
Effects of Plant Elicitors on Growth and Gypenosides Biosynthesis in Cell Culture of Giao co lam (Gynostemma pentaphyllum)

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Determinaton of Rb1 in Suspension Cell
2.2. Effect of Elicitor Concentrations
2.3. Effect of Elicitation Time
3. Discussion
4. Materials and Methods
4.1. Cell Suspension Cultivation
4.2. Elicitation
4.3. Gypenosides Quantification
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Sample Availability
References
- Cui, W.Y.; Jin, Y.; Liu, H.; Zu, M.L.; Zhai, X.F.; Yang, C.; Gu, Y.L.; Cheng, Y.; Piao, X.L. Dammarane-type saponins from Gynostemma pentaphyllum and their cytotoxicities. Nat. Prod. Res. 2021, 35, 4433–4441. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.Y.; Chang, C.C.; Huang, H.C.; Zhang, L.J.; Liaw, C.C.; Lin, Y.C.; Nguyen, N.L.; Vo, T.H.; Cheng, Y.Y.; Morris-Natschke, S.L.; et al. New Dammarane-type Saponins from Gynostemma pentaphyllum. Molecules 2019, 24, 1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, T.; Zou, L.; Sun, S.; Kuang, X.; Wei, J.; Wang, L.; Li, Y.; Sun, C. Hybrid sequencing of the Gynostemma pentaphyllum transcriptome provides new insights into gypenoside biosynthesis. BMC Genom. 2019, 20, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Balan, P.; Popovich, D.G. Chapter 6—Comparison of the ginsenoside composition of Asian ginseng (Panax ginseng) and American ginseng (Panax quinquefolius L.) and their transformation pathways. In Studies in Natural Products Chemistry; Atta Ur, R., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 161–195. [Google Scholar]
- Ahmed, T.; Raza, S.H.; Maryam, A.; Setzer, W.N.; Braidy, N.; Nabavi, S.F.; de Oliveira, M.R.; Nabavi, S.M. Ginsenoside Rb1 as a neuroprotective agent: A review. Brain Res. Bull. 2016, 125, 30–43. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Xie, W.; Sun, Y.; Dai, Z.; Li, G.; Sun, G.; Sun, X. Ginsenoside Rb1 and mitochondria: A short review of the literature. Mol. Cell. Probes 2019, 43, 1–5. [Google Scholar] [CrossRef]
- Shi, Y.H.; Li, Y.; Wang, Y.; Xu, Z.; Fu, H.; Zheng, G.Q. Ginsenoside-Rb1 for ischemic stroke: A systematic review and meta-analysis of preclinical evidence and possible mechanisms. Front. Pharmacol. 2020, 11, 285. [Google Scholar] [CrossRef]
- Huang, P.; Xia, L.; Zhou, L.; Liu, W.; Wang, P.; Qing, Z.; Zeng, J. Influence of different elicitors on BIA production in Macleaya cordata. Sci. Rep. 2021, 11, 619. [Google Scholar] [CrossRef]
- Luthra, R.; Roy, A.; Pandit, S.; Prasad, R. Biotechnological methods for the production of ginsenosides. S. Afr. J. Bot. 2021, 141, 25–36. [Google Scholar] [CrossRef]
- Davey, M. Secondary Metabolism in Plant Cell Cultures. In Encyclopedia of Applied Plant Sciences, 2nd ed.; Thomas, B., Murray, B.G., Murphy, D.J., Eds.; Academic Press: Oxford, UK, 2017; pp. 462–467. [Google Scholar]
- Farag, M.A.; Al-Mahdy, D.A.; Meyer, A.; Westphal, H.; Wessjohann, L.A. Metabolomics reveals biotic and abiotic elicitor effects on the soft coral Sarcophyton ehrenbergi terpenoid content. Sci. Rep. 2017, 7, 648. [Google Scholar] [CrossRef] [Green Version]
- Nhan, N.H.; Loc, N.H. Enhancement of eurycomanone biosynthesis in cell culture of longjack (Eurycoma longifolia) by elicitor treatment. J. Plant Biotechnol. 2018, 45, 340–346. [Google Scholar] [CrossRef] [Green Version]
- Loc, N.H.; Anh, N.H.T.; Khuyen, L.T.; An, T.N.T. Effects of yeast extract and methyl jasmonate on the enhancement of solasodine biosynthesis in cell cultures of Solanum hainanense Hance. J. BioScience Biotechnol. 2014, 3, 1–6. [Google Scholar]
- Kochan, E.; Szymczyk, P.; Kuźma, Ł.; Lipert, A.; Szymańska, G. Yeast Extract Stimulates Ginsenoside Production in Hairy Root Cultures of American Ginseng Cultivated in Shake Flasks and Nutrient Sprinkle Bioreactors. Molecules 2017, 22, 880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Khayri, J.M.; Naik, P.M. Elicitor-Induced Production of Biomass and Pharmaceutical Phenolic Compounds in Cell Suspension Culture of Date Palm (Phoenix dactylifera L.). Molecules 2020, 25, 4669. [Google Scholar] [CrossRef] [PubMed]
- Loc, N.H.; Giang, N.T.; Huy, N.D.; Lan, T.T.P. Accumulation of madecassoside—A major component of centelloside—In centella (Centella asiatica (L.) Urban) cells elicited by salicylic acid. Period. Biol. 2017, 119, 81–85. [Google Scholar]
- Azeez, H.; Ibrahim, K. Effect of Biotic Elicitors on Secondary Metabolite Production in Cell Suspensions of Hypericum triquetrifolium Turra. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca Hortic. 2013, 70, 26–33. [Google Scholar]
- Endah, R.; Beyene, G.; Kiggundu, A.; van den Berg, N.; Schlüter, U.; Kunert, K.; Chikwamba, R. Elicitor and Fusarium-induced expression of NPR1-like genes in banana. Plant Physiol. Biochem. 2008, 46, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Anh, T.T.N.; Thi, P.T.D.; Lan, T.T.; Dung, T.Q.; Vu, N.Q.H.; Phuong, T.T.B.; Quang, H.T. Affect of culture conditions on growth and accumulation of saponin Rb1 in Gynostemma pentaphyllum (Thunb.) Makino suspension cells. Proc. Vietnam. Natl. Conf. Biotechnol. 2020, 885–890. (In Vietnamese) [Google Scholar]
- Quang, H.T.; Thi, P.T.D.; Lan, T.T.; Thuy, P.M.T.; Hoang, V.D.; Kim, P.T.A. Effects of culture medium on callus growth of Gynostemma pentaphyllum (Thunb.) Makino—A valuable medicinal plant. Hue Univ. J. Sci. Nat. Sci. 2021, 130, 127–137. (In Vietnamese) [Google Scholar]
- Namdeo, A.G. Plant cell elicitation for production of secondary metabolites: A review. Pharmacogn. Rev. 2007, 1, 69–79. [Google Scholar]
- Naik, P.M.; Al-Khayri, J.M. Abiotic and Biotic Elicitors–Role in Secondary Metabolites Production through In Vitro Culture of Medicinal Plants. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives; Shanker, A.K., Shanker, C., Eds.; IntechOpen: London, UK, 2016. [Google Scholar]
- Patel, Z.M.; Mahapatra, R.; Jampala, S.S.M. Chapter 11—Role of fungal elicitors in plant defense mechanism. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Sharma, V., Salwan, R., Al-Ani, L.K.T., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 143–158. [Google Scholar]
- Vijendra, P.D.; Jayanna, S.G.; Kumar, V.; Sannabommaji, T.; Rajashekar, J.; Gajula, H. Product enhancement of triterpenoid saponins in cell suspension cultures of Leucas aspera Spreng. Ind. Crops Prod. 2020, 156, 112857. [Google Scholar] [CrossRef]
- Raghavendra, S.; Ramesh, C.K.; Kumar, V.; Moinuddin Khan, M.H. Elicitors and precursor induced effect on L-Dopa production in suspension cultures of Mucuna pruriens L. Front. Life Sci. 2011, 5, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Sliwińska, A.; Syklowska, K.; Kosmider, A.; Granica, S.; Miszczak, K.; Nowicka, G.; Kasztelan, A.; Podsadni, P.; Turlo, J. Stimulation of phenolic compounds production in the in vitro cultivated Polyscias filicifolia Bailey shoots and evaluation of the antioxidant and cytotoxic potential of plant extracts. Acta Soc. Bot. Pol. 2018, 87, 289–316. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, Q.; Huang, Y.; Zhao, R.; Sun, J.; Yuan, X.; Xu, H.; Liu, H.; Wu, Y. Gene excavation and expression analysis of CYP and UGT related to the post modifying stage of gypenoside biosynthesis in Gynostemma pentaphyllum (Thunb.) Makino by comprehensive analysis of RNA and proteome sequencing. PLoS ONE 2021, 16, e0260027. [Google Scholar] [CrossRef] [PubMed]
- Krzyzanowska, J.; Czubacka, A.; Pecio, L.; Przybys, M.; Doroszewska, T.; Stochmal, A.; Oleszek, W. The effects of jasmonic acid and methyl jasmonate on rosmarinic acid production in Mentha × piperita cell suspension cultures. Plant Cell Tissue Organ Cult. 2012, 108, 73–81. [Google Scholar] [CrossRef]
- Cai, Z.; Kastell, A.; Smetanska, I. Chitosan or yeast extract enhance the accumulation of eight phenolic acids in cell suspension cultures of Malus × domestica Borkh. J. Hortic. Sci. Biotechnol. 2014, 89, 93–99. [Google Scholar] [CrossRef]
- Chandran, H.; Meena, M.; Barupal, T.; Sharma, K. Plant tissue culture as a perpetual source for production of industrially important bioactive compounds. Biotechnol. Rep. 2020, 26, e00450. [Google Scholar] [CrossRef]
- Tung, N.T.; Thi, P.T.D.; Sang, D.N.; Thao, D.T.; Quang, H.T. Biomass accumulation of Gynostemma pentaphyllum (Thunb.) Makino in cell suspension cultures inhibiting human cancer cell growth. Res. J. Biotechnol. 2022, 17, 61–68. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar]
- Huy, N.D.; Ha, D.T.T.; Khoo, K.S.; Lan, P.T.N.; Quang, H.T.; Loc, N.H.; Park, S.-M.; Veeramuthu, A.; Show, P.L. Synthetic dyes removal by Fusarium oxysporum HUIB02 and stimulation effect on laccase accumulation. Environ. Technol. Innov. 2020, 19, 101027. [Google Scholar] [CrossRef]
- Wu, Q.; Jang, M.; Piao, X.L. Determination by UPLC-MS of four dammarane-type saponins from heat-processed Gynostemma pentaphyllum. Biosci. Biotechnol. Biochem. 2014, 78, 311–316. [Google Scholar] [CrossRef]
- 35 Goel, N.G.; Sirohi, S.K.; Dwivedi, J. Estimation of total saponins and evaluate their effect on in vitro methanogenesis and rumen fermentation pattern in wheat straw based diet. J. Adv. Vet. Res. 2012, 2, 120–126. [Google Scholar]
- Hiai, S.; Oura, H.; Nakajima, T. Color reaction of some sapogenins and saponins with vanillin and sulfuric acid. Planta Med. 1976, 29, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ren, D.; Guo, D.A.; Pan, Y.; Zhang, H.; Hu, P. Method Development for Gypenosides Fingerprint by High Performance Liquid Chromatography with Diode-Array Detection and the Addition of Internal Standard. Chem. Pharm. Bull. 2008, 56, 389–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
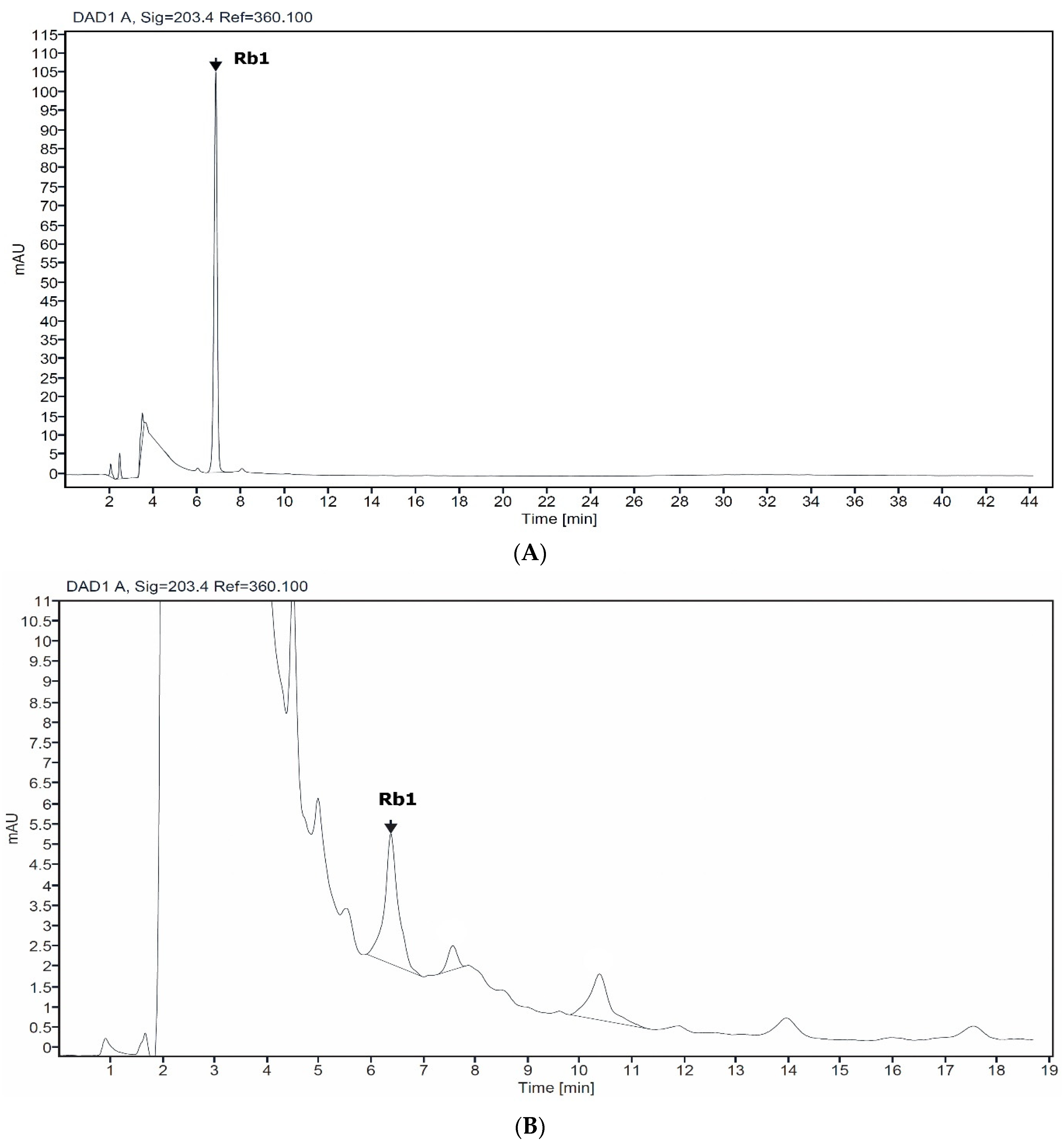


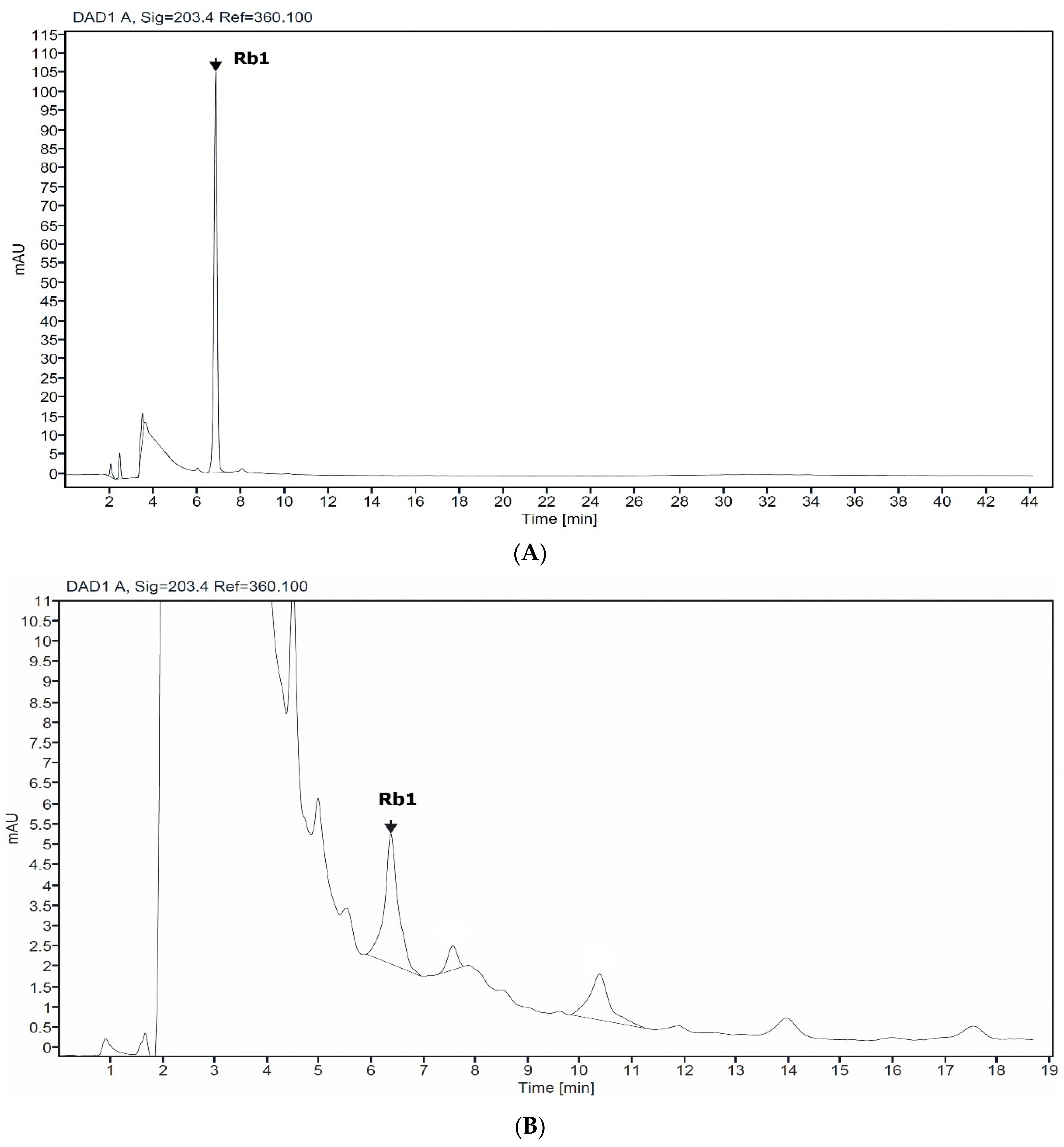{kind=link}
{kind=link}
| Concentrations (mM) | Elicitors | Fresh Weight (g) | Dry Weight (g) | Gypenosides (mg/g Dry Weight) | Rb1 (mg/g Dry Weight) |
|---|---|---|---|---|---|
| 0 | - | 5.732 a | 0.190 a | 50.545 d | 0.048 cde |
| 25 | MeJA | 5.043 bcd | 0.165 bc | 54.546 cd | 0.052 bcd |
| SA | 5.763 a | 0.204 a | 58.475 bc | 0.060 bc | |
| 50 | MeJA | 5.003 bcd | 0.163 bc | 58.116 bcd | 0.053 bcd |
| SA | 5.456 ab | 0.163 bc | 63.980 b | 0.062 b | |
| 100 | MeJA | 4.876 bcde | 0.094 de | 53.617 cd | 0.041 de |
| SA | 5.187 abc | 0.141 c | 76.809 a | 0.075 a | |
| 150 | MeJA | 4.854 bcde | 0.088 de | 32.552 e | 0.037 f |
| SA | 4.659 cde | 0.107 d | 32.100 e | 0.061 b | |
| 200 | MeJA | 4.472 de | 0.063 e | 29.514 e | 0.037 f |
| SA | 4.331 e | 0.026 f | 9.724 f | 0.056 bc |
| Concentrations (g/L) | Elicitors | Fresh Weight (g) | Dry Weight (g) | Gypenosides (mg/g Dry Weight) | Rb1 (mg/g Dry Weight) |
|---|---|---|---|---|---|
| 0 | - | 5.433 a | 0.171 abc | 50.478 de | 0.052 b |
| 1 | YE | 5.499 a | 0.170 abc | 54.974 d | 0.061 a |
| Fox | 4.597 bc | 0.181 a | 36.790 f | 0.049 bc | |
| 2 | YE | 5.411 a | 0.131 de | 67.135 b | 0.063 a |
| Fox | 4.885 ab | 0.174 ab | 34.924 f | 0.050 bc | |
| 3 | YE | 5.410 a | 0.137 de | 73.767 a | 0.066 a |
| Fox | 4.521 bc | 0.170 abc | 31.115 fg | 0.043 cd | |
| 4 | YE | 5.424 a | 0.123 ef | 60.915 c | 0.063 a |
| Fox | 4.076 cd | 0.165 abcd | 27.411 gh | 0.040 d | |
| 5 | YE | 5.387 a | 0.105 f | 46.338 e | 0.062 a |
| Fox | 3.931 d | 0.142 bcde | 23.066 h | 0.039 d |
| Treatment Days | Elicitors | Fresh Weight (g) | Dry Weight (g) | Total Gypenosides (mg/g Dry Weight) | Rb1 (mg/g Dry Weight) |
|---|---|---|---|---|---|
| 0 | MeJA | 4.968 cde | 0.207 fg | 57.852 hi | 0.052 g |
| SA | 4.908 cde | 0.148 l | 76.480 ab | 0.075 c | |
| YE | 4.855 e | 0.186 i | 74.579 bc | 0.061 f | |
| 3 | MeJA | 5.110 bcde | 0.212 ef | 65.633 ef | 0.055 ghi |
| SA | 4.906 de | 0.149 l | 76.661 ab | 0.088 b | |
| YE | 4.980 cde | 0.192 h | 61.202 gh | 0.056 gh | |
| 6 | MeJA | 5.168 bcde | 0.216 de | 73.461 bc | 0.055 ghi |
| SA | 4.907 bde | 0.157 k | 79.721 a | 0.093 a | |
| YE | 5.041 bcde | 0.204 g | 57.543 hi | 0.035 k | |
| 9 | MeJA | 5.302 bc | 0.244 b | 73.901 bc | 0.066 de |
| SA | 5.025 cde | 0.178 j | 73.558 bc | 0.063 ef | |
| YE | 5.137 bcde | 0.209 f | 54.163 ij | 0.031 ij | |
| 12 | MeJA | 5.254 bcd | 0.229 c | 68.622 de | 0.067 d |
| SA | 5.259 bcd | 0.211 f | 70.547 cd | 0.059 fg | |
| YE | 5.424 b | 0.221 d | 51.321 j | 0.027 jk | |
| 15 | MeJA | 5.018 cde | 0.191 hi | 60.814 gh | 0.045 j |
| SA | 5.327 bc | 0.221 d | 62.628 fg | 0.054 ghi | |
| YE | 6.048 a | 0.252 a | 49.989 j | 0.024 k |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quang, H.T.; Thi, P.T.D.; Sang, D.N.; Tram, T.T.N.; Huy, N.D.; Dung, T.Q.; The, Q.T.T. Effects of Plant Elicitors on Growth and Gypenosides Biosynthesis in Cell Culture of Giao co lam (Gynostemma pentaphyllum). Molecules 2022, 27, 2972. https://doi.org/10.3390/molecules27092972
Quang HT, Thi PTD, Sang DN, Tram TTN, Huy ND, Dung TQ, The QTT. Effects of Plant Elicitors on Growth and Gypenosides Biosynthesis in Cell Culture of Giao co lam (Gynostemma pentaphyllum). Molecules. 2022; 27(9):2972. https://doi.org/10.3390/molecules27092972
Chicago/Turabian StyleQuang, Hoang Tan, Pham Thi Diem Thi, Dang Ngoc Sang, Tran Thi Ngoc Tram, Nguyen Duc Huy, Tran Quoc Dung, and Quach Thi Thu The. 2022. "Effects of Plant Elicitors on Growth and Gypenosides Biosynthesis in Cell Culture of Giao co lam (Gynostemma pentaphyllum)" Molecules 27, no. 9: 2972. https://doi.org/10.3390/molecules27092972
APA StyleQuang, H. T., Thi, P. T. D., Sang, D. N., Tram, T. T. N., Huy, N. D., Dung, T. Q., & The, Q. T. T. (2022). Effects of Plant Elicitors on Growth and Gypenosides Biosynthesis in Cell Culture of Giao co lam (Gynostemma pentaphyllum). Molecules, 27(9), 2972. https://doi.org/10.3390/molecules27092972