
LC/ESI/TOF-MS Characterization, Anxiolytic and Antidepressant-like Effects of Mitragyna speciosa Korth Extract in Diabetic Rats



Abstract
:1. Introduction
2. Results
2.1. LC-ESI-QTOF-MS Analysis for the Identification of Metabolites in MSE
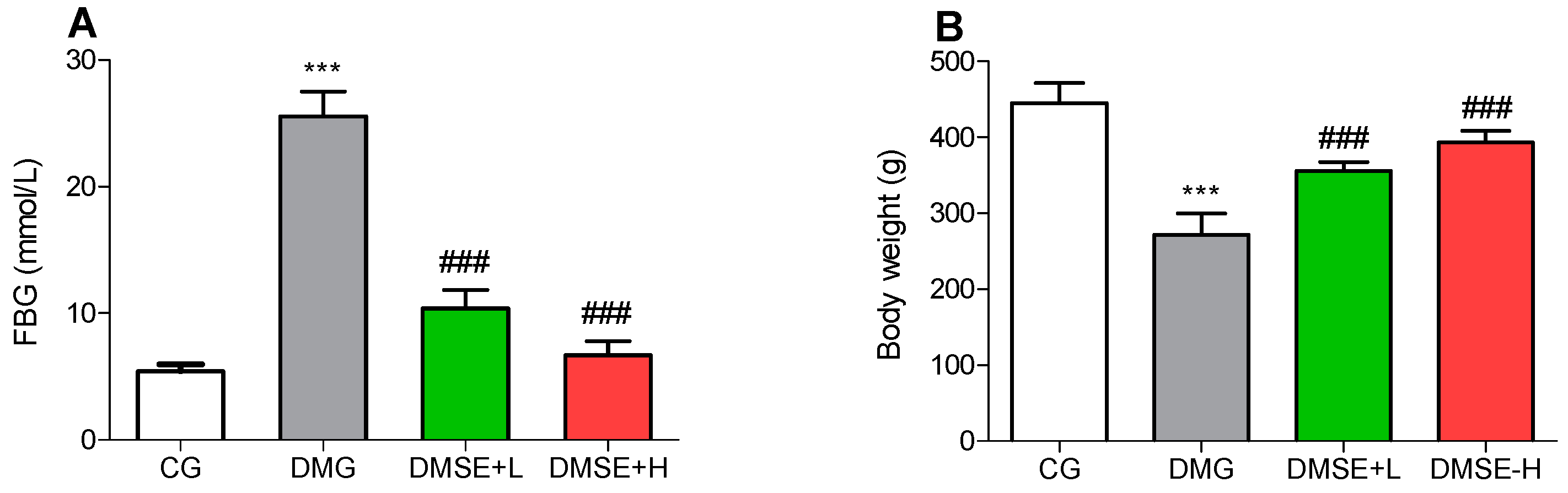
2.2. Effect of MSE on Blood Glucose
2.3. Effect of MSE on Body Weight
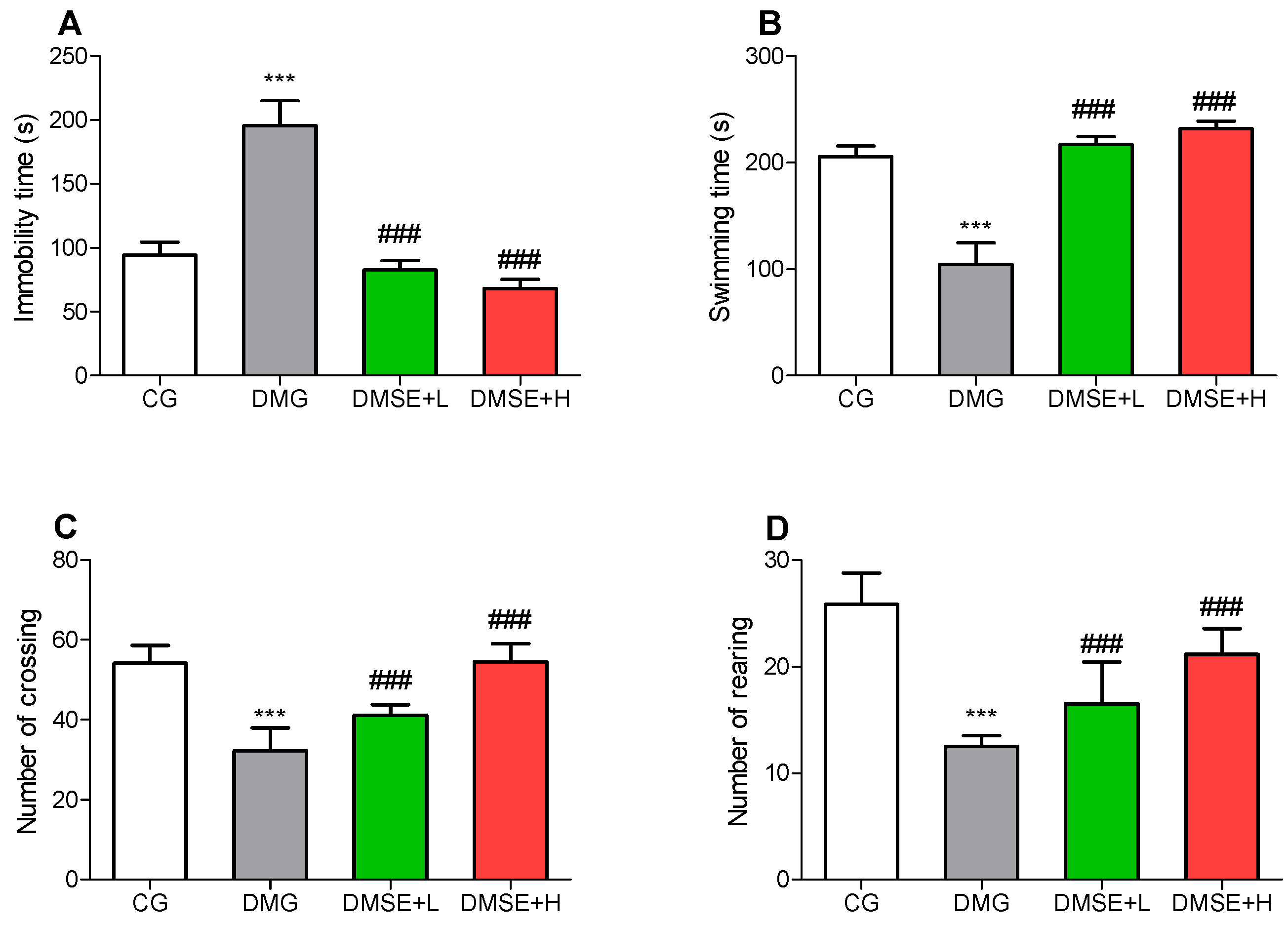
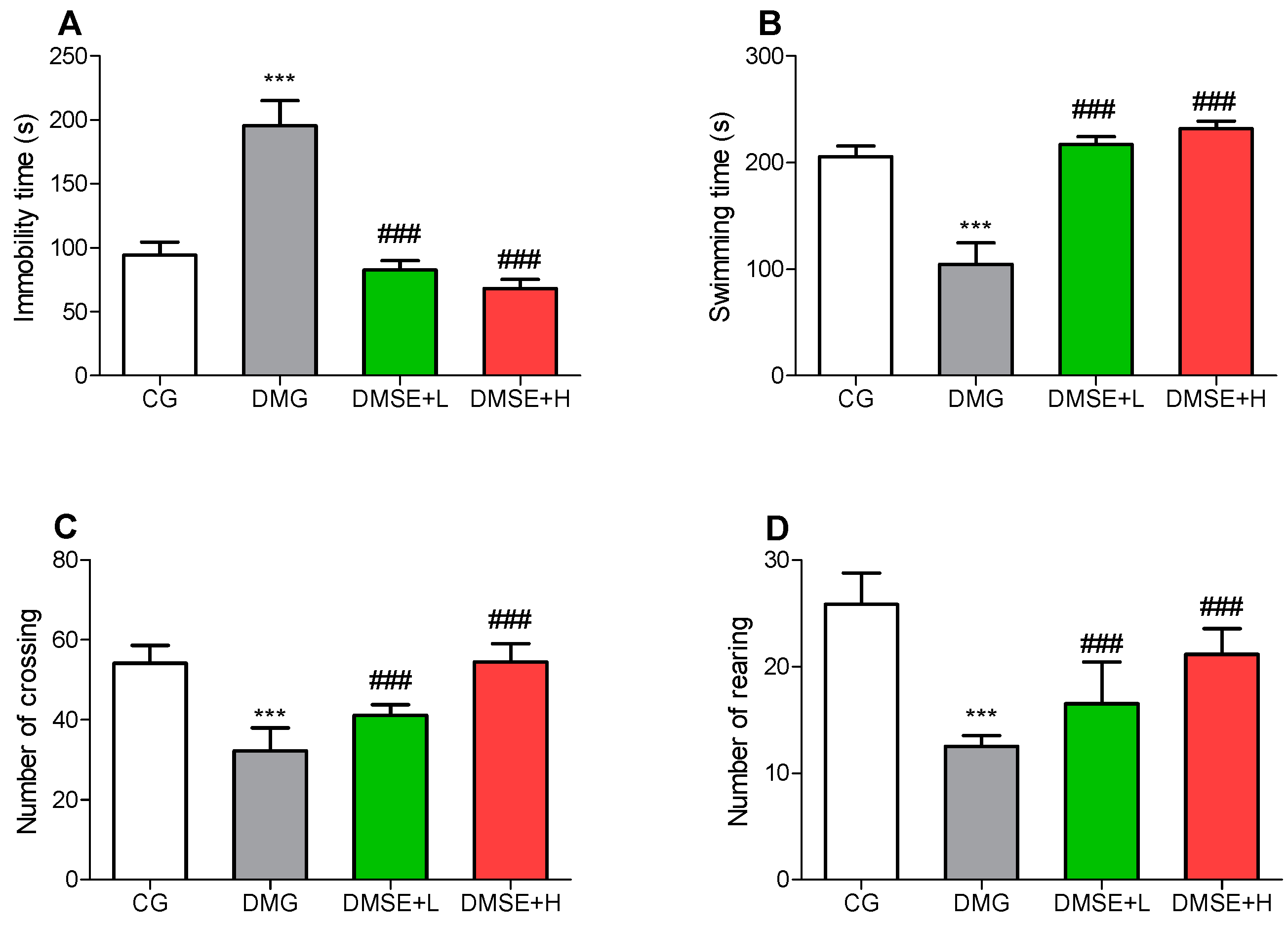
2.4. Effect of MSE on Depressive-like Behaviors
2.5. Effect of MSE on Locomotor Activity
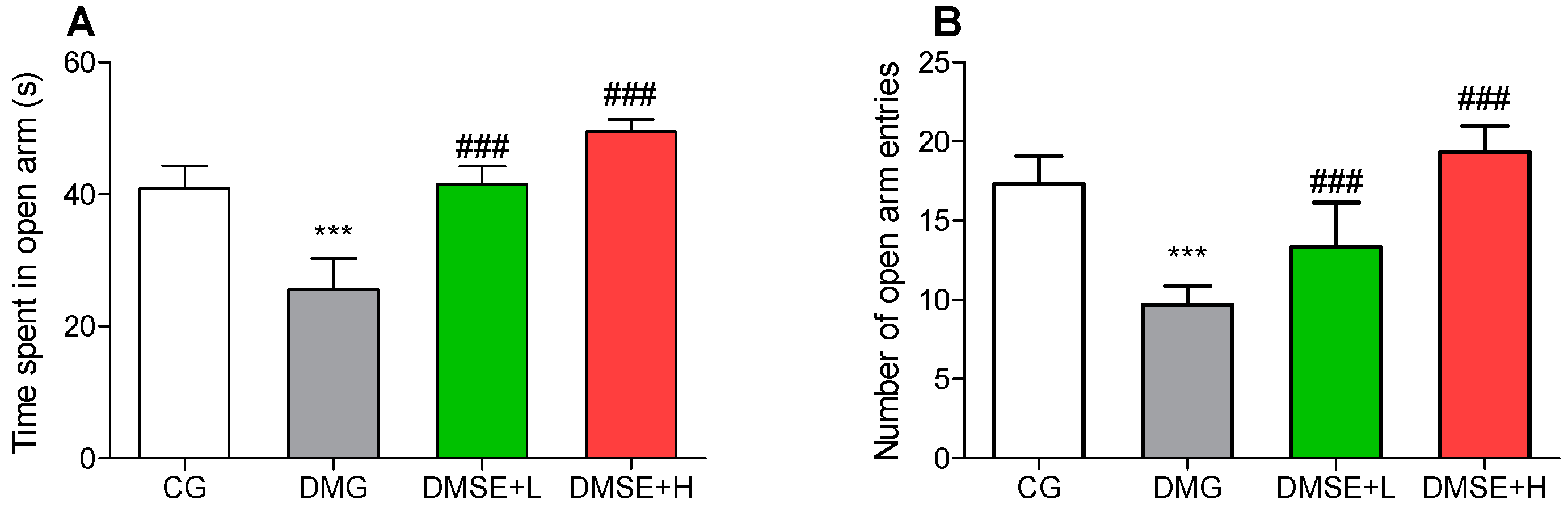
2.6. Effect of MSE on Anxiety-like Behavior
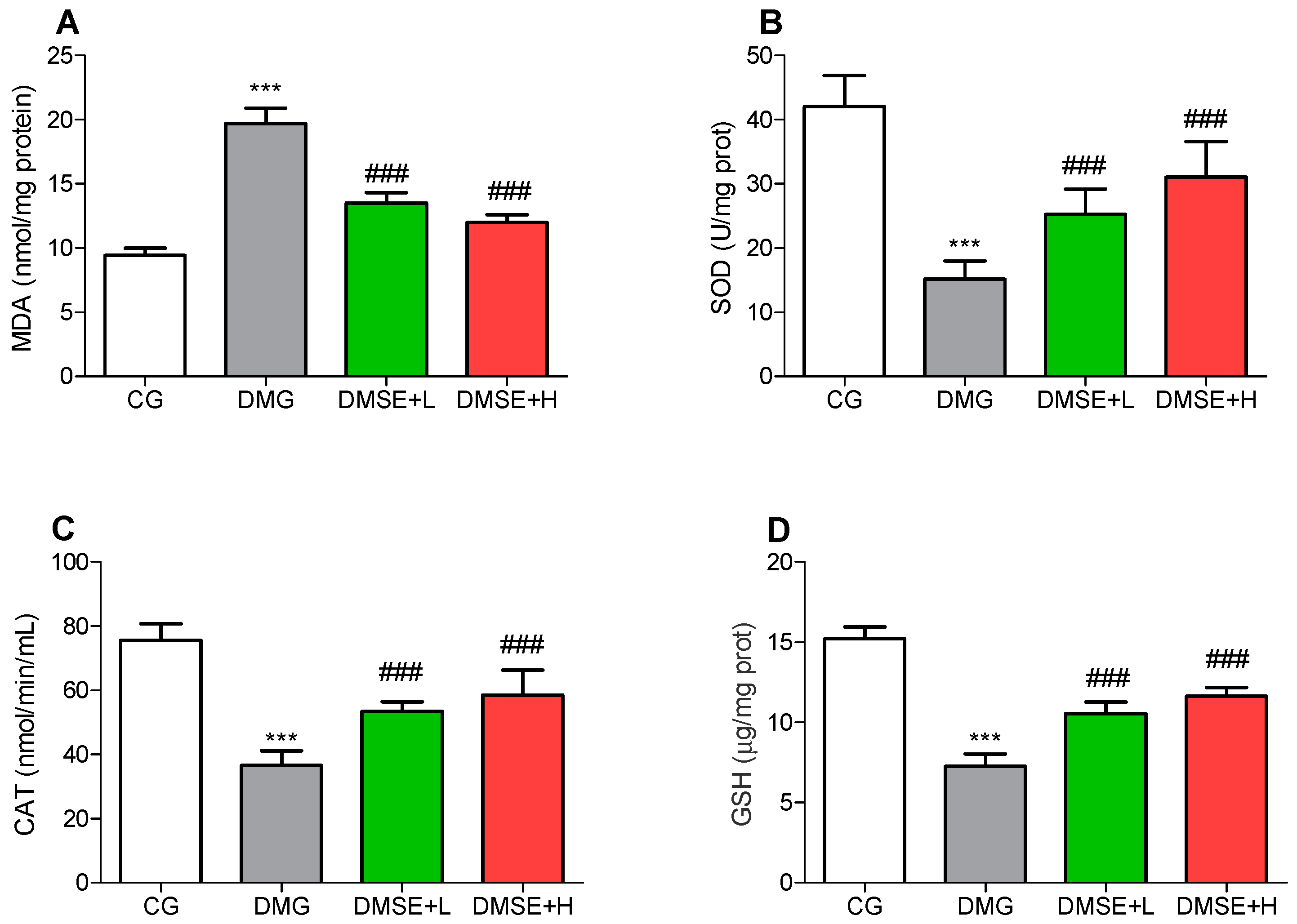
2.7. Effect of MSE on Oxidative Stress Parameters
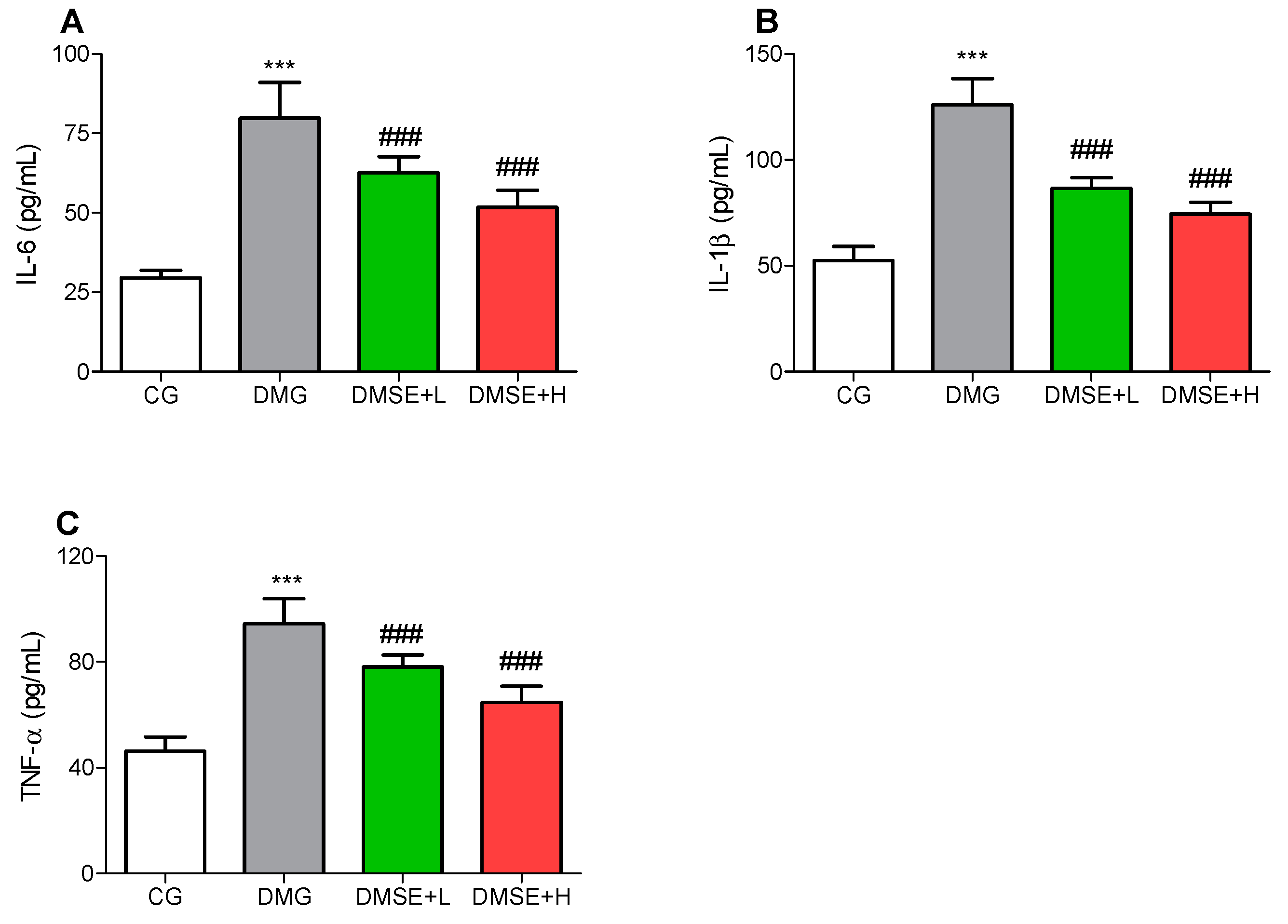
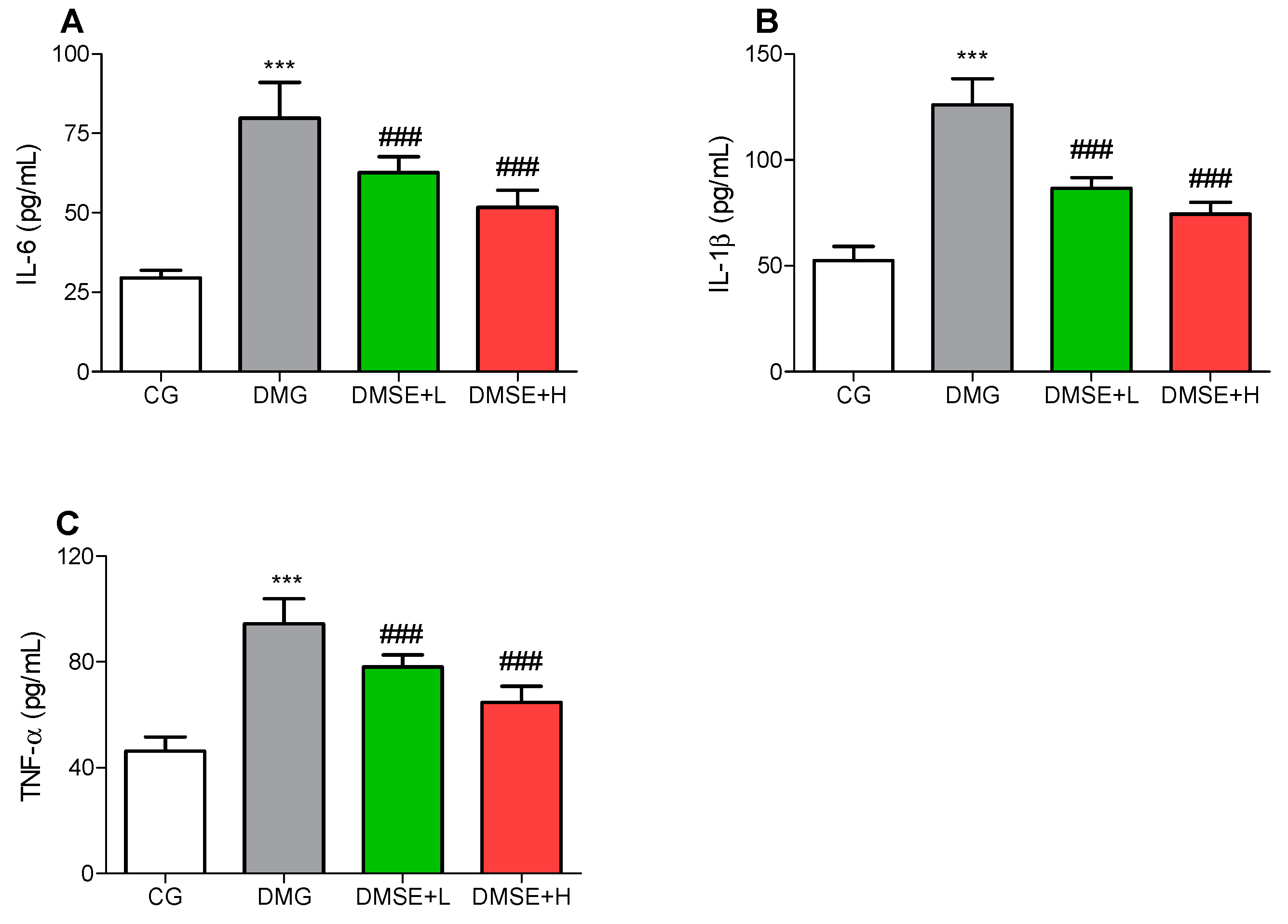
2.8. Effect of MSE on Proinflammatory Parameters
3. Discussion
4. Materials and Methods
4.1. Preparation of M. speciosa Extract
4.2. Metabolite Profiling Using Ultra-High-Performance Liquid Chromatography Coupled to Electrospray Time-of-Flight Tandem Mass Spectrometry (UPLC/ESI/TOF-MS)
4.3. Animals and Experimental Design
4.4. Diabetes Induction
4.5. Open Field Test
4.6. Elevated Plus Maze Test
4.7. Forced Swimming Test
4.8. Sacrifice and Sample Collection
4.9. Analysis of Oxidative and Inflammatory Biomarkers
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Makinde, E.A.; Radenahmad, N.; Adekoya, A.E.; Olatunji, O.J. Tiliacora triandra extract possesses antidiabetic effects in high fat diet/streptozotocin-induced diabetes in rats. J. Food Biochem. 2020, 44, e13239. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Sun, C.; Li, J.; Long, T.; Yan, Y.; Qin, H.; Makinde, E.A.; Famurewa, A.C.; Jaisi, A.; Nie, Y.; et al. Tiliacora triandra extract and its major constituent attenuates diabetic kidney and testicular impairment by modulating redox imbalance and pro-inflammatory responses in rats. J. Sci. Food Agric. 2021, 101, 1598–1608. [Google Scholar] [CrossRef] [PubMed]
- Erukainure, O.L.; Oyebode, O.A.; Ijomone, O.M.; Chukwuma, C.I.; Koorbanally, N.A.; Islam, M.S. Raffia palm (Raphia hookeri G. Mann & H. Wendl) wine modulates glucose homeostasis by enhancing insulin secretion and inhibiting redox imbalance in a rat model of diabetes induced by high fructose diet and streptozotocin. J. Ethnopharmacol. 2019, 237, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Olatunji, O.J.; Chen, H.; Zhou, Y. Lycium chinense leaves extract ameliorates diabetic nephropathy by suppressing hyperglycemia mediated renal oxidative stress and inflammation. Biomed. Pharmacother. 2018, 102, 1145–1151. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.J.; Freedland, K.E.; Clouse, R.E.; Lustman, P.J. The prevalence of comorbid depression in adults with diabetes: A meta-analysis. Diabetes Care 2001, 24, 1069–1078. [Google Scholar] [CrossRef] [Green Version]
- Moulton, C.D.; Pickup, J.C.; Ismail, K. The link between depression and diabetes: The search for shared mechanisms. Lancet Diabetes Endocrinol. 2015, 3, 461–471. [Google Scholar] [CrossRef]
- Zimath, P.L.; Dalmagro, A.P.; Mota da Silva, L.; Malheiros, A.; Maria de Souza, M. Myrsinoic acid B from Myrsine coriacea reverses depressive-like behavior and brain oxidative stress in streptozotocin-diabetic rats. Chem. Biol. Interact. 2021, 347, 109603. [Google Scholar] [CrossRef]
- Rahmani, G.; Farajdokht, F.; Mohaddes, G.; Babri, S.; Ebrahimi, V.; Ebrahimi, H. Garlic (Allium sativum) improves anxiety- and depressive-related behaviors and brain oxidative stress in diabetic rats. Arch. Physiol. Biochem. 2020, 126, 95–100. [Google Scholar] [CrossRef]
- Siddiqui, S. Depression in type 2 diabetes mellitus-a brief review. Diabetes Metab. Syndr. Clin. Res. Rev. 2014, 8, 62–65. [Google Scholar] [CrossRef]
- de Morais, H.; de Souza, C.P.; da Silva, L.M.; Ferreira, D.M.; Werner, M.F.; Andreatini, R.; da Cunha, J.M.; Zanoveli, J.M. Increased oxidative stress in prefrontal cortex and hippocampus is related to depressive-like behavior in streptozotocin-diabetic rats. Behav. Brain Res. 2014, 258, 52–64. [Google Scholar] [CrossRef]
- Menezes Zanoveli, J.; de Morais, H.; Caroline da Silva Dias, I.; Karoline Schreiber, A.; Pasquini de Souza, C.; Maria da Cunha, J. Depression associated with diabetes: From pathophysiology to treatment. Curr. Diabetes Rev. 2016, 12, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Makinde, E.A.; Eze, F.N.; Olatunji, O.J. Securidaca inappendiculata polyphenol rich extract counteracts cognitive deficits, neuropathy, neuroinflammation and oxidative stress in diabetic encephalopathic rats via p38 MAPK/Nrf2/HO-1 pathways. Front. Pharmacol. 2021, 12, 737764. [Google Scholar] [CrossRef] [PubMed]
- Makinde, E.A.; Ovatlarnporn, C.; Adekoya, A.E.; Nwabor, O.F.; Olatunji, O.J. Antidiabetic, antioxidant and antimicrobial activity of the aerial part of Tiliacora triandra. S. Afr. J. Bot. 2019, 125, 337–343. [Google Scholar] [CrossRef]
- Ahmad, I.; Prabowo, W.C.; Arifuddin, M.; Fadraersada, J.; Indriyanti, N.; Herman, H.; Purwoko, R.Y.; Nainu, F.; Rahmadi, A.; Paramita, S.; et al. Mitragyna species as pharmacological agents: From abuse to promising pharmaceutical products. Life 2022, 12, 193. [Google Scholar] [CrossRef]
- Domnic, G.; Jeng-Yeou Chear, N.; Abdul Rahman, S.F.; Ramanathan, S.; Lo, K.-W.; Singh, D.; Mohana-Kumaran, N. Combinations of indole based alkaloids from Mitragyna speciosa (Kratom) and cisplatin inhibit cell proliferation and migration of nasopharyngeal carcinoma cell lines. J. Ethnopharmacol. 2021, 279, 114391. [Google Scholar] [CrossRef] [PubMed]
- Meireles, V.; Rosado, T.; Barroso, M.; Soares, S.; Gonçalves, J.; Luís, Â.; Caramelo, D.; Simão, A.Y.; Fernández, N.; Duarte, A.P.; et al. Mitragyna speciosa: Clinical, toxicological aspects and analysis in biological and non-biological samples. Medicines 2019, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Narayanan, S.; Vicknasingam, B. Traditional and non-traditional uses of Mitragynine (Kratom): A survey of the literature. Brain Res. Bull. 2016, 126, 41–46. [Google Scholar] [CrossRef]
- Goh, Y.S.; Karunakaran, T.; Murugaiyah, V.; Santhanam, R.; Abu Bakar, M.H.; Ramanathan, S. Accelerated solvent extractions (ASE) of Mitragyna speciosa Korth. (Kratom) leaves: Evaluation of its cytotoxicity and antinociceptive activity. Molecules 2021, 26, 3704. [Google Scholar] [CrossRef]
- Srichana, K.; Janchawee, B.; Prutipanlai, S.; Raungrut, P.; Keawpradub, N. Effects of mitragynine and a crude alkaloid extract derived from Mitragyna speciosa korth. On permethrin elimination in rats. Pharmaceutics 2015, 7, 10–26. [Google Scholar] [CrossRef] [Green Version]
- Parthasarathy, S.; Bin Azizi, J.; Ramanathan, S.; Ismail, S.; Sasidharan, S.; Mohd, M.I.; Mansor, S.M. Evaluation of antioxidant and antibacterial activities of aqueous, methanolic and alkaloid extracts from Mitragyna speciosa (rubiaceae family) leaves. Molecules 2009, 14, 3964–3974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Idayu, N.F.; Hidayat, M.T.; Moklas, M.A.M.; Sharida, F.; Raudzah, A.R.N.; Shamima, A.R.; Apryani, E. Antidepressant-like effect of mitragynine isolated from Mitragyna speciosa Korth in mice model of depression. Phytomedicine 2011, 18, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.N.; Lund, J.A.; Murch, S.J. A botanical, phytochemical and ethnomedicinal review of the genus Mitragyna Korth: Implications for products sold as kratom. J. Ethnopharmacol. 2017, 202, 302–325. [Google Scholar] [CrossRef]
- Zakaria, F.; Tan, J.-K.; Faudzi, S.M.M.; Rahman, M.B.A.; Ashari, S.E. Ultrasound-assisted extraction condition optimization using response surface methodology from Mitragyna speciosa (Korth.) Havil leaves. Ultrason. Sonochem. 2021, 81, 105851. [Google Scholar] [CrossRef] [PubMed]
- Gerhard, D.M.; Wohleb, E.S.; Duman, R.S. Emerging treatment mechanisms for depression: Focus on glutamate and synaptic plasticity. Drug Discov. Today 2016, 21, 454–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olatunji, O.J.; Zuo, J.; Olatunde, O.O. Securidaca inappendiculata stem extract confers robust antioxidant and antidiabetic effects against high fructose/streptozotocin induced type 2 diabetes in rats. Exploration of bioactive compounds using UHPLC-ESI-QTOF-MS. Arch. Physiol. Biochem. 2021, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bampi, S.R.; Casaril, A.M.; Fronza, M.G.; Domingues, M.; Vieira, B.; Begnini, K.R.; Seixas, F.K.; Collares, T.V.; Lenardão, E.J.; Savegnago, L. The selenocompound 1-methyl-3-(phenylselanyl)-1H-indole attenuates depression-like behavior, oxidative stress, and neuroinflammation in streptozotocin-treated mice. Brain Res. Bull. 2020, 161, 158–165. [Google Scholar] [CrossRef]
- Gasparin, A.T.; Rosa, E.S.; Jesus, C.H.A.; Guiloski, I.C.; da Silva de Assis, H.C.; Beltrame, O.C.; Dittrich, R.L.; Pacheco, S.D.G.; Zanoveli, J.M.; da Cunha, J.M. Bixin attenuates mechanical allodynia, anxious and depressive-like behaviors associated with experimental diabetes counteracting oxidative stress and glycated hemoglobin. Brain Res. 2021, 1767, 147557. [Google Scholar] [CrossRef]
- Tyagi, A.; Pugazhenthi1, S. Targeting insulin resistance to treat cognitive dysfunction. Mol. Neurobiol. 2021, 58, 2672–2691. [Google Scholar] [CrossRef]
- Kong, S.H.; Park, Y.J.; Lee, J.Y.; Cho, N.H.; Moon, M.K. Insulin resistance is associated with cognitive decline among older Koreans with normal baseline cognitive function: A prospective community-based cohort study. Sci. Rep. 2018, 8, 650. [Google Scholar] [CrossRef] [Green Version]
- Leonard, B.E.; Wegener, G. Inflammation, insulin resistance and neuroprogression in depression. Acta Neuropsychiatr. 2020, 32, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.A.; Habila, J.D.; Koorbanally, N.A.; Islam, M.S. Butanol fraction of Parkia biglobosa (Jacq.) G. Don leaves enhance pancreatic β-cell functions, stimulates insulin secretion and ameliorates other type 2 diabetes-associated complications in rats. J. Ethnopharmacol. 2016, 183, 103–111. [Google Scholar] [CrossRef]
- Bikri, S.; Aboussaleh, Y.; Berrani, A.; Louragli, I.; Hafid, A.; Chakib, S.; Ahami, A. Effects of date seeds administration on anxiety and depressive symptoms in streptozotocin-induced diabetic rats: Biochemical and behavioral evidences. J. Basic Clin. Physiol. Pharmacol. 2021, 32, 1031–1040. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Hong, T.W.; Wang, Y.J.; Chen, K.C.; Pei, J.C.; Chuang, T.Y.; Lai, W.S.; Tsai, S.H.; Chu, R.; Chen, W.C.; et al. Ophiocordyceps formosana improves hyperglycemia and depression-like behavior in an STZ-induced diabetic mouse model. BMC Complement. Altern. Med. 2016, 16, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Wang, J.; Song, L.; Guan, Y.; Cao, C.; Cui, Y.; Zhang, Y.; Liu, C. Catalpol weakens depressive-like behavior in mice with streptozotocin-induced hyperglycemia via PI3K/AKT/Nrf2/HO-1 signaling pathway. Neuroscience 2021, 473, 102–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, C.; Elsheikh, N.A.H.; Li, C.; Yang, F.; Wang, G.; Li, L. HO-1 reduces heat stress-induced apoptosis in bovine granulosa cells by suppressing oxidative stress. Aging 2019, 11, 5535–5547. [Google Scholar] [CrossRef] [PubMed]
- Reus, G.Z.; Carlessi, A.S.; Silva, R.H.; Ceretta, L.B.; Quevedo, J. Relationship of oxidative stress as a link between diabetes mellitus and major depressive disorder. Oxid. Med. Cell. Longev. 2019, 2019, 1–6. [Google Scholar] [CrossRef]
- Gu, F.; Chauhan, V.; Chauhan, A. Glutathione redox imbalance in brain disorders. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 89–95. [Google Scholar] [CrossRef]
- Ceriello, A. New insights on oxidative stress and diabetic complications may lead to a “causal” antioxidant therapy. Diabetes Care 2003, 26, 1589–1596. [Google Scholar] [CrossRef] [Green Version]
- Moylan, S.; Berk, M.; Dean, O.M.; Samuni, Y.; Williams, L.J.; O’Neil, A.; Hayley, A.C.; Pasco, J.A.; Anderson, G.; Jacka, F.N.; et al. Oxidative & nitrosative stress in depression: Why so much stress? Neurosci. Biobehav. Rev. 2014, 45, 46–62. [Google Scholar]
- Zborowski, V.A.; Heck, S.O.; Vencato, M.; Pinton, S.; Marques, L.S.; Nogueira, C.W. Keap1/Nrf2/HO-1 signaling pathway contributes to p-chlorodiphenyl diselenide antidepressant-like action in diabetic mice. Psychopharmacology 2020, 237, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Jawale, A.; Datusalia, A.K.; Bishnoi, M.; Sharma, S.S. Reversal of diabetes-induced behavioral and neurochemical deficits by cinnamaldehyde. Phytomedicine 2016, 23, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.S.; Ray, R.S.; Sharma, A.; Mehta, V.; Katyal, A.; Udayabanu, M. Antidepressant and anxiolytic like effects of Urtica dioica leaves in streptozotocin induced diabetic mice. Metab. Brain Dis. 2018, 33, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Eze, F.N.; Jayeoye, T.J. Chromolaena odorata (Siam weed): A natural reservoir of bioactive compounds with potent anti-fibrillogenic, antioxidative, and cytocompatible properties. Biomed. Pharmacother. 2021, 141, 111811. [Google Scholar] [CrossRef] [PubMed]
- Pathak, L.; Agrawal, Y.; Dhir, A. Natural polyphenols in the management of major depression. Expert Opin. Investig. Drugs. 2013, 22, 863–880. [Google Scholar] [CrossRef] [PubMed]
- Flores-Bocanegra, L.; Raja, H.A.; Graf, T.N.; Augustinović, M.; Wallace, E.D.; Hematian, S.; Kellogg, J.J.; Todd, D.A.; Cech, N.B.; Oberlies, N.H. The chemistry of kratom [Mitragyna speciosa]: Updated characterization data and methods to elucidate indole and oxindole alkaloids. J. Nat. Prod. 2020, 83, 2165–2177. [Google Scholar] [CrossRef]
- Alzahrani, S.; Said, E.; Ajwah, S.M.; Alsharif, S.Y.; El-Bayoumi, K.S.; Zaitone, S.A.; Qushawy, M.; Elsherbiny, N.M. Isoliquiritigenin attenuates inflammation and modulates Nrf2/caspase-3 signalling in STZ-induced aortic injury. J. Pharm. Pharmacol. 2021, 73, 193–205. [Google Scholar] [CrossRef]
- Sun, L.; Yang, Z.; Zhang, J.; Wang, J. Isoliquiritigenin attenuates acute renal injury through suppressing oxidative stress, fibrosis and JAK2/STAT3 pathway in streptozotocin-induced diabetic rats. Bioengineered 2021, 12, 11188–11200. [Google Scholar] [CrossRef]
- Prajapati, R.; Seong, S.H.; Park, S.E.; Paudel, P.; Jung, H.A.; Choi, J.S. Isoliquiritigenin, a potent human monoamine oxidase inhibitor, modulates dopamine D1, D3, and vasopressin V1A receptors. Sci. Rep. 2021, 11, 23528. [Google Scholar] [CrossRef]
- Silvestro, S.; Bramanti, P.; Mazzon, E. Role of quercetin in depressive-like behaviors: Findings from animal models. Appl. Sci. 2021, 11, 7116. [Google Scholar] [CrossRef]
- Prabakaran, D.; Ashokkumar, N. Protective effect of esculetin on hyperglycemia-mediated oxidative damage in the hepatic and renal tissues of experimental diabetic rats. Biochimie 2013, 95, 366–373. [Google Scholar] [CrossRef]
- Zhu, L.; Nang, C.; Luo, F.; Pan, H.; Zhang, K.; Liu, J.; Zhou, R.; Gao, J.; Chang, X.; He, H.; et al. Esculetin attenuates lipopolysaccharide (LPS)-induced neuroinflammatory processes and depressive-like behavior in mice. Physiol. Behav. 2016, 163, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Jaishree, V.; Narsimha, S. Swertiamarin and quercetin combination ameliorates hyperglycemia, hyperlipidemia and oxidative stress in streptozotocin-induced type 2 diabetes mellitus in wistar rats. Biomed. Pharmacother. 2020, 130, 110561. [Google Scholar] [CrossRef] [PubMed]
- Parwani, K.; Patel, F.; Bhagwat, P.; Dilip, H.; Patel, D.; Thiruvenkatam, V.; Mandal, P. Swertiamarin mitigates nephropathy in high-fat diet/streptozotocin-induced diabetic rats by inhibiting the formation of advanced glycation end products. Arch. Physiol. Biochem. 2021, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Eze, F.N.; Tola, A.J. Protein glycation and oxidation inhibitory activity of Centella asiatica phenolics (CAP) in glucose-mediated bovine serum albumin glycoxidation. Food Chem. 2020, 332, 127302. [Google Scholar] [CrossRef]
- Reanmongkol, W.; Keawpradub, N.; Sawangjaroen, K. Effects of the extracts from Mitragyna speciosa Korth. leaves on analgesic and behavioral activities in experimental animals. Songklanakarin J. Sci. Technol. 2007, 29, 1. [Google Scholar]
- Vijeepallam, K.; Pandy, V.; Kunasegaran, T.; Murugan, D.D.; Naidu, M. Mitragyna speciosa leaf extract exhibits antipsychotic-like effect with the potential to alleviate positive and negative symptoms of psychosis in mice. Front. Pharmacol. 2016, 7, 464. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RT (min) | Mass (m/z) | Elemental Composition | DB Diff (ppm) | Tentative Metabolite Identity |
|---|---|---|---|---|---|
| 1 | 1.639 | 426.0767 | C19H16F2O9 | −1.01 | Diflunisal phenolic glucuronide |
| 2 | 1.662 | 252.0953 | C8H16N2O7 | 1.81 | Cycasin |
| 3 | 1.719 | 556.1377 | C31H24O10 | −1.39 | Dianhydroaurasperone C |
| 4 | 1.725 | 370.09 | C16H18O10 | 0.01 | Isoferuloyl C1-glucuronide |
| 5 | 1.729 | 654.2207 | C36H34N2O10 | 1 | Citbismine F |
| 6 | 1.778 | 128.0467 | C6H8O3 | 5.11 | Osmundalactone |
| 7 | 1.787 | 518.1835 | C19H34O16 | 2.32 | Ciceritol |
| 8 | 1.81 | 696.1922 | C31H36O18 | −2.9 | Malvidin 3-(6″-acetylglucoside)-5-glucoside |
| 9 | 1.831 | 366.1128 | C18H22O6S | 2.57 | 4-Hydroxyestrone sulfate |
| 10 | 1.885 | 470.088 | C20H22O11S | 0.55 | (Z)-Resveratrol 3-(3″-sulfoglucoside) |
| 11 | 1.907 | 238.1043 | C9H18O7 | 4.07 | (x)-1,2-Propanediol 1-O-β-d-glucopyranoside |
| 12 | 1.922 | 291.095 | C11H17NO8 | 1.32 | Sarmentosin epoxide |
| 13 | 2.216 | 135.0544 | C5H5N5 | 0.87 | Adenine |
| 14 | 2.35 | 636.0096 | C22H20O18S2 | −0.8 | Isoscutellarein 4′-methyl ether 8-(2″,4″-disulfatoglucuronide) |
| 15 | 2.354 | 408.1068 | C19H20O10 | −2.76 | Khellol glucoside |
| 16 | 2.75 | 283.091 | C10H13N5O5 | 2.51 | Crotonoside |
| 17 | 4.43 | 406.1469 | C17H26O11 | 1.42 | Shanzhiside methyl ester |
| 18 | 5.101 | 390.1158 | C16H22O11 | 1.14 | Theveside |
| 19 | 5.284 | 402.1157 | C17H22O11 | 1.26 | 4-Hydroxy-5-(3′,5′-dihydroxyphenyl)-valeric acid-O-glucuronide |
| 20 | 5.311 | 316.0787 | C13H16O9 | 2.34 | Protocatechuic acid-3-glucoside |
| 21 | 5.379 | 332.1101 | C14H20O9 | 1.91 | Leonuriside A |
| 22 | 5.453 | 464.0948 | C21H20O12 | 1.48 | Herbacetin 7-glucoside |
| 23 | 5.529 | 356.073 | C15H16O10 | 1.51 | Caffeoyl C1-glucuronide |
| 24 | 6.221 | 300.0839 | C13H16O8 | 1.91 | Salicylic acid β-d-glucoside |
| 25 | 6.417 | 360.105 | C15H20O10 | 1.89 | Glucosyringic acid |
| 26 | 6.435 | 376.1363 | C16H24O10 | 1.72 | Mussaenosidic acid |
| 27 | 6.456 | 436.1 | C20H20O11 | 1.22 | Taxifolin 3-arabinoside |
| 28 | 6.722 | 112.0159 | C5H4O3 | 0.99 | Pyromeconic acid |
| 29 | 7.546 | 404.1315 | C17H24O11 | 1.02 | Secoxyloganin |
| 30 | 7.663 | 286.0685 | C12H14O8 | 1.4 | Uralenneoside |
| 31 | 7.665 | 342.0947 | C15H18O9 | 1.23 | Glucocaffeic acid |
| 32 | 7.813 | 406.2193 | C19H34O9 | 2.53 | (3S,5R,6R,7E,9S)-Megastigman-7-ene-3,5,6,9-tetrol 9-O-β-d-glucopyranoside |
| 33 | 8.017 | 376.1365 | C16H24O10 | 1.3 | Loganic acid |
| 34 | 8.135 | 596.1521 | C30H28O13 | 1.44 | Eriodictyol 7-(6-trans-p-coumaroylglucoside) |
| 35 | 8.259 | 342.0948 | C15H18O9 | 0.88 | Glucocaffeic acid |
| 36 | 8.286 | 444.199 C | C21H32O10 | 1.26 | Cynaroside A |
| 37 | 8.879 | 446.1419 | C19H26O12 | 1.16 | Lucuminic acid |
| 38 | 8.884 | 418.1461 | C18H26O11 | 3.32 | 2-C-Methyl-d-erythritol 1-O-β-d-(6-O-4-hydroxybenzoyl)glucopyranoside |
| 39 | 8.928 | 402.1893 | C19H30O9 | −0.8 | d-Linalool 3-(6″-malonylglucoside) |
| 40 | 8.976 | 372.105 | C16H20O10 | 1.67 | Dihydroferulic acid 4-O-glucuronide |
| 41 | 9.166 | 354.0948 | C16H18O9 | 0.92 | 5-Caffeoylquinic acid |
| 42 | 9.263 | 772.205 | C33H40O21 | 1.58 | Kaempferol-3,7,4′-triglucoside |
| 43 | 9.309 | 290.0785 | C15H14O6 | 1.81 | Epifisetinidol-4α-ol |
| 44 | 9.322 | 452.1312 | C21H24O11 | 1.47 | 7-Hydroxybutylidenephthalide 7-(6-malonylglucoside) |
| 45 | 9.364 | 692.1939 | C32H36O17 | 1.97 | 6-Methoxykaempferol 3,7-bis(3-acetylrhamnoside) |
| 46 | 9.379 | 390.1159 | C16H22O11 | 0.74 | Deacetyl asperulosidic acid |
| 47 | 9.435 | 426.173 | C17H30O12 | 1.64 | 3-Methylbutanoyl-6-O-α-d-glucopyranosyl-β-d-fructofuranoside |
| 48 | 9.522 | 625.1409 | C27H29O17 | −0.76 | Cyanidin 3-(2″-glucuronosylglucoside) |
| 49 | 9.533 | 306.0732 | C15H14O7 | 2.57 | Epigallocatechin |
| 50 | 9.576 | 326.0997 | C15H18O8 | 1.48 | o-Coumaric acid-β-d-glucoside |
| 51 | 9.584 | 392.1312 | C16H24O11 | 1.59 | Caryoptosidic acid |
| 52 | 9.796 | 374.1207 | C16H22O10 | 1.54 | Swertiamarin |
| 53 | 9.81 | 330.0944 | C14H18O9 | 2.02 | 3′-Glucosyl-2′,4′,6′-trihydroxyacetophenone |
| 54 | 9.841 | 742.231 | C33H42O19 | 1.43 | Naringin 4′-glucoside |
| 55 | 9.91 | 722.1473 | C35H30O17 | 1.43 | Thonningianin B |
| 56 | 9.976 | 460.1575 | C20H28O12 | 1.27 | Methyl salicylate O-[rhamnosyl-(1→6)-glucoside] |
| 57 | 9.977 | 528.1446 | C30H24O9 | −4.83 | Mahuangnin D |
| 58 | 10.129 | 756.2101 | C33H40O20 | 1.64 | Kaempferol 3-rutinoside-4′-glucoside |
| 59 | 10.14 | 316.1152 | C14H20O8 | 1.80 | 3,4-Dihydroxyphenylethyl alcohol glucoside |
| 60 | 10.196 | 178.0262 | C9H6O4 | 2.35 | Aesculetin |
| 61 | 10.219 | 326.1002 | C15H18O8 | −0.06 | cis-β-d-Glucosyl-2-hydroxycinnamate |
| 62 | 10.329 | 866.2047 | C45H38O18 | 1.34 | Cinnamtannin A1 |
| 63 | 10.4 | 402.152 | C18H26O10 | 1.41 | Benzyl O-[arabinofuranosyl-(1→6)-glucoside] |
| 64 | 10.401 | 516.1443 | C29H24O9 | −4.35 | Thelephantin A |
| 65 | 10.403 | 578.142 | C30H26O12 | 0.71 | Apigenin 7-(4″-E-p-coumarylglucoside) |
| 66 | 10.566 | 430.1468 | C19H26O11 | 1.58 | Bungeiside C |
| 67 | 10.609 | 566.1839 | C23H34O16 | 1.32 | 7-Glucosyl-11-methylodeoside |
| 68 | 10.635 | 1154.2717 | C60H50O24 | −2.14 | Cinnamtannin A2 |
| 69 | 10.759 | 386.121 | C17H22O10 | 0.78 | 3′-O-β-glucopyranosyl plumbagic acid |
| 70 | 10.796 | 458.1417 | C20H26O12 | 1.64 | 7-Hydroxy-4-methylphthalide O-[arabinosyl- (1→6)-glucoside] |
| 71 | 10.882 | 886.2728 | C39H50O23 | 1.67 | Reiniose E |
| 72 | 10.882 | 886.2728 | C16H18O8 | 1.4 | 1,2,4-Trihydroxynaphthalene-4-glucoside |
| 73 | 10.986 | 388.1364 | C17H24O10 | 1.35 | Geniposide |
| 74 | 11.042 | 490.1681 | C21H30O13 | 1.15 | Phloroacetophenone 6′-[xylosyl-(1→6)- glucoside] |
| 75 | 11.057 | 630.1941 | C31H34O14 | 1.23 | (R)-Rutaretin 1′-(6″-sinapoylglucoside) |
| 76 | 11.156 | 390.1522 | C17H26O10 | 0.98 | Todatriol glucoside |
| 77 | 11.161 | 386.1935 | C19H30O8 | 0.93 | Roseoside |
| 78 | 11.276 | 630.1943 | C31H34O14 | 0.87 | (R)-Rutaretin 1′-(6″-sinapoylglucoside) |
| 79 | 11.345 | 520.1785 | C22H32O14 | 1.30 | Swertiapunimarin |
| 80 | 11.372 | 590.1388 | C38H22O7 | −3.75 | 4′-Hydroxyanigorootin |
| 81 | 11.372 | 582.1731 | C30H30O12 | 1.15 | Epicatechin 3-O-(3-trans-cinnamoyl-β-d-allopyranoside) |
| 82 | 11.392 | 307.0686 | C14H13NO7 | 1.82 | Lycoricidinol |
| 83 | 11.398 | 246.0886 | C14H14O4 | 2.65 | Columbianetin |
| 84 | 11.408 | 648.2043 | C31H36O15 | 1.73 | Embigenin 2″-(2′″-acetylrhamnoside) |
| 85 | 11.412 | 404.1316 | C17H24O11 | 0.73 | Secoxyloganin |
| 86 | 11.425 | 704.2306 | C34H40O16 | 1.49 | Amorphigenin O-vicianoside |
| 87 | 11.501 | 462.1724 | C20H30O12 | 2.87 | Verbasoside |
| 88 | 11.641 | 344.1464 | C16H24O8 | 1.95 | Dihydroconiferin |
| 89 | 11.665 | 461.1524 | C19H27NO12 | 2 | 4-(2-Nitroethyl)phenyl primeveroside |
| 90 | 11.668 | 444.1626 | C20H28O11 | 1.23 | Sibiricaphenone |
| 91 | 11.734 | 450.1158 | C21H22O11 | 0.90 | Sinensin |
| 92 | 11.882 | 864.1891 | C45H36O18 | 1.21 | Pavetannin B2 |
| 93 | 12.058 | 356.1105 | C16H20O9 | 0.61 | Gentiopicrin |
| 94 | 12.5 | 434.0844 | C20H18O11 | 1.20 | Quercetin 3-β-l-arabinopyranoside |
| 95 | 12.539 | 400.1363 | C18H24O10 | 1.51 | 3′-O-β-Glucopyranosyl plumbagic acid methyl ester |
| 96 | 12.788 | 372.1049 | C16H20O10 | 1.98 | Veranisatin C |
| 97 | 12.807 | 404.1315 | C17H24O11 | 0.79 | Theviridoside |
| 98 | 12.867 | 370.1256 | C17H22O9 | 2.07 | Perilloside E |
| 99 | 12.991 | 414.1517 | C19H26O10 | 2.18 | Ptelatoside A |
| 100 | 13.052 | 394.1832 | C17H30O10 | 1.67 | cis-3-Hexenyl β-primeveroside |
| 101 | 13.102 | 514.194 | C26H30N2O9 | 2.28 | 10-Hydroxystrictosamide |
| 102 | 13.247 | 346.1623 | C16H26O8 | 1.27 | Villoside |
| 103 | 13.266 | 739.2082 | C33H39O19 | 0.46 | Malvidin 3-glucoside-5-(6″-malonylglucoside) |
| 104 | 13.381 | 224.1405 | C13H20O3 | 3.14 | Vomifoliol |
| 105 | 13.966 | 476.1885 | C21H32O12 | 1.75 | Kanokoside A |
| 106 | 14.095 | 368.11 | C17H20O9 | 1.99 | 3-O-Caffeoyl-4-O-methylquinic acid |
| 107 | 14.375 | 436.1726 | C22H28O9 | 1.79 | Icaride A2 |
| 108 | 14.491 | 740.173 | C39H32O15 | 1.55 | 3″,6″-Di-O-p-coumaroyltrifolin |
| 109 | 15.137 | 384.1414 | C18H24O9 | 1.76 | Tenuifoliside D |
| 110 | 15.901 | 516.2101 | C26H32N2O9 | 1.38 | Isomitraphyllic acid (16→1)-β-d-glucopyranosyl ester |
| 111 | 15.921 | 464.0948 | C21H20O12 | 1.39 | Robinetin 7-glucoside |
| 112 | 15.978 | 452.1102 | C24H20O9 | 1.07 | Epigallocatechin 3-O-p-coumarate |
| 113 | 16.185 | 348.1777 | C16H28O8 | 2.19 | Foeniculoside VIII |
| 114 | 16.234 | 304.0578 | C15H12O7 | 1.6 | 2′,3,5,6′,7-Pentahydroxyflavanone |
| 115 | 16.288 | 594.1582 | C27H30O15 | 0.4 | Isoorientin 7-O-rhamnoside |
| 116 | 16.29 | 582.194 | C27H34O14 | 1.45 | 10-Acetoxyligustroside |
| 117 | 16.296 | 1064.2994 | C48H56O27 | 1.44 | Capilliposide I |
| 118 | 16.329 | 464.2259 | C21H36O11 | −0.22 | Linalool 3,7-oxide β-primeveroside |
| 119 | 16.334 | 460.1583 | C20H28O12 | −0.57 | Aplopaeonoside |
| 120 | 16.335 | 576.1261 | C30H24O12 | 1.17 | Epicatechin-(2β→5,4β→6)-entepicatechin |
| 121 | 16.343 | 592.1571 | C31H28O12 | 1.62 | 8,8′-Methylenebiscatechin |
| 122 | 16.366 | 552.1838 | C26H32O13 | 0.89 | (Z)-Resveratrol 3,4′-diglucoside |
| 123 | 16.417 | 754.1892 | C40H34O15 | 0.71 | Chrysoeriol 7-(3″,6″-di-(E)-p-coumaroylglucoside) |
| 124 | 16.424 | 522.162 | C25H30O12 | 22.4 | Melampodinin |
| 125 | 16.435 | 484.1363 | C25H24O10 | 1.31 | Silidianin |
| 126 | 16.479 | 518.1992 | C23H34O13 | 1.37 | Jioglutoside B |
| 127 | 16.487 | 612.2029 | C28H36O15 | 4.1 | Neohesperidin dihydrochalcone |
| 128 | 16.518 | 410.1578 | C20H26O9 | −0.32 | Yadanziolide C |
| 129 | 16.525 | 448.1001 | C21H20O11 | 1.03 | Petunidin-3-O-arabinoside |
| 130 | 16.55 | 1048.3048 | C48H56O26 | 1.11 | Quercetin 3-rhamnosyl-(1→6)-[rhamnosyl-(1→2)-(3″-(E)-p-coumaroylgalactoside)]-7-Rhamnoside |
| 131 | 16.575 | 520.1938 | C26H32O11 | 1.18 | Tetracentronside B |
| 132 | 16.621 | 580.2147 | C28H36O13 | 1.58 | (+)-7-epi-Syringaresinol 4′-glucoside |
| 133 | 16.655 | 420.1634 | C18H28O11 | −0.61 | Lamioside |
| 134 | 16.663 | 412.1988 | C23H28N2O5 | 2.52 | Reserpiline |
| 135 | 16.766 | 528.1625 | C27H28O11 | 1.34 | Tremulacin |
| 136 | 16.821 | 548.2251 | C28H36O11 | 1.19 | Bruceantin |
| 137 | 16.822 | 504.1839 | C22H32O13 | 0.84 | (S)-Multifidol 2-[apiosyl-(1→6)-glucoside] |
| 138 | 16.85 | 448.1365 | C22H24O10 | 1.04 | Licoagroside D |
| 139 | 16.927 | 436.1155 | C24H20O8 | 0.76 | (−)-Epigallocatechin 3-cinnamate |
| 140 | 16.934 | 498.1517 | C26H26O10 | 1.79 | Dukunolide B |
| 141 | 16.946 | 772.1847 | C36H36O19 | 0.49 | Cyanidin 3-(2-glucosyl-6-caffeoylglucoside) |
| 142 | 16.949 | 902.2468 | C42H46O22 | 1.4 | Kaempferol 3-caffeylrobinobioside-7-Rhamnoside |
| 143 | 16.978 | 342.0732 | C18H14O7 | 2.09 | Iriskashmirianin |
| 144 | 16.998 | 828.4492 | C42H68O16 | 1.9 | Centellasaponin B |
| 145 | 17.017 | 436.1362 | C21H24O10 | 1.75 | Phlorhizin |
| 146 | 17.025 | 317.0665 | C16H13O7 | −1.09 | Petunidin |
| 147 | 17.029 | 942.4798 | C47H74O19 | 2.77 | Phytolaccoside I |
| 148 | 17.052 | 466.1259 | C25H22O9 | 1.12 | Silymonin |
| 149 | 17.067 | 756.1896 | C36H36O18 | 0.68 | Kaempferol 3-glucoside-7-p-coumarylglucoside |
| 150 | 17.11 | 856.4445 | C43H68O17 | 1.35 | Laxogenin 3-O-{O-β-d-xylopyranosyl-(1→4)-O-[α-l-arabinopyranosyl-(1→6)-β-d-glucopyranoside |
| 151 | 17.114 | 766.4492 | C41H66O13 | 1.48 | Soyasaponin IV |
| 152 | 17.135 | 532.2311 | C28H36O10 | −0.47 | Nomilinic acid |
| 153 | 17.232 | 812.4543 | C42H68O15 | 1.86 | Kudzusaponin SA1 |
| 154 | 17.237 | 956.4974 | C48H76O19 | 0.76 | Chikusetsusaponin V |
| 155 | 17.293 | 550.1678 | C26H30O13 | 1.45 | Neoliquiritin 2″-apioside |
| 156 | 17.331 | 194.0574 | C10H10O4 | 2.81 | Isoferulic acid |
| 157 | 17.356 | 780.4285 | C41H64O14 | 1.37 | Cynarasaponin F |
| 158 | 17.411 | 808.4241 | C42H64O15 | 0.58 | Licoricesaponin B2 |
| 159 | 17.411 | 926.4899 | C47H74O18 | −2.55 | Tarasaponin I |
| 160 | 17.425 | 842.4645 | C43H70O16 | 2.28 | Aspafilioside B |
| 161 | 17.487 | 750.4547 | C41H66O12 | 1.03 | Scabioside B |
| 162 | 17.514 | 666.3966 | C36H58O11 | 1.98 | Arjunglucoside I |
| 163 | 17.548 | 878.3963 | C44H62O18 | −3.03 | Hancoside A |
| 164 | 17.559 | 638.3654 | C34H54O11 | 1.91 | 26-Glucosyl-1,3,11,22-tetrahydroxyergosta-5,24-dien-26-oate |
| 165 | 17.619 | 422.2144 | C19H34O10 | 1.84 | 1-Octen-3-yl primeveroside |
| 166 | 17.656 | 396.2142 | C21H32O7 | 1.64 | Isopetasoside |
| 167 | 17.748 | 288.0622 | C15H12O6 | 4.12 | 3,4,2′,4′,α-Pentahydroxychalcone |
| 168 | 17.748 | 446.2143 | C21H34O10 | 1.95 | Dendroside G |
| 169 | 17.796 | 302.0416 | C15H10O7 | 3.59 | Quercetin |
| 170 | 17.8 | 396.2044 | C23H28N2O4 | 1.38 | Echitovenine |
| 171 | 17.8 | 530.2511 | C29H38O9 | 0.99 | Angeloylgomisin Q |
| 172 | 17.813 | 516.2356 | C28H36O9 | 0.73 | Kupitengester 4 |
| 173 | 17.822 | 398.2201 | C23H30N2O4 | 1.04 | Mitragynine |
| 174 | 17.918 | 648.3862 | C36H56O10 | 1.73 | 2α-Hydroxygypsogenin 3-O-β-d-glucoside |
| 175 | 17.992 | 792.4281 | C42H64O14 | 1.86 | Mabioside C |
| 176 | 18.112 | 680.3765 | C36H56O12 | 0.99 | Trachelosperoside A1 |
| 177 | 18.133 | 208.0731 | C11H12O4 | 2.28 | 6-Methoxymellein |
| 178 | 18.443 | 650.4016 | C36H58O10 | 2.16 | Lucyoside N |
| 179 | 18.577 | 450.2607 | C25H38O7 | 2.27 | Laserpitin |
| 180 | 18.82 | 632.3912 | C36H56O9 | 1.99 | Lucyoside K |
| 181 | 18.821 | 796.459 | C42H68O14 | 2.35 | Saikosaponin L |
| 182 | 18.868 | 192.0781 | C11H12O3 | 2.88 | (R)-Shinanolone |
| 183 | 19.065 | 250.1198 | C14H18O4 | 2.69 | Helinorbisabone |
| 184 | 19.189 | 252.0992 | C13H16O5 | 2.30 | Methyl 3,4,5-trimethoxycinnamate |
| 185 | 19.514 | 594.136 | C30H26O13 | 2.22 | Tiliroside |
| 186 | 19.597 | 256.0728 | C15H12O4 | 2.92 | Isoliquiritigenin |
| 187 | 19.677 | 222.1611 | C14H22O2 | 3.96 | Isokobusone |
| 188 | 19.678 | 294.1826 | C17H26O4 | 1.64 | Myrsinone |
| 189 | 20.303 | 488.3487 | C30H48O5 | 2.97 | Uncaric acid |
| 190 | 20.674 | 276.1716 | C17H24O3 | 3.36 | Moxartenone |
| 191 | 20.849 | 292.1665 | C17H24O4 | 3.2 | 6-Hydroxyshogaol |
| 192 | 21.329 | 234.1613 | C15H22O2 | 2.88 | Sclerosporin |
| 193 | 23.648 | 430.2005 | C24H30O7 | −3.20 | Schisanlignone A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Fei, S.; Olatunji, O.J. LC/ESI/TOF-MS Characterization, Anxiolytic and Antidepressant-like Effects of Mitragyna speciosa Korth Extract in Diabetic Rats. Molecules 2022, 27, 2208. https://doi.org/10.3390/molecules27072208
Chen L, Fei S, Olatunji OJ. LC/ESI/TOF-MS Characterization, Anxiolytic and Antidepressant-like Effects of Mitragyna speciosa Korth Extract in Diabetic Rats. Molecules. 2022; 27(7):2208. https://doi.org/10.3390/molecules27072208
Chicago/Turabian StyleChen, Lin, Shizao Fei, and Opeyemi Joshua Olatunji. 2022. "LC/ESI/TOF-MS Characterization, Anxiolytic and Antidepressant-like Effects of Mitragyna speciosa Korth Extract in Diabetic Rats" Molecules 27, no. 7: 2208. https://doi.org/10.3390/molecules27072208
APA StyleChen, L., Fei, S., & Olatunji, O. J. (2022). LC/ESI/TOF-MS Characterization, Anxiolytic and Antidepressant-like Effects of Mitragyna speciosa Korth Extract in Diabetic Rats. Molecules, 27(7), 2208. https://doi.org/10.3390/molecules27072208