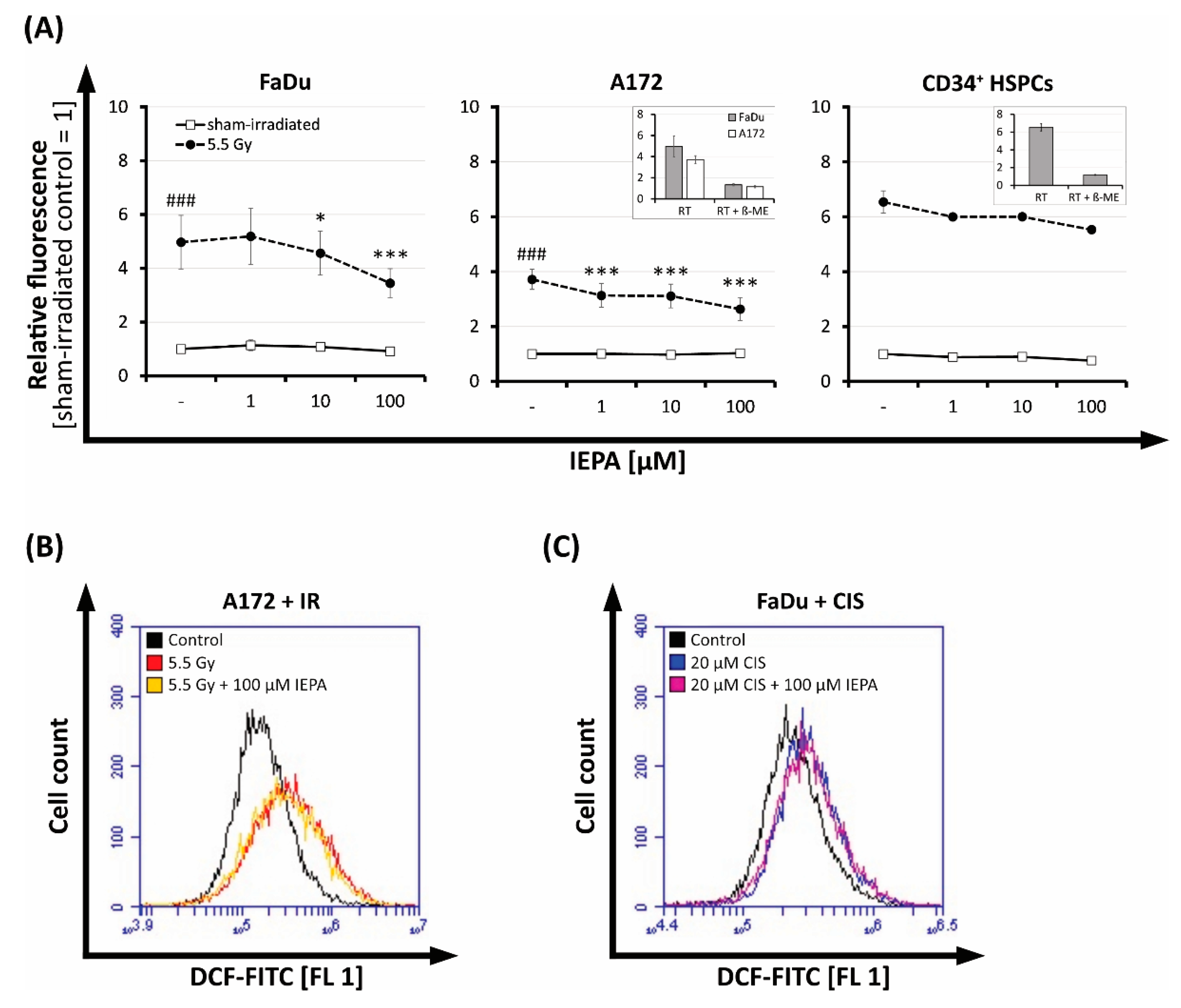
2.5. Effect of IEPA on IR- or ChT-Induced Reactive Oxygen Species (ROS) in Tumor Cells and CD34+ HSPCs
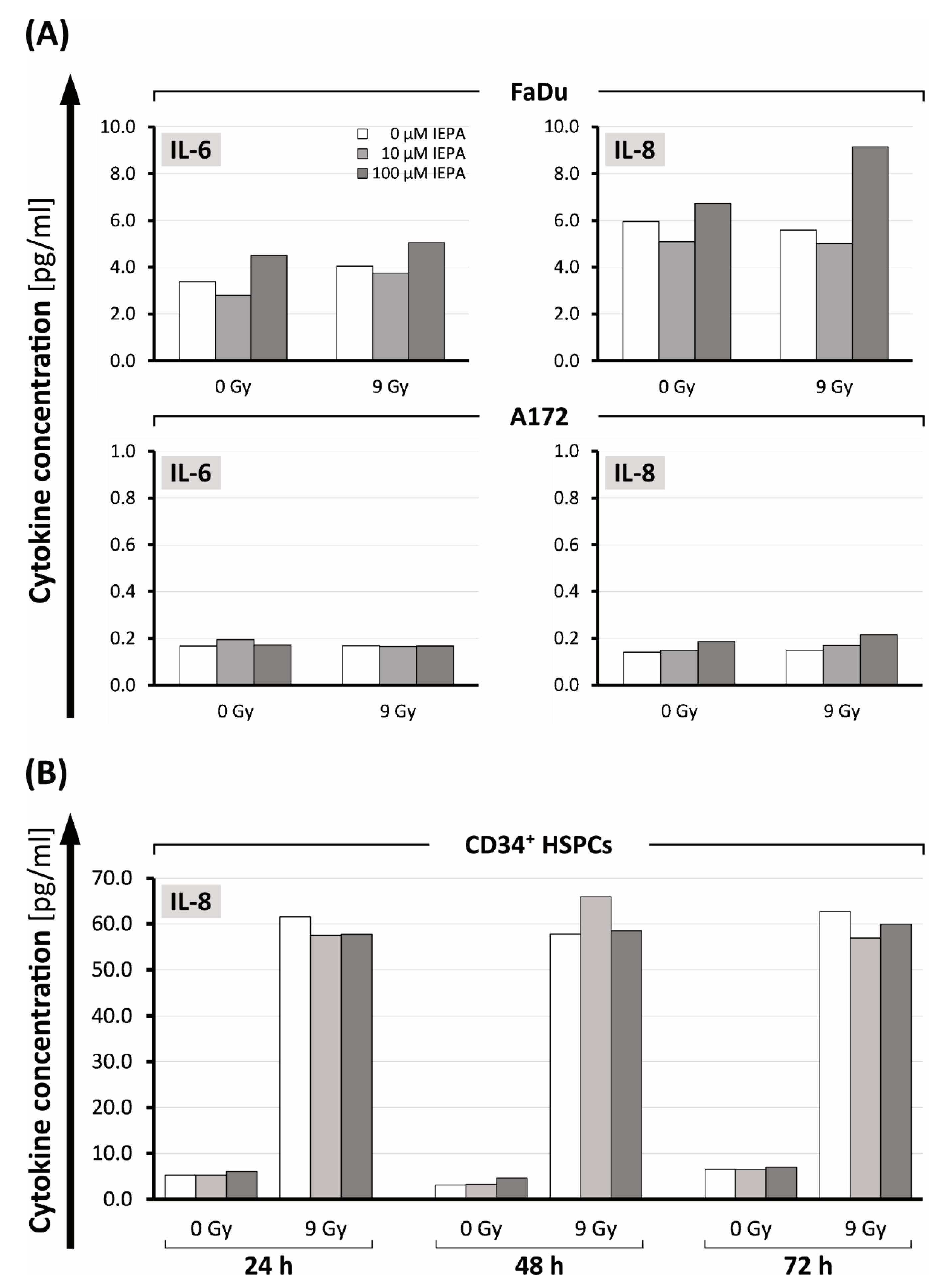
2.6. Effects of IEPA on Cytokine Release after IR of Tumor Cells and CD34+ HSPCs
2.7. Effects of IEPA and IR or ChT Treatment on the Differentiation Behavior of CD34+ HSCPs after Treatment
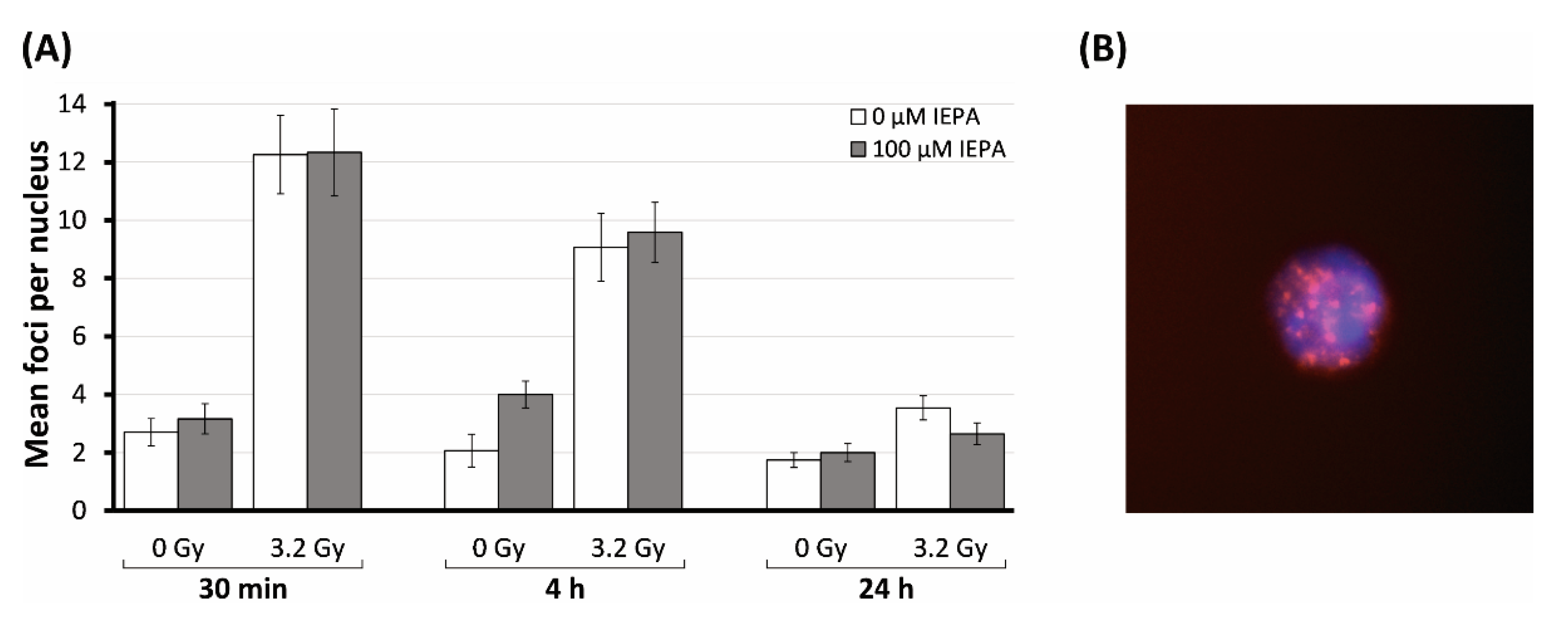
2.8. Effects of IEPA on Initialization and Repair Kinetic of IR-Induced DNA Double-Strand Breaks
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture of Tumor Cell Lines
4.3. Isolation and Culture of Human CD34+ Cells
4.4. Ionizing Radiation (IR)
1st binominal formula |
4.5. Treatment Schedule
4.6. Metabolic Activity (WST-1)
4.7. Cell Proliferation (EdU, BrdU)
4.8. Apoptosis (Annexin-V)
4.9. Clonogenic Survival of Tumor Cells (Clonogenic Assay)
4.10. Reactive Oxygen Species (DCFDA)
4.11. Cytokine Release (CBA)
4.12. Differentiation of CD34+ HSCPs (CFU Assay)
4.13. DNA Double-Strand Breaks by γH2AX Assay
4.14. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Stone, H.B.; Moulder, J.E.; Coleman, C.N.; Ang, K.K.; Anscher, M.S.; Barcellos-Hoff, M.H.; Dynan, W.S.; Fike, J.R.; Grdina, D.J.; Greenberger, J.S.; et al. Models for evaluating agents intended for the prophylaxis, mitigation and treatment of radiation injuries. Report of an NCI Workshop, December 3–4, 2003. Radiat. Res. 2004, 162, 711–728. [Google Scholar] [CrossRef] [PubMed]
- Aliper, A.M.; Bozdaganyan, M.E.; Sarkisova, V.A.; Veviorsky, A.P.; Ozerov, I.V.; Orekhov, P.S.; Korzinkin, M.B.; Moskalev, A.; Zhavoronkov, A.; Osipov, A.N. Radioprotectors.org: An open database of known and predicted radioprotectors. Aging 2020, 12, 15741–15755. [Google Scholar] [CrossRef]
- Singh, V.K.; Hanlon, B.K.; Santiago, P.T.; Seed, T.M. A review of radiation countermeasures focusing on injury-specific medicinals and regulatory approval status: Part III. Countermeasures under early stages of development along with ‘standard of care’ medicinal and procedures not requiring regulatory approval for use. Int. J. Radiat. Biol. 2017, 93, 885–906. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Seed, T.M. The efficacy and safety of amifostine for the acute radiation syndrome. Expert Opin. Drug Saf. 2019, 18, 1077–1090. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Probin, V.; Zhou, D. Cancer therapy-induced residual bone marrow injury-Mechanisms of induction and implication for therapy. Curr. Cancer Ther. Rev. 2006, 2, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Machiels, J.-P.; René Leemans, C.; Golusinski, W.; Grau, C.; Licitra, L.; Gregoire, V. Squamous cell carcinoma of the oral cavity, larynx, oropharynx and hypopharynx: EHNS-ESMO-ESTRO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2020, 31, 1462–1475. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; van den Bent, M.; Preusser, M.; Le Rhun, E.; Tonn, J.C.; Minniti, G.; Bendszus, M.; Balana, C.; Chinot, O.; Dirven, L.; et al. EANO guidelines on the diagnosis and treatment of diffuse gliomas of adulthood. Nat. Rev. Clin. Oncol. 2021, 18, 170–186. [Google Scholar] [CrossRef]
- Oosting, S.F.; Haddad, R.I. Best Practice in Systemic Therapy for Head and Neck Squamous Cell Carcinoma. Front. Oncol. 2019, 9, 815. [Google Scholar] [CrossRef]
- Noronha, V.; Joshi, A.; Patil, V.M.; Agarwal, J.; Ghosh-Laskar, S.; Budrukkar, A.; Murthy, V.; Gupta, T.; D’Cruz, A.K.; Banavali, S.; et al. Once-a-Week Versus Once-Every-3-Weeks Cisplatin Chemoradiation for Locally Advanced Head and Neck Cancer: A Phase III Randomized Noninferiority Trial. J. Clin. Oncol. 2018, 36, 1064–1072. [Google Scholar] [CrossRef]
- Saran, F.; Chinot, O.L.; Henriksson, R.; Mason, W.; Wick, W.; Cloughesy, T.; Dhar, S.; Pozzi, E.; Garcia, J.; Nishikawa, R. Bevacizumab, temozolomide, and radiotherapy for newly diagnosed glioblastoma: Comprehensive safety results during and after first-line therapy. Neuro Oncol. 2016, 18, 991–1001. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; Le Rhun, E. How did lomustine become standard of care in recurrent glioblastoma? Cancer Treat. Rev. 2020, 87, 102029. [Google Scholar] [CrossRef] [PubMed]
- Attia, S.M. The impact of quercetin on cisplatin-induced clastogenesis and apoptosis in murine marrow cells. Mutagenesis 2010, 25, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Go, R.S.; Adjei, A.A. Review of the comparative pharmacology and clinical activity of cisplatin and carboplatin. J. Clin. Oncol. 1999, 17, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Ang, K.; Zhang, Q.; Wheeler, R.H.; Rosenthal, D.I.; Nguyen-Tan, F.; Kim, H.; Lu, C.; Axelrod, R.S.; Silverman, C.I.; Weber, R.S. A phase III trial (RTOG 0129) of two radiation-cisplatin regimens for head and neck carcinomas (HNC): Impact of radiation and cisplatin intensity on outcome. JCO 2010, 28, 5507. [Google Scholar] [CrossRef]
- Bhattacharjee, A.; Noronha, V.; Patil, V.M.; Abhayankar, A.; Joshi, A.; Menon, N.S.; Prabhash, K. Optimal cumulative cisplatin dose for radio-sensitization in locally advanced head and neck cancer. JCO 2020, 38, e18553. [Google Scholar] [CrossRef]
- Ludin, A.; Gur-Cohen, S.; Golan, K.; Kaufmann, K.B.; Itkin, T.; Medaglia, C.; Lu, X.-J.; Ledergor, G.; Kollet, O.; Lapidot, T. Reactive oxygen species regulate hematopoietic stem cell self-renewal, migration and development, as well as their bone marrow microenvironment. Antioxid. Redox Signal. 2014, 21, 1605–1619. [Google Scholar] [CrossRef]
- Winkler, I.G.; Barbier, V.; Wadley, R.; Zannettino, A.C.W.; Williams, S.; Lévesque, J.-P. Positioning of bone marrow hematopoietic and stromal cells relative to blood flow in vivo: Serially reconstituting hematopoietic stem cells reside in distinct nonperfused niches. Blood 2010, 116, 375–385. [Google Scholar] [CrossRef]
- Al-Samkari, H.; Parnes, A.D.; Goodarzi, K.; Weitzman, J.I.; Connors, J.M.; Kuter, D.J. A multicenter study of romiplostim for chemotherapy-induced thrombocytopenia in solid tumors and hematologic malignancies. Haematologica 2021, 106, 1148–1157. [Google Scholar] [CrossRef]
- Klastersky, J.; de Naurois, J.; Rolston, K.; Rapoport, B.; Maschmeyer, G.; Aapro, M.; Herrstedt, J. Management of febrile neutropaenia: ESMO Clinical Practice Guidelines. Ann. Oncol. 2016, 27, v111–v118. [Google Scholar] [CrossRef]
- Tassi, C.; Tazzari, P.L.; Bonifazi, F.; Giudice, V.; Nannetti, A.; Ricci, F.; Rizzi, S.; Bandini, G.; Conte, R. Short- and long-term haematological surveillance of healthy donors of allogeneic peripheral haematopoietic progenitors mobilized with G-CSF: A single institution prospective study. Bone Marrow Transplant. 2005, 36, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Nebolsin, V.E.; Zhdanov, V.V.; Zyuzkov, G.N.; Udut, E.V.; Khrichkova, T.Y.; Simanina, E.V.; Stavrova, L.A.; Miroshnichenko, L.A.; Chaikovskiy, A.V.; Dygai, A.M. Mechanisms of protective effect of Dicarbamin on the blood system in cytostatic treatment. Bull. Exp. Biol. Med. 2011, 150, 343–347. [Google Scholar] [CrossRef]
- Zarubaev, V.V.; Garshinina, A.V.; Kalinina, N.A.; Shtro, A.A.; Belyaevskaya, S.V.; Slita, A.V.; Nebolsin, V.E.; Kiselev, O.I. Activity of Ingavirin (6-2-(1H-Imidazol-4-yl)ethylamino-5-oxo-hexanoic Acid) Against Human Respiratory Viruses in in Vivo Experiments. Pharmacwutics 2011, 4, 1518–1534. [Google Scholar] [CrossRef] [PubMed]
- Gershanovich, M.L.; Filatova, L.V. Dicarbamin—An effective protector of myelodepression in combined treatment for Hodgkin’s disease. Vopr. Onkol. 2007, 53, 589–595. [Google Scholar] [PubMed]
- Moiseeva, I.I.; Zinov’ev, A.I.; Nikishin, S.A.; Nebol’sin, V.E. Hematoprotective effectiveness of dicarbamin in experimental post-radiation bone marrow syndrome. Vopr. Onkol. 2012, 58, 81–84. [Google Scholar] [PubMed]
- Lee, F.Y.; Workman, P.; Roberts, J.T.; Bleehen, N.M. Clinical pharmacokinetics of oral CCNU (lomustine). Cancer Chemother. Pharmacol. 1985, 14, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Agarwala, S.S.; Kirkwood, J.M. Temozolomide, a novel alkylating agent with activity in the central nervous system, may improve the treatment of advanced metastatic melanoma. Oncologist 2000, 5, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, P.; Mathew, B.S.; Das, S.; Isaiah, R.; John, S.; Prabha, R.; Fleming, D.H. Cisplatin Concentrations in Long and Short Duration Infusion: Implications for the Optimal Time of Radiation Delivery. J. Clin. Diagn. Res. 2016, 10, XC01–XC04. [Google Scholar] [CrossRef] [PubMed]
- Pleimes, D. Efficacy and Safety Phase IIa Study of Myelo001 in Chemotherapy-Induced Neutropenia (MyeloConcept): ClinicalTrials.gov Identifier: NCT02692742. 2016. Available online: https://www.clinicaltrials.gov/ct2/show/NCT02692742 (accessed on 20 January 2023).
- Pleimes, D.; Möbus, V.; Mayer, F.; Schmidt, M.; Asang, C.; Flunkert, K.; Mikus, G. Induced Myelosuppression and Recommendation for Monitoring of Absolute Neutrophil Counts in Patients Treated with Epirubicin and Cyclophosphamide in Early Breast Cancer: Results from a Phase 2a Collaborative Trial of the AGO-B and CESAR Study Groups. Blood 2018, 132, 5106. [Google Scholar] [CrossRef]
- Zakharova, E.; Lobzin, Y.V. Efficacy and Safety Study of Ingavirin® to Treat Influenza and Other Acute Respiratory Viral Infections in Patients 3-6 y.o. (ACCORD): ClinicalTrials.gov Identifier: NCT02644018. 2015. Available online: https://clinicaltrials.gov/ct2/show/NCT02644018 (accessed on 20 January 2023).
- Gordeev, I.; Kazey, V.; Kapashin, A.; Luchinkina, E.; Globenko, A.; Vladykin, A.; Jain, E.; Cherenkova, A.; Kokorin, I.; Levchuk, N.; et al. Pharmacokinetics of Pentanedioic Acid Imidazolyl Ethanamide in Healthy Volunteers. Antibiot. Chemother. 2021, 66, 19–25. [Google Scholar] [CrossRef]
- Matsui, T.; Nuryadi, E.; Komatsu, S.; Hirota, Y.; Shibata, A.; Oike, T.; Nakano, T. Robustness of Clonogenic Assays as a Biomarker for Cancer Cell Radiosensitivity. Int. J. Mol. Sci. 2019, 20, 4148. [Google Scholar] [CrossRef] [PubMed]
- Kiyota, N.; Tahara, M.; Mizusawa, J.; Kodaira, T.; Fujii, H.; Yamazaki, T.; Mitani, H.; Iwae, S.; Fujimoto, Y.; Onozawa, Y.; et al. Weekly Cisplatin Plus Radiation for Postoperative Head and Neck Cancer (JCOG1008): A Multicenter, Noninferiority, Phase II/III Randomized Controlled Trial. J. Clin. Oncol. 2022, 40, 1980–1990. [Google Scholar] [CrossRef] [PubMed]
- Herrlinger, U.; Rieger, J.; Koch, D.; Loeser, S.; Blaschke, B.; Kortmann, R.-D.; Steinbach, J.P.; Hundsberger, T.; Wick, W.; Meyermann, R.; et al. Phase II trial of lomustine plus temozolomide chemotherapy in addition to radiotherapy in newly diagnosed glioblastoma: UKT-03. J. Clin. Oncol. 2006, 24, 4412–4417. [Google Scholar] [CrossRef] [PubMed]
- Kaneyuki, Y.; Yoshino, H.; Kashiwakura, I. Involvement of intracellular reactive oxygen species and mitochondria in the radiosensitivity of human hematopoietic stem cells. J. Radiat. Res. 2012, 53, 145–150. [Google Scholar] [CrossRef]
- Azzam, E.I.; Jay-Gerin, J.-P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef]
- Floratou, K.; Giannopoulou, E.; Antonacopoulou, A.; Karakantza, M.; Adonakis, G.; Kardamakis, D.; Matsouka, P. Oxidative stress due to radiation in CD34(+) hematopoietic progenitor cells: Protection by IGF-1. J. Radiat. Res. 2012, 53, 672–685. [Google Scholar] [CrossRef]
- Gupta, S.C.; Hevia, D.; Patchva, S.; Park, B.; Koh, W.; Aggarwal, B.B. Upsides and downsides of reactive oxygen species for cancer: The roles of reactive oxygen species in tumorigenesis, prevention, and therapy. Antioxid. Redox Signal. 2012, 16, 1295–1322. [Google Scholar] [CrossRef]
- Kim, W.; Lee, S.; Seo, D.; Kim, D.; Kim, K.; Kim, E.; Kang, J.; Seong, K.M.; Youn, H.; Youn, B. Cellular Stress Responses in Radiotherapy. Cells 2019, 8, 1105. [Google Scholar] [CrossRef]
- Kawamura, K.; Qi, F.; Kobayashi, J. Potential relationship between the biological effects of low-dose irradiation and mitochondrial ROS production. J. Radiat. Res. 2018, 59, ii91–ii97. [Google Scholar] [CrossRef]
- Marullo, R.; Werner, E.; Degtyareva, N.; Moore, B.; Altavilla, G.; Ramalingam, S.S.; Doetsch, P.W. Cisplatin induces a mitochondrial-ROS response that contributes to cytotoxicity depending on mitochondrial redox status and bioenergetic functions. PLoS ONE 2013, 8, e81162. [Google Scholar] [CrossRef]
- Poljsak, B.; Milisav, I. The Role of Antioxidants in Cancer, Friends or Foes? Curr. Pharm. Des. 2018, 24, 5234–5244. [Google Scholar] [CrossRef] [PubMed]
- Schaue, D.; Kachikwu, E.L.; McBride, W.H. Cytokines in radiobiological responses: A review. Radiat. Res. 2012, 178, 505–523. [Google Scholar] [CrossRef] [PubMed]
- Cachaço, A.S.; Carvalho, T.; Santos, A.C.; Igreja, C.; Fragoso, R.; Osório, C.; Ferreira, M.; Serpa, J.; Correia, S.; Pinto-do-O, P.; et al. TNF-alpha regulates the effects of irradiation in the mouse bone marrow microenvironment. PLoS ONE 2010, 5, e8980. [Google Scholar] [CrossRef]
- Duan, L.; Aoyagi, M.; Tamaki, M.; Yoshino, Y.; Morimoto, T.; Wakimoto, H.; Nagasaka, Y.; Hirakawa, K.; Ohno, K.; Yamamoto, K. Impairment of both apoptotic and cytoprotective signalings in glioma cells resistant to the combined use of cisplatin and tumor necrosis factor alpha. Clin. Cancer Res. 2004, 10, 234–243. [Google Scholar] [CrossRef]
- Guglielmetti, S.; Taverniti, V.; Minuzzo, M.; Arioli, S.; Zanoni, I.; Stuknyte, M.; Granucci, F.; Karp, M.; Mora, D. A dairy bacterium displays in vitro probiotic properties for the pharyngeal mucosa by antagonizing group A streptococci and modulating the immune response. Infect. Immun. 2010, 78, 4734–4743. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Kashiwakura, I. Role of reactive oxygen species in the radiation response of human hematopoietic stem/progenitor cells. PLoS ONE 2013, 8, e70503. [Google Scholar] [CrossRef]
- Hayashi, T.; Hayashi, I.; Shinohara, T.; Morishita, Y.; Nagamura, H.; Kusunoki, Y.; Kyoizumi, S.; Seyama, T.; Nakachi, K. Radiation-induced apoptosis of stem/progenitor cells in human umbilical cord blood is associated with alterations in reactive oxygen and intracellular pH. Mutat. Res. 2004, 556, 83–91. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, L.; Pazhanisamy, S.K.; Li, H.; Meng, A.; Zhou, D. Total body irradiation causes residual bone marrow injury by induction of persistent oxidative stress in murine hematopoietic stem cells. Free Radic. Biol. Med. 2010, 48, 348–356. [Google Scholar] [CrossRef]
- Milyavsky, M.; Gan, O.I.; Trottier, M.; Komosa, M.; Tabach, O.; Notta, F.; Lechman, E.; Hermans, K.G.; Eppert, K.; Konovalova, Z.; et al. A distinctive DNA damage response in human hematopoietic stem cells reveals an apoptosis-independent role for p53 in self-renewal. Cell Stem Cell 2010, 7, 186–197. [Google Scholar] [CrossRef]
- Fabbrizi, M.R.; Meyer, B.; Misri, S.; Raj, S.; Zobel, C.L.; Hallahan, D.E.; Sharma, G.G. Transient PP2A inhibition alleviates normal tissue stem cell susceptibility to cell death during radiotherapy. Cell Death Dis. 2018, 9, 492. [Google Scholar] [CrossRef]
- Itkin, T.; Gur-Cohen, S.; Spencer, J.A.; Schajnovitz, A.; Ramasamy, S.K.; Kusumbe, A.P.; Ledergor, G.; Jung, Y.; Milo, I.; Poulos, M.G.; et al. Distinct bone marrow blood vessels differentially regulate haematopoiesis. Nature 2016, 532, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Jež, M.; Rožman, P.; Ivanović, Z.; Bas, T. Concise review: The role of oxygen in hematopoietic stem cell physiology. J. Cell. Physiol. 2015, 230, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhang, Y.; Miao, W.; Cheng, T. Reactive Oxygen Species and Nrf2: Functional and Transcriptional Regulators of Hematopoiesis. Oxid. Med. Cell. Longev. 2019, 2019, 5153268. [Google Scholar] [CrossRef] [PubMed]
- Simsek, T.; Kocabas, F.; Zheng, J.; Deberardinis, R.J.; Mahmoud, A.I.; Olson, E.N.; Schneider, J.W.; Zhang, C.C.; Sadek, H.A. The distinct metabolic profile of hematopoietic stem cells reflects their location in a hypoxic niche. Cell Stem Cell 2010, 7, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, J.; Hayashi, N.; Yamaguchi, M.; Monzen, S.; Kashiwakura, I. Characteristics of human CD34+ cells exposed to ionizing radiation under cytokine-free conditions. J. Radiat. Res. 2015, 56, 678–690. [Google Scholar] [CrossRef]
- Barcellos-Hoff, M.H.; Park, C.; Wright, E.G. Radiation and the microenvironment—Tumorigenesis and therapy. Nat. Rev. Cancer 2005, 5, 867–875. [Google Scholar] [CrossRef]
- Pasi, F.; Facoetti, A.; Nano, R. IL-8 and IL-6 bystander signalling in human glioblastoma cells exposed to gamma radiation. Anticancer Res. 2010, 30, 2769–2772. [Google Scholar]
- Shan, Y.; He, X.; Song, W.; Han, D.; Niu, J.; Wang, J. Role of IL-6 in the invasiveness and prognosis of glioma. Int. J. Clin. Exp. Med. 2015, 8, 9114–9120. [Google Scholar]
- McKelvey, K.J.; Hudson, A.L.; Back, M.; Eade, T.; Diakos, C.I. Radiation, inflammation and the immune response in cancer. Mamm. Genome 2018, 29, 843–865. [Google Scholar] [CrossRef]
- Hormaechea-Agulla, D.; Le, D.T.; King, K.Y. Common Sources of Inflammation and Their Impact on Hematopoietic Stem Cell Biology. Curr. Stem Cell Rep. 2020, 6, 96–107. [Google Scholar] [CrossRef]
- Sokolova, T.; Poloskov, V.; Shuvalov, A.; Burova, O.; Sokolova, Z. Signaling TLR/RLR-mechanisms of immunomodulating action of ingavirin and thymogen preparations. Russ. J. Biother. 2019, 18, 60–66. [Google Scholar] [CrossRef]
- Behringer, D.; Kresin, V.; Henschler, R.; Mertelsmann, R.; Lindemann, A. Cytokine and chemokine production by CD34+ haemopoietic progenitor cells: Detection in single cells. Br. J. Haematol. 1997, 97, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Majka, M.; Janowska-Wieczorek, A.; Ratajczak, J.; Ehrenman, K.; Pietrzkowski, Z.; Kowalska, M.A.; Gewirtz, A.M.; Emerson, S.G.; Ratajczak, M.Z. Numerous growth factors, cytokines, and chemokines are secreted by human CD34(+) cells, myeloblasts, erythroblasts, and megakaryoblasts and regulate normal hematopoiesis in an autocrine/paracrine manner. Blood 2001, 97, 3075–3085. [Google Scholar] [CrossRef]
- Laterveer, L.; Lindley, I.J.; Hamilton, M.S.; Willemze, R.; Fibbe, W.E. Interleukin-8 induces rapid mobilization of hematopoietic stem cells with radioprotective capacity and long-term myelolymphoid repopulating ability. Blood 1995, 85, 2269–2275. [Google Scholar] [CrossRef] [PubMed]
- McMahon, S.J. The linear quadratic model: Usage, interpretation and challenges. Phys. Med. Biol. 2018, 64, 01TR01. [Google Scholar] [CrossRef]
- van Leeuwen, C.M.; Oei, A.L.; Crezee, J.; Bel, A.; Franken, N.A.P.; Stalpers, L.J.A.; Kok, H.P. The alfa and beta of tumours: A review of parameters of the linear-quadratic model, derived from clinical radiotherapy studies. Radiat. Oncol. 2018, 13, 96. [Google Scholar] [CrossRef]
- Patties, I.; Kortmann, R.-D.; Glasow, A. Inhibitory effects of epigenetic modulators and differentiation inducers on human medulloblastoma cell lines. J. Exp. Clin. Cancer Res. 2013, 32, 27. [Google Scholar] [CrossRef]
- Patties, I.; Kortmann, R.-D.; Menzel, F.; Glasow, A. Enhanced inhibition of clonogenic survival of human medulloblastoma cells by multimodal treatment with ionizing irradiation, epigenetic modifiers, and differentiation-inducing drugs. J. Exp. Clin. Cancer Res. 2016, 35, 94. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50: Cytostatic Agents | FaDu | A 172 | ||
|---|---|---|---|---|
| CIS | 0.5 µM | - | ||
| TMZ daily | - | 4 × 20 µM | ||
| CCNU | - | 30 µM | ||
| ID50: Irradiation | FaDu + A172 combined | |||
| ID50 (4 fractions) Experimentally determined | 1.8 Gy/fraction | |||
| ↪ ID[BED]50 (biologically effective dose) Calculated, α/β = 10 | 8.5 Gy | |||
| ↪ ID50 (single dose) Calculated | 5.5 Gy | |||
| Multiples of ID50 | Single dose | BED | 4 fractions | |
| 0.25 × ID50 | 2.1 Gy | 2.1 Gy | 4 × 0.5 Gy | |
| 0.5 × ID50 | 3.2 Gy | 4.3 Gy | 4 × 1.0 Gy | |
| 1 × ID50 | 5.5 Gy | 8.5 Gy | 4 × 1.8 Gy | |
| 2 × ID50 | 9.0 Gy | 17.1 Gy | 4 × 3.2 Gy | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pfau, L.C.; Glasow, A.; Seidel, C.; Patties, I. Imidazolyl Ethanamide Pentandioic Acid (IEPA) as Potential Radical Scavenger during Tumor Therapy in Human Hematopoietic Stem Cells. Molecules 2023, 28, 2008. https://doi.org/10.3390/molecules28052008
Pfau LC, Glasow A, Seidel C, Patties I. Imidazolyl Ethanamide Pentandioic Acid (IEPA) as Potential Radical Scavenger during Tumor Therapy in Human Hematopoietic Stem Cells. Molecules. 2023; 28(5):2008. https://doi.org/10.3390/molecules28052008
Chicago/Turabian StylePfau, Lucas C., Annegret Glasow, Clemens Seidel, and Ina Patties. 2023. "Imidazolyl Ethanamide Pentandioic Acid (IEPA) as Potential Radical Scavenger during Tumor Therapy in Human Hematopoietic Stem Cells" Molecules 28, no. 5: 2008. https://doi.org/10.3390/molecules28052008
APA StylePfau, L. C., Glasow, A., Seidel, C., & Patties, I. (2023). Imidazolyl Ethanamide Pentandioic Acid (IEPA) as Potential Radical Scavenger during Tumor Therapy in Human Hematopoietic Stem Cells. Molecules, 28(5), 2008. https://doi.org/10.3390/molecules28052008