
Knockout of KLF10 Ameliorated Diabetic Renal Fibrosis via Downregulation of DKK-1

Abstract
:1. Introduction
2. Results
2.1. Biochemical Properties of Experimental Animals
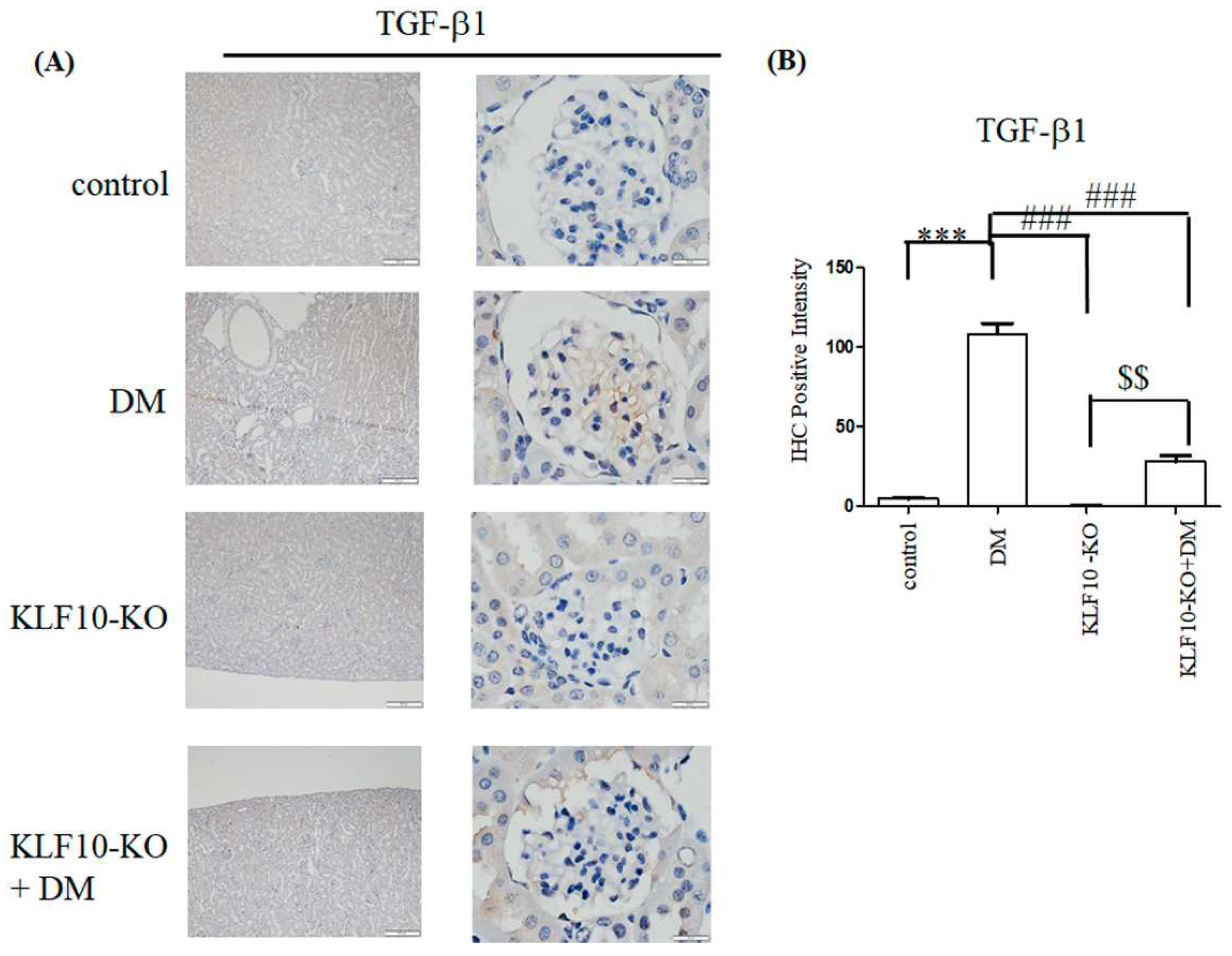
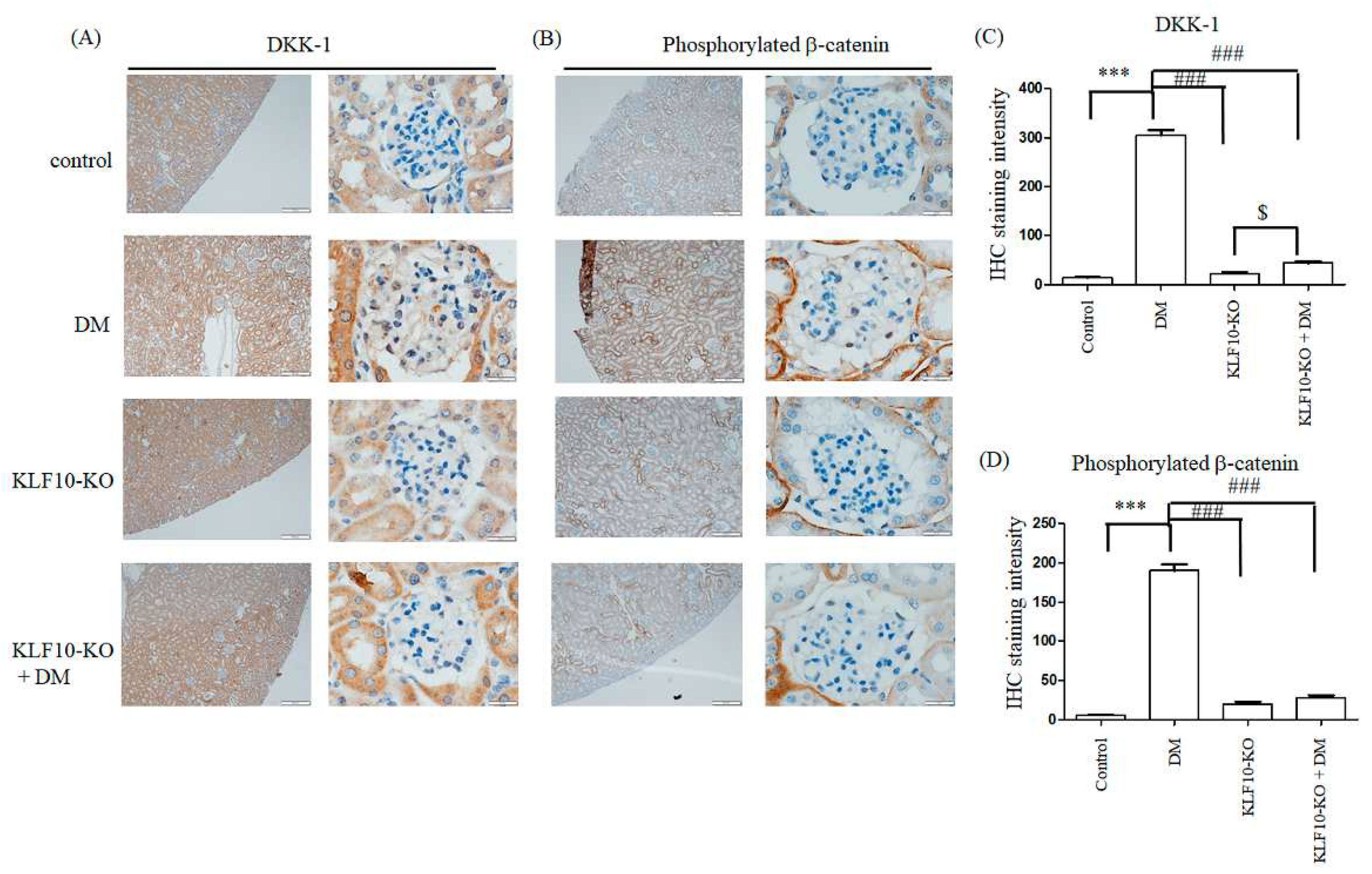
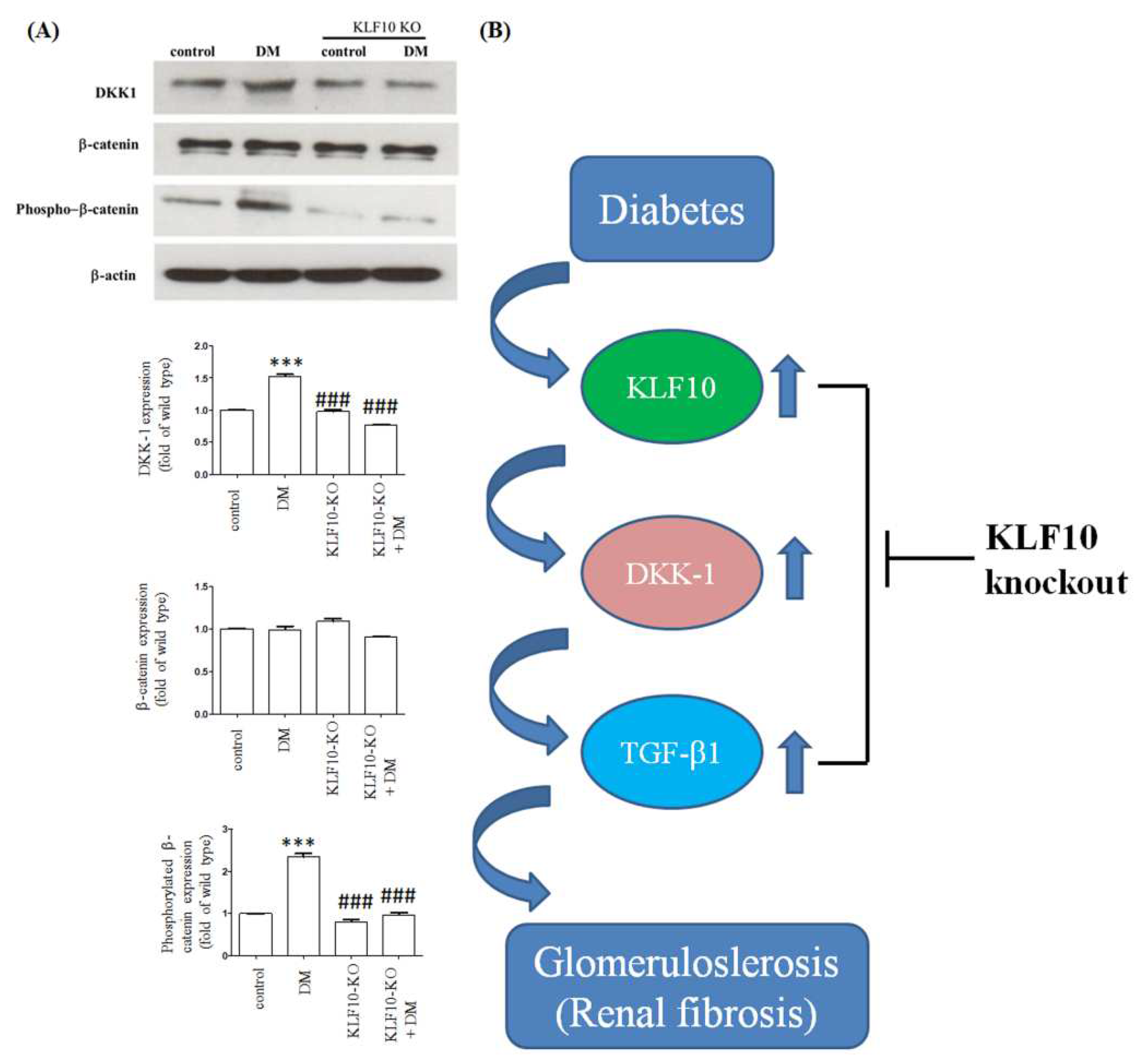
2.2. Knockout of KLF10 and Its Downstream Targets in Diabetic Mice
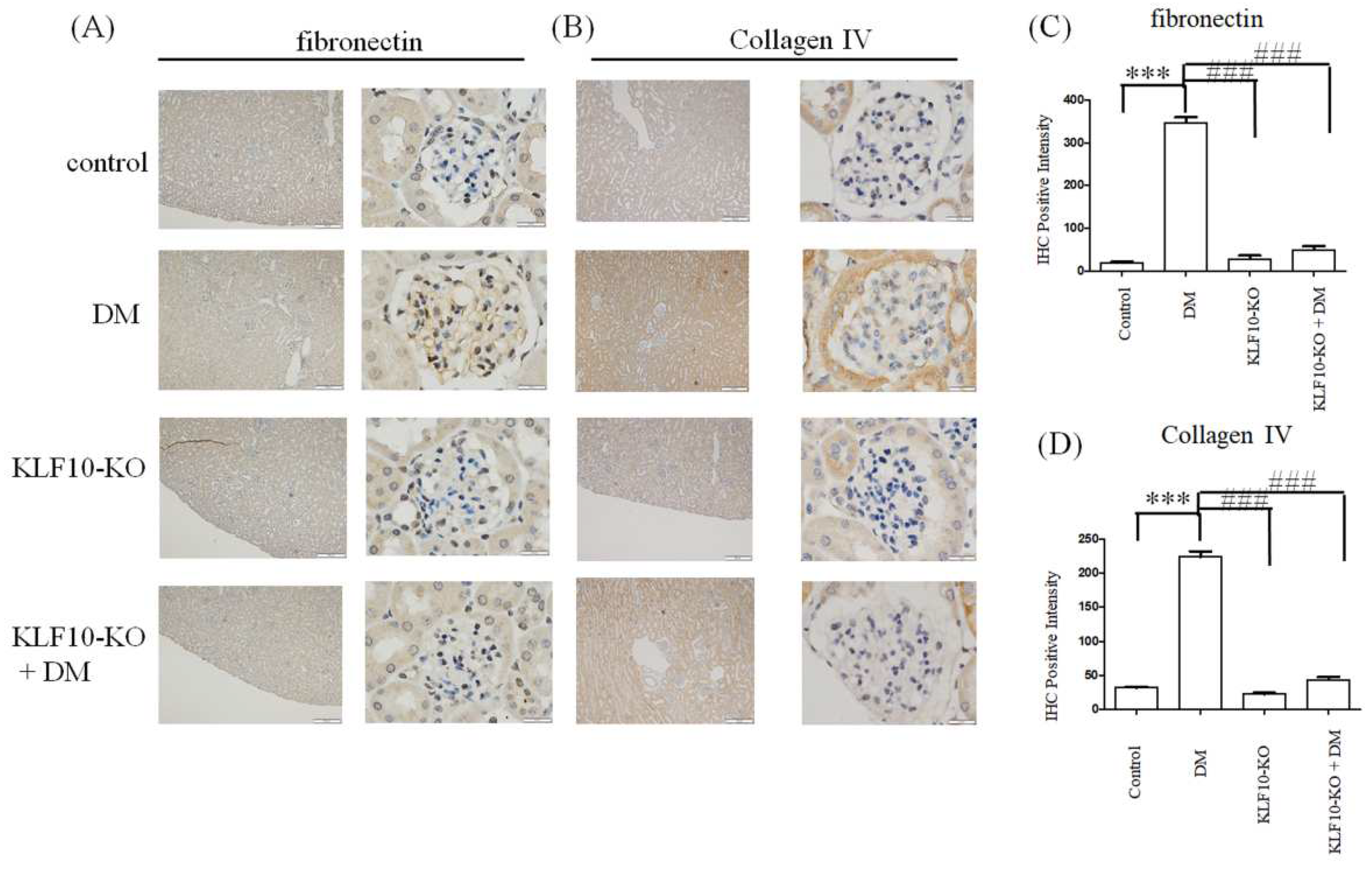
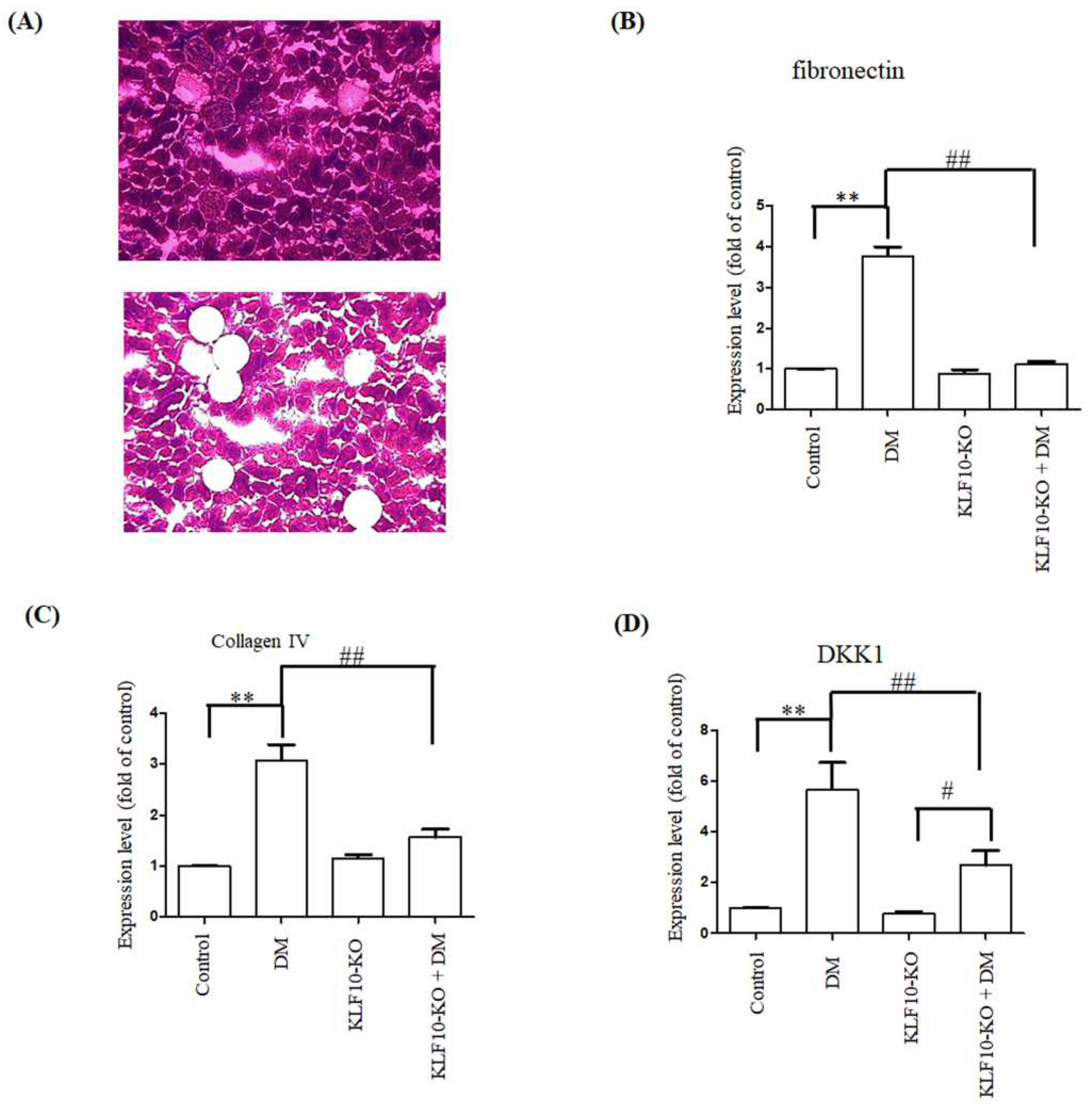
2.3. Effects of KLF10 Knockout on the Expression of Fibrosis-Related Genes
3. Discussion
4. Materials and Methods
4.1. Animal Model
4.2. Immunohistochemical (IHC) and Masson’s Trichrome Staining
4.3. Laser Capture Microdissection
4.4. Quantitative Reverse Transcription–Polymerase Chain Reaction
4.5. Western Blot Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kanter, J.E.; Bornfeldt, K.E. Impact of Diabetes Mellitus. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1049–1053. [Google Scholar] [CrossRef] [Green Version]
- Goraya, T.Y.; Leibson, C.L.; Palumbo, P.J.; Weston, S.A.; Killian, J.M.; Pfeifer, E.A.; Jacobsen, S.J.; Frye, R.L.; Roger, V.L. Coronary atherosclerosis in diabetes mellitus: A population-based autopsy study. J. Am. Coll. Cardiol. 2002, 40, 946–953. [Google Scholar] [CrossRef] [Green Version]
- Sai Laxmi, M.; Prabhakar, O. Inflammatory biomarkers as a part of diagnosis in diabetic peripheral neuropathy. J. Diabetes Metab Disord 2021, 20, 869–882. [Google Scholar]
- Badal, S.S.; Danesh, F.R. New insights into molecular mechanisms of diabetic kidney disease. Am. J. Kidney Dis. 2014, 63, S63–S83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glassock, R.J.; Warnock, D.G.; Delanaye, P. The global burden of chronic kidney disease: Estimates, variability and pitfalls. Nat. Rev. Nephrol. 2017, 13, 104–114. [Google Scholar] [CrossRef]
- Isaka, Y. Targeting TGF-beta Signaling in Kidney Fibrosis. Int J. Mol. Sci. 2018, 19, 2532. [Google Scholar] [CrossRef] [Green Version]
- Tang, P.C.; Chan, A.S.; Zhang, C.B.; Garcia Cordoba, C.A.; Zhang, Y.Y.; To, K.F.; Leung, K.T.; Lan, H.Y.; Tang, P.M. TGF-beta1 Signaling: Immune Dynamics of Chronic Kidney Diseases. Front. Med. Lausanne 2021, 106, 628519. [Google Scholar] [CrossRef]
- Ribeiro, A.; Bronk, S.F.; Roberts, P.J.; Urrutia, R.; Gores, G.J. The transforming growth factor beta(1)-inducible transcription factor TIEG1, mediates apoptosis through oxidative stress. Hepatology 1999, 30, 1490–1497. [Google Scholar] [CrossRef]
- Hu, Z.C.; Shi, F.; Liu, P.; Zhang, J.; Guo, D.; Cao, X.-L.; Chen, C.-F.; Qu, S.Q.; Zhu, J.Y.; Tang, B. TIEG1 Represses Smad7-Mediated Activation of TGF-beta1/Smad Signaling in Keloid Pathogenesis. J. Investig. Dermatol. 2017, 137, 1051–1059. [Google Scholar] [CrossRef] [Green Version]
- Johnsen, S.A.; Subramaniam, M.; Janknecht, R.; Spelsberg, T.C. TGFbeta inducible early gene enhances TGFbeta/Smad-dependent transcriptional responses. Oncogene 2002, 21, 5783–5790. [Google Scholar] [CrossRef] [Green Version]
- Subramaniam, M.; Hawse, J.R.; Johnsen, S.A.; Spelsberg, T.C. Role of TIEG1 in biological processes and disease states. J. Cell Biochem. 2007, 102, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, S.V.; Ivanova, A.V.; Salnikow, K.; Timofeeva, O.; Subramaniam, M.; Lerman, M.I. Two novel VHL targets, TGFBI (BIGH3) and its transactivator KLF10, are up-regulated in renal clear cell carcinoma and other tumors. Biochem. Biophys. Res. Commun. 2008, 370, 536–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.K.; Lee, K.S.; Chang, H.Y.; Lee, W.K.; Lee, J.I. Progression of diet induced nonalcoholic steatohepatitis is accompanied by increased expression of Kruppel-like-factor 10 in mice. J. Transl. Med. 2014, 12, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiMario, J.X. KLF10 Gene Expression Modulates Fibrosis in Dystrophic Skeletal Muscle. Am. J. Pathol. 2018, 188, 1263–1275. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jin, D.; Kang, X.; Zhou, R.; Sun, Y.; Lian, F.; Tong, X. Signaling Pathways Involved in Diabetic Renal Fibrosis. Front. Cell Dev. Biol. 2021, 9, 1728. [Google Scholar] [CrossRef]
- Tan, R.J.; Zhou, D.; Zhou, L.; Liu, Y. Wnt/beta-catenin signaling and kidney fibrosis. Kidney Int. Suppl. 2014, 4, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Klavdianou, K.; Liossis, S.N.; Daoussis, D. Dkk1: A key molecule in joint remodelling and fibrosis. Mediterr. J. Rheumatol. 2017, 28, 174–182. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Dai, C.; Li, Y.; Zeng, G.; Monga, S.P.; Liu, Y. Wnt/beta-catenin signaling promotes renal interstitial fibrosis. J. Am. Soc. Nephrol. 2009, 20, 765–776. [Google Scholar] [CrossRef] [Green Version]
- Dai, C.; Stolz, D.B.; Kiss, L.P.; Monga, S.P.; Holzman, L.B.; Liu, Y. Wnt/beta-catenin signaling promotes podocyte dysfunction and albuminuria. J. Am. Soc. Nephrol. 2009, 20, 1997–2008. [Google Scholar] [CrossRef] [Green Version]
- Hou, N.N.; Kan, C.X.; Huang, N.; Liu, Y.P.; Mao, E.W.; Ma, Y.T.; Han, F.; Sun, H.X.; Sun, X.D. Relationship between serum Dickkopf-1 and albuminuria in patients with type 2 diabetes. World J. Diabetes 2021, 12, 47–55. [Google Scholar]
- Lin, C.L.; Wang, J.Y.; Ko, J.Y.; Huang, Y.T.; Kuo, Y.H.; Wang, F.S. Dickkopf-1 promotes hyperglycemia-induced accumulation of mesangial matrix and renal dysfunction. J. Am. Soc. Nephrol. 2010, 21, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Killick, R.; Ribe, E.M.; Al-Shawi, R.; Malik, B.; Hooper, C.; Fernandes, C.; Dobson, R.; Nolan, P.M.; Lourdusamy, A.; Furney, S.; et al. Clusterin regulates beta-amyloid toxicity via Dickkopf-1-driven induction of the wnt-PCP-JNK pathway. Mol. Psychiatry 2014, 19, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.C.; Chang, P.J.; Lin, S.J.; Liaw, C.C.; Shih, Y.H.; Chen, L.W.; LIn, C.L. Curcumin Reinforces MiR-29a Expression, Reducing Mesangial Fibrosis in a Model of Diabetic Fibrotic Kidney via Modulation of CB1R Signaling. Processes 2021, 9, 694. [Google Scholar] [CrossRef]
- Memon, A.; Lee, W.K. KLF10 as a Tumor Suppressor Gene and Its TGF-beta Signaling. Cancers 2018, 10, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Oh, A.R.; Lee, H.Y.; Moon, Y.A.; Lee, H.J.; Cha, J.Y. Deletion of KLF10 Leads to Stress-Induced Liver Fibrosis upon High Sucrose Feeding. Int. J. Mol. Sci. 2020, 22, 331. [Google Scholar] [CrossRef]
- Wahab, N.A.; Weston, B.S.; Mason, R.M. Modulation of the TGFbeta/Smad signaling pathway in mesangial cells by CTGF/CCN2. Exp. Cell Res. 2005, 307, 305–314. [Google Scholar] [CrossRef]
- Akhmetshina, A.; Palumbo, K.; Dees, C.; Bergmann, C.; Venalis, P.; Zerr, P.; Horn, A.; Kireva, T.; Beyer, C.; Zwerina, J.; et al. Activation of canonical Wnt signalling is required for TGF-beta-mediated fibrosis. Nat. Commun. 2012, 3, 735. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.H.; Hsu, C.F.; Lin, C.Y.; Guo, J.Y.; Yu, W.C.; Chang, V.H. Kruppel-like factor 10 upregulates the expression of cyclooxygenase 1 and further modulates angiogenesis in endothelial cell and platelet aggregation in gene-deficient mice. Int. J. Biochem. Cell Biol. 2013, 45, 419–428. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Chang, P.J.; Ho, C.; Huang, Y.T.; Shih, Y.H.; Wang, C.J.; Lin, C.L. Protective effects of miR-29a on diabetic glomerular dysfunction by modulation of DKK1/Wnt/beta-catenin signaling. Sci. Rep. 2016, 6, 30575. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.L.; Hsu, Y.C.; Huang, Y.T.; Shih, Y.H.; Wang, C.J.; Chiang, W.C.; Chang, P.J. A KDM6A-KLF10 reinforcing feedback mechanism aggravates diabetic podocyte dysfunction. EMBO Mol. Med. 2019, 11, e9828. [Google Scholar]
- Subramaniam, M.; Hawse, J.R.; Rajamannan, N.M.; Ingle, J.N.; Spelsberg, T.C. Functional role of KLF10 in multiple disease processes. Biofactors 2010, 36, 8–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | DM | KLF10-KO | KLF10-KO + DM | |
|---|---|---|---|---|
| Fasting blood glucose (mg/dL) | 156 ± 21 | 487 ± 70 * | 153 ± 17 | 496 ± 56 * |
| HbA1C (%) | 4.1 ± 0.9 | 8.1 ± 0.9 * | 4.3 ± 1.2 | 8.4 ± 0.3 * |
| Body weight (BW, g) | 29.1 ± 1.4 | 24.1 ± 0.7 * | 33.0 ± 1.9 | 23.9 ± 1.2 * |
| Kidney/BW (%) | 0.76 ± 0.21 | 1.10 ± 0.21 * | 0.82 ± 0.23 | 0.89 ± 0.07 *# |
| Urine (TP/CRE) | 5.9 ± 0.7 | 11.2 ± 1.2 | 6.1 ± 0.4 | 8.4 ± 0.7 *# |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, Y.-C.; Ho, C.; Shih, Y.-H.; Ni, W.-C.; Li, Y.-C.; Chang, H.-C.; Lin, C.-L. Knockout of KLF10 Ameliorated Diabetic Renal Fibrosis via Downregulation of DKK-1. Molecules 2022, 27, 2644. https://doi.org/10.3390/molecules27092644
Hsu Y-C, Ho C, Shih Y-H, Ni W-C, Li Y-C, Chang H-C, Lin C-L. Knockout of KLF10 Ameliorated Diabetic Renal Fibrosis via Downregulation of DKK-1. Molecules. 2022; 27(9):2644. https://doi.org/10.3390/molecules27092644
Chicago/Turabian StyleHsu, Yung-Chien, Cheng Ho, Ya-Hsueh Shih, Wen-Chiu Ni, Yi-Chen Li, Hsiu-Ching Chang, and Chun-Liang Lin. 2022. "Knockout of KLF10 Ameliorated Diabetic Renal Fibrosis via Downregulation of DKK-1" Molecules 27, no. 9: 2644. https://doi.org/10.3390/molecules27092644
APA StyleHsu, Y.-C., Ho, C., Shih, Y.-H., Ni, W.-C., Li, Y.-C., Chang, H.-C., & Lin, C.-L. (2022). Knockout of KLF10 Ameliorated Diabetic Renal Fibrosis via Downregulation of DKK-1. Molecules, 27(9), 2644. https://doi.org/10.3390/molecules27092644