
Comparative Analysis of the Metabolic Profiles of Strains of Tribolium castaneum (Herbst) Adults with Different Levels of Phosphine Resistance Based on Direct Immersion Solid-Phase Microextraction and Gas Chromatography-Mass Spectrometry



Abstract
:1. Introduction
2. Results and Discussion
2.1. Phosphine Susceptibility Tests
2.2. Metabolite Expression in Response to Tribolium Castaneum (Herbst) Adults of Different Phosphine Resistance Levels
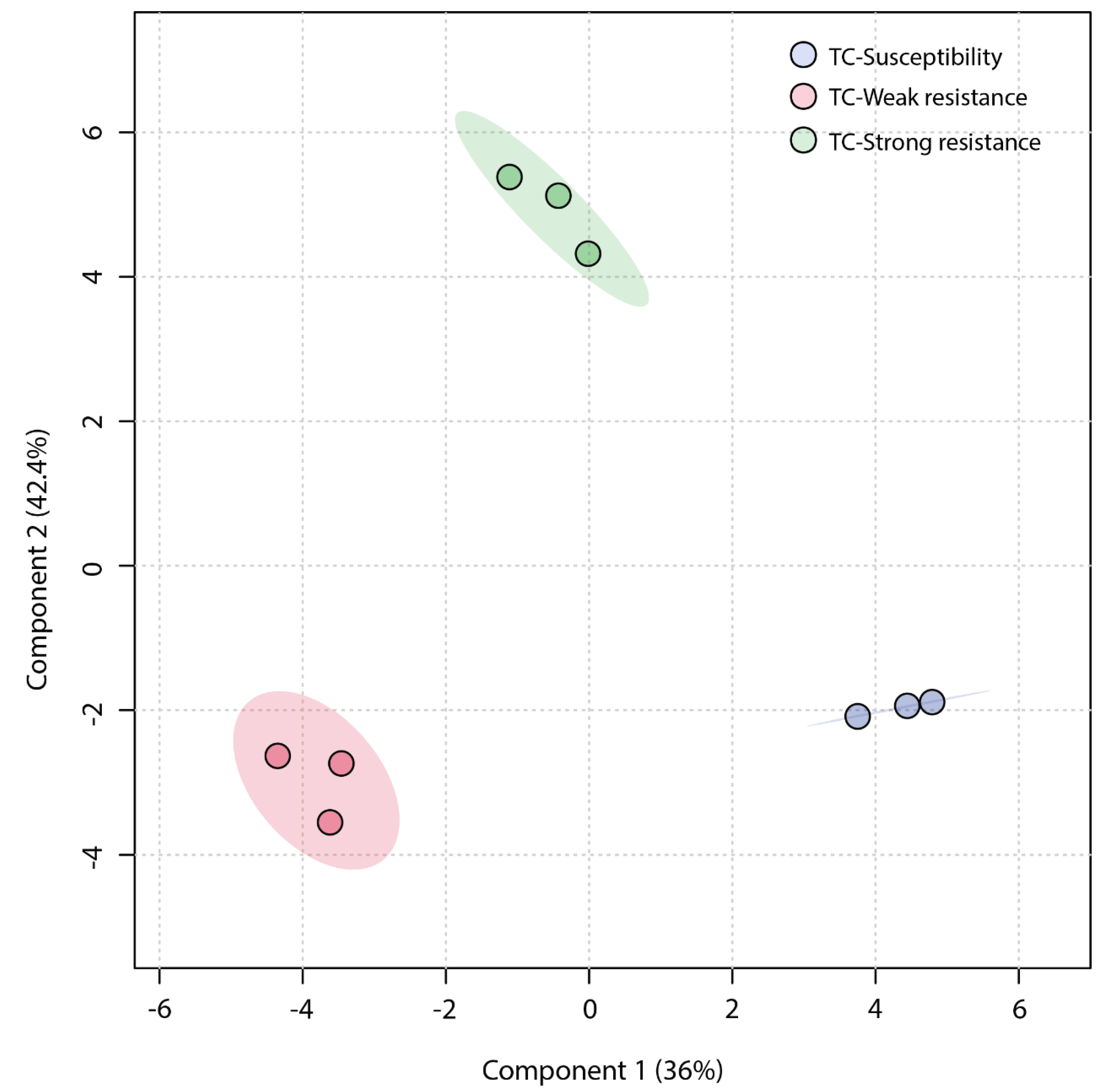
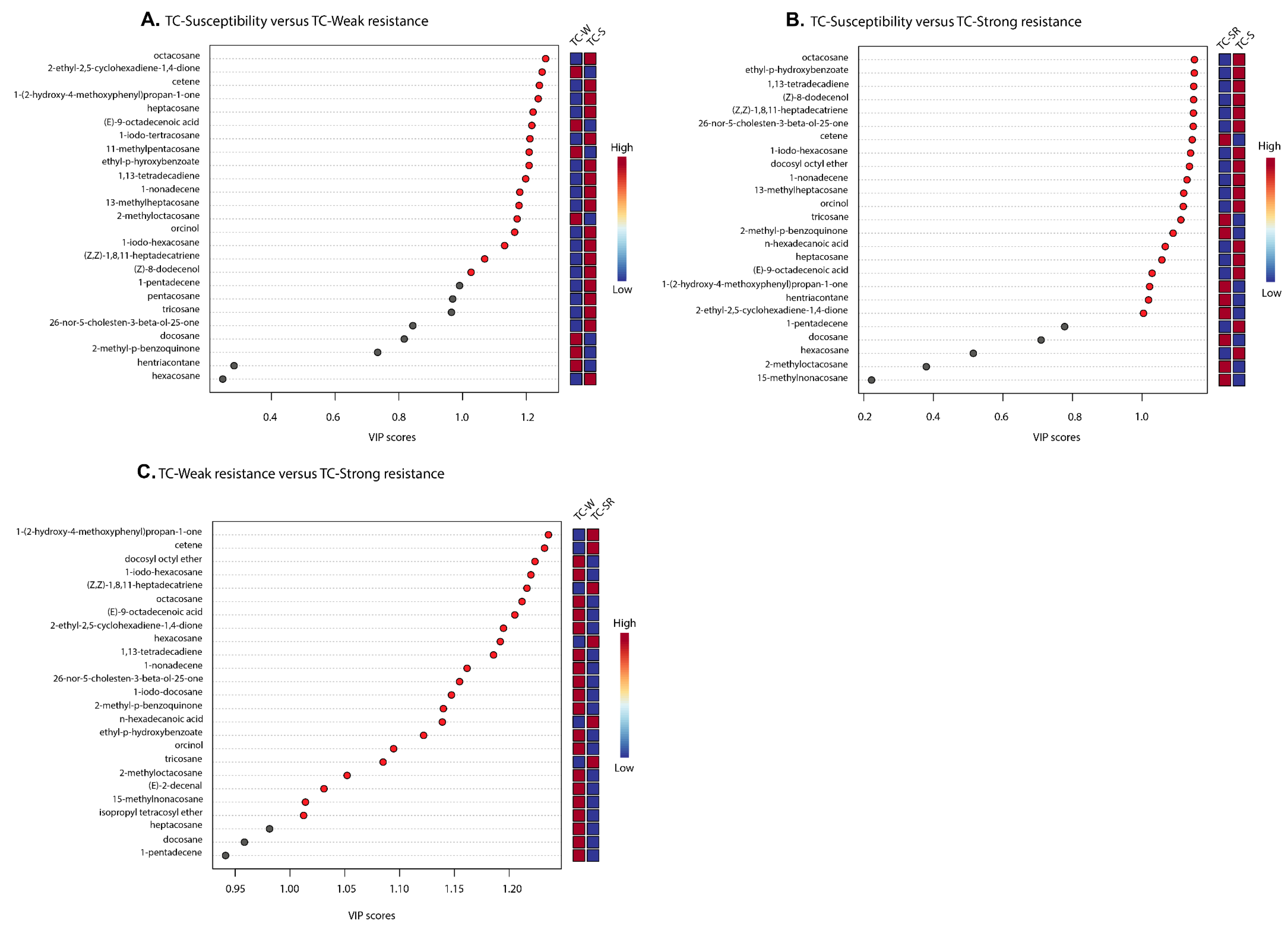
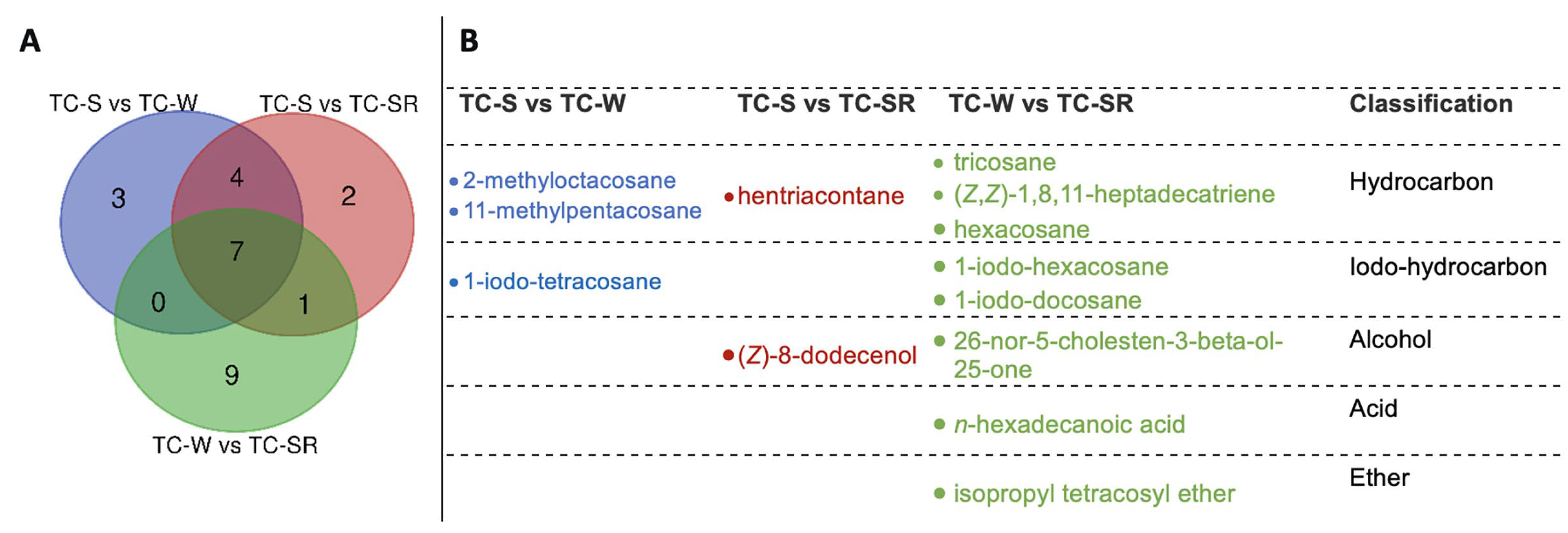
2.3. Statistical Analysis and Differentially Regulated Metabolites
3. Materials and Methods
3.1. The Insect Culture
3.2. Fumigation Bioassay
3.3. Sample Preparation and Extraction Using DI-SPME
3.4. Gas Chromatography-Mass Spectrometry (GC-MS) Conditions
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Atta, B.; Rizwan, M.; Sabir, A.M.; Gogi, M.D.; Ali, K. Damage potential of Tribolium castaneum (Herbst)(Coleoptera: Tenebrionidae) on wheat grains stored in hermetic and non-hermetic storage bags. Int. J. Trop. Insect Sci. 2020, 40, 27–37. [Google Scholar] [CrossRef]
- Cook, S.A. Evaluation of Sealed Storage Silos for Grain Fumigation. Doctoral Dissertation, Kansas State University, Manhattan, KS, USA, 2016. [Google Scholar]
- Kim, K.; Yang, J.O.; Sung, J.Y.; Lee, J.Y.; Park, J.S.; Lee, H.S.; Lee, S.E. Minimization of energy transduction confers resistance to phosphine in the rice weevil. Sitophilus oryzae. Sci. Rep. 2019, 9, 14605. [Google Scholar] [PubMed]
- Jagadeesan, R.; Singarayan, V.T.; Nayak, M.K. A co-fumigation strategy utilizing reduced rates of phosphine (PH3) and sulfuryl fluoride (SF) to control strongly resistant rusty grain beetle, Cryptolestes ferrugineus (Stephens)(Coleoptera: Laemophloeidae). Pest Manag. Sci. 2021, 77, 4009–4015. [Google Scholar] [CrossRef] [PubMed]
- Daglish, G.J.; Nayak, M.K.; Arthur, F.H.; Athanassiou, C.G. Insect pest management in stored grain. In Recent Advances in Stored Product Protection; Springer: Berlin/Heidelberg, Germany, 2018; pp. 45–63. [Google Scholar]
- Rajendran, S. Insect pest management in stored products. Outlooks Pest Manag. 2020, 31, 24–35. [Google Scholar] [CrossRef]
- FAO. Recommended methods for the detection and measurement of resistance of agricultural pests to pesticides. Tentative method for adults of some major species of stored cereals with methyl bromide and phosphine—FAO Method No 16. In FAO Plant Protection Bulletin; FAO: Rome, Italy, 1975; Volume 23, pp. 12–25. [Google Scholar]
- Cao, Y.; Wang, D. Relationship between phosphine resistance and narcotic knockdown in Tribolium castaneum (Herbst), Sitophilus oryzae (L.), and S. zeamais (Motsch). In Proceedings of the Conference Controlled Atmosphere and Fumigation in Stored-products, Fresno, CA, USA, 29 October–3 November 2000. [Google Scholar]
- Afful, E.; Cato, A.; Nayak, M.K.; Phillips, T.W. A rapid assay for the detection of resistance to phosphine in the lesser grain borer, Rhyzopertha dominica (F.)(Coleoptera: Bostrichidae). J. Stored Prod. Res. 2021, 91, 101776. [Google Scholar] [CrossRef]
- Chen, Z.; Schlipalius, D.; Opit, G.; Subramanyam, B.; Phillips, T.W. Diagnostic molecular markers for phosphine resistance in US populations of Tribolium castaneum and Rhyzopertha dominica. PLoS ONE 2015, 10, e0121343. [Google Scholar]
- Schlipalius, D.I.; Valmas, N.; Tuck, A.G.; Jagadeesan, R.; Ma, L.; Kaur, R.; Ebert, P.R. A core metabolic enzyme mediates resistance to phosphine gas. Science 2012, 338, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Nath, N.S.; Bhattacharya, I.; Tuck, A.G.; Schlipalius, D.I.; Ebert, P.R. Mechanisms of phosphine toxicity. J. Toxicol. 2011, 2011, 494168. [Google Scholar] [CrossRef]
- Schlipalius, D.I.; Tuck, A.G.; Jagadeesan, R.; Nguyen, T.; Kaur, R.; Subramanian, S.; Ebert, P.R. Variant linkage analysis using de novo transcriptome sequencing identifies a conserved phosphine resistance gene in insects. Genetics 2018, 209, 281–290. [Google Scholar] [CrossRef]
- Du, X. The Physiochemical Responses of Stored Grain Insect Pests to Synthetic Amorphous Silica (SAS) Powders. Doctoral Dissertation, Murdoch University, Murdoch, Australia, 2021. [Google Scholar]
- Kim, D.; Kim, K.; Lee, Y.H.; Lee, S.E. Transcriptome and Micro–CT analysis unravels the cuticle modification in phosphine-resistant stored grain insect pest, Tribolium castaneum (Herbst). Chem. Biol. Technol. Agric. 2023, 10, 88. [Google Scholar] [CrossRef]
- Alnajim, I.; Du, X.; Lee, B.; Agarwal, M.; Liu, T.; Ren, Y. New method of analysis of lipids in Tribolium castaneum (Herbst) and Rhyzopertha dominica (Fabricius) insects by direct immersion solid-phase microextraction (DI-SPME) coupled with GC–MS. Insects 2019, 10, 363. [Google Scholar] [CrossRef] [PubMed]
- Alnajim, I.; Agarwal, M.; Liu, T.; Li, B.; Du, X.; Ren, Y. Preliminary study on the differences in hydrocarbons between phosphine-susceptible and-resistant strains of Rhyzopertha dominica (Fabricius) and Tribolium castaneum (Herbst) using direct immersion solid-phase microextraction coupled with GC-MS. Molecules 2020, 25, 1565. [Google Scholar] [CrossRef] [PubMed]
- Alnajim, I.; Agarwal, M.; Liu, T.; Ren, Y. A novel method for the analysis of Volatile Organic Compounds (VOCs) from red flour beetle Tribolium castaneum (H.) using Headspace-SPME Technology. Curr. Anal. Chem. 2020, 16, 404–412. [Google Scholar] [CrossRef]
- Shan, C.; Li, B.; Li, L.; Li, B.; Ren, Y.; Liu, T. Correlation between irradiation treatment and metabolite changes in Bactrocera dorsalis (Diptera: Tephritidae) Larvae using solid-phase microextraction (SPME) coupled with gas chromatography-mass spectrometry (GC-MS). Molecules 2022, 27, 4641. [Google Scholar] [CrossRef] [PubMed]
- Renou, M.; Guerrero, A. Insect parapheromones in olfaction research and semiochemical-based pest control strategies. Ann. Rev. Entomol. 2000, 45, 605–630. [Google Scholar] [CrossRef] [PubMed]
- Witzgall, P.; Kirsch, P.; Cork, A. Sex pheromones and their impact on pest management. J. Chem. Ecol. 2010, 36, 80–100. [Google Scholar] [CrossRef]
- Al-Khshemawee, H.; Du, X.; Agarwal, M.; Yang, J.O.; Ren, Y.L. Application of direct immersion Solid-Phase Microextraction (DI-SPME) for understanding biological changes of Mediterranean fruit fly (Ceratitis capitata) during mating procedures. Molecules 2018, 23, 2951. [Google Scholar] [CrossRef]
- Opit, G.P.; Phillips, T.W.; Aikins, M.J.; Hasan, M.M. Phosphine resistance in Tribolium castaneum and Rhyzopertha dominica from stored wheat in Oklahoma. J. Econ. Entomol. 2012, 105, 1107–1114. [Google Scholar] [CrossRef]
- Villaverde, M.L.; Juárez, M.P.; Mijailovsky, S. Detection of Tribolium castaneum (Herbst) volatile defensive secretions by solid phase microextraction–capillary gas chromatography (SPME-CGC). J. Stored Prod. Res. 2007, 43, 540–545. [Google Scholar] [CrossRef]
- Tian, X.; Hao, J.; Wu, F.; Hu, H.; Zhou, G.; Liu, X.; Zhang, T. 1-Pentadecene, a volatile biomarker for the detection of Tribolium castaneum (Herbst)(Coleoptera: Tenebrionidae) infested brown rice under different temperatures. J. Stored Prod. Res. 2022, 97, 101981. [Google Scholar] [CrossRef]
- Wang, C.; Feng, Y.; Zhang, S.; Fu, T.; Sheng, Y.; Zhang, Y.; Zhang, D. Effects of storage on brown rice (Oryza sativa L.) metabolites, analyzed using gas chromatography and mass spectrometry. Food Sci. Nutr. 2020, 8, 2882–2894. [Google Scholar]
- Mon, K.K.; Zhu, Y.; Chanthavixay, G.; Kern, C.; Zhou, H. Integrative analysis of gut microbiome and metabolites revealed novel mechanisms of intestinal Salmonella carriage in chicken. Sci. Rep. 2020, 10, 4809. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.M.; Xu, X.; He, Y.; Wei, G.B.; Sima, Y.H.; Shi, Q.X. Metabonomic analysis of Bombyx mori (Heterocera: Bombysidae) treated with acetaminophen. J. Insect Sci. 2014, 14, 225. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, J.; Malinski, E.; Szafranek, J. Function and Composition of Cuticular Hydrocarbons of Stored-Product Insects. In Proceedings of the 6th International Working Conference on Stored-Product Protection, Canberra, Australia, 17–23 April 1994; CAB International: Wallingford, UK, 1994. [Google Scholar]
- Alnajim, I.; Aldosary, N.; Agarwal, M.; Liu, T.; Du, X.; Ren, Y. Role of lipids in phosphine resistant stored-grain insect pests Tribolium castaneum and Rhyzopertha dominica. Insects 2022, 13, 798. [Google Scholar] [CrossRef] [PubMed]
- Malekpour, R.; Rafter, M.A.; Daglish, G.J.; Walter, G.H. The movement abilities and resource location behaviour of Tribolium castaneum: Phosphine resistance and its genetic influences. J. Pest Sci. 2018, 91, 739–749. [Google Scholar] [CrossRef]
- Sakka, M.K.; Romano, D.; Stefanini, C.; Canale, A.; Benelli, G.; Athanassiou, C.G. Mobility parameters of Tribolium castaneum and Rhyzopertha dominica populations with different susceptibility to phosphine. J. Stored Prod. Res. 2020, 87, 101593. [Google Scholar] [CrossRef]
- Ridley, A.W.; Magabe, S.; Schlipalius, D.I.; Rafter, M.A.; Collins, P.J. Sublethal Exposure to Phosphine Decreases Offspring Production in Strongly. PLoS ONE 2012, 7, e53356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Liu, L.; Steffen, D.; Ye, T.; Raftery, D. Metabolic profiling of gender: Headspace-SPME/GC–MS and 1-HNMR analysis of urine. Metabolomics 2012, 8, 323–334. [Google Scholar] [CrossRef]
- Pathak, R.; Kumari, A.; Mohsin, M.; Bisht, G.; Bala, M. Phytochemical assessment and in vitro antioxidant potential of Camelina sativa L. seed cake. Asian J. Res. Chem. 2020, 13, 38–43. [Google Scholar] [CrossRef]
- Jagadeesan, R.; Collins, P.J.; Daglish, G.J.; Ebert, P.R.; Schlipalius, D.I. Phosphine resistance in the rust red flour beetle, Tribolium castaneum (Coleoptera: Tenebrionidae): Inheritance, gene interactions and fitness costs. PLoS ONE 2012, 7, e31582. [Google Scholar] [CrossRef]
- Nayak, M.K.; Daglish, G.J.; Phillips, T.W.; Ebert, P.R. Resistance to the fumigant phosphine and its management in insect pests of stored products: A global perspective. Annu. Rev. Entomol. 2020, 65, 333–350. [Google Scholar] [CrossRef] [PubMed]

 represent significant compounds selected based on the p-value threshold (<0.05), and the
represent significant compounds selected based on the p-value threshold (<0.05), and the  represent non-significant compounds (left). The number of metabolites in each chemical classification (right).
represent significant compounds selected based on the p-value threshold (<0.05), and the represent non-significant compounds (left). The number of metabolites in each chemical classification (right).
represent non-significant compounds (left). The number of metabolites in each chemical classification (right).
represent significant compounds selected based on the p-value threshold (<0.05), and the represent non-significant compounds (left). The number of metabolites in each chemical classification (right).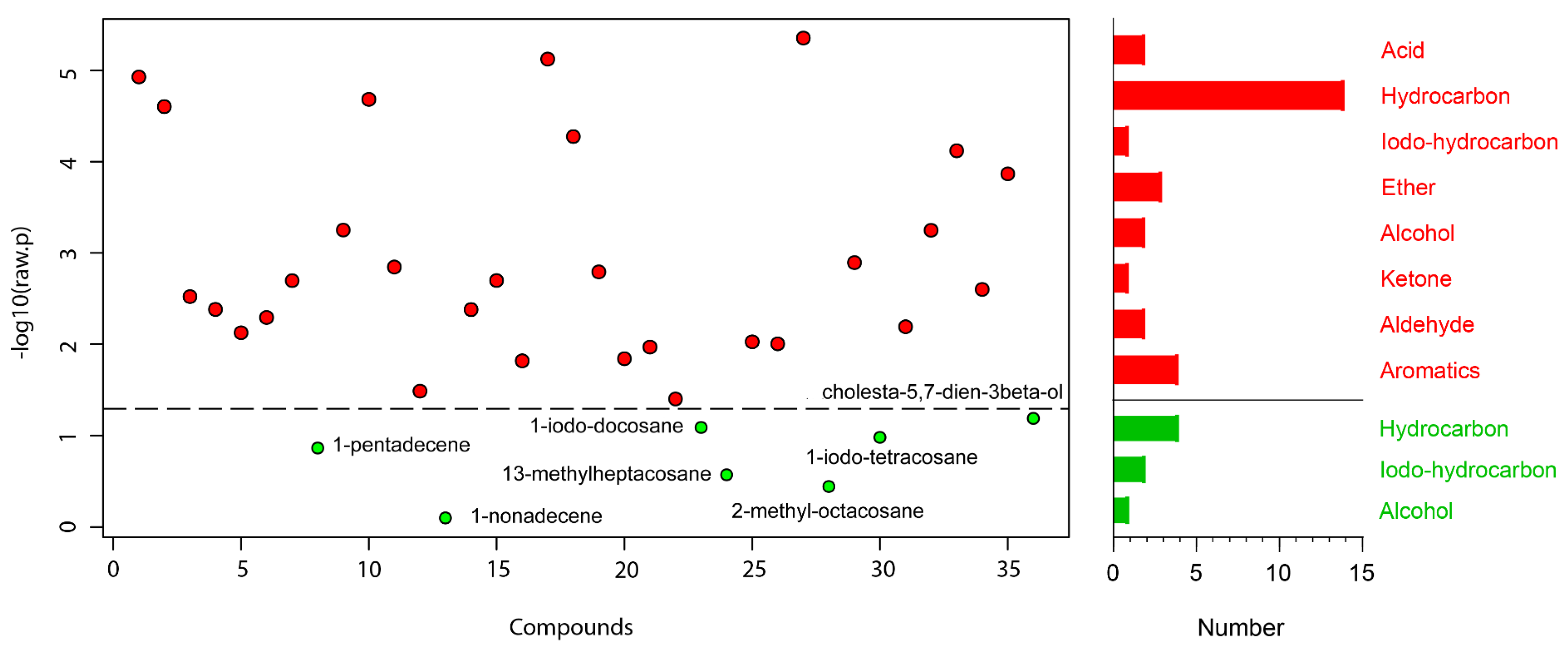




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | 1 N | Slope ± SE | LC50 (mg/L) (95% 2 FL) | LC99 (mg/L) | Heterogeneity Factor | 3 df | 4 G -Factor | 5 Mean Deviance Ratio | 6 RR (7 CL) | Classification |
|---|---|---|---|---|---|---|---|---|---|---|
| Wuhan (TC-S) | 1050 | 1.667 ± 0.242 | 0.011 (0.003, 0.014) | 0.223 | 0.520 | 16 | 0.063 | 487.084 (p < 0.001) | - | Susceptibility |
| Qihe (TC-W) | 900 | 2.037 ± 0.207 | 0.508 (0.443, 0.570) | 7.049 | 0.401 | 16 | 0.284 | 676.357 (p < 0.001) | 46.194 (40.298, 54.267) | Weak |
| Zibo (TC-SR) | 900 | 9.084 ± 0.673 | 3.114 (2.988, 3.234) | 5.615 | 0.570 | 16 | 0.027 | 585.656 (p < 0.001) | 283.050 (271.678, 294.072) | Strong |
| Metabolites | 1 RT | 2 RI (Exp) | 3 RI (Lit) | RA (%) ± 5 SD | p-Value | CAS | ||
|---|---|---|---|---|---|---|---|---|
| TC-S | TC-W | TC-SR | ||||||
| 2-methyl-p-benzoquinone | 6.961 | 1042 | 1018 | 0.18 ± 0.02 b | 0.47 ± 0.02 a | 0.49 ± 0.02 a | 0.0000118 | 553-97-9 |
| 2-ethyl-2,5-cyclohexadiene-1,4-dione | 8.915 | 1129 | 1215 | 0.3 ± 0.01 c | 0.69 ± 0.01 a | 0.54 ± 0.05 b | 0.0000250 | 4754-26-1 |
| (E)-2-decenal | 12.137 | 1225 | 1263 | 4 N.D. c | 0.3 ± 0.01 b | 0.38 ± 0.05 a | 0.0030101 | 3913-81-3 |
| 2-undecenal | 13.708 | 1363 | 1367 | N.D. c | 0.29 ± 0.04 b | 0.41 ± 0.05 a | 0.0041508 | 2463-77-6 |
| orcinol | 14.108 | 1421 | 1374 | 0.95 ± 0.16 a | 0.58 ± 0.10 b | 0.41 ± 0.06 b | 0.0074732 | 504-15-4 |
| ethyl-p-hydroxybenzoate | 15.254 | 1502 | 1438 | 1.92 ± 0.09 a | 1.23 ± 0.11 b | 1.17 ± 0.15 b | 0.0050822 | 2349-70-4 |
| (Z)-8-dodecenol | 16.104 | 1508 | 1468 | 0.77 ± 0.03 a | 0.46 ± 0.16 b | N.D. c | 0.0020082 | 40642-40-8 |
| 1-pentadecene | 16.461 | 1527 | 1515 | 15.93 ± 0.66 a | 15.56 ± 1.71 a | 15.63 ± 3.01 a | 0.1367729 | 13360-61-7 |
| 1-(2-hydroxy-4-methoxyphenyl)propan-1-one | 17.338 | 1653 | 1538 * | 0.96 ± 0.04 b | 0.75 ± 0.04 b | 1.33 ± 0.20 a | 0.0005612 | 6270-44-6 |
| cetene | 18.098 | 1612 | 1592 | 0.50 ± 0.03 b | 0.39 ± 0.01 c | 0.83 ± 0.08 a | 0.0000209 | 629-73-2 |
| (Z,Z)-1,8,11-heptadecatriene | 19.271 | 1620 | 1665 | 0.28 ± 0.04 a | 0.50 ± 0.04 b | 0.54 ± 0.01 b | 0.0014243 | 56134-03-3 |
| 1,13-tetradecadiene | 19.41 | 1709 | 1867 | 4.81 ± 0.46 b | 5.63 ± 0.41 b | 7.32 ± 0.40 a | 0.032606 | 21964-49-8 |
| 1-nonadecene | 19.756 | 1717 | 1892 | 5.87 ± 0.31 b | 5.86 ± 0.57 b | 7.14 ± 0.32 a | 0.7961594 | 18435-45-5 |
| n-hexadecanoic acid | 23.809 | 2016 | 1968 | 1.13 ± 0.05 a | 1.28 ± 0.18 a | 1.65 ± 0.28 a | 0.0041685 | 57-10-3 |
| (E)-9-octadecenoic acid | 26.506 | 2154 | 2139 | 0.81 ± 0.02 b | 1.62 ± 0.31 a | 0.77 ± 0.03 b | 0.0020028 | 112-79-8 |
| docosane | 31.904 | 2402 | 2200 | 0.16 ± 0.01 b | 0.46 ± 0.08 a | 0.43 ± 0.02 a | 0.015176 | 629-97-0 |
| tricosane | 32.981 | 2443 | 2300 | 0.90 ± 0.05 b | 0.85 ± 0.12 b | 1.68 ± 0.02 a | 0.00000754 | 638-67-5 |
| tetracosane | 33.358 | 2505 | 2400 | N.D. c | 0.27 ± 0.01 b | 0.44 ± 0.00 a | 0.0000532 | 646-31-1 |
| pentacosane | 33.496 | 2507 | 2500 | 0.37 ± 0.03 a | 0.33 ± 0.06 a | 0.47 ± 0.01 a | 0.0016095 | 629-99-2 |
| hexacosane | 33.869 | 2703 | 2600 | 1.45 ± 0.27 a | 1.68 ± 0.08 a | 1.58 ± 0.21 a | 0.014387 | 630-01-3 |
| heptacosane | 34.275 | 2711 | 2700 | 0.78 ± 0.11 a | 0.45 ± 0.04 b | 0.48 ± 0.06 b | 0.010739 | 593-49-7 |
| isopropyl tetracosyl ether | 34.458 | 2720 | 2724 | N.D. b | 0.37 ± 0.07 a | 0.33 ± 0.01 a | 0.039729 | 6 N.A. |
| 1-iodo-docosane | 34.675 | 2735 | 2730 | 1.57 ± 0.13 b | 1.38 ± 0.21 b | N.D. a | 0.0813 | 1000406-31-9 |
| 13-methylheptacosane | 35.376 | 2805 | 2731 | 0.22 ± 0.02 b | 0.42 ± 0.05 a | 0.45 ± 0.01 a | 0.2679168 | 15689-72-2 |
| 11-methylpentacosane | 35.487 | 2809 | 2734 | N.D. c | 7.12 ± 0.17 b | 13.03 ± 2.13 a | 0.0094031 | 15689-71-1 |
| 2-methylheptacosane | 35.561 | 2829 | 2762 | 1.40 ± 0.23 a | 1.70 ± 0.21 a | 1.87 ± 0.34 a | 0.0098973 | 1561-00-8 |
| octacosane | 35.756 | 2856 | 2800 | 3.12 ± 0.23 b | 3.11 ± 0.38 b | 7.36 ± 0.09 a | 0.00000444 | 630-02-4 |
| 2-methyloctacosane | 35.958 | 2870 | 2859 | N.D. c | 6.65 ± 0.65 b | 8.34 ± 0.25 a | 0.361 | 1560-88-9 |
| 15-methylnonacosane | 36.128 | 2804 | 2923 | 1.76 ± 0.08 b | 2.25 ± 0.28 a | 2.18 ± 0.14 ab | 0.0012749 | 65820-60-2 |
| 1-iodo-tetracosane | 36.818 | 2873 | 2942 | 5.01 ± 0.11 a | 3.75 ± 0.35 b | N.D. c | 0.104 | 1000406-32-0 |
| docosyl heptyl ether | 37.238 | 2910 | 2966 | N.D. a | 0.27 ± 0.05 b | 0.39 ± 0.02 a | 0.0064187 | N.A. |
| docosyl octyl ether | 37.983 | 3046 | 3056 | 1.2 ± 0.04 b | 1.50 ± 0.11 a | 1.23 ± 0.14 ab | 0.0005646 | N.A. |
| hentriacontane | 38.315 | 3003 | 3100 | 0.72 ± 0.13 b | 0.95 ± 0.15 b | 1.44 ± 0.04 a | 0.0000762 | 630-04-6 |
| 26-nor-5-cholesten-3-beta-ol-25-one | 38.540 | 3022 | 3131 | 1.89 ± 0.02 a | 1.90 ± 0.25 a | 1.33 ± 0.12 b | 0.0025098 | 7494-34-0 |
| 1-iodo-hexacosane | 40.449 | 3172 | 3147 | 1.99 ± 0.13 a | 1.79 ± 0.18 a | 1.05 ± 0.14 b | 0.0001359 | 52644-81-2 |
| cholesta-5,7-dien-3beta-ol | 40.958 | 3201 | 3158 | N.D. c | 0.19 ± 0.03 b | 0.29 ± 0.03 a | 0.0647143 | 434-16-2 |
| TC-S versus TC-W | p-Value | log2(FC) | TC-S versus TC-SR | p-Value | log2(FC) | TC-W versus TC-SR | p-Value | log2(FC) |
|---|---|---|---|---|---|---|---|---|
| octacosane | 0.0000309 | 1.5416 | octacosane | 0.00000236 | 1.5416 | cetene | 0.000143 | −0.6049 |
| 2-ethyl-2,5-cyclohexadiene-1,4-dione | 0.000264 | 0.90981 | ethyl-p-hydroxybenzoate | 0.00000355 | −0.93771 | docosyl octyl ether | 0.00042 | 0.79877 |
| cetene | 0.000599 | −0.674 | 1,13-tetradecadiene | 0.0000138 | 0.67309 | 1-iodo-hexacosane | 0.000583 | −1.248 |
| 1-(2-hydroxy-4-methoxyphenyl)propan-1-one | 0.000784 | −0.66323 | (Z)-8-dodecenol | 0.0000174 | −0.7814 | (Z,Z)-1,8,11-heptadecatriene | 0.000764 | 1.3119 |
| heptacosane | 0.001893 | −1.1068 | cetene | 0.0000687 | −0.674 | octacosane | 0.001025 | 0.51017 |
| (E)-9-octadecenoic acid | 0.002226 | 0.67888 | 1-nonadecene | 0.000585 | 0.5782 | (E)-9-octadecenoic acid | 0.001478 | 1.5648 |
| 1-iodo-tetracosane | 0.002793 | −0.71705 | 13-methylheptacosane | 0.001161 | 0.99779 | 2-ethyl-2,5-cyclohexadiene-1,4-dione | 0.002354 | 0.88627 |
| 11-methylpentacosane | 0.003041 | 1.1804 | orcinol | 0.001259 | −1.0116 | hexacosane | 0.002634 | 0.61333 |
| ethyl-p-hydroxybenzoate | 0.00307 | −0.93771 | 2-methyl-p-benzoquinone | 0.004376 | 0.78326 | 1,13-tetradecadiene | 0.003282 | 0.72677 |
| 1,13-tetradecadiene | 0.004294 | 0.67309 | heptacosane | 0.009924 | −1.1068 | 26-nor-5-cholesten-3-beta-ol-25-one | 0.007607 | 0.99856 |
| 1-nonadecene | 0.006906 | 0.5782 | (E)-9-octadecenoic acid | 0.016531 | 0.67888 | 1-iodo-docosane | 0.00889 | −1.6578 |
| 13-methylheptacosane | 0.007303 | 0.99779 | 1-(2-hydroxy-4-methoxyphenyl)propan-1-one | 0.018477 | −0.66323 | 2-methyl-p-benzoquinone | 0.010275 | 0.5094 |
| 2-methyloctacosane | 0.008263 | 0.62565 | hentriacontane | 0.019355 | 0.097868 | n-hexadecanoic acid | 0.01046 | 1.044 |
| orcinol | 0.009666 | −1.0116 | 2-ethyl-2,5-cyclohexadiene-1,4-dione | 0.023668 | 0.90981 | ethyl-p-hydroxybenzoate | 0.014055 | 0.5571 |
| orcinol | 0.020883 | 0.96149 | ||||||
| tricosane | 0.023572 | 0.5086 | ||||||
| isopropyl tetracosyl ether | 0.050881 | −0.68621 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Shan, C.; Liu, Q.; Li, B.; Liu, T. Comparative Analysis of the Metabolic Profiles of Strains of Tribolium castaneum (Herbst) Adults with Different Levels of Phosphine Resistance Based on Direct Immersion Solid-Phase Microextraction and Gas Chromatography-Mass Spectrometry. Molecules 2023, 28, 7721. https://doi.org/10.3390/molecules28237721
Li L, Shan C, Liu Q, Li B, Liu T. Comparative Analysis of the Metabolic Profiles of Strains of Tribolium castaneum (Herbst) Adults with Different Levels of Phosphine Resistance Based on Direct Immersion Solid-Phase Microextraction and Gas Chromatography-Mass Spectrometry. Molecules. 2023; 28(23):7721. https://doi.org/10.3390/molecules28237721
Chicago/Turabian StyleLi, Li, Changyao Shan, Qun Liu, Baishu Li, and Tao Liu. 2023. "Comparative Analysis of the Metabolic Profiles of Strains of Tribolium castaneum (Herbst) Adults with Different Levels of Phosphine Resistance Based on Direct Immersion Solid-Phase Microextraction and Gas Chromatography-Mass Spectrometry" Molecules 28, no. 23: 7721. https://doi.org/10.3390/molecules28237721
APA StyleLi, L., Shan, C., Liu, Q., Li, B., & Liu, T. (2023). Comparative Analysis of the Metabolic Profiles of Strains of Tribolium castaneum (Herbst) Adults with Different Levels of Phosphine Resistance Based on Direct Immersion Solid-Phase Microextraction and Gas Chromatography-Mass Spectrometry. Molecules, 28(23), 7721. https://doi.org/10.3390/molecules28237721