
Hibiscus Acid from Hibiscus sabdariffa L. Inhibits Flagellar Motility and Cell Invasion in Salmonella enterica

 , ,
, ,  , and
, and 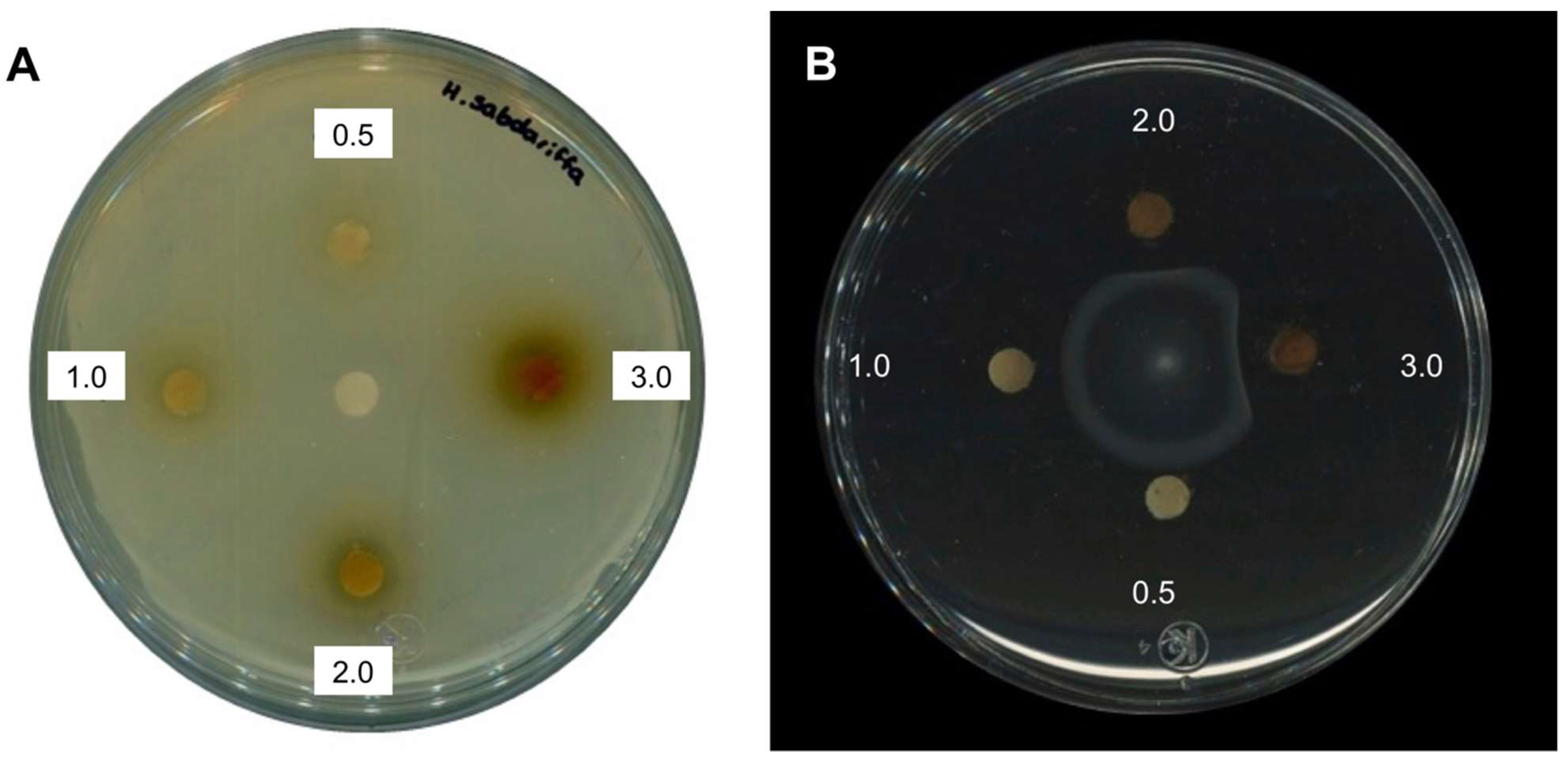{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
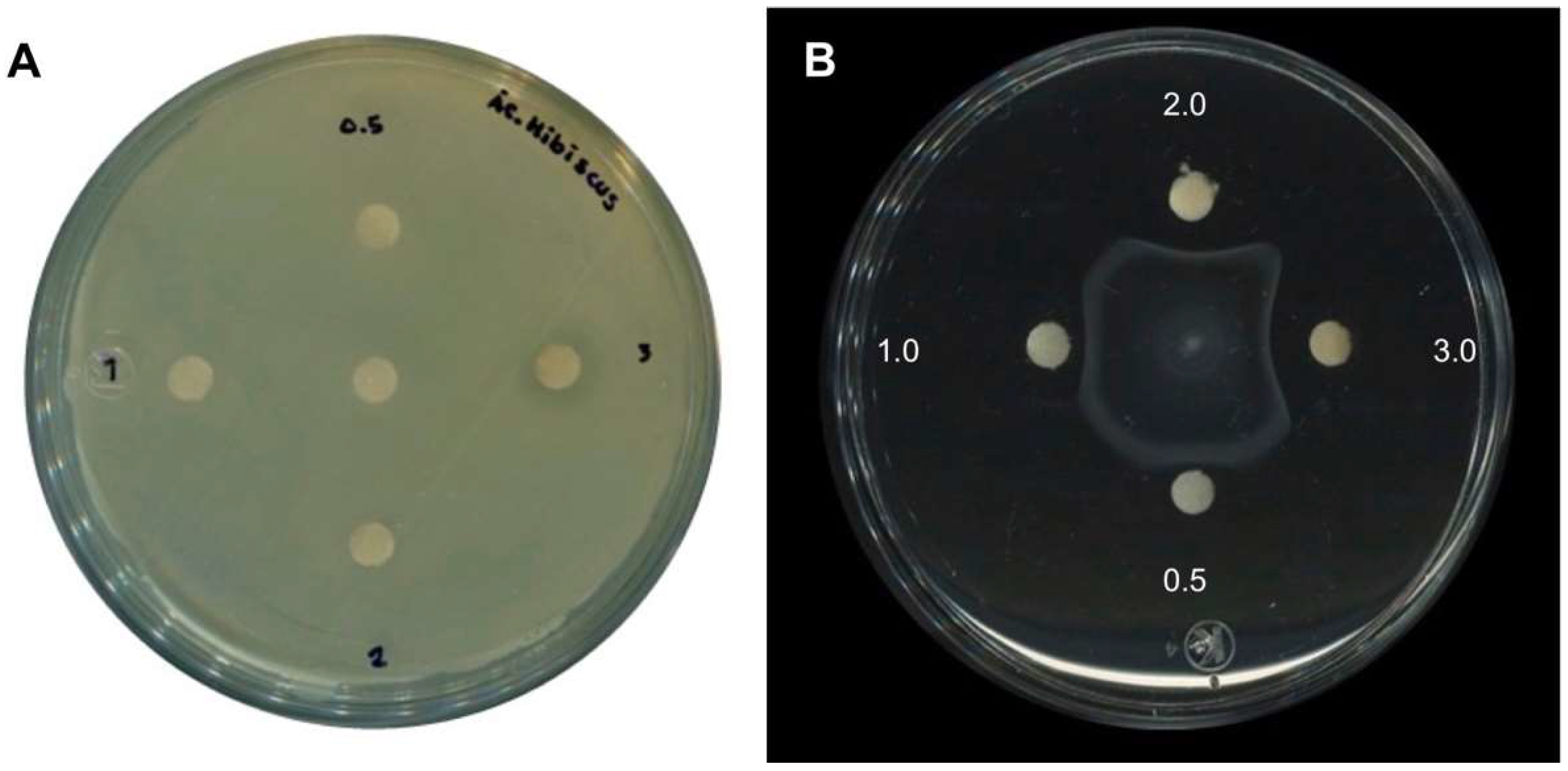
2.1. Antimicrobial Activity of H. sabdariffa L. Extracts
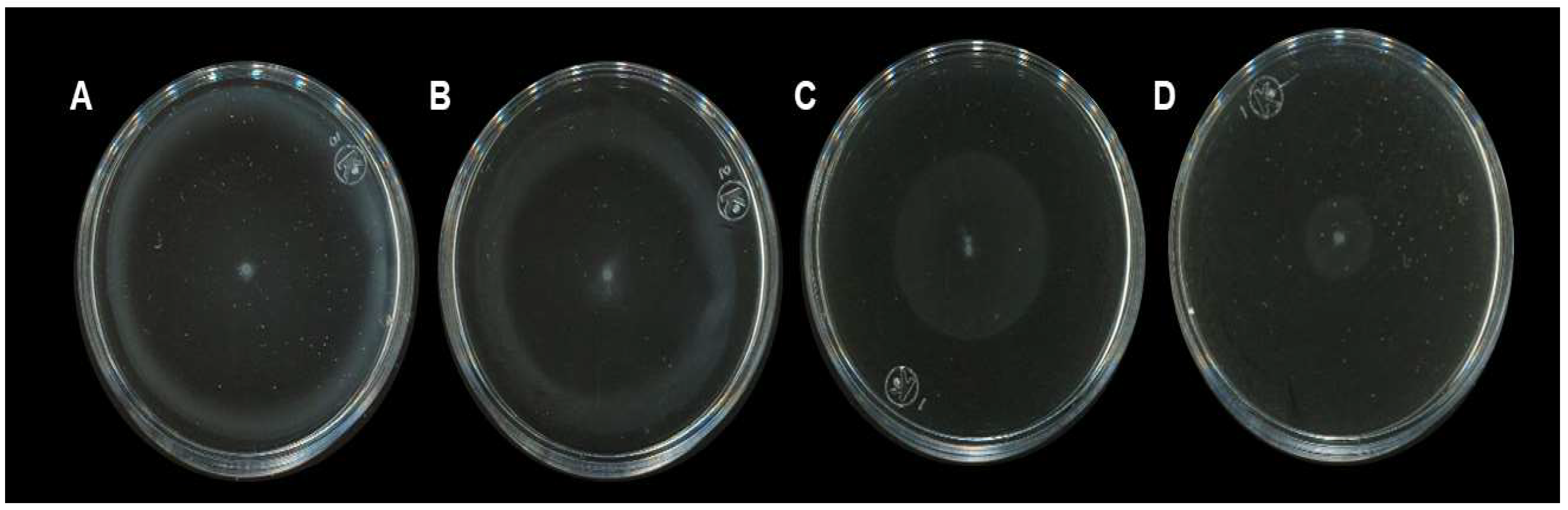
2.2. Anti-Motility Activity of H. sabdariffa L. Calyxes
2.3. Isolation of Secondary Metabolites from H. sabdariffa L.
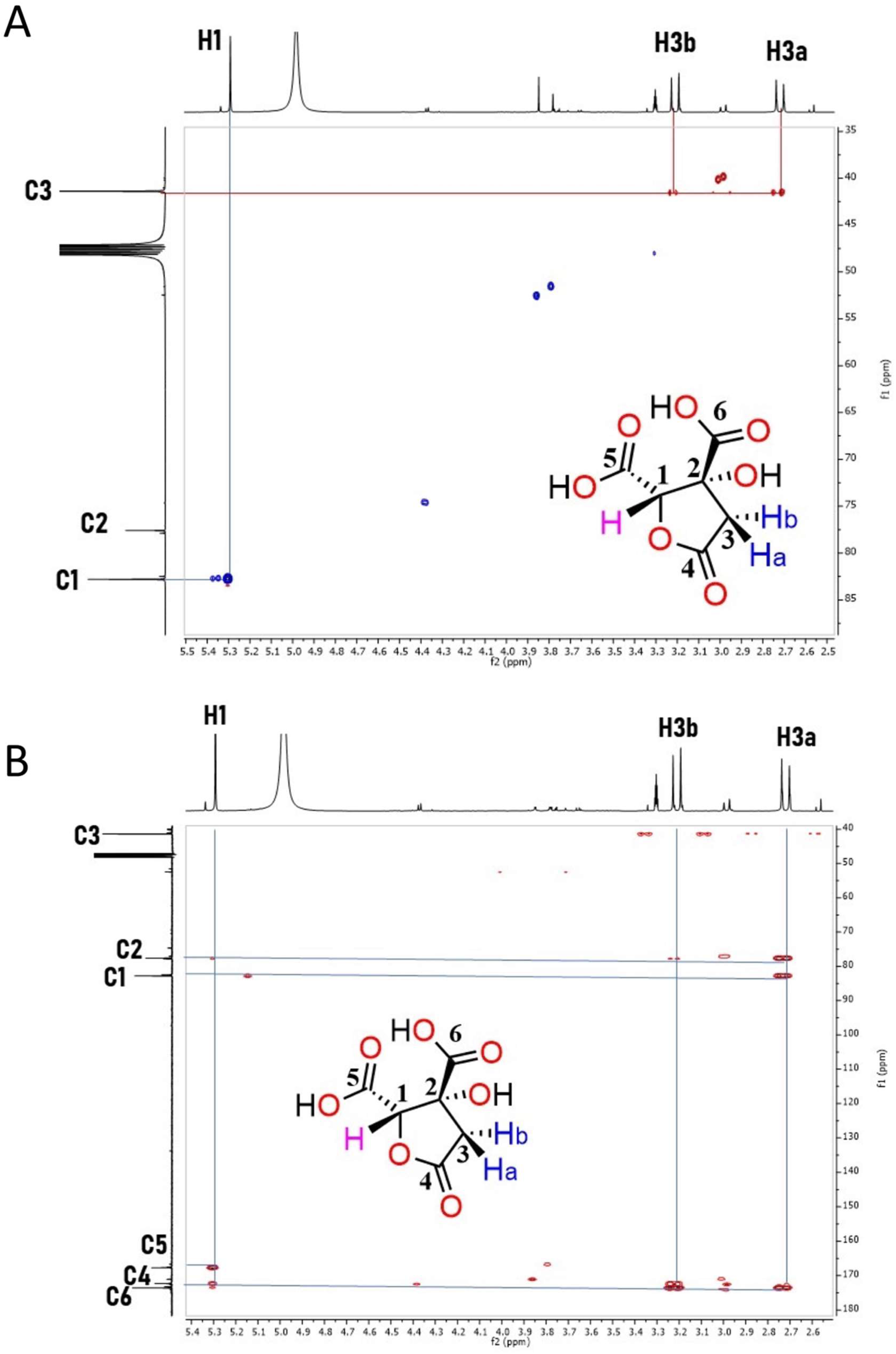
Characterization of HA by NMR and Mass Spectrometry
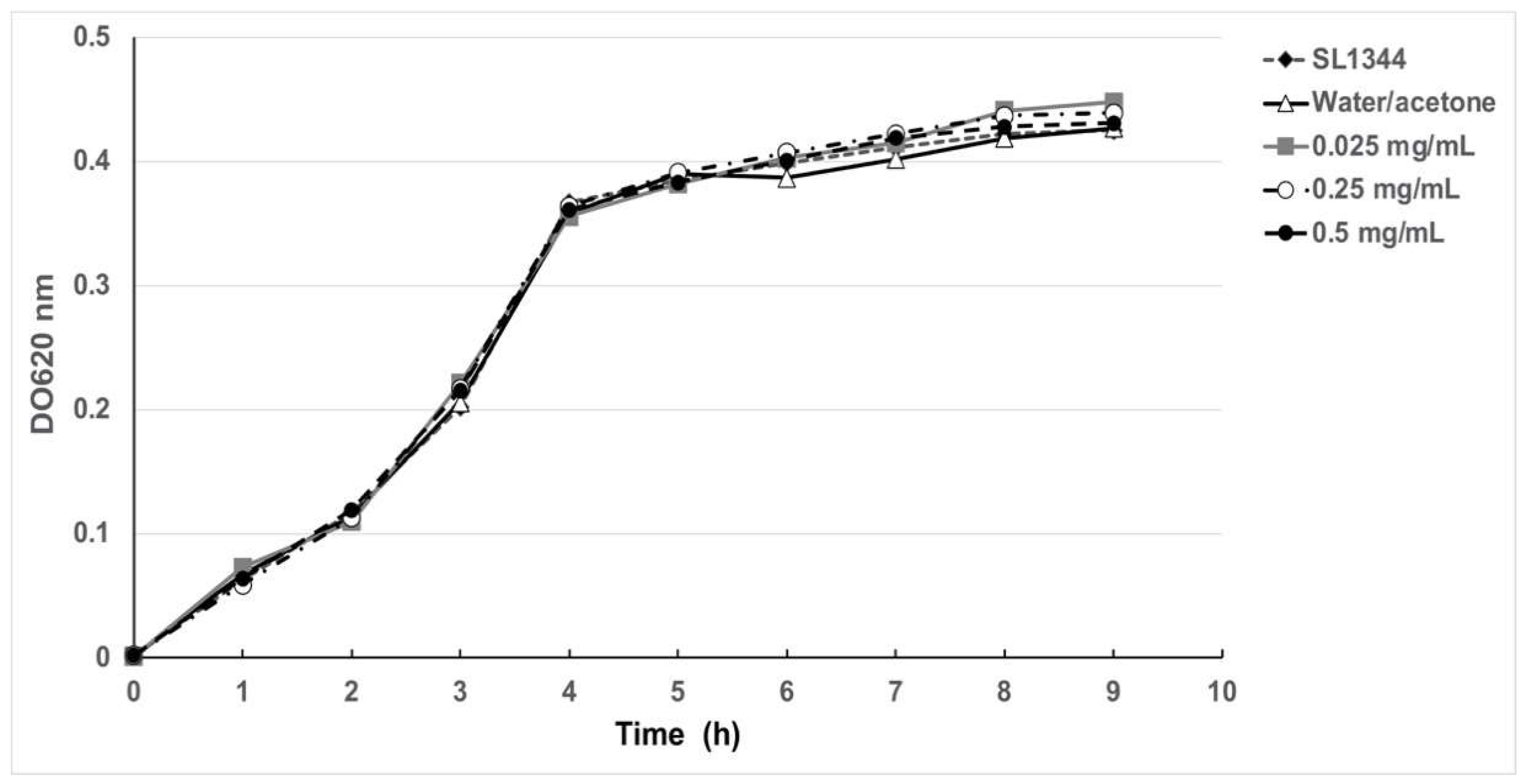
2.4. Antimicrobial and Anti-Motility Activities of HA
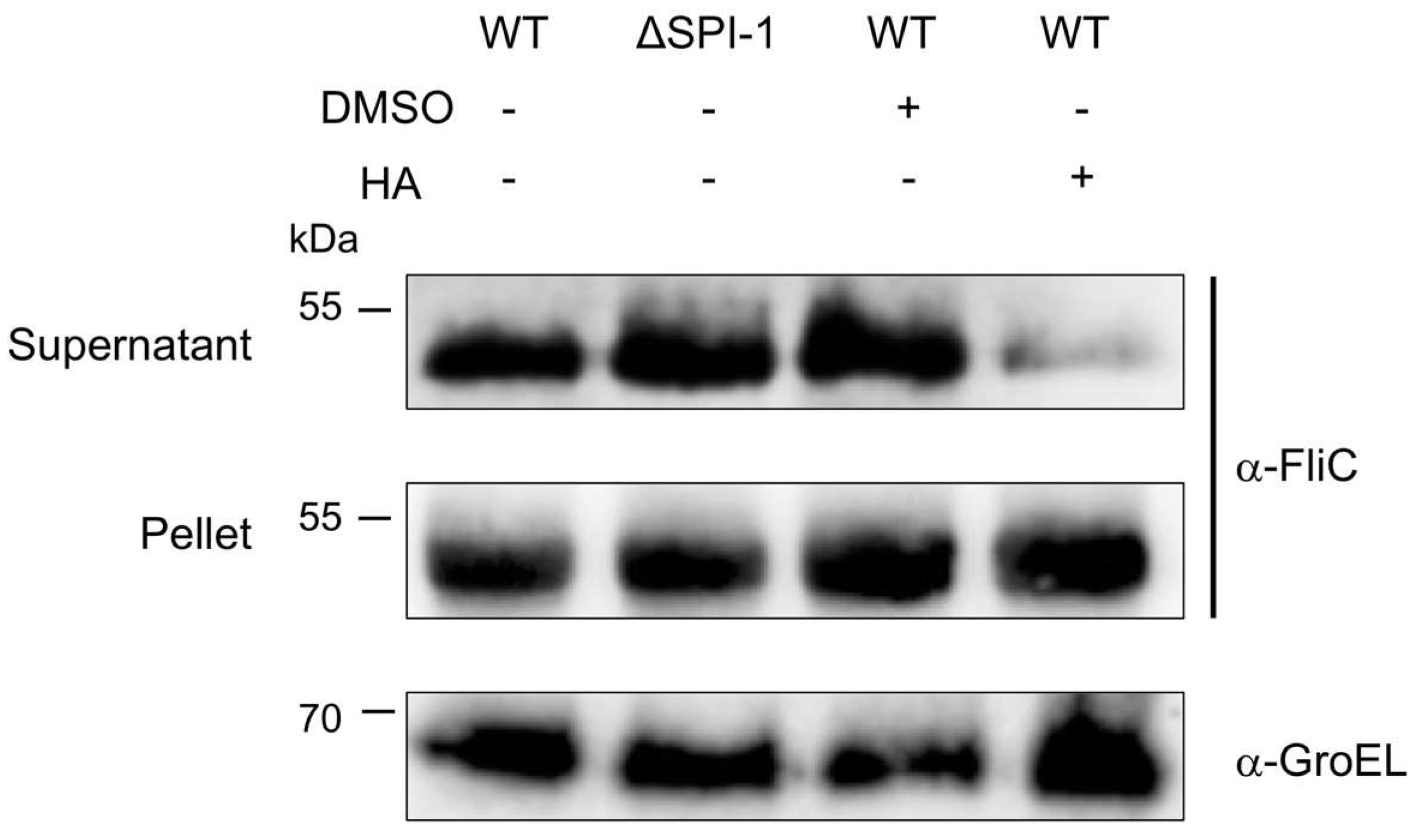
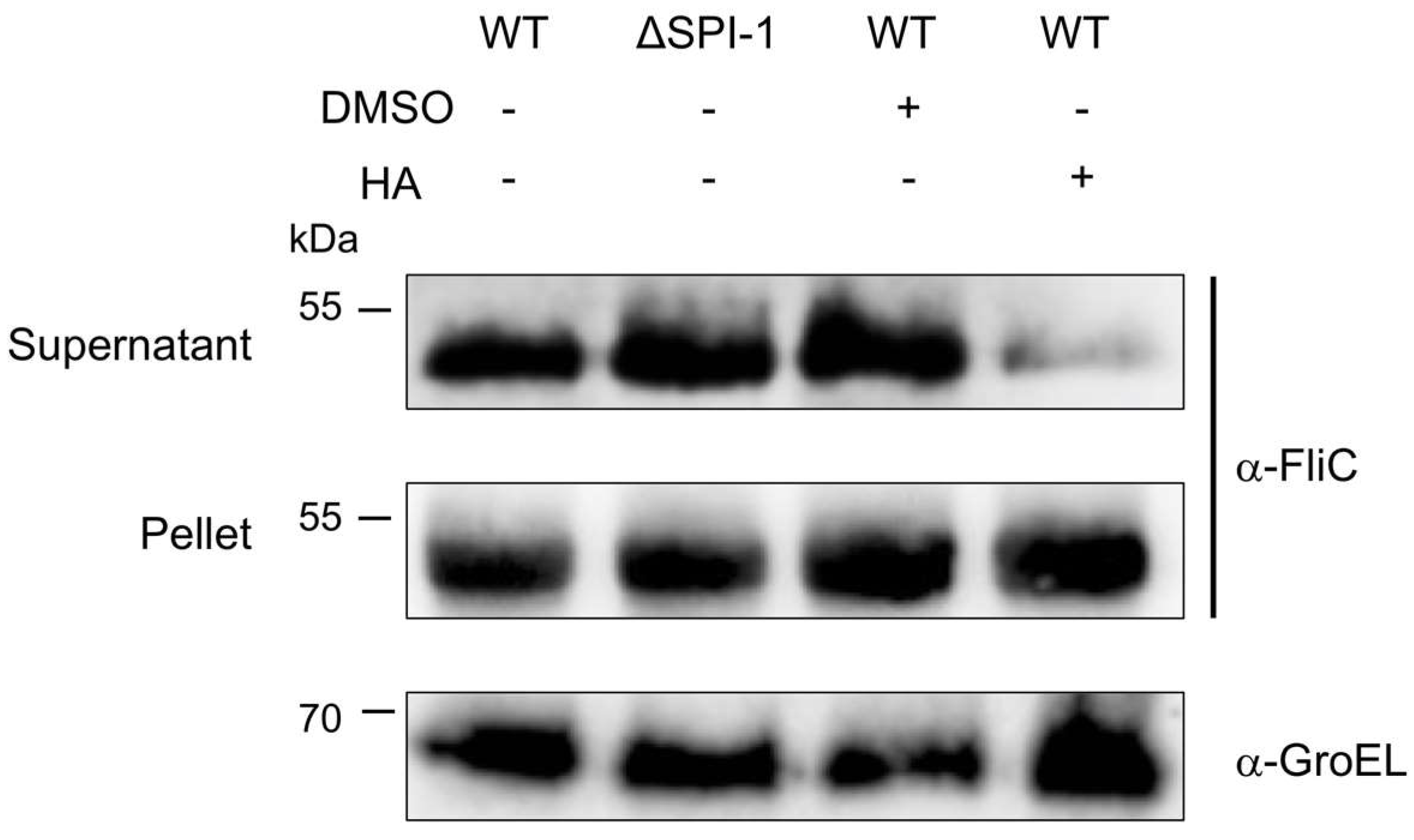
2.5. HA Partially Affects Flagellin Secretion
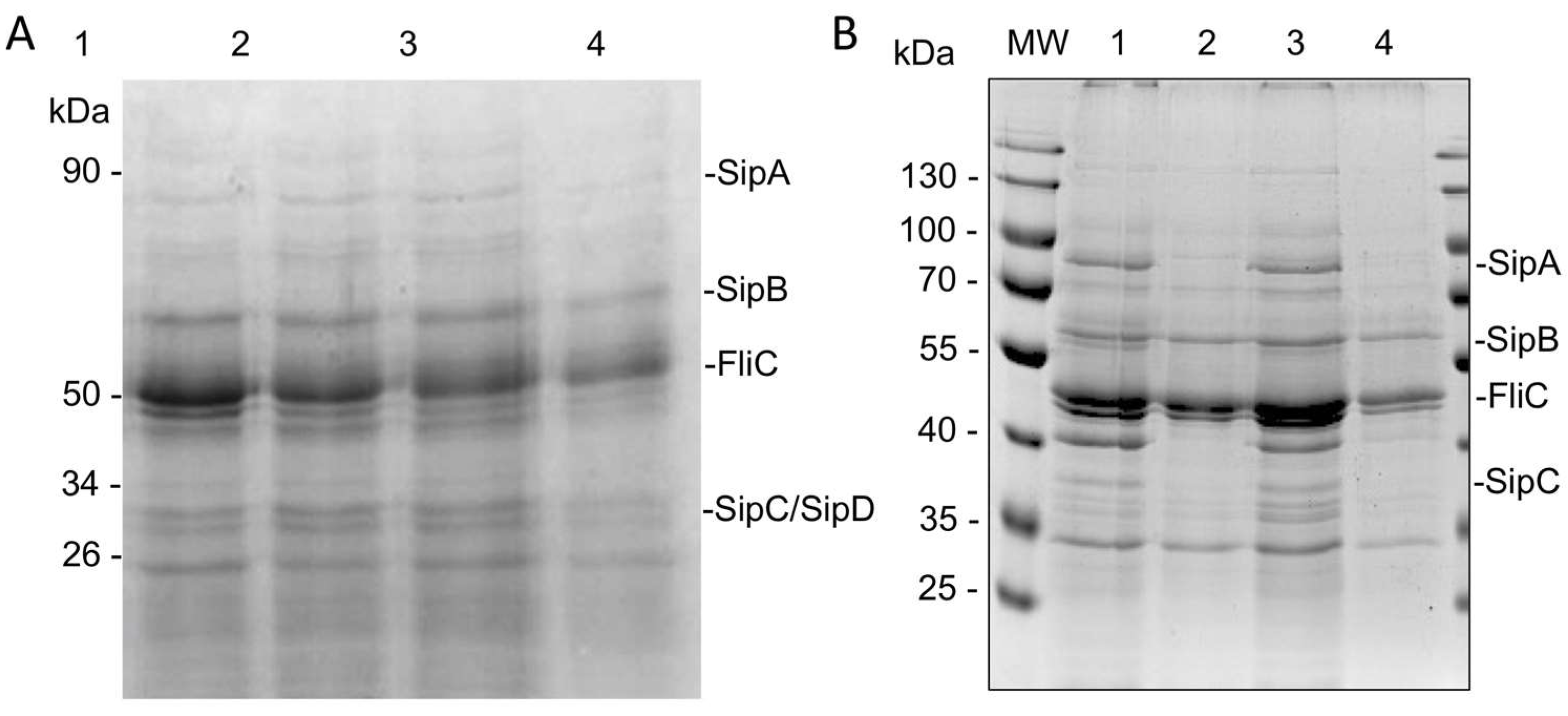
2.6. HA Partially Inhibits Secretion of T3SS Effector Proteins
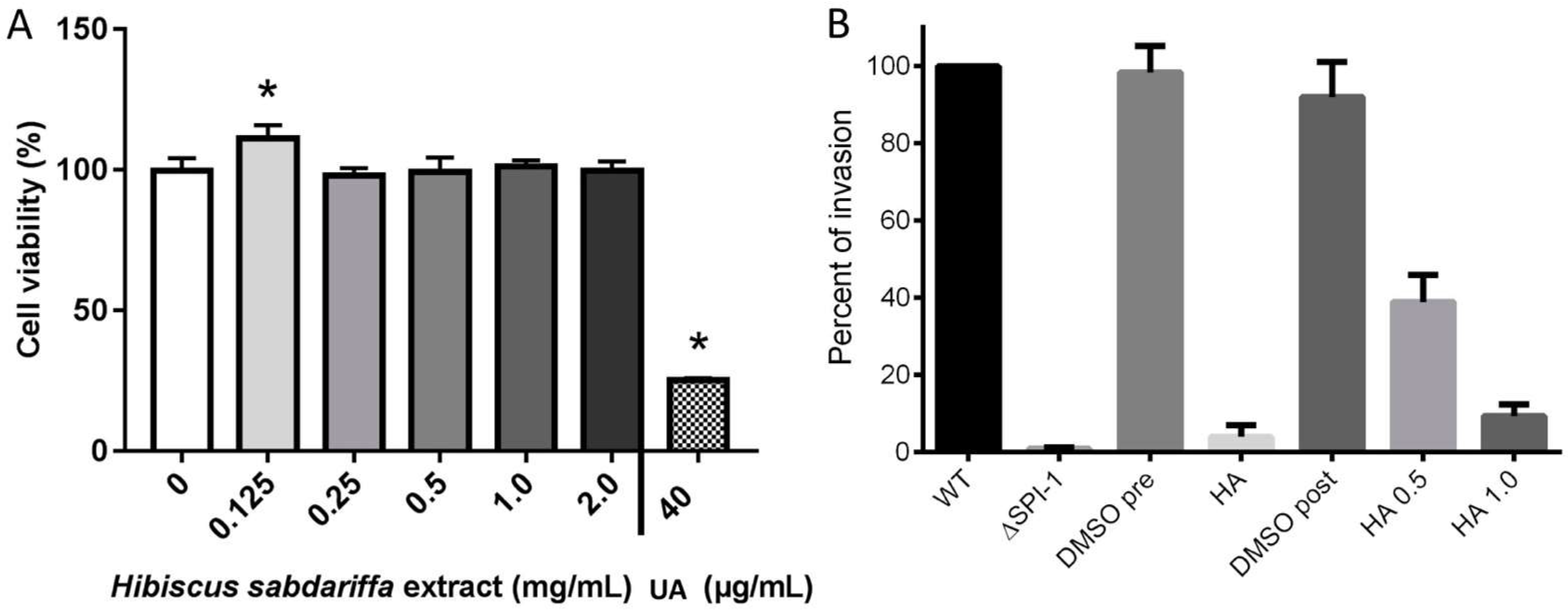
2.7. The Hibiscus Acid Is Not Cytotoxic for Human Epithelial Cells
2.8. HA Reduces Invasion of Salmonella
2.9. HA Induces Metabolic Alterations on Salmonella
3. Discussion
4. Materials and Methods
4.1. H. sabdariffa L. Extract
4.2. Isolation of Secondary Metabolites from H. sabdariffa L.
4.3. NMR and Low-Resolution Mass Spectrometry
4.4. Bacterial Strains and Growth Conditions
4.5. Antimicrobial Activity and Growth Curves
4.6. Motility Assay
4.7. Secretion Assay
4.8. Cytotoxicity Assays
4.9. Invasion Assays
4.10. Western Blot for Flagellin
4.11. Metabolic Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Mulvey, M.R.; Simor, A.E. Antimicrobial resistance in hospitals: How concerned should we be? Can. Med Assoc. J. 2009, 180, 408–415. [Google Scholar] [CrossRef] [Green Version]
- McEwen, S.A.; Collignon, P.J. Antimicrobial Resistance: A One Health Perspective. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Ferri, M.; Ranucci, E.; Romagnoli, P.; Giaccone, V. Antimicrobial resistance: A global emerging threat to public health systems. Crit. Rev. Food Sci. Nutr. 2017, 57, 2857–2876. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol. Rev. 2018, 42, fux053. [Google Scholar] [CrossRef]
- Dhingra, S.; Rahman, N.A.A.; Peile, E.; Rahman, M.; Sartelli, M.; Hassali, M.A.; Islam, T.; Islam, S.; Haque, M. Microbial Resistance Movements: An Overview of Global Public Health Threats Posed by Antimicrobial Resistance, and How Best to Counter. Front. Public Health 2020, 8, 535668. [Google Scholar] [CrossRef]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad Bugs, No Drugs: No ESKAPE! An Update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug Discov. 2010, 9, 117–128. [Google Scholar] [CrossRef]
- Fernebro, J. Fighting bacterial infections—Future treatment options. Drug Resist. Updat. 2011, 14, 125–139. [Google Scholar] [CrossRef]
- Gill, E.E.; Franco, O.L.; Hancock, R.E. Antibiotic Adjuvants: Diverse Strategies for Controlling Drug-Resistant Pathogens. Chem. Biol. Drug Des. 2015, 85, 56–78. [Google Scholar] [CrossRef]
- Sharma, A.K.; Dhasmana, N.; Dubey, N.; Kumar, N.; Gangwal, A.; Gupta, M.; Singh, Y. Bacterial Virulence Factors: Secreted for Survival. Indian J. Microbiol. 2017, 57, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.N.; Zimmer, K.R.; Macedo, A.J.; Trentin, D. Plant Natural Products Targeting Bacterial Virulence Factors. Chem. Rev. 2016, 116, 9162–9236. [Google Scholar] [CrossRef]
- Lamas, A.; Miranda, J.M.; Regal, P.; Vazquez, B.; Franco, C.M.; Cepeda, A. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol. Res. 2018, 206, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, A.M.P.; Ferrari, R.G.; Conte-Junior, C.A. Virulence Factors in Salmonella Typhimurium: The Sagacity of a Bacterium. Curr. Microbiol. 2019, 76, 762–773. [Google Scholar] [CrossRef]
- Ibarra, J.A.; Knodler, L.A.; Sturdevant, D.E.; Virtaneva, K.; Carmody, A.B.; Fischer, E.R.; Porcella, S.F.; Steele-Mortimer, O. Induction of Salmonella pathogenicity island 1 under different growth conditions can affect Salmonella–host cell interactions in vitro. Microbiology 2010, 156, 1120–1133. [Google Scholar] [CrossRef] [Green Version]
- De Jong, H.K.; Parry, C.M.; Van Der Poll, T.; Wiersinga, W.J. Host–Pathogen Interaction in Invasive Salmonellosis. PLoS Pathog. 2012, 8, e1002933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero-Fresno, A.; Olsen, J.E. Salmonella Typhimurium metabolism affects virulence in the host—A mini-review. Food Microbiol. 2018, 71, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Hsieh, Y.H.; Powers, Z.M.; Kao, C.Y. Defeating Antibiotic-Resistant Bacteria: Exploring Alternative Therapies for a Post-Antibiotic Era. Int. J. Mol. Sci. 2020, 21, 1061. [Google Scholar] [CrossRef] [Green Version]
- Vasudeva, N.; Sharma, S.K. Biologically Active Compounds from the Genus Hibiscus. Pharm. Biol. 2008, 46, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Da-Costa-Rocha, I.; Bonnlaender, B.; Sievers, H.; Pischel, I.; Heinrich, M. Hibiscus sabdariffa L.—A phytochemical and pharmacological review. Food Chem. 2014, 165, 424–443. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.-S.; Tsao, S.-M.; Yin, M.-C. In vitro antibacterial activity of roselle calyx and protocatechuic acid. Phytother. Res. 2005, 19, 942–945. [Google Scholar] [CrossRef]
- Gutiérrez-Alcántara, E.J.; Rangel-Vargas, E.; Gómez-Aldapa, C.A.; Cortes, R.N.F.; Rodriguez, M.L.R.M.; Godínez-Oviedo, A.; Acevedo-Sandoval, O.A.; Castro-Rosas, J. Attachment of 13 Types of Foodborne Bacteria to Jalapeño and Serrano Peppers and Antibacterial Effect of Roselle Calyx Extracts, Sodium Hypochlorite, Colloidal Silver, and Acetic Acid against These Foodborne Bacteria on Peppers. J. Food Prot. 2017, 80, 406–413. [Google Scholar] [CrossRef]
- Gutiérrez-Alcántara, E.J.; Rangel-Vargas, E.; Gómez-Aldapa, C.A.; Cortes, R.N.F.; Rodriguez, M.L.R.M.; Godínez-Oviedo, A.; Cortes-López, H.; Castro-Rosas, J. Antibacterial effect of roselle extracts (Hibiscus sabadariffa), sodium hypochlorite and acetic acid against multidrug-resistant Salmonella strains isolated from tomatoes. Lett. Appl. Microbiol. 2015, 62, 177–184. [Google Scholar] [CrossRef]
- Portillo-Torres, L.A.; Bernardino-Nicanor, A.; Gómez-Aldapa, C.A.; González-Montiel, S.; Rangel-Vargas, E.; Villagómez-Ibarra, J.R.; González-Cruz, L.; Cortés-López, H.; Castro-Rosas, J. Hibiscus Acid and Chromatographic Fractions from Hibiscus sabdariffa Calyces: Antimicrobial Activity against Multidrug-Resistant Pathogenic Bacteria. Antibiotics 2019, 8, 218. [Google Scholar] [CrossRef] [Green Version]
- Dunlop, D.M. The use of 2:4-dinitrophenol as a metabolic stimulant. Brit. Med. J. 1934, 1, 524–527. [Google Scholar] [CrossRef]
- Sánchez-Clemente, R.; Igeño, M.I.; Población, A.G.; Guijo, M.I.; Merchán, F.; Blasco, R. Study of pH Changes in Media during Bacterial Growth of Several Environmental Strains. Proceedings 2018, 2, 1297. [Google Scholar] [CrossRef] [Green Version]
- Geisler, J.G. 2,4 Dinitrophenol as Medicine. Cells 2019, 8, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, D.; Carmo, H.; Roque Bravo, R.; Carvalho, F.; Bastos, M.D.L.; De Pinho, P.G.; Da Silva, D.D. Diet aid or aid to die: An update on 2,4-dinitrophenol (2,4-DNP) use as a weight-loss product. Arch. Toxicol. 2020, 94, 1071–1083. [Google Scholar] [CrossRef]
- Abdallah, E.M. Antibacterial efficiency of the Sudanese Roselle (Hibiscus sabdariffa L.), a famous beverage from Sudanese folk medicine. J. Intercult. Ethnopharmacol. 2016, 5, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Baena-Santillán, E.S.; Piloni-Martini, J.; Santos-López, E.M.; Gómez-Aldapa, C.A.; Rangel-Vargas, E.; Castro-Rosas, J. Comparison of the Antimicrobial Activity of Hibiscus sabdariffa Calyx Extracts, Six Commercial Types of Mouthwashes, and Chlorhexidine on Oral Pathogenic Bacteria, and the Effect of Hibiscus sabdariffa Extracts and Chlorhexidine on Permeability of the Bacterial Membrane. J. Med. Food 2021, 24, 67–76. [Google Scholar] [CrossRef]
- Hansawasdi, C.; Kawabata, J.; Kasai, T. Hibiscus Acid as an Inhibitor of Starch Digestion in the Caco-2 Cell Model System. Biosci. Biotechnol. Biochem. 2001, 65, 2087–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheoat, A.M.; Gray, A.I.; Igoli, J.O.; Kennedy, A.R.; Ferro, V.A. Crystal structures of hibiscus acid and hibiscus acid dimethyl ester isolated from Hibiscus sabdariffa (Malvaceae). Acta Crystallogr. E Crystallogr. Commun. 2017, 73, 1368–1371. [Google Scholar] [CrossRef] [Green Version]
- Zheoat, A.M.; Gray, A.I.; Igoli, J.O.; Ferro, V.; Drummond, R.M. Hibiscus acid from Hibiscus sabdariffa (Malvaceae) has a vasorelaxant effect on the rat aorta. Fitoterapia 2019, 134, 5–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansawasdi, C.; Kawabata, J.; Kasai, T. Alpha-Amylase Inhibitors from Roselle (Hibiscus sabdariffa Linn.) Tea. Biosci. Biotechnol. Biochem. 2000, 64, 1041–1043. [Google Scholar] [CrossRef] [PubMed]
- Cortes-López, H.; Castro-Rosas, J.; García-Contreras, R.; Rodríguez-Zavala, J.S.; González-Pedrajo, B.; Díaz-Guerrero, M.; Hernández-Morales, J.; Muñoz-Cazares, N.; Soto-Hernández, M.; Ruíz-Posadas, L.D.M.; et al. Antivirulence Activity of a Dietary Phytochemical: Hibiscus Acid Isolated from Hibiscus sabdariffa L. Reduces the Virulence of Pseudomonas aeruginosa in a Mouse Infection Model. J. Med. Food 2021, 24, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Shin, D.; Kim, M.; Park, J.; Lim, S.; Ryu, S. LsrR-Mediated Quorum Sensing Controls Invasiveness of Salmonella typhimurium by Regulating SPI-1 and Flagella Genes. PLoS ONE 2012, 7, e37059. [Google Scholar] [CrossRef] [PubMed]
- Diepold, A.; Wagner, S. Assembly of the bacterial type III secretion machinery. FEMS Microbiol. Rev. 2014, 38, 802–822. [Google Scholar] [CrossRef] [Green Version]
- Jin, Q.; Kirk, M.F. pH as a Primary Control in Environmental Microbiology: 1. Thermodynamic Perspective. Front. Environ. Sci. 2018, 6, 21. [Google Scholar] [CrossRef]
- Berg, H.; Turner, L. Torque generated by the flagellar motor of Escherichia coli. Biophys. J. 1993, 65, 2201–2216. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Brown, D.G. Variation in Bacterial ATP Level and Proton Motive Force Due to Adhesion to a Solid Surface. Appl. Environ. Microbiol. 2020, 75, 2346–2353. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, G.F.; Fleet, G.H.; Lyons, M.J.; Walker, R.A. Method for the isolation of highly purified Salmonella flagellins. J. Clin. Microbiol. 1985, 22, 1040–1044. [Google Scholar] [CrossRef] [Green Version]
- Maurer, L.M.; Yohannes, E.; Bondurant, S.S.; Radmacher, M.; Slonczewski, J.L. pH Regulates Genes for Flagellar Motility, Catabolism, and Oxidative Stress in Escherichia coli K-12. J. Bacteriol. 2005, 187, 304–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseini, A.; Bakhtiari, E.; Mousavi, S.H. Protective Effect of Hibiscus sabdariffa on Doxorubicin-induced Cytotoxicity in H9c2 Cardiomyoblast Cells. Iran J. Pharm. Res. 2017, 16, 708–713. [Google Scholar]
- CLSI. Procedure for Optimizing Disk Contents (Potencies) for Disk Diffusion Testing of Antimicrobial Agents Using Harmonized CLSI and EUCAST Criteria. In CLSI Document M23S, 1st ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020; pp. 1–50. [Google Scholar]
- Banda, M.M.; López, C.; Manzo, R.; Rico-Pérez, G.; García, P.; Rosales-Reyes, R.; De la Cruz, M.A.; Soncini, F.C.; Portillo, F.G.-D.; Bustamante, V.H. HilD and PhoP independently regulate the expression of grhD1, a novel gene required for Salmonella Typhimurium invasion of host cells. Sci. Rep. 2018, 8, 4841, Erratum in Sci. Rep. 2018, 8, 7697. [Google Scholar] [CrossRef] [PubMed]
- Castrejón-Jiménez, N.S.; Leyva-Paredes, K.; Baltierra-Uribe, S.L.; Castillo-Cruz, J.; Campillo-Navarro, M.; Hernández-Pérez, A.D.; Luna-Angulo, A.B.; Chacón-Salinas, R.; Coral-Vázquez, R.M.; Estrada-García, I.; et al. Ursolic and Oleanolic Acids Induce Mitophagy in A549 Human Lung Cancer Cells. Molecules 2019, 24, 3444. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sedillo-Torres, I.Y.; Hernández-Rangel, Á.O.; Gómez-y-Gómez, Y.; Cortés-Avalos, D.; García-Pérez, B.E.; Villalobos-Rocha, J.C.; Hernández-Rodríguez, C.H.; Zepeda-Vallejo, L.G.; Estrada-de los Santos, P.; Vargas-Díaz, M.E.; et al. Hibiscus Acid from Hibiscus sabdariffa L. Inhibits Flagellar Motility and Cell Invasion in Salmonella enterica. Molecules 2022, 27, 655. https://doi.org/10.3390/molecules27030655
Sedillo-Torres IY, Hernández-Rangel ÁO, Gómez-y-Gómez Y, Cortés-Avalos D, García-Pérez BE, Villalobos-Rocha JC, Hernández-Rodríguez CH, Zepeda-Vallejo LG, Estrada-de los Santos P, Vargas-Díaz ME, et al. Hibiscus Acid from Hibiscus sabdariffa L. Inhibits Flagellar Motility and Cell Invasion in Salmonella enterica. Molecules. 2022; 27(3):655. https://doi.org/10.3390/molecules27030655
Chicago/Turabian StyleSedillo-Torres, Ixchell Y., Álvaro O. Hernández-Rangel, Yolanda Gómez-y-Gómez, Daniel Cortés-Avalos, Blanca Estela García-Pérez, Juan C. Villalobos-Rocha, César H. Hernández-Rodríguez, Luis Gerardo Zepeda-Vallejo, Paulina Estrada-de los Santos, María Elena Vargas-Díaz, and et al. 2022. "Hibiscus Acid from Hibiscus sabdariffa L. Inhibits Flagellar Motility and Cell Invasion in Salmonella enterica" Molecules 27, no. 3: 655. https://doi.org/10.3390/molecules27030655
APA StyleSedillo-Torres, I. Y., Hernández-Rangel, Á. O., Gómez-y-Gómez, Y., Cortés-Avalos, D., García-Pérez, B. E., Villalobos-Rocha, J. C., Hernández-Rodríguez, C. H., Zepeda-Vallejo, L. G., Estrada-de los Santos, P., Vargas-Díaz, M. E., & Ibarra, J. A. (2022). Hibiscus Acid from Hibiscus sabdariffa L. Inhibits Flagellar Motility and Cell Invasion in Salmonella enterica. Molecules, 27(3), 655. https://doi.org/10.3390/molecules27030655