
Preparation of Herbal Extracts for Intestinal Immune Modulation Activity Based on In Vitro Screening and In Vivo Evaluation of Zingiber officinale Rosc. Extracts


Abstract
:1. Introduction
2. Results and Discussion
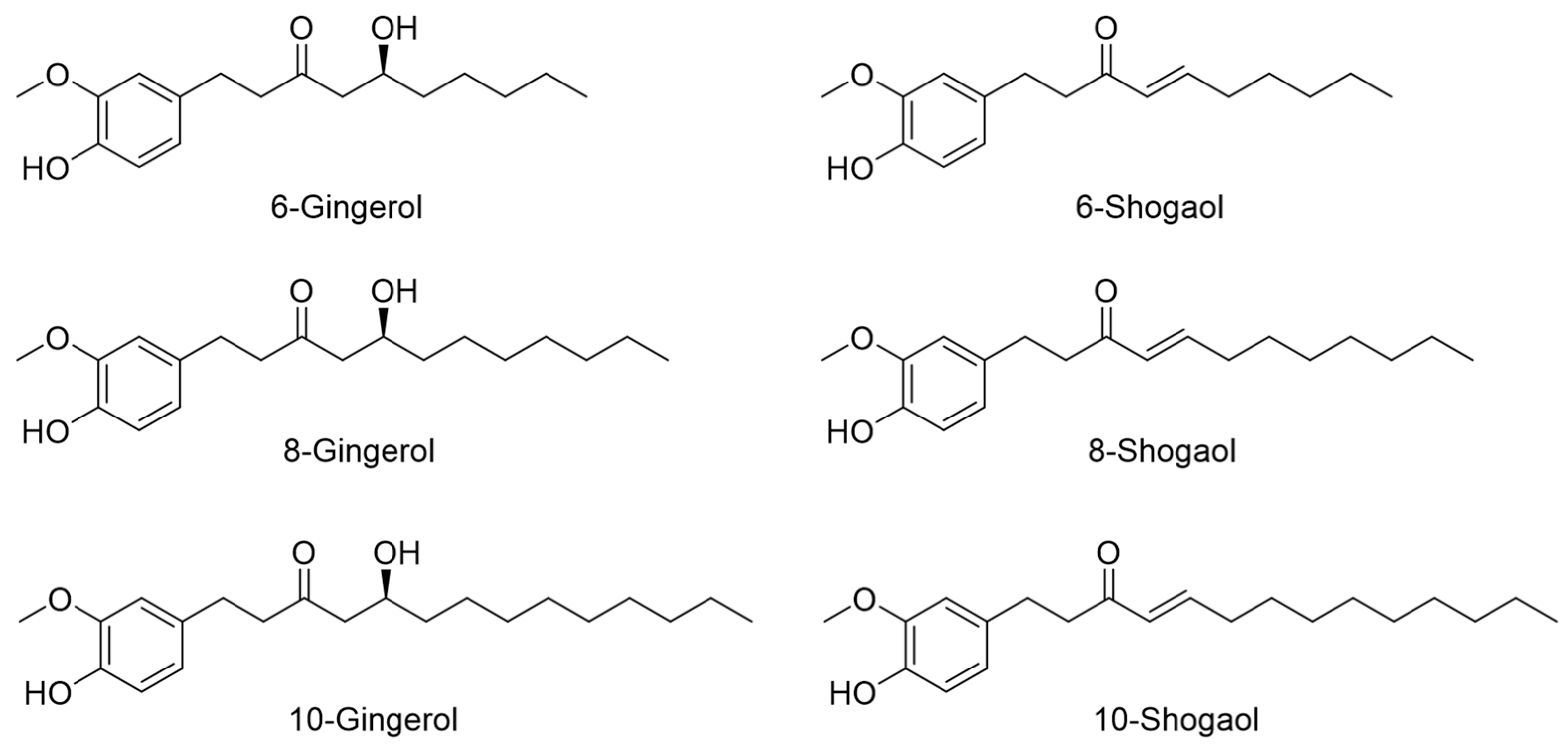
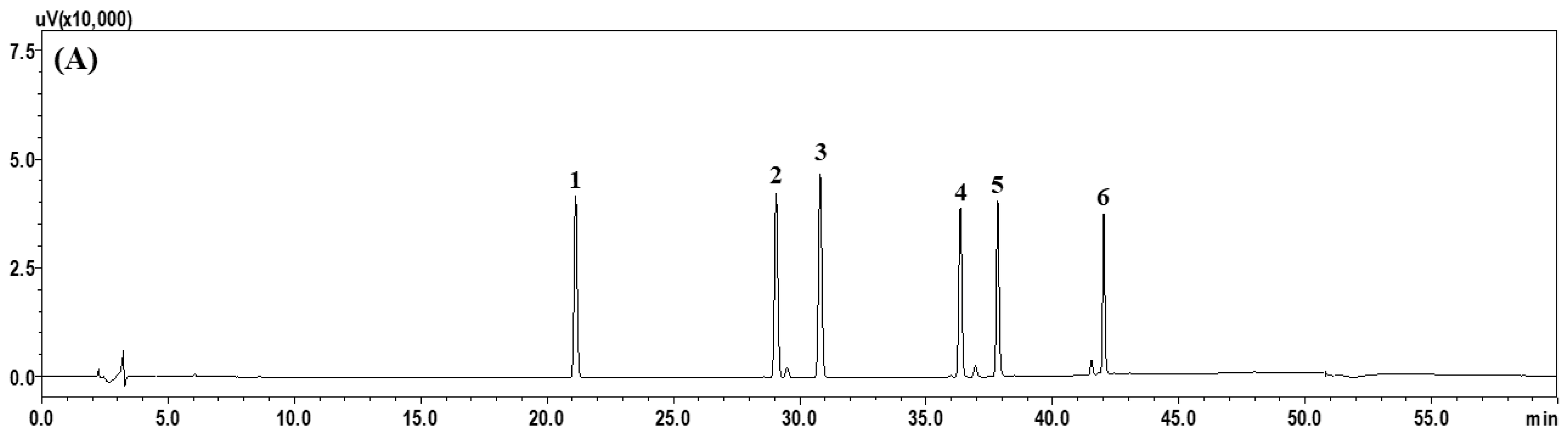
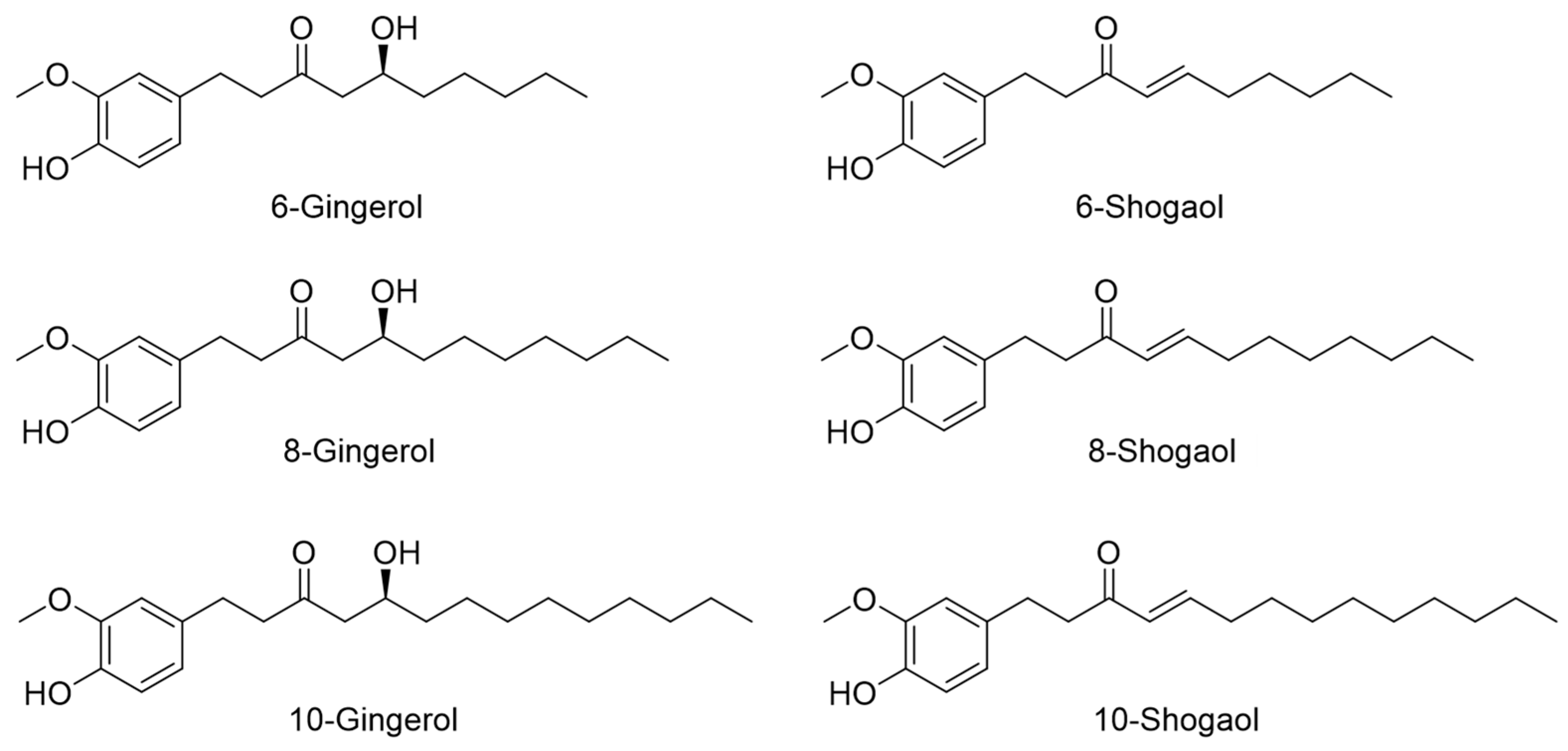
2.1. HPLC Profiling of Six Index Compounds of ZO Extract
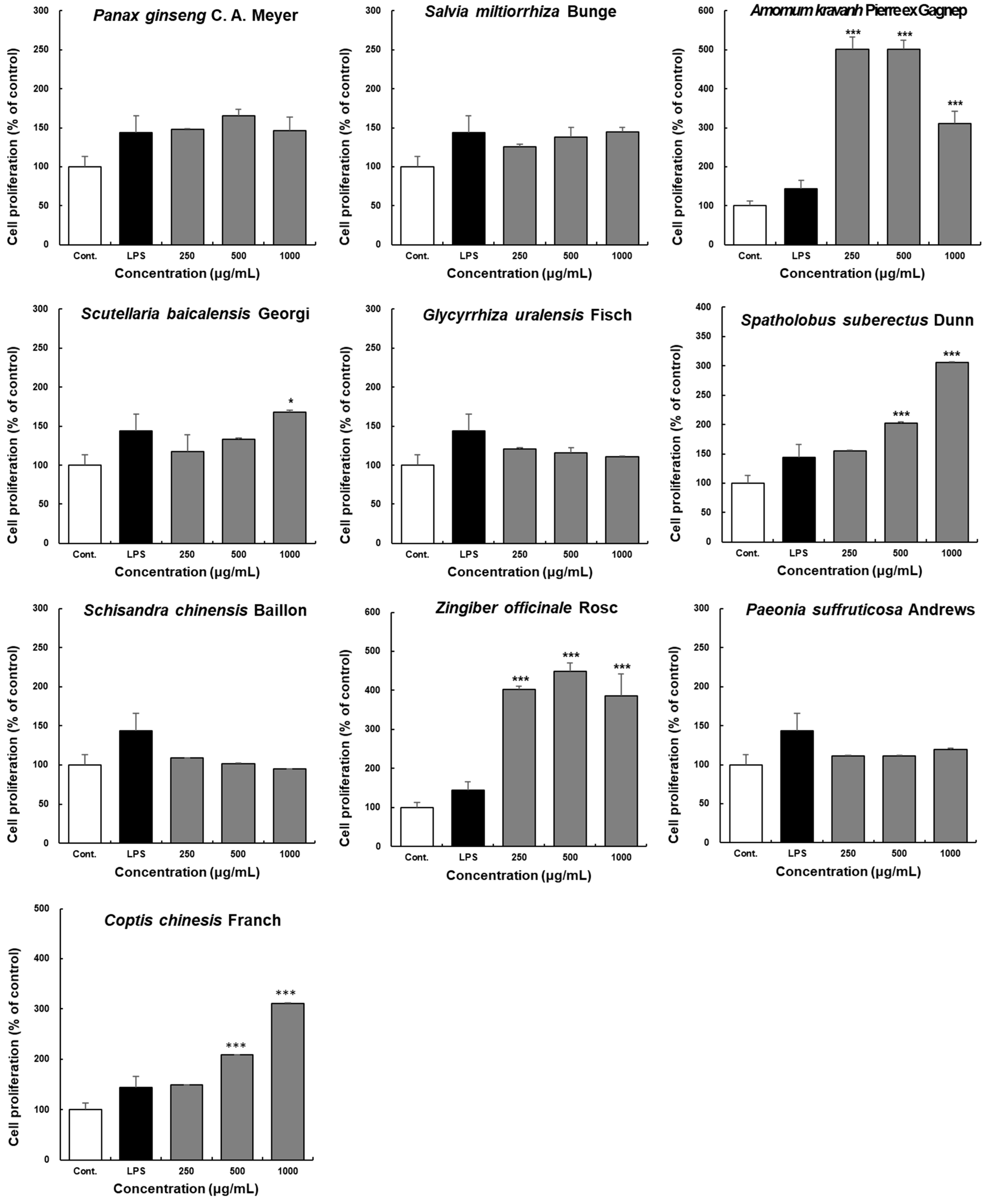
2.2. Effects of the Ten Herbal Medicine Extracts on Peyer’s Patch Cell Proliferation in Ex Vivo Experiments
2.3. Effects of Ten Herbal Medicine Extracts on IgA Production by Peyer’s Patch Cells Ex Vivo
2.4. Effects of the Ten Extracts on GM-CSF Production by Peyer’s Patch Cells in Ex Vivo Experiment
2.5. Determination of Peyer’s Patch Cell Proliferation and IgA Production Induced by Six Major Index Compounds Present in ZO Extract
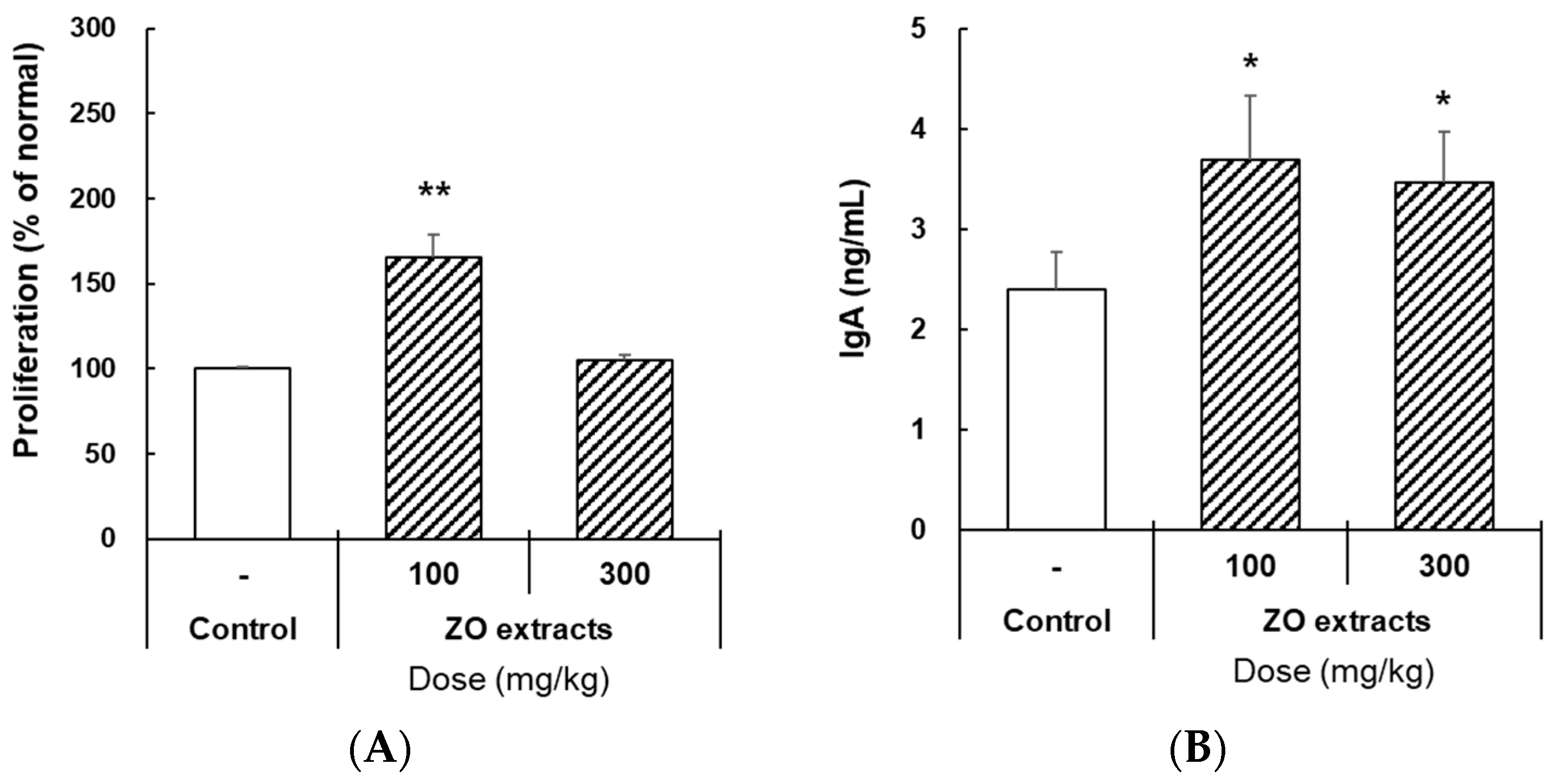
2.6. Effects of ZO Extract Administered Orally on Mouse IgA Production
3. Materials and Methods
3.1. Preparation of Crude Extract from Herbal Medicines
3.2. Chemicals and Reagents
3.3. HPLC Profiling of the Six Major Index Compounds in ZO Extract
3.4. Animals
3.5. Ex Vivo Proliferation of Peyer’s Patch Lymphocytes
3.6. Determination of IgA and GM-CSF Production in Peyer’s Patch Cells Ex Vivo
3.7. Determination of IgA Production in Peyer’s Patches In Vivo
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Janeway, C.A.; Traver, P. Immunobiology: The Immune System in Health and Disease, 1st ed.; Current Biology Ltd.; Garland Publishing Inc.: New York, NY, USA, 1994. [Google Scholar]
- Mowat, A.M. Anatomical basis of tolerance and immunity to intestinal antigens. Nat. Rev. Immunol. 2003, 3, 331–341. [Google Scholar] [CrossRef]
- Brandtzaeg, P. Mucosal immunity: Induction, dissemination, and effector functions. Scand. J. Immunol. 2009, 70, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Kiyono, H. The mucosal immune system of the respiratory tract. Curr. Opin. Virol. 2012, 2, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.; Hugot, J.P.; Barreau, F. Peyer’s patches: The immune sensors of the intestine. Int. J. Inflam. 2010, 2010, 823710. [Google Scholar] [CrossRef] [PubMed]
- Mantis, N.J.; Cheung, M.C.; Chintalacharuvu, K.R.; Rey, J.; Corthésy, B.; Neutra, M.R. Selective adherence of IgA to murine Peyer’s patch M cells: Evidence for a novel IgA receptor. J. Immunol. 2002, 169, 1844–1851. [Google Scholar] [CrossRef]
- Reboldi, A.; Arnon, T.I.; Rodda, L.B.; Atakilit, A.; Sheppard, D.; Cyster, J.G. IgA production requires B cell interaction with subepithelial dendritic cells in Peyer’s patches. Science 2016, 352, aaf4822. [Google Scholar] [CrossRef]
- Mosmann, T.R.; Coffman, R.L. TH1 and TH2 cells: Different patterns of lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 1989, 7, 145–173. [Google Scholar] [CrossRef]
- Ceuppens, J.L.; Baroja, M.L.; Lorre, K.; Van Damme, J.; Billiau, A. Human T cell activation with phytohemagglutinin. The function of IL-6 as an accessory signal. J. Immunol. 1988, 141, 3868–3874. [Google Scholar] [CrossRef]
- Li, Y.; Jin, L.; Chen, T. The effects of secretory IgA in the mucosal immune system. BioMed Res. Int. 2020, 2020, 2032057. [Google Scholar] [CrossRef]
- Yang, Y. Chinese Herbal Medicines: Comparisons and Characteristics, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2009; ISBN 9780702047817. [Google Scholar]
- Liu, C.; Tseng, A.; Yang, S. Chinese Herbal Medicine Modern Applications of Traditional Formulas; CRC Press: Boca Raton, FL, USA, 2004; ISBN 9780849315688. [Google Scholar]
- Kim, J.H.; Do, E.J.; Lee, G. Investigation of anti-microbial activity of herbal medicines used as natural preservatives based on the analysis of papers and patents. J. Physiol. Pathol. Korean Med. 2015, 29, 101–113. [Google Scholar] [CrossRef]
- Kang, B.H.; Park, D.H.; Lee, M.J.; Jeon, C.Y.; Kang, K.S.; Choi, Y.K. Beneficial effect of paeonol on antibiotic-associated inflammatory response in mice with diarrhea. Biomolecules 2022, 12, 1634. [Google Scholar] [CrossRef]
- Ha, H.; Kim, Y.; Kweon, K.; Kim, J. Review of the domestic research trends in the study of Korean herbal medicine with anti-inflammation effects. Kor. J. Herbol. 2011, 26, 15–22. [Google Scholar]
- Yamaguchi, M.; Yahagi, N.; Kato, H.; Takano, F.; Ohta, T. Capsicum extract and its constituents modulated the production of immunoglobulins in Peyer’s patch cells ex vivo. J. Funct. Foods 2010, 2, 255–262. [Google Scholar] [CrossRef]
- Ota, N.; Takano, F.; Muroga, S.; Kawabata, T.; Ishigaki, Y.; Yahagi, N.; Ohta, T. Garlic extract and its selected organosulphur constituents promote ileal immune responses ex vivo. J. Funct. Foods 2012, 4, 243–252. [Google Scholar] [CrossRef]
- Park, D.H.; Han, B.; Shin, M.S.; Hwang, G.S. Enhanced intestinal immune response in mice after oral administration of Korea red ginseng-derived polysaccharide. Polymers 2020, 12, 2186. [Google Scholar] [CrossRef] [PubMed]
- Suh, H.J.; Yang, H.S.; Ra, K.S.; Noh, D.O.; Kwon, K.H.; Hwang, J.H.; Yu, K.W. Peyer’s patch-mediated intestinal immune system modulating activity of pectic-type polysaccharide from peel of Citrus unshiu. Food Chem. 2013, 138, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Chaiyama, V.; Keawsompong, S.; LeBlanc, J.G.; de Moreno de LeBlanc, A.D.M.; Chatel, J.M.; Chanput, W. Action modes of the immune modulating activities of crude mushroom polysaccharide from Phallus atrovolvatus. Bioact. Carbohydr. Diet. Fibre 2020, 23, 100216. [Google Scholar] [CrossRef]
- Kim, S.H.; Jeung, W.; Choi, I.D.; Jeong, J.W.; Lee, D.E.; Huh, C.S.; Kim, G.B.; Hong, S.S.; Shim, J.J.; Lee, J.L.; et al. Lactic acid bacteria improves Peyer’s patch cell-mediated immunoglobulin A and tight-junction expression in a destructed gut microbial environment. J. Microbiol. Biotechnol. 2016, 26, 1035–1045. [Google Scholar] [CrossRef]
- Shin, M.S.; Yu, K.W.; Shin, K.S.; Lee, H. Enhancement of immunological activity in mice with oral administration of cell wall components of Bifidobacterium bifidum. Food Sci. Biotechnol. 2004, 13, 85–89. [Google Scholar]
- Reboldi, A.; Cyster, J.G. Peyer’s patches: Organizing B-cell responses at the intestinal frontier. Immunol. Rev. 2016, 271, 230–245. [Google Scholar] [CrossRef]
- Rey, J.; Garin, N.; Spertini, F.; Corthésy, B. Targeting of secretory IgA to Peyer’s patch dendritic and T cells after transport by intestinal M cells. J. Immunol. 2004, 172, 3026–3033. [Google Scholar] [CrossRef] [PubMed]
- Gurung, P.; Lim, J.; Thapa Magar, T.B.; Shrestha, R.; Kim, Y.W. Modulation of local and systemic immune responses by fermented garlic extract. Int. J. Funct. Nutr. 2022, 3, 6. [Google Scholar] [CrossRef]
- Hong, T.; Matsumoto, T.; Kiyohara, H.; Yamada, H. Enhanced production of hematopoietic growth factors through T cell activation in Peyer’s patches by oral administration of Kampo (Japanese herbal) medicine, “Juzen-Taiho-To”. Phytomedicine 1998, 5, 353–360. [Google Scholar] [CrossRef]
- Takano, F.; Yamaguchi, M.; Takada, S.; Shoda, S.; Yahagi, N.; Takahashi, T.; Ohta, T. Capsicum ethanol extracts and capsaicin enhance interleukin-2 and interferon-gamma production in cultured murine Peyer’s patch cells ex vivo. Life Sci. 2007, 80, 1553–1563. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Liu, C.H.; Roberts, A.I.; Das, J.; Xu, G.; Ren, G.; Zhang, Y.; Zhang, L.; Yuan, Z.R.; Tan, H.S.; et al. Granulocyte-macrophage colony-stimulating factor (GM-CSF) and T-cell responses: What we do and don’t know. Cell Res. 2006, 16, 126–133. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Amount (mg/g) | ||
|---|---|---|---|
| Mean | SD | RSD (%) | |
| 6-Gingerol | 6.64 | 0.02 | 0.36 |
| 8-Gingerol | 0.22 | 0.00 | 1.62 |
| 6-Shogaol | 0.28 | 0.00 | 0.42 |
| 10-Gingerol | 0.16 | 0.00 | 0.52 |
| 8-Shogaol | 0.01 | 0.00 | 1.84 |
| 10-Shogaol | <LOQ * | - | - |
| Viability (%) | Control | 10 μM | 25 μM | 50 μM | ||||
|---|---|---|---|---|---|---|---|---|
| Average | SD | Average | SD | Average | SD | Average | SD | |
| Control | 100.0 | 1.2 | - | - | - | - | - | - |
| LPS | 220.5 *** | 8.4 | - | - | - | - | - | - |
| 6-Gingerol | - | - | 95.3 | 1.7 | 93.8 | 0.4 | 92.1 | 0.7 |
| 8-Gingerol | - | - | 96.6 | 1.2 | 93.6 | 1.0 | 90.9 | 0.7 |
| 10-Gingerol | - | - | 96.8 | 0.2 | 93.9 | 1.3 | 93.2 | 2.4 |
| 6-Shogaol | - | - | 94.2 | 1.2 | 88.4 | 5.2 | 94.9 | 2.1 |
| 8-Shogaol | - | - | 94.0 | 2.4 | 92.7 | 2.6 | 91.4 | 2.0 |
| 10-Shogaol | - | - | 96.9 | 0.4 | 94.4 | 1.8 | 95.7 | 2.4 |
| IgA (pg/mL) | Control | 10 μM | 25 μM | 50 μM | ||||
| IgA | SD | IgA | SD | IgA | SD | IgA | SD | |
| Control | 3246.3 | 19.9 | - | - | - | - | - | - |
| LPS | 5260.4 *** | 57.8 | - | - | - | - | - | - |
| 6-Gingerol | - | - | 2951.9 | 142.5 | 2868.0 | 227.4 | 2995.1 | 54.3 |
| 8-Gingerol | - | - | 2052.1 | 17.0 | 2481.6 | 156.1 | 2654.4 | 74.7 |
| 10-Gingerol | - | - | 1579.4 | 156.1 | 2606.4 | 203.6 | 2767.2 | 234.2 |
| 6-Shogaol | - | - | 1452.2 | 193.4 | 1672.9 | 84.8 | 1812.1 | 125.6 |
| 8-Shogaol | - | - | 1622.5 | 54.3 | 2548.8 | 95.0 | 2724.0 | 254.5 |
| 10-Shogaol | - | - | 1900.9 | 54.3 | 2064.1 | 142.5 | 2896.8 | 57.7 |
| Number | Herbal Medicine | Origin | Freeze-Dried Extract (g) | Yield (%) |
|---|---|---|---|---|
| 1 | Panax ginseng C. A. Meyer | Republic of Korea | 8.3 | 27.66 |
| 2 | Salvia miltiorrhiza Bunge | China | 12.2 | 40.6 |
| 3 | Amomum kravanh Pierre ex Gagnep. | Indonesia | 9.9 | 33.0 |
| 4 | Scutellaria baicalensis Georgi | China | 1.5 | 5.0 |
| 5 | Glycyrrhiza uralensis Fisch. | China | 11.8 | 39.3 |
| 6 | Spatholobus suberectus Dunn | Vietnam | 9.1 | 30.3 |
| 7 | Schisandra chinensis Baillon | China | 11.7 | 39.0 |
| 8 | Zingiber officinale Rosc. | Peru | 10.3 | 34.3 |
| 9 | Paeonia suffruticosa Andrews | China | 6.6 | 22.0 |
| 10 | Coptis chinesis Franch. | China | 10.2 | 34.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Min, S.J.; Kim, S.J.; Park, J.Y.; Seo, C.-S.; Choi, Y.-K. Preparation of Herbal Extracts for Intestinal Immune Modulation Activity Based on In Vitro Screening and In Vivo Evaluation of Zingiber officinale Rosc. Extracts. Molecules 2023, 28, 6743. https://doi.org/10.3390/molecules28186743
Min SJ, Kim SJ, Park JY, Seo C-S, Choi Y-K. Preparation of Herbal Extracts for Intestinal Immune Modulation Activity Based on In Vitro Screening and In Vivo Evaluation of Zingiber officinale Rosc. Extracts. Molecules. 2023; 28(18):6743. https://doi.org/10.3390/molecules28186743
Chicago/Turabian StyleMin, Su Ji, Sung Jin Kim, Jun Yeon Park, Chang-Seob Seo, and You-Kyong Choi. 2023. "Preparation of Herbal Extracts for Intestinal Immune Modulation Activity Based on In Vitro Screening and In Vivo Evaluation of Zingiber officinale Rosc. Extracts" Molecules 28, no. 18: 6743. https://doi.org/10.3390/molecules28186743
APA StyleMin, S. J., Kim, S. J., Park, J. Y., Seo, C.-S., & Choi, Y.-K. (2023). Preparation of Herbal Extracts for Intestinal Immune Modulation Activity Based on In Vitro Screening and In Vivo Evaluation of Zingiber officinale Rosc. Extracts. Molecules, 28(18), 6743. https://doi.org/10.3390/molecules28186743