
Lowering the Intraocular Pressure in Rats and Rabbits by Cordyceps cicadae Extract and Its Active Compounds

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethanol Extract of C. cicadae Mycelium (CCME) Preparation
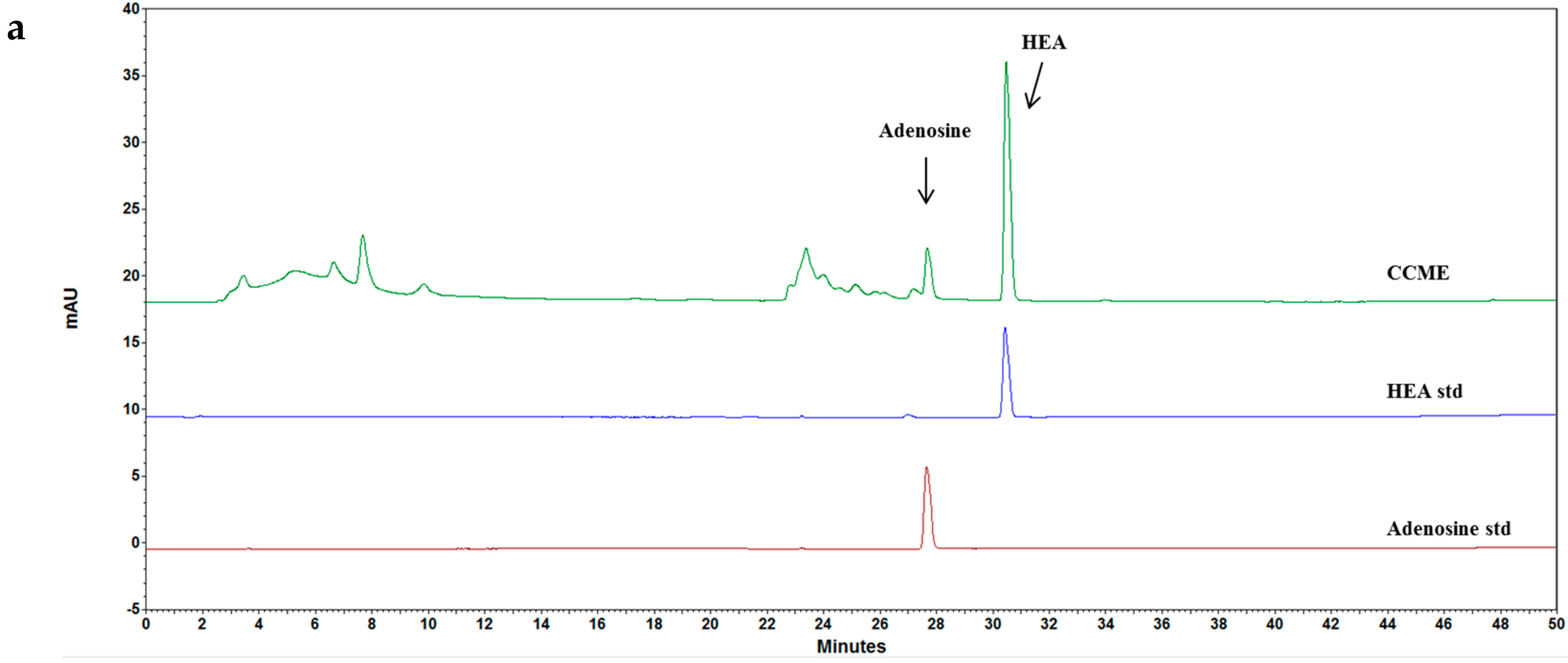
2.2. High-Performance Liquid Chromatography (HPLC) Analysis
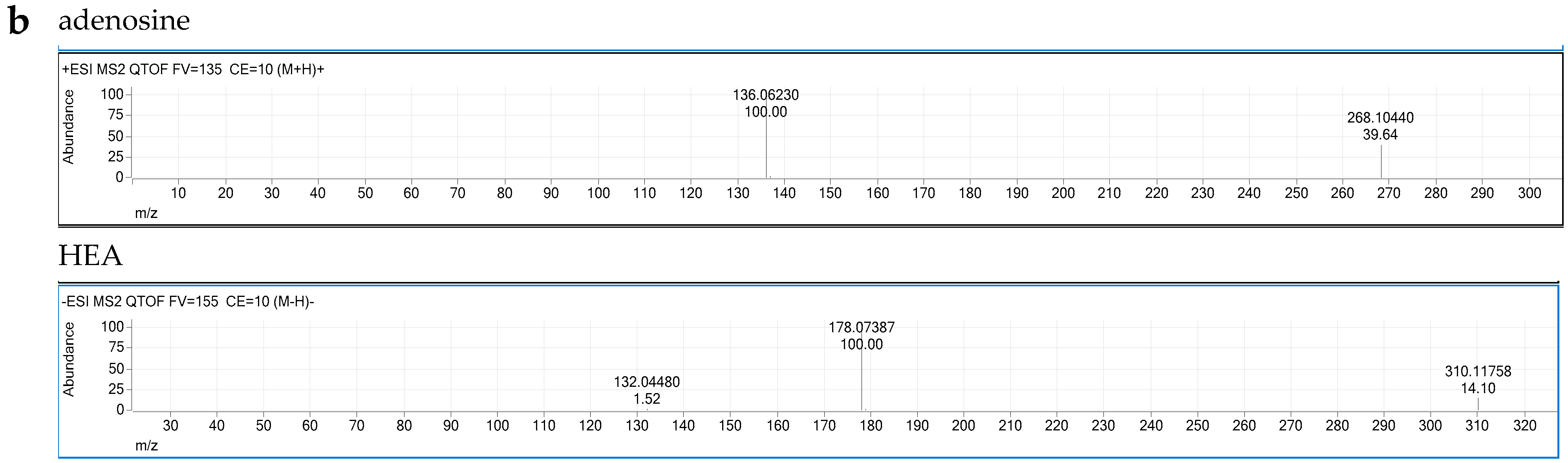
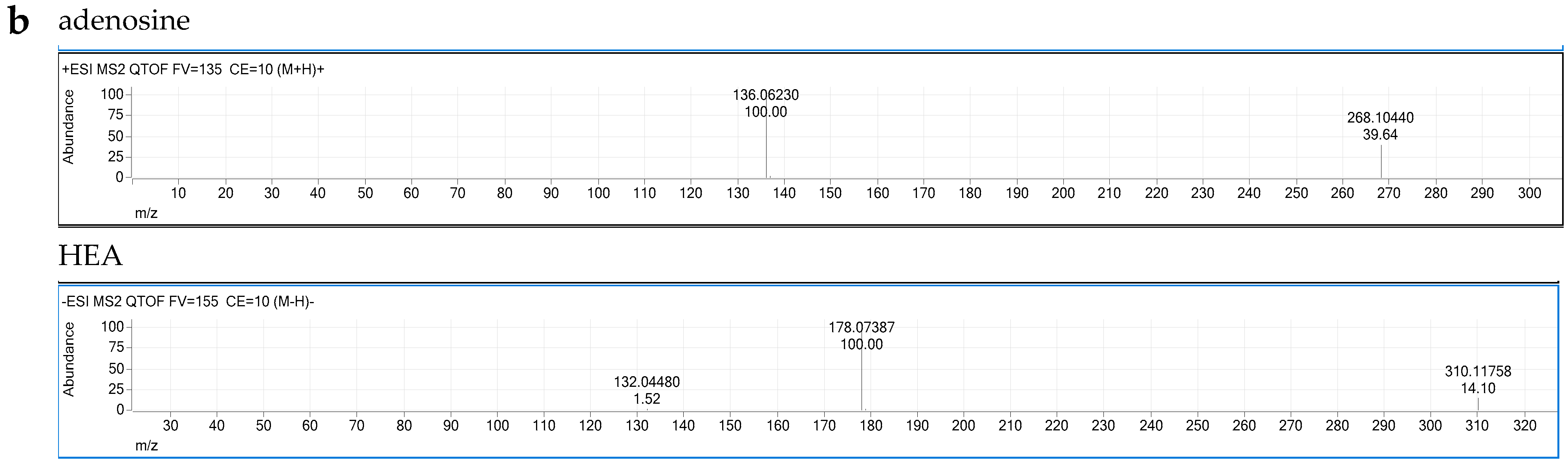
2.3. Liquid Chromatography–Quadrupole-Time of Flight Mass Spectrometer (LC–QTOF/MS) Analysis
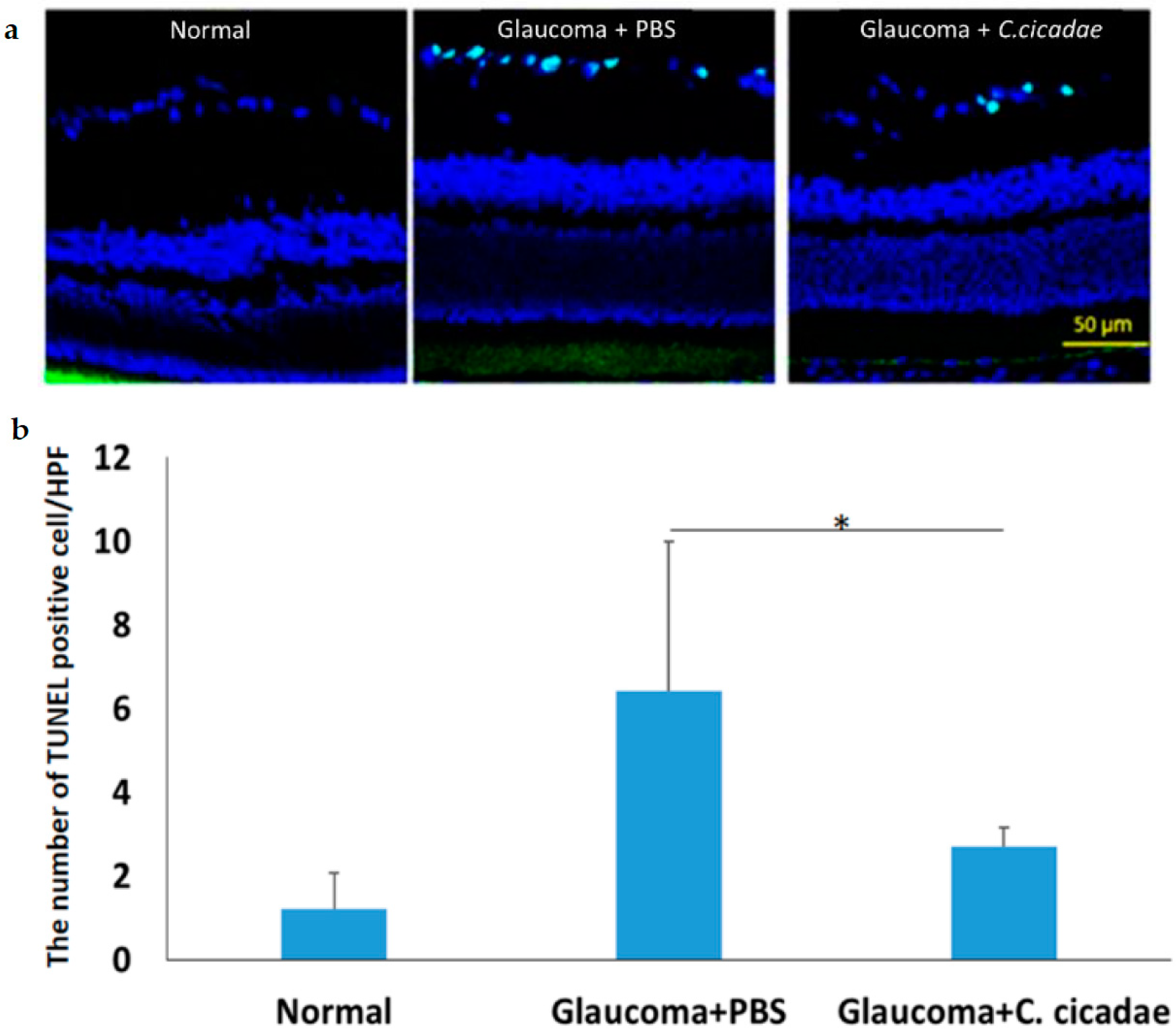
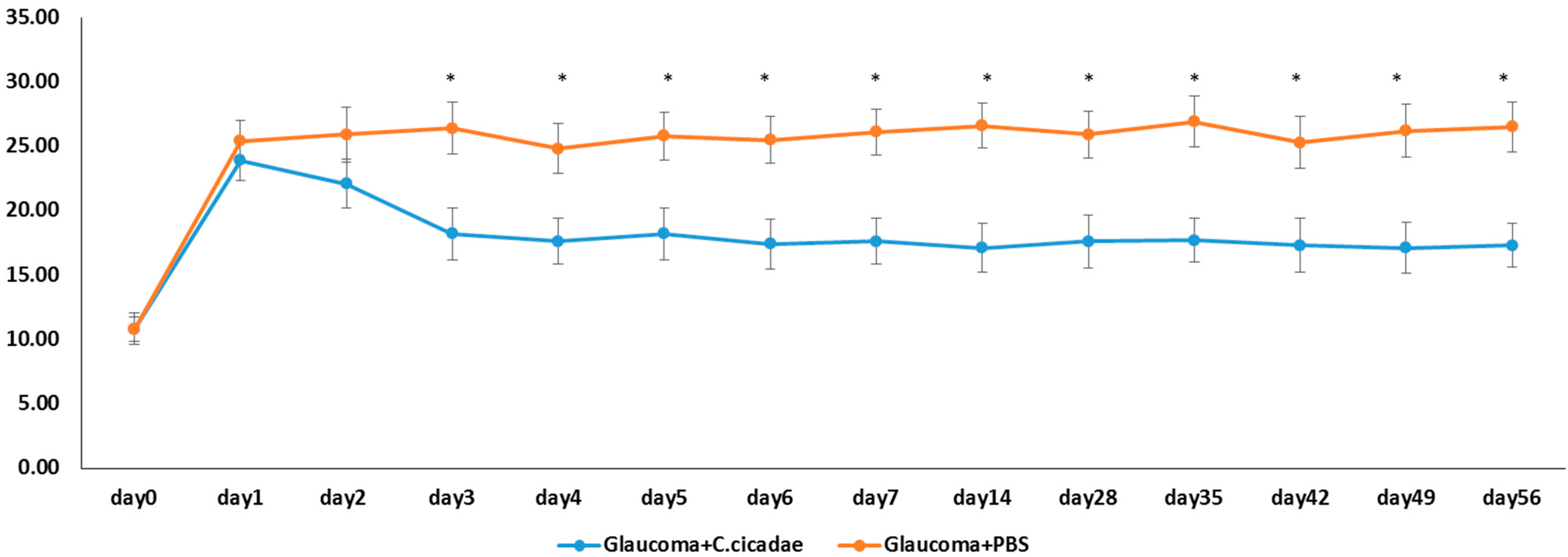
2.4. Microbead Occlusion in Rat Model
2.5. ROCK Kinase Assay
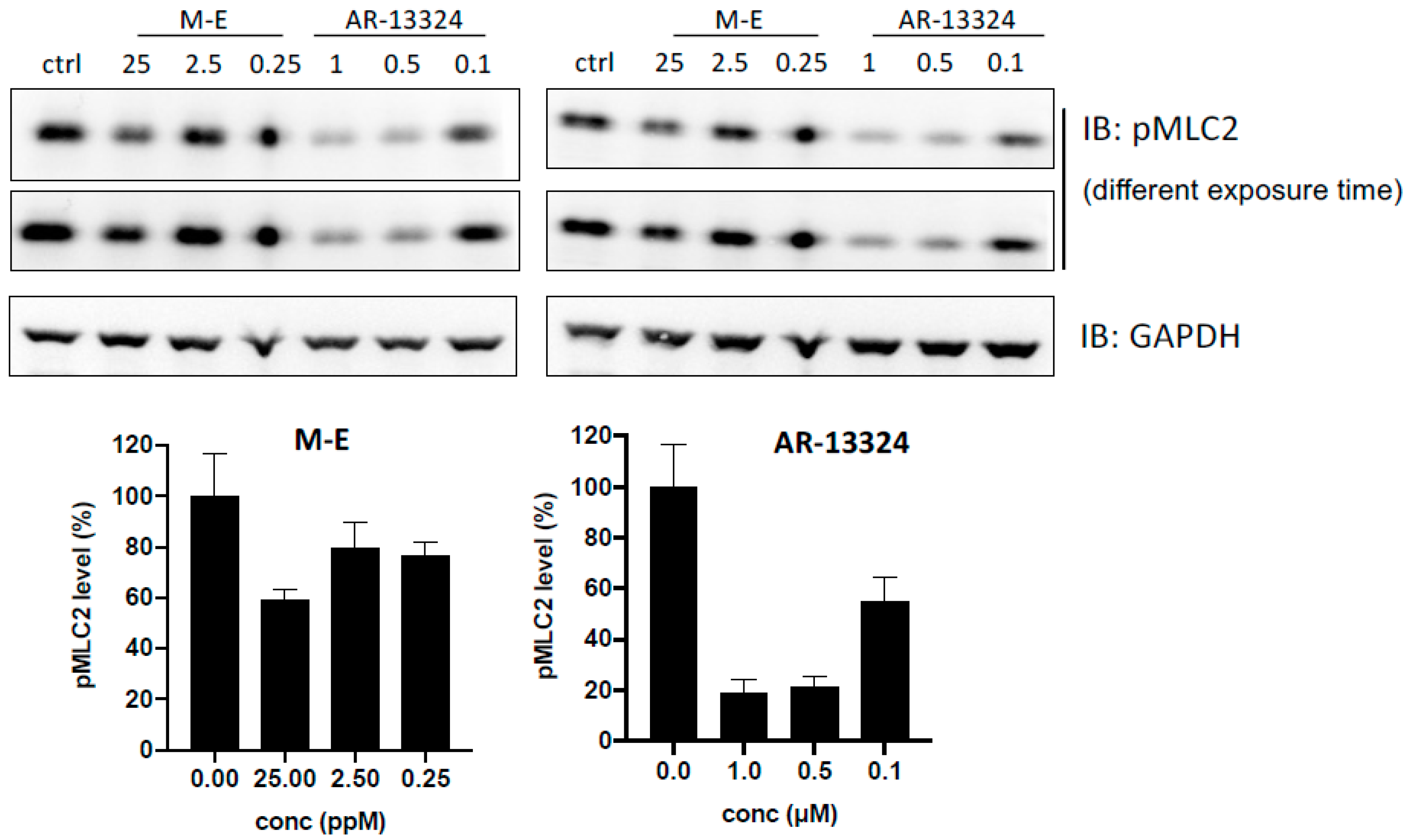
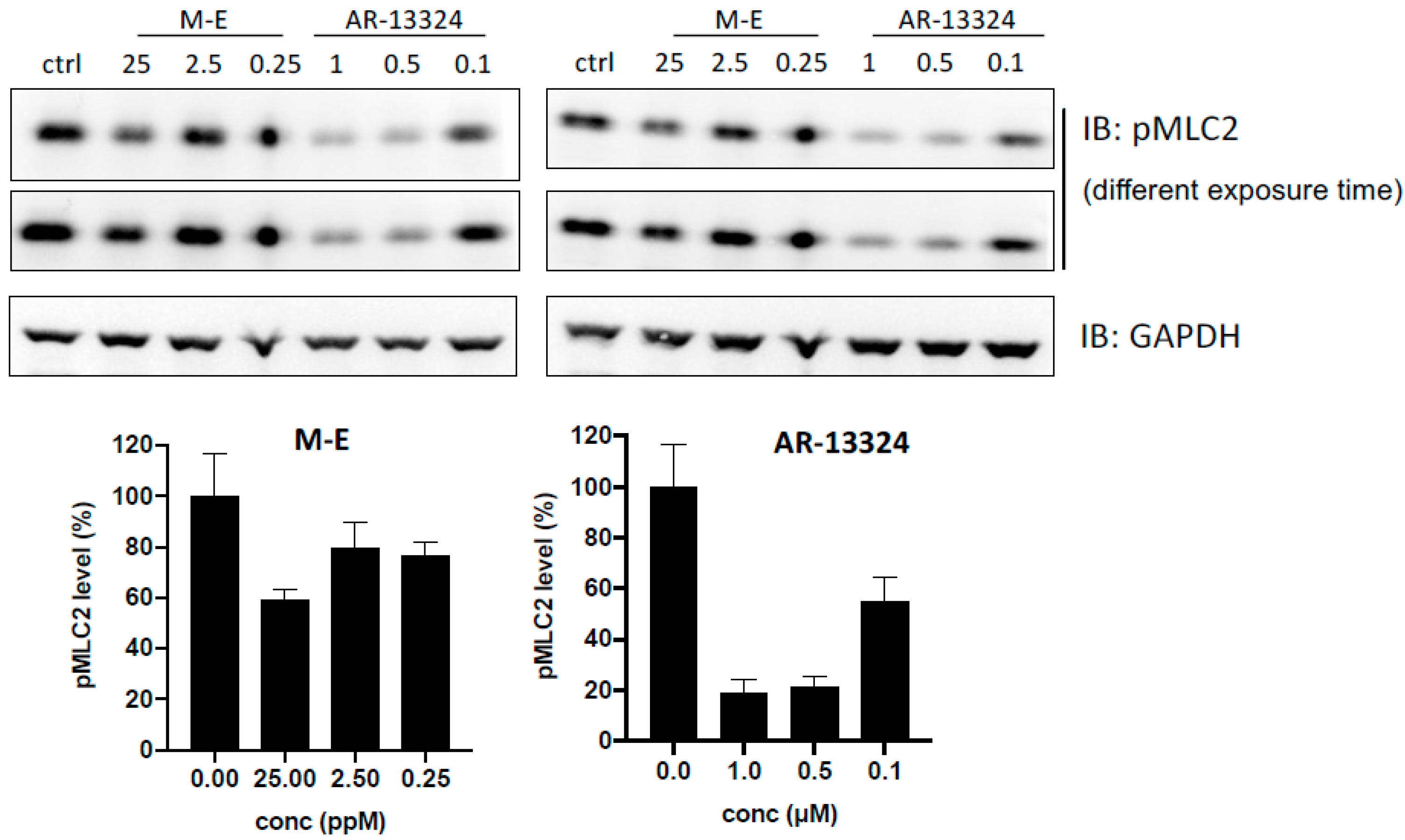
2.6. MLC Phosphorylation Inhibition (p MLC) Assay
2.7. Normal Rabbit Model
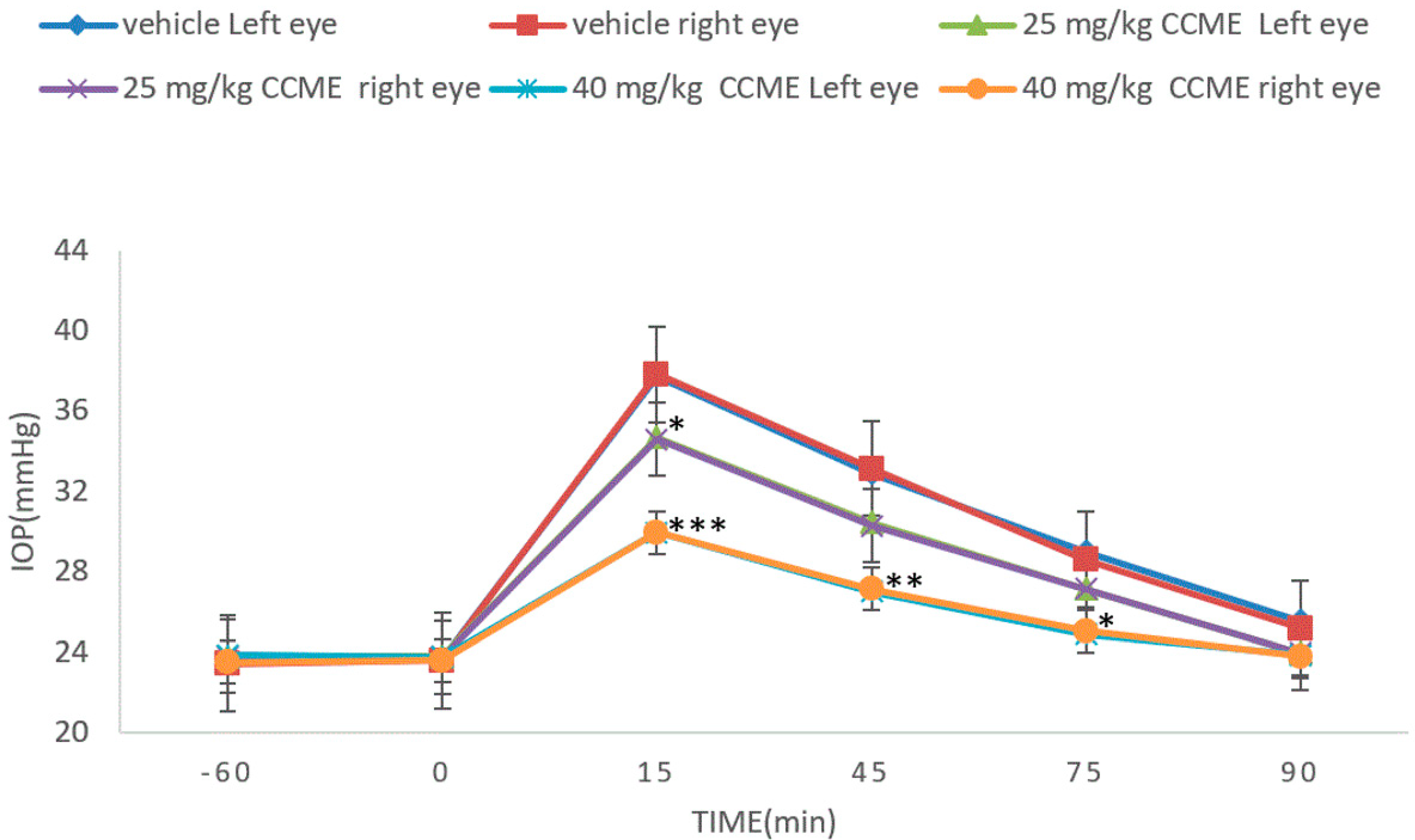
2.8. Dextrose-Induced Acute Glaucoma Model
3. Results
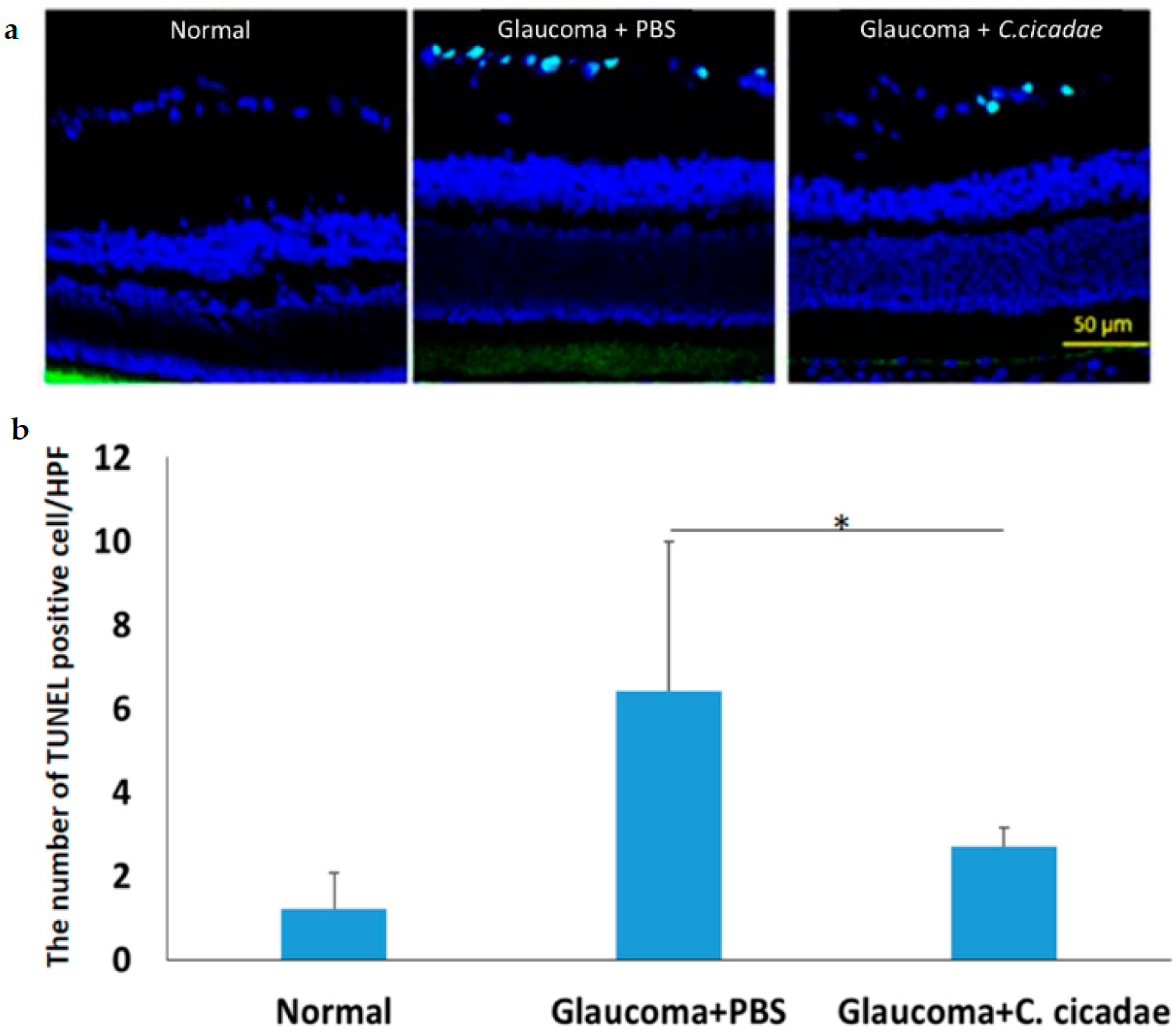
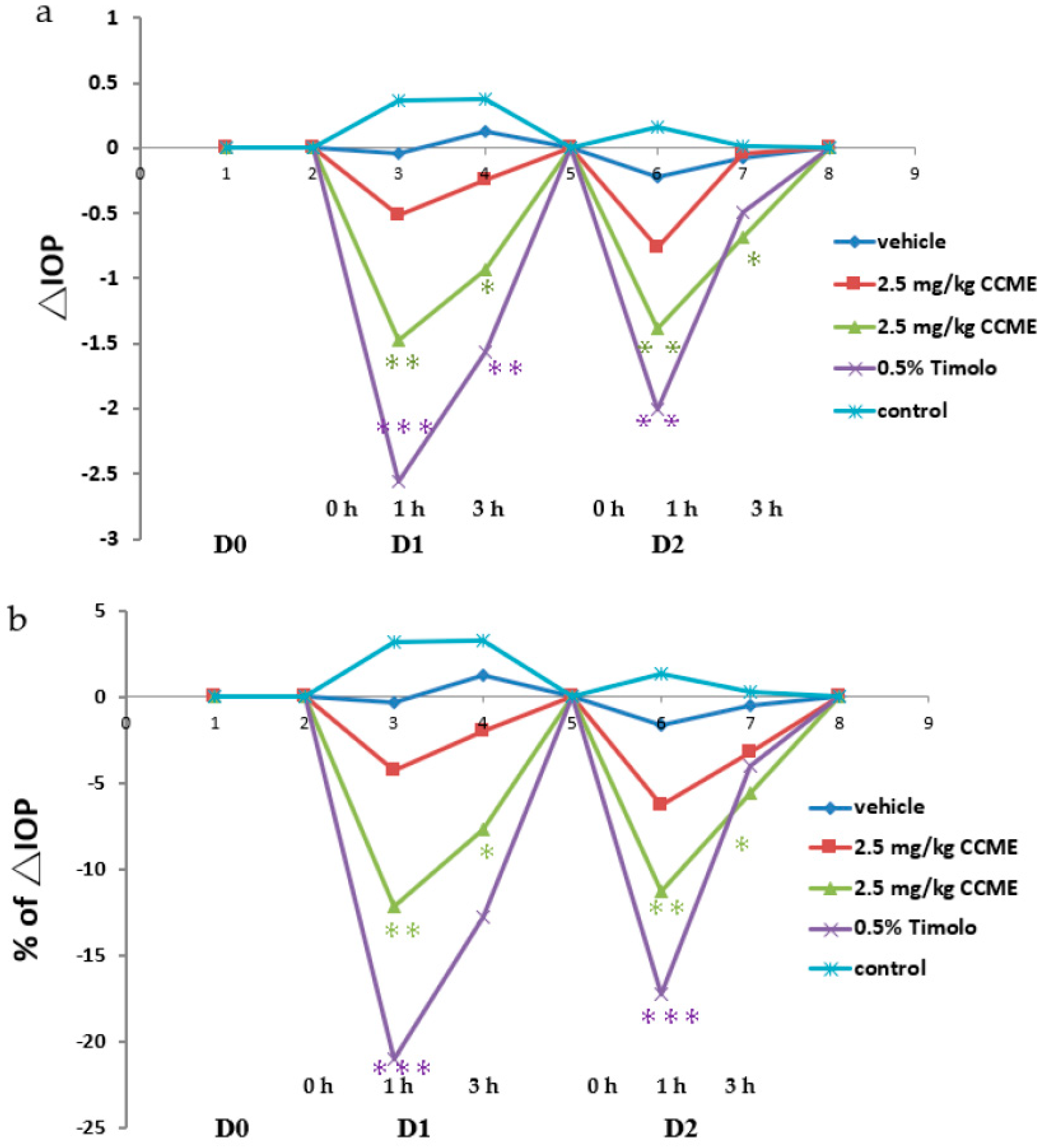
3.1. Glaucoma Model
3.2. ROCK Kinase Assay
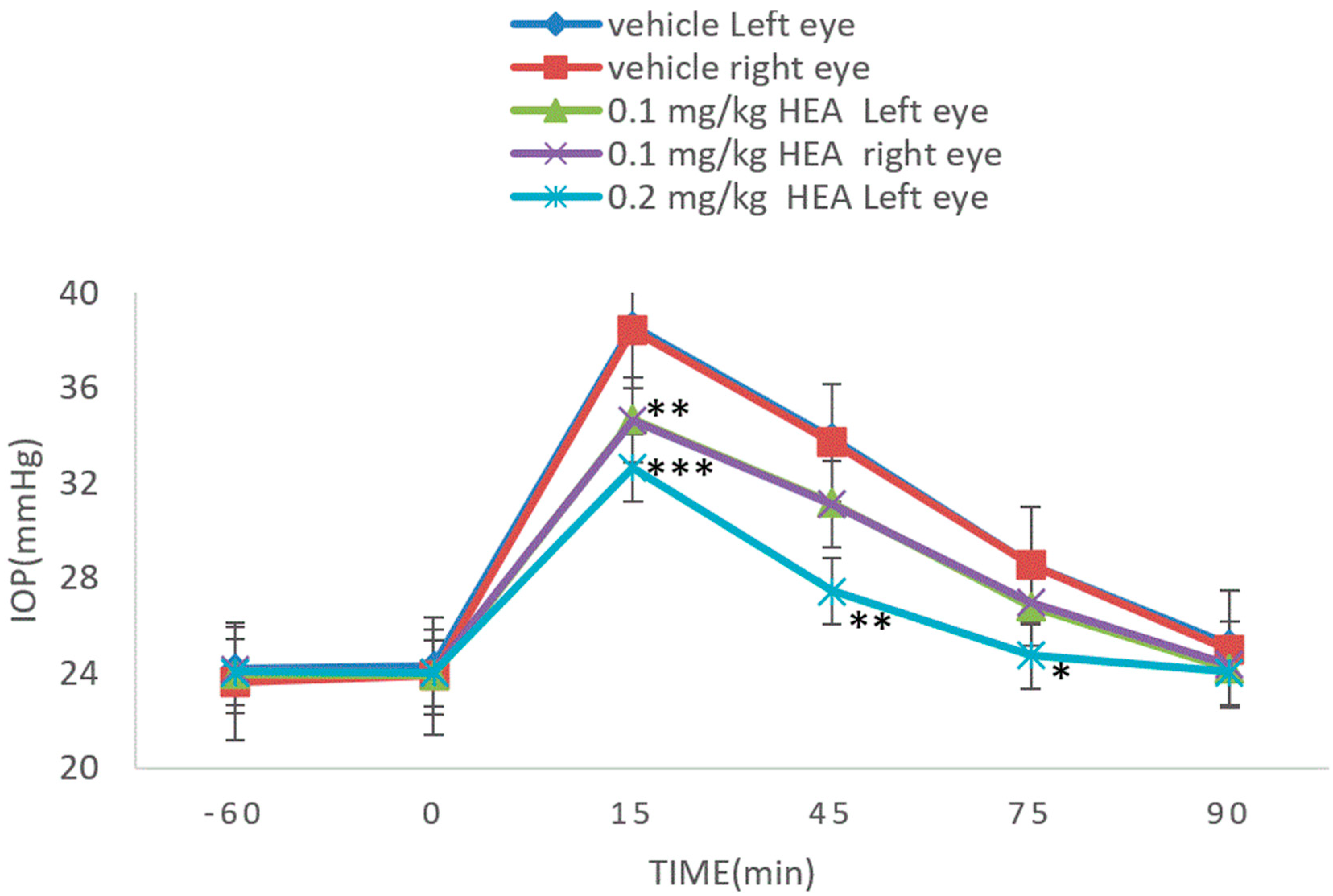
3.3. Normal Rabbit Model
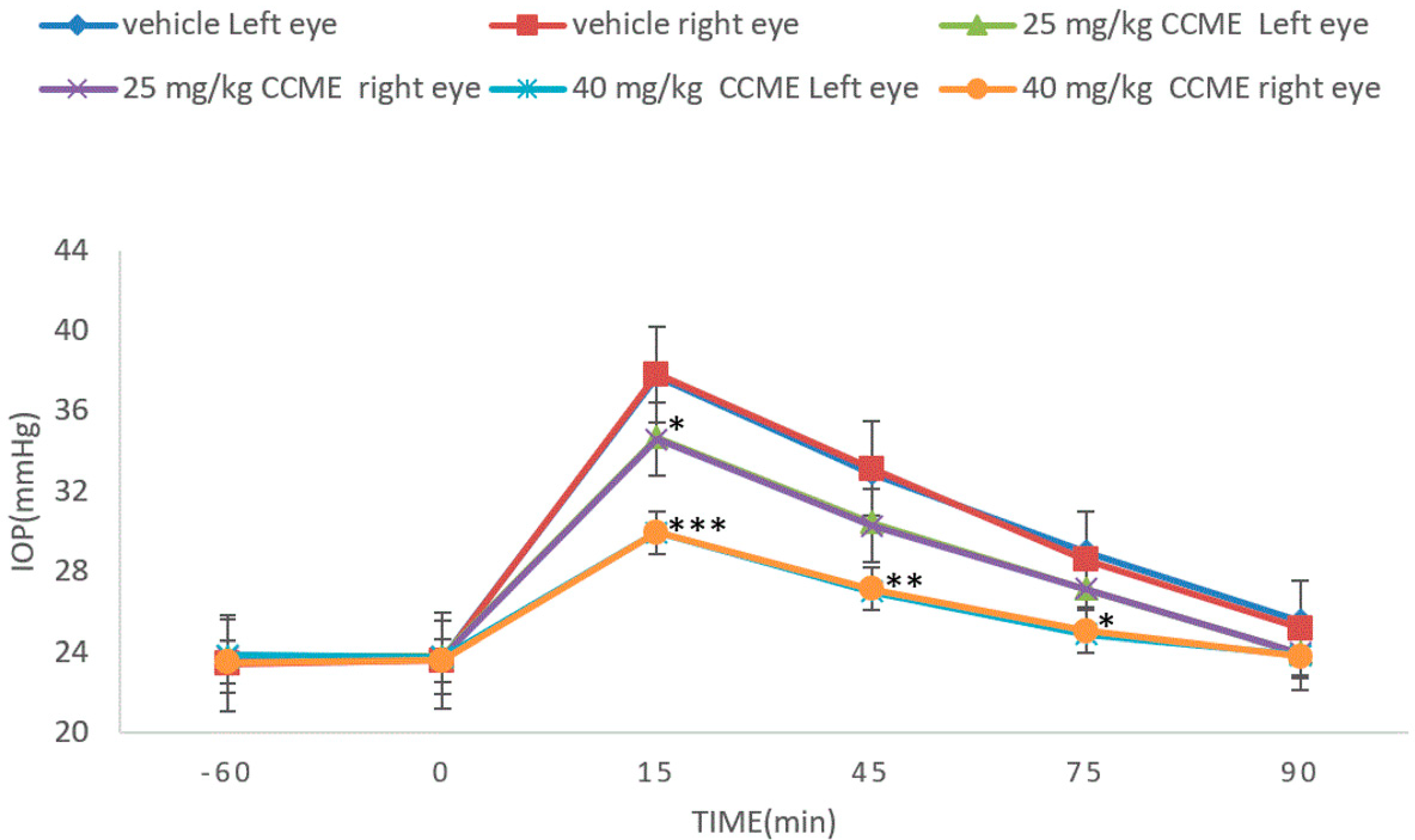
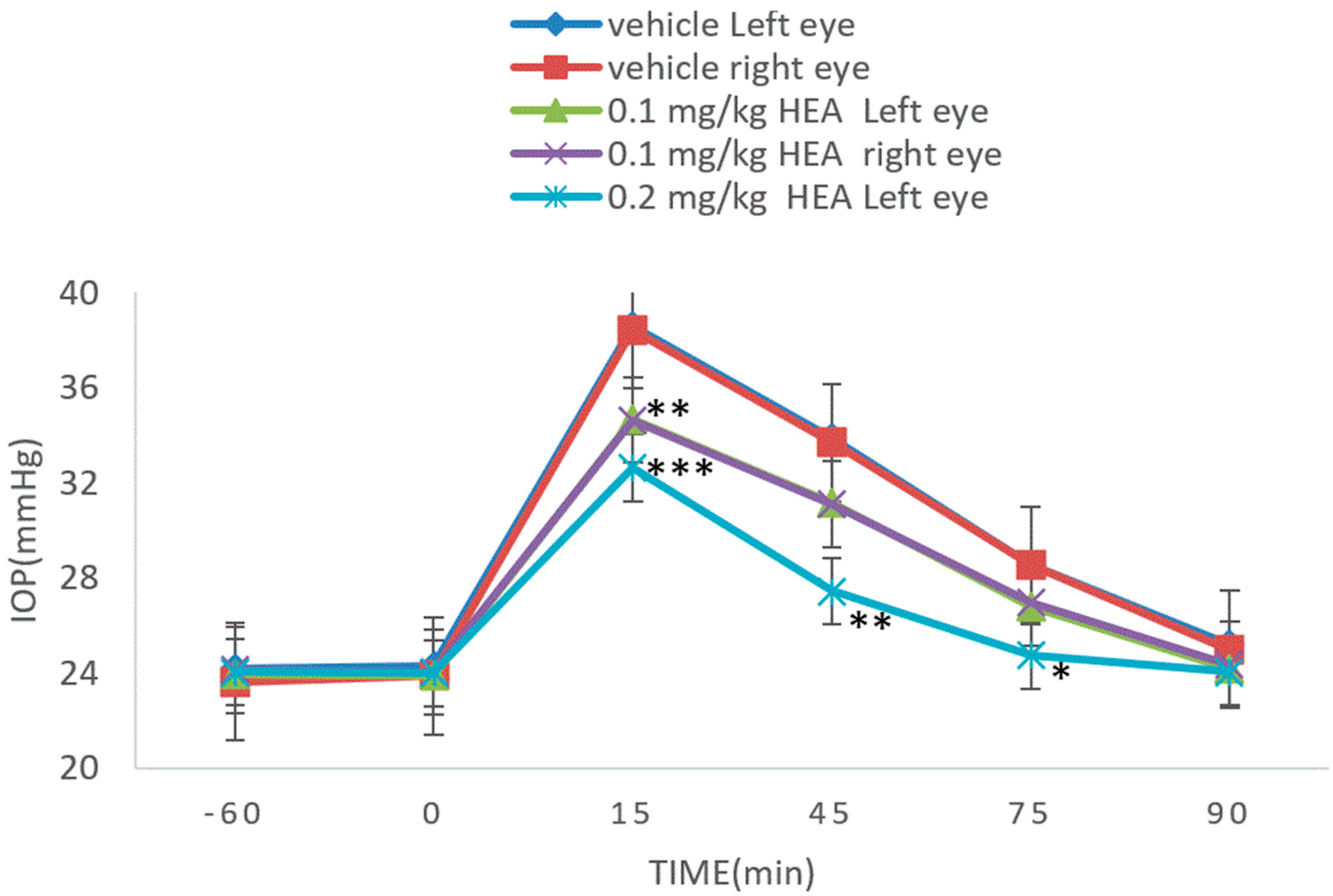
3.4. Dextrose-Induced Acute Glaucoma Model
3.5. CCME Active Components
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Yu, H.M.; Wang, B.S.; Huang, S.C.; Duh, P.D. Comparison of protective effects between cultured Cordyceps militaris and natural Cordyceps sinensis against oxidative damage. J. Agric. Food Chem. 2006, 54, 3132–3138. [Google Scholar] [CrossRef]
- Zhu, Y.; Yu, X.; Ge, Q.; Li, J.; Wang, D.; Wei, Y.; Ouyang, Z. Antioxidant and anti-aging activities of polysaccharides from Cordyceps cicadae. Int. J. Biol. Macromol. 2020, 15, 394–400. [Google Scholar] [CrossRef]
- Won, S.Y.; Park, E.H. Anti-inflammatory and related pharmacological activities of cultured mycelia and fruiting bodies of Cordyceps militaris. J. Ethnopharmacol. 2005, 96, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Weng, S.C.; Chou, C.J.; Lin, L.C.; Tsai, W.J.; Kuo, Y.C. Immunomodulatory functions of extracts from the Chinese medicinal fungus Cordyceps cicadae. J. Ethnopharmacol. 2002, 83, 79–85. [Google Scholar] [CrossRef]
- Zhang, Q.; Olatunji, O.J.; Chen, H.; Tola, A.; Oluwaniyi, O.-O. Evaluation of the Anti-Diabetic Activity of Polysaccharide from Cordyceps cicadae in Experimental Diabetic Rats. Chem. Biodivers. 2018, 15, e1800219. [Google Scholar] [CrossRef]
- Li, L.; Zhang, T.; Li, C.; Xie, L.; Li, N.; Hou, T.; Wang, Y.; Wang, B. Potential therapeutic effects of Cordyceps cicadae and Paecilomyces cicadae on adenine-induced chronic renal failure in rats and their phytochemical analysis. Drug Des. Dev. Ther. 2018, 13, 103–117. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-S.; Wang, X.; Feng, Z.; Cui, H.; Zhu, Z.; Xia, C.; Han, X.; Liu, W.J.; Liu, Y.N. Cordyceps cicadae Prevents Renal Tubular Epithelial Cell Apoptosis by Regulating the SIRT1/p53 Pathway in Hypertensive Renal Injury. Evid. Based Complement. Alternat. Med. 2020, 2020, 7202519. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Zhu, R.; Li, X.L.; Li, X.Y.; Shen, L.I.; Chen, Y.; Zhong, Y.F.; Deng, Y.Y. N6-(2-Hydroxyethyl) Adenosine From Cordyceps cicadae Ameliorates Renal Interstitial Fibrosis and Prevents Inflammation via TGF-β1/Smad and NF-κB Signaling Pathway. Front. Physiol. 2018, 9, 1229. [Google Scholar] [CrossRef]
- Wang, X.H.; Qin, A.O.; Xiao, F.; Olatunji, O.J.; Zhang, S.Y.; Pan, D.; Han, W.Z.; Wang, D.Q.; Ni, Y.H. N6-(2-hydroxyethyl)-adenosine from Cordyceps cicadae protects against diabetic kidney disease via alleviation of oxidative stress and inflammation. J. Food Biochem. 2019, 43, e12727. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Yeh, S.H.; Lin, T.W.; Chen, C.C.; Chen, C.S.; Kuo, C.F. A 90-Day Subchronic Toxicity Study of Submerged Mycelial Culture of Cordyceps cicadae (Ascomycetes) in Rats. Int. J. Med. Mushrooms 2015, 17, 771–781. [Google Scholar] [CrossRef]
- Hsu, J.H.; Yeh, S.H.; Huang, W.C.; Chen, C.C. Isolation, Identification and Genotoxicity Test of Cordyceps cicadae mycelium. J. Test. Qual. Assur. 2015, 4, 114–127. [Google Scholar]
- Jhou, B.Y.; Hsu, J.H.; Yeh, S.H.; Wang, Y.H.; Chen, C.C. A Subacute Toxicity Study of Cordyceps cicadae Mycelium in High-Glucose Diet-Fed LY Pigs. Hans. J. Food Nutr. Sci. 2016, 5, 13–20. [Google Scholar]
- Chen, C.C.; Wong, T.C.; Huang, C.L.; Jhou, B.Y.; Hsu, J.H.; Yeh, S.H.; Tsai, Y.T.; Yang, S.H. Clinical Evaluation of Blood Glucose Regulation and Safety of Cordyceps cicadae Mycelium. J. Food Nutr. Res. 2017, 5, 137–143. [Google Scholar]
- Li, I.C.; Hsu, J.H.; Lin, T.W.; Lin, W.H.; Chen, C.C. Prenatal developmental toxicity study of HEA-enriched Cordyceps cicadae mycelia in Sprague-Dawley rats. SDRP J. Food Sci. Technol. 2017, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.Y.; Hsu, J.H.; Wu, J.J.; Shih, C.H.; Chen, C.C. A Acute Toxicity Study of Cordyceps cicadae Mycelium in ICR Mice and SD Rats. Hans J. Food Nutr. Sci. 2017, 6, 96–105. [Google Scholar] [CrossRef]
- Tsai, Y.S.; Hsu, J.H.; David Lin, P.C.; Chang, H.H.; Chang, W.J.; Chen, Y.L.; Chen, C.C. Safety Assessment of HEA-Enriched Cordyceps cicadae Mycelium: A Randomized Clinical Trial. J. Am. Coll. Nutr. 2021, 40, 127–132. [Google Scholar] [CrossRef]
- Hsu, J.H.; Chen, C.C. Intraocular Pressure-Lowering Efficacy and Safety of Fermented Cordyceps cicadae Mycelia. Hans J. Food Nutr. Sci. 2021, 10, 33–39. [Google Scholar]
- Horng, C.T.; Yang, Y.L.; Chen, C.C.; Huang, Y.S.; Chen, C.; Chen, F.A. Intraocular pressure-lowering effect of Cordyceps cicadae mycelia extract in a glaucoma rat model. Int. J. Med. Sci. 2021, 18, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.I.; Hsu, J.H.; Li, T.J.; Yeh, S.H.; Chen, C.C. Safety assessment of HEA-enriched Cordyceps cicadae mycelia on the central nervous system (CNS), cardiovascular system, and respiratory system in ICR male mice. Food Sci. Nutr. 2021, 9, 4905–4915. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.U.; Chang, H.H.; Tang, U.J.; Chen, C.C.; Lee, Y.L.; David Lin, P.C. Fermented Cordyceps cicadae Mycelia Extracts Ameliorate Dry Eye Symptoms through Reduction of Cornea Epithelial Cell Apoptosis and Maintenance of Conjunctival Goblet Cells in a Mouse Dry Eye Model. Food Nutr. Res. 2017, 5, 320–330. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Albarral, J.A.; Ramírez, A.I.; De Hoz, R.; López-Villarín, N.; Salobrar-García, E.; López-Cuenca, I.; Licastro, E.; Inarejos-García, A.M.; Almodóvar, P.; Pinazo-Durán, M.D.; et al. Neuroprotective and Anti-Inflammatory Effects of a Hydrophilic Saffron Extract in a Model of Glaucoma. Int. J. Mol. Sci. 2019, 22, 4110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonomi, L.; Tomazzoli, L.; Jaria, D. An improved model of experimentally induced ocular hypertension in the rabbit. Investig. Ophthalmol. Vis. Sci. 1976, 15, 781–784. [Google Scholar]
- Tsai, R.K.; Chang, C.H.; Wang, H.Z. Neuroprotective effects of recombinant human granulocyte colony-stimulating factor (G-CSF) in neurodegeneration after optic nerve crush in rats. Exp. Eye Res. 2008, 87, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Albrecht-Küpper, B.E.; Leineweber, K.; Nell, P.G. Partial adenosine A1 receptor agonists for cardiovascular therapies. Purinergic Signal 2012, 8, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Varma, R.; Lee, P.; Glodberg, I.; Kotaks, S. An assessment of the health and economic burdens of glaucoma. Am. J. Opthalmol. 2011, 152, 515–522. [Google Scholar] [CrossRef] [Green Version]
- Kish, T. Old and New Drug Classes Expanding To Include Glaucoma Treatments. Pharm. Ther. 2018, 43, 489–502. [Google Scholar]
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucoma and projections ofglaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2089. [Google Scholar] [CrossRef]
- Bradley, J.M.; Vranka, J.; Colvis, C.M.; Conger, D.M.; Alexander, J.P.; Fisk, A.S.; Samples, J.R.; Acott, T.S. Effect of matrix metalloproteinase activity on outflow in perfused human organ culture. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2649–2658. [Google Scholar]
- Tian, B.; Geiger, B.; Epstein, D.-L.; Kaufman, P.-L. Cytoskeletal involvement in the regulation of aqueous humor outflow. Investig. Ophthalmol. Vis. Sci. 2000, 41, 619–623. [Google Scholar]
- Tian, B.; Gabelt, B.T.; Geiger, B.; Kaufman, P.L. The role of the actomyosin system in regulating trabecular fluid outflow. Exp. Eye Res. 2009, 88, 713–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Yang, Z.; Huang, W.C.; Luo, X. Adenosine, adenosine receptors and glaucoma: An updated overview. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2013, 1830, 2882–2890. [Google Scholar] [CrossRef]
- Jacobson, K.-A.; Tosh, D.K.; Jain, S.; Gao, Z.G. Historical and current adenosine receptor agonists in preclinical and clinical development. Front. Cell. Neurosci. 2019, 13, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, T.-W.; Crosson, C.E. Adenosine A1 receptor modulation of MMP-2 secretion by trabecular meshwork cells. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3016–3020. [Google Scholar]
- Shearer, T.; Crosson, C.E. Activation of extracellular signal-regulated kinase in trabecular meshwork cells. Exp. Eye Res. 2001, 73, 7325–7335. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Leung, C.T.; Peterson-Yantorno, K.; Stamer, W.D.; Mitchell, C.H.; Civan, M.M. Mechanisms of ATP release by human trabecular meshwork cells, the enabling step in purinergic regulation of aqueous humor outflow. J. Cell. Physiol. 2012, 227, 172–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, A.R.; Madeira, M.H.; Boia, R.; Aires, I.D.; Rodrigues-Neves, A.C.; Santos, P.F. Keep an eye on adenosine: Its role in retinal inflammation. Pharmacol. Ther. 2020, 210, 107513. [Google Scholar] [CrossRef]
- Hasko, G.; Linden, J.; Cronstein, B.; Pacher, P.A. Adenosine receptors: Therapeutic aspects for inflammatory and immune diseases. Nat. Rev. Drug Discov. 2008, 7, 759–770. [Google Scholar] [CrossRef]
- Fishman, P.; Bar-Yehuda, S.; Liang, B.T.; Jacobson, K.A. Pharmacological and therapeutic effects of A3 adenosine receptor agonists. Drug Discov. Today 2012, 17, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Crosson, C.E. Adenosine receptor activation modulates intraocular pressure in rabbits. J. Pharm. Exp. Ther. 1995, 273, 320–326. [Google Scholar]
- Crosson, C.E. Ocular hypotensive activity of the adenosine agonist (R)-phenylisopropyladenosine in rabbits. Curr. Eye Res. 1992, 11, 453–458. [Google Scholar] [CrossRef]
- Polska, E.; Ehrlich, P.; Luksch, A.; Fuchsjager-Mayrl, G.; Schmetterer, L. Effects of adenosine on intraocular pressure, optic nerve head blood flow, and choroidal blood flow in healthy humans. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3110–3114. [Google Scholar] [CrossRef] [PubMed]
- Ming, F.; Chai, Y.; Chen, G.; Wang, H.; Huang, B. N6-(2-Hydroxyethyl)-adenosine exhibits Insecticidal activity against patella xylostella via Adenosine receptors. PLoS ONE 2016, 11, e0162859. [Google Scholar]
- Peng, X.X.; Chai, Y.Q.; Zhu, B.C.; Jin, Y.W.; Li, X.L.; Yu, L.S. The protective effects of N6-(2-hydroxyethyl)-adenosine extracted from Ophiocordyceps sobolifera on renal ischemia reperfusion injury (IRI) in mice. Mycosystema 2015, 34, 11–320. [Google Scholar]
- Hu, H.; Lu, W.; Laties, A.M.; Mitchell, C.H. Stimulation of the p2x7 receptor kills rat retinal ganglion cells in vivo. Exp. Eye Res. 2010, 91, 425–432. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conc | Inhibition of % |
|---|---|
| CCME 0.25 μg/mL | 4 ± 4 |
| CCME 2.5 μg/mL | 29 ± 4 |
| CCME 25 μg/mL | 4 ± 2 |
| K-115 21.4 ± 1.6 nM | 50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, L.-Y.; Hsu, J.-H.; Fu, H.-I.; Chen, C.-C.; Tung, K.-C. Lowering the Intraocular Pressure in Rats and Rabbits by Cordyceps cicadae Extract and Its Active Compounds. Molecules 2022, 27, 707. https://doi.org/10.3390/molecules27030707
Lee L-Y, Hsu J-H, Fu H-I, Chen C-C, Tung K-C. Lowering the Intraocular Pressure in Rats and Rabbits by Cordyceps cicadae Extract and Its Active Compounds. Molecules. 2022; 27(3):707. https://doi.org/10.3390/molecules27030707
Chicago/Turabian StyleLee, Li-Ya, Jui-Hsia Hsu, Hsin-I Fu, Chin-Chu Chen, and Kwong-Chung Tung. 2022. "Lowering the Intraocular Pressure in Rats and Rabbits by Cordyceps cicadae Extract and Its Active Compounds" Molecules 27, no. 3: 707. https://doi.org/10.3390/molecules27030707
APA StyleLee, L.-Y., Hsu, J.-H., Fu, H.-I., Chen, C.-C., & Tung, K.-C. (2022). Lowering the Intraocular Pressure in Rats and Rabbits by Cordyceps cicadae Extract and Its Active Compounds. Molecules, 27(3), 707. https://doi.org/10.3390/molecules27030707