
Uncovering the Anticancer Potential of Polydatin: A Mechanistic Insight

 ,
,  ,
,  , , , , , , ,
, , , , , , ,  and
and 
Abstract
1. Introduction
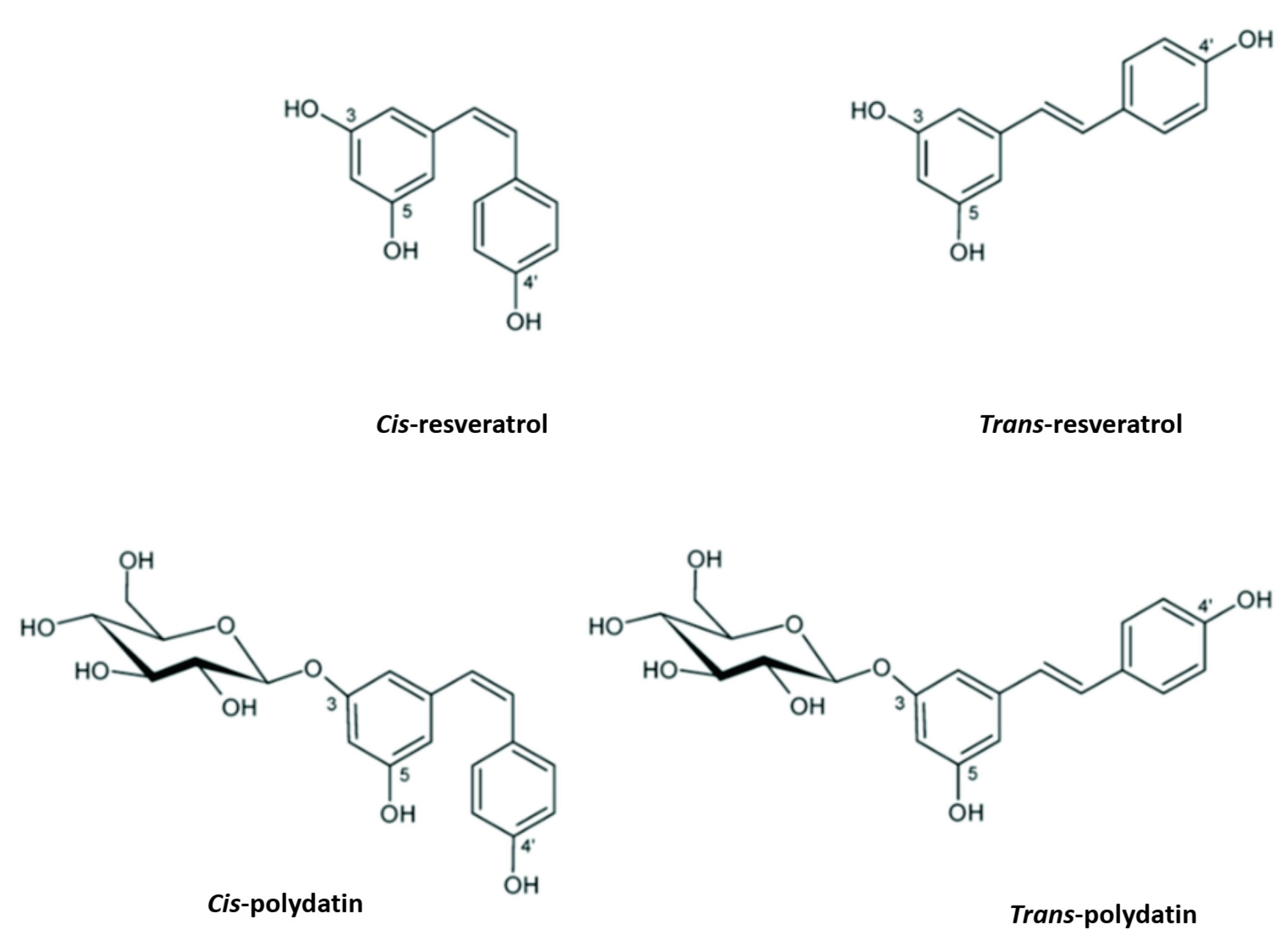
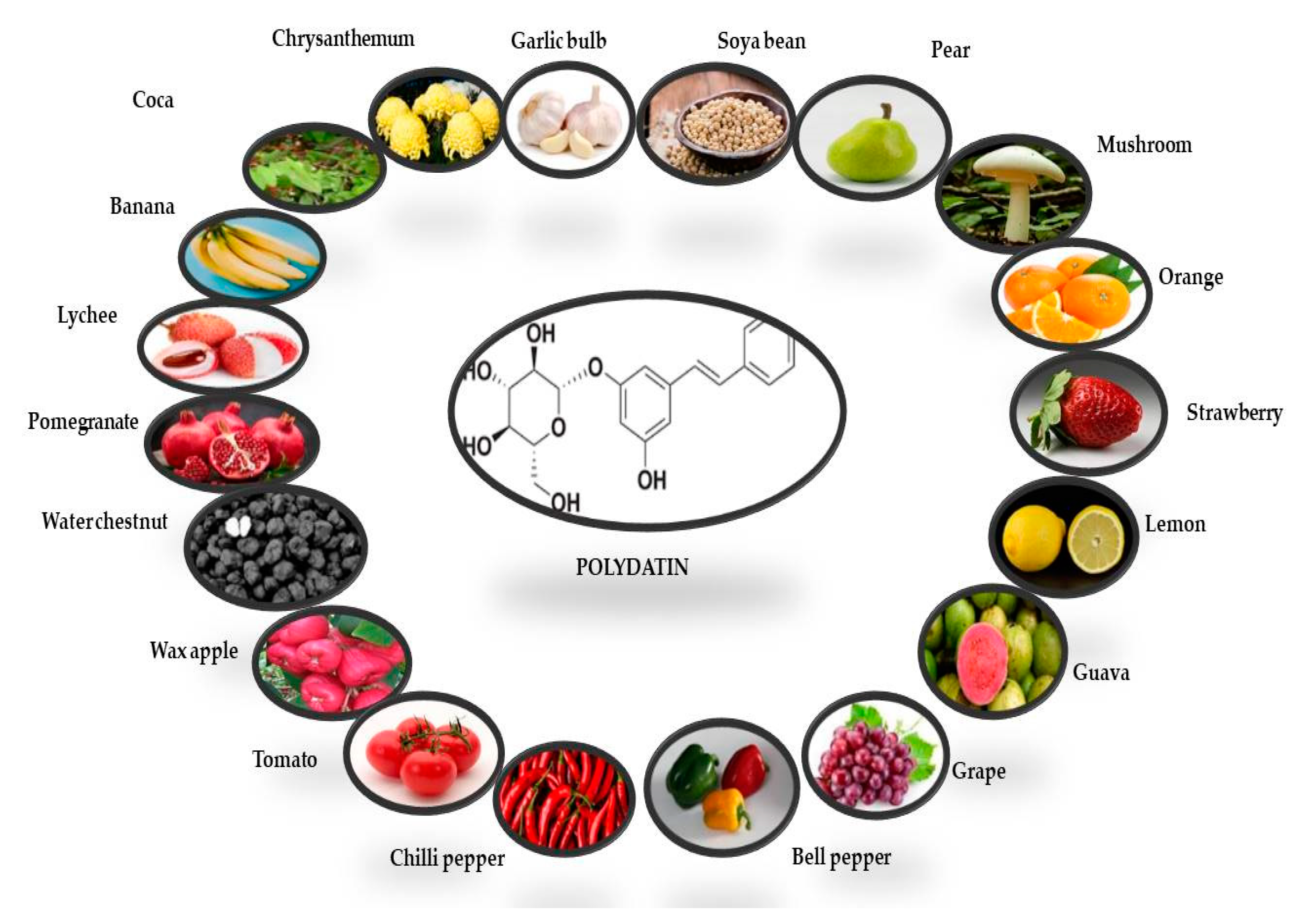
2. Polydatin Chemistry and Biosynthesis
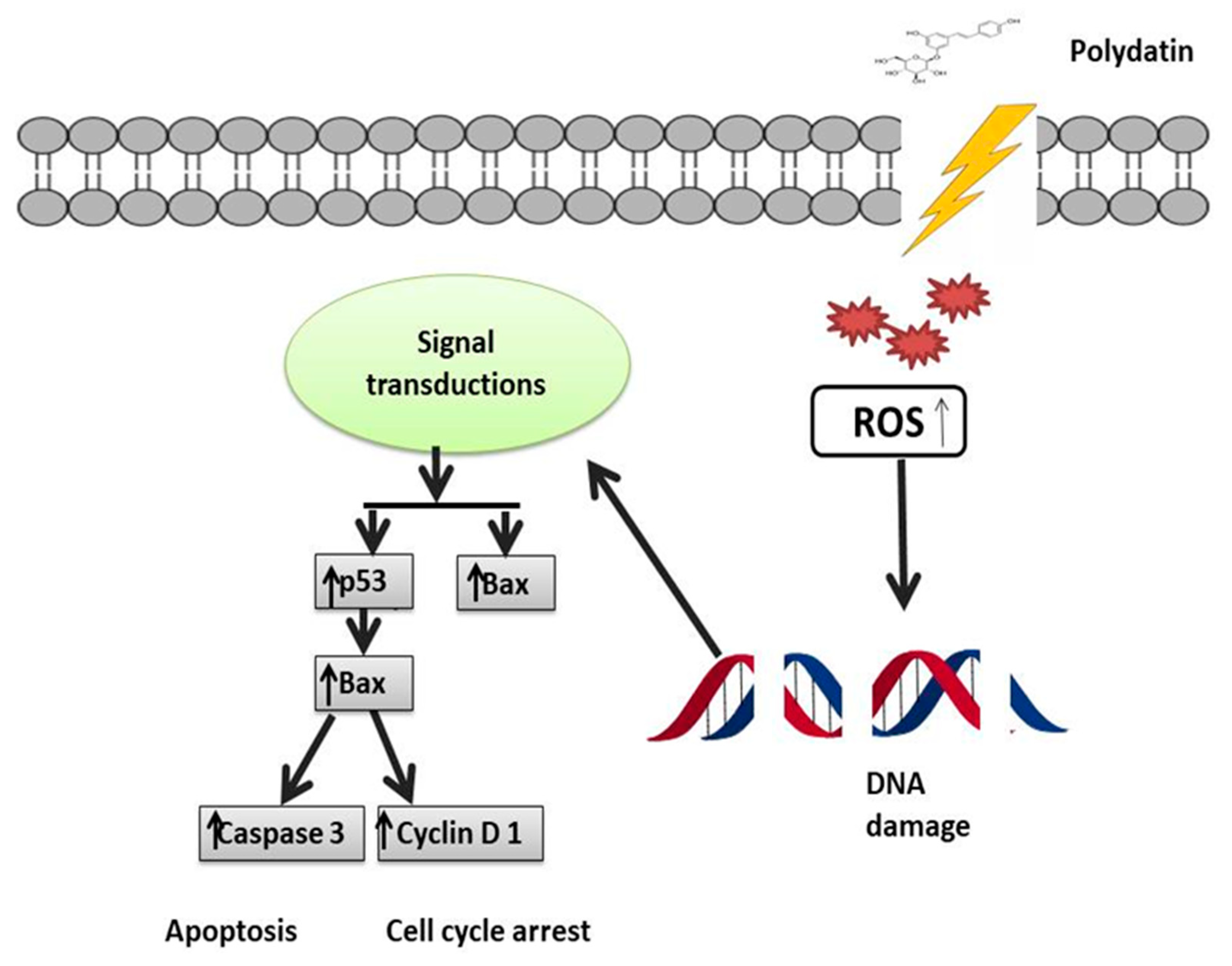
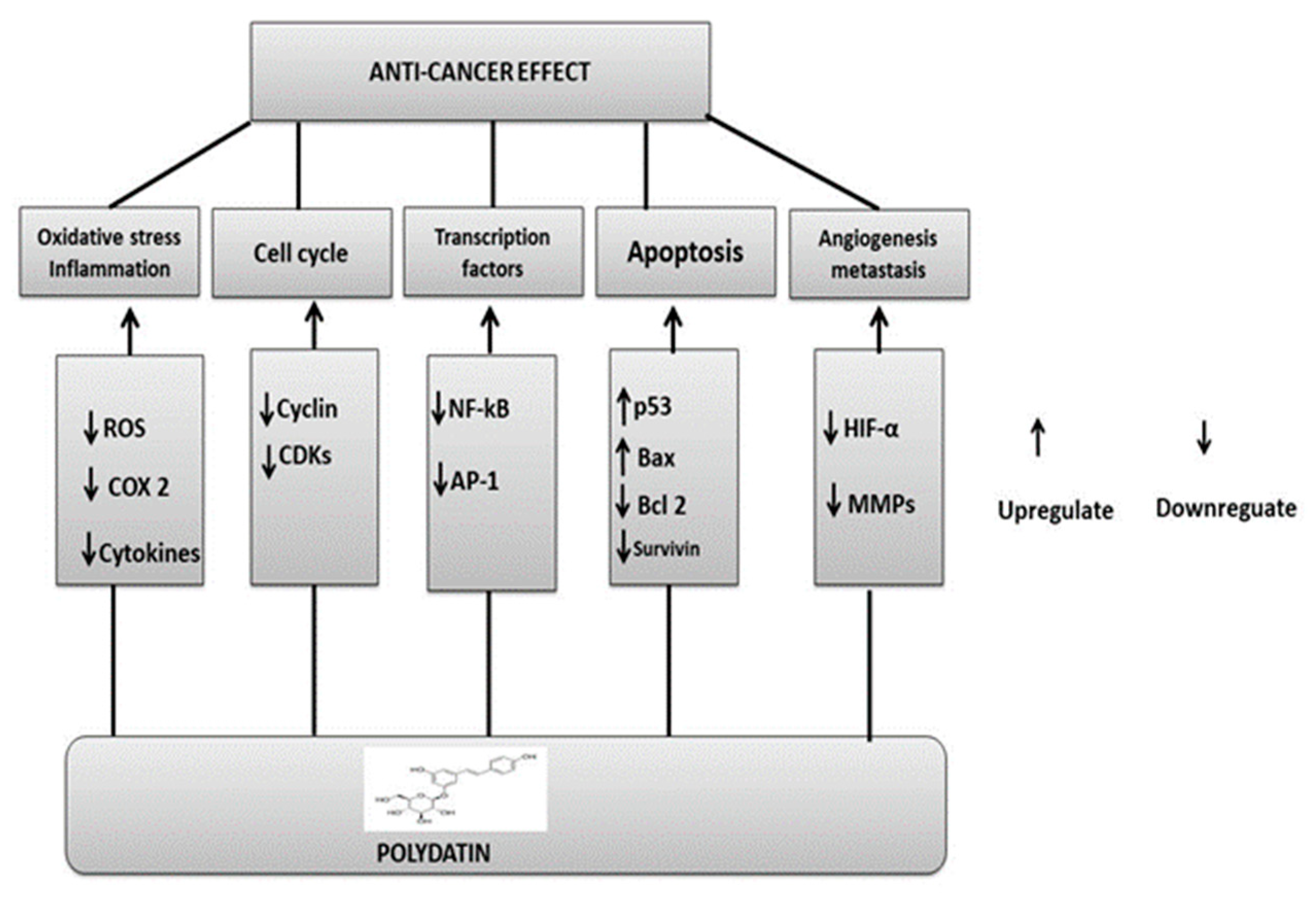
3. Role of Polydatin in Cancer
4. Anticancer Activity of Polydatin on Liver, Colon, Bone, Breast, Lung, Cervical, and Ovarian Cancer Proliferation
4.1. Liver Cancer
4.2. Colon Cancer
4.3. Bone Cancer
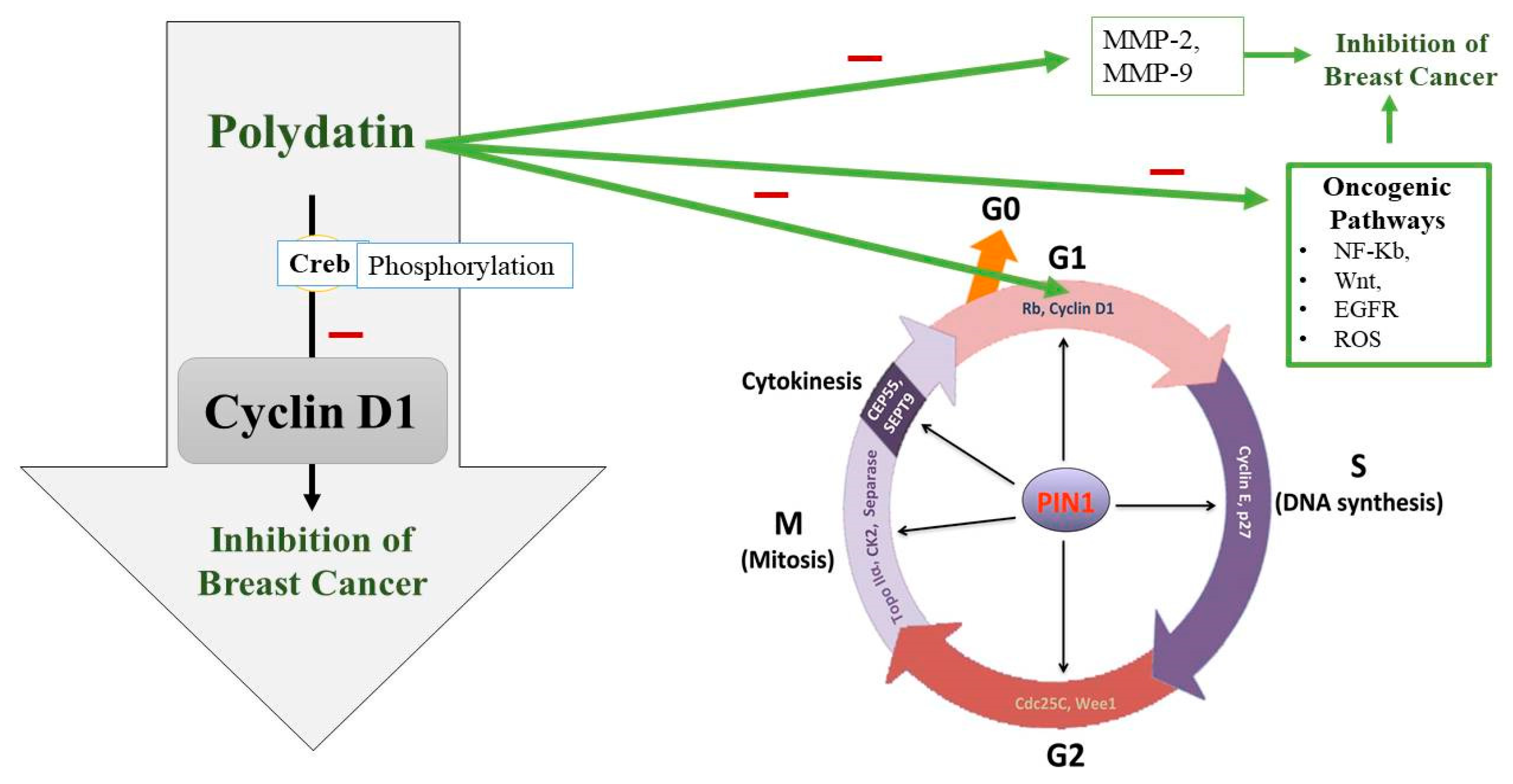
4.4. Breast Cancer
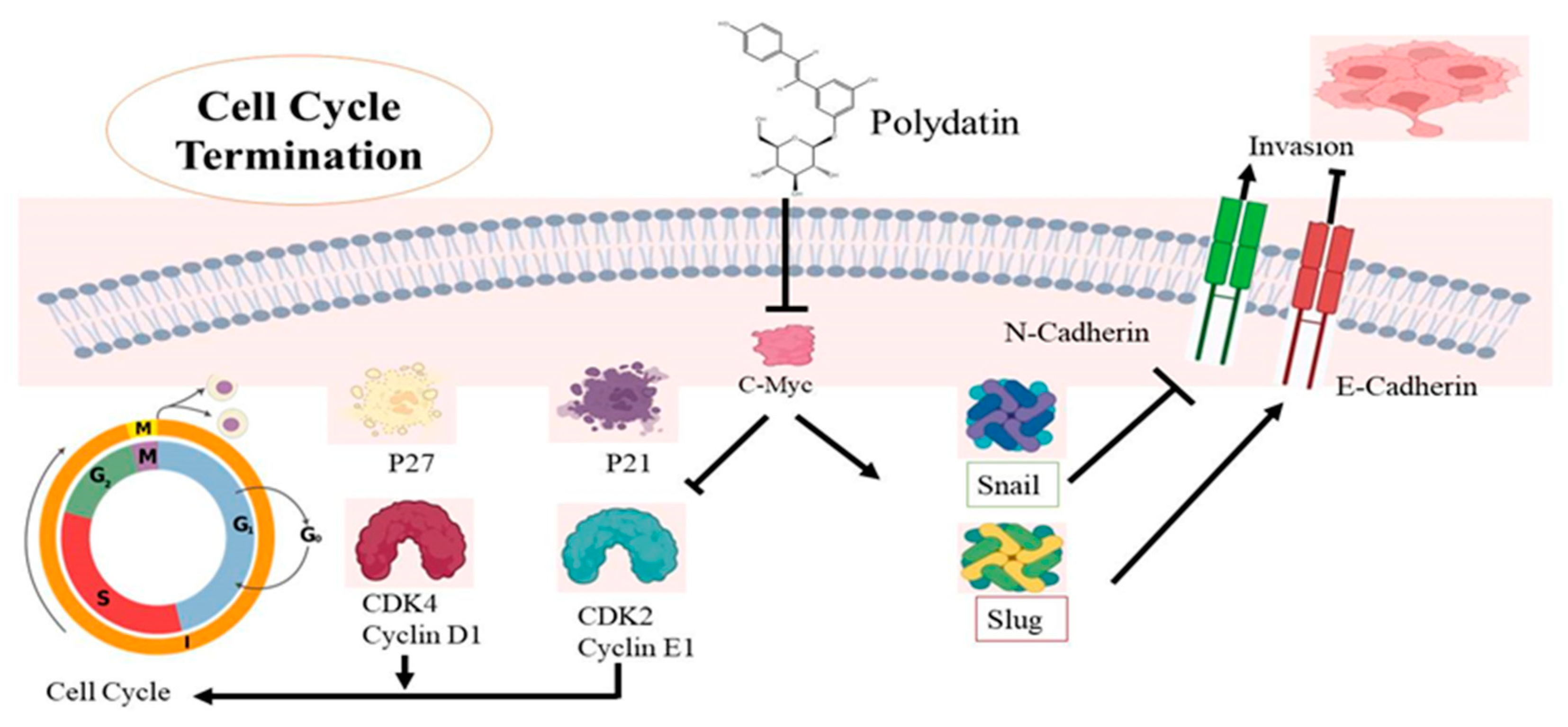
4.5. Cervical Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Cell Line | Type of Study | Concentrations of PD | Molecular Targets | Mechanism of Action | References |
|---|---|---|---|---|---|---|
| Breast cancer | MDA-MB-231 MCF-7 | In vitro | 2, 4, 6 µM | ↑ p38 ↑ JUN ↑ ERK ↑ AKT | Promotes apoptosis by MAPK/ERK & P13K/AKT pathways | [122] |
| 4T1 MCF-7 | In vitro | 5.53 mmol/L 8.67 mmol/L | ↓ p-PI3K/PI3K ↓ p-AKT/AKT | Inhibits P13K/AKT pathways | [80] | |
| Cervical cancer | CaSki C33A | In vitro | 0.1, 10,100, 500 µM | ↑ p21 ↑ p27 ↓ Cdk4 ↓ Cdk2 Cyclin D1 ↓ Cyclin E1 | Inhibits growth promoter proteins and cell cycle arrest | [105] |
| HeLa | In vitro | 50, 100, 150 μmol/L | ↓ PI3K ↓ AKT ↓ mTOR P70S6K ↓ c-Myc | Induced apoptosis by suppression of PI3K/AKT/mTOR signaling | [15] | |
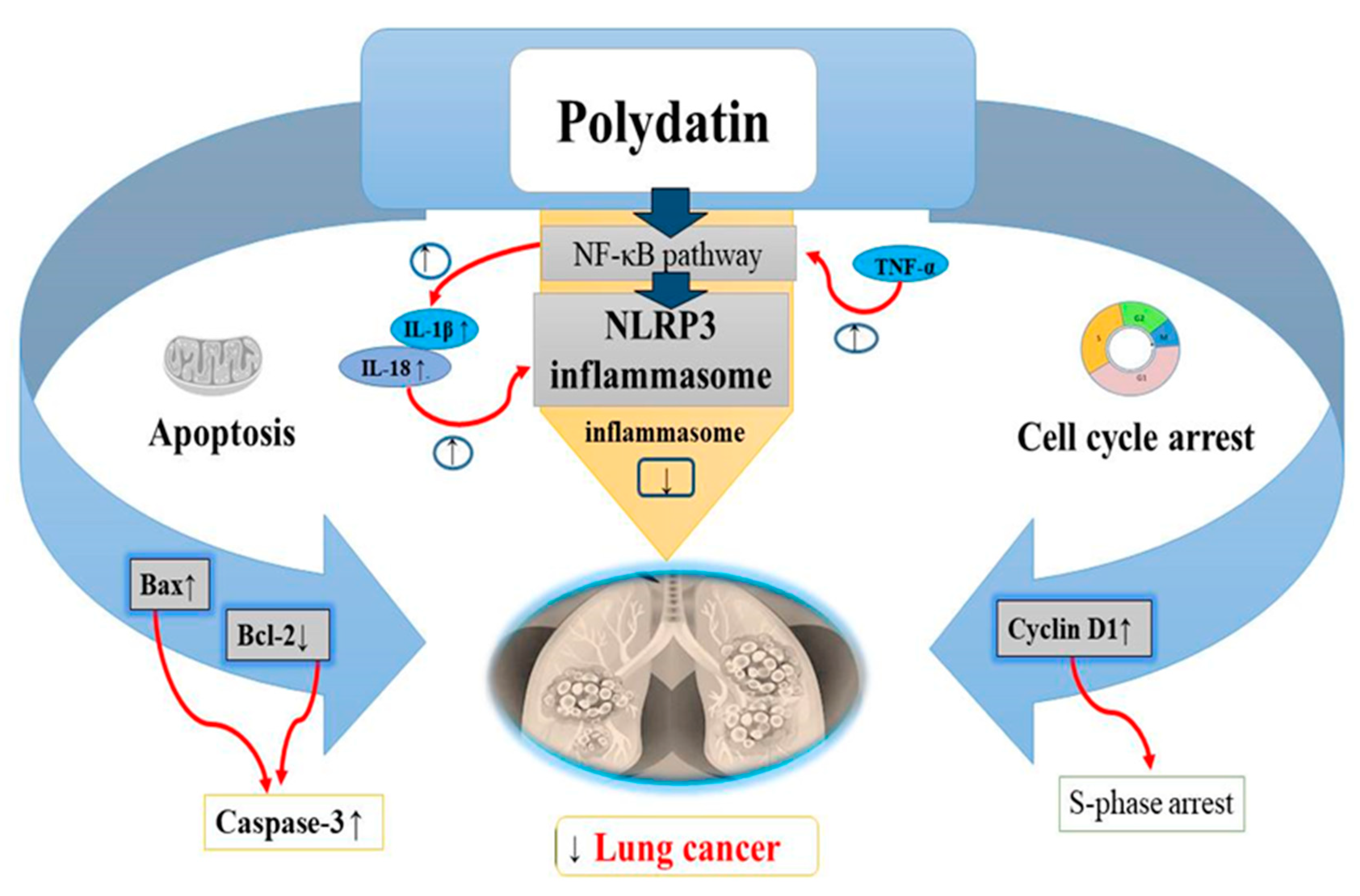
| Lung cancer | A549 NCI-H1975 | In vitro | 6 µ mol/L | ↓ Bcl 2 ↑ Bax ↑ Cyclin D1 | Cell cycle arrest and apoptotic pathway | [123] |
| A549 and H1299 cells | In vitro | ↓ NLRP3 ↓ ASC ↑ pro-caspase-1 ↑ NF-kB ↑ p56 | Promotes apoptosis and NLRP3 inflammasome inhibition by NF-kB | [20] | ||
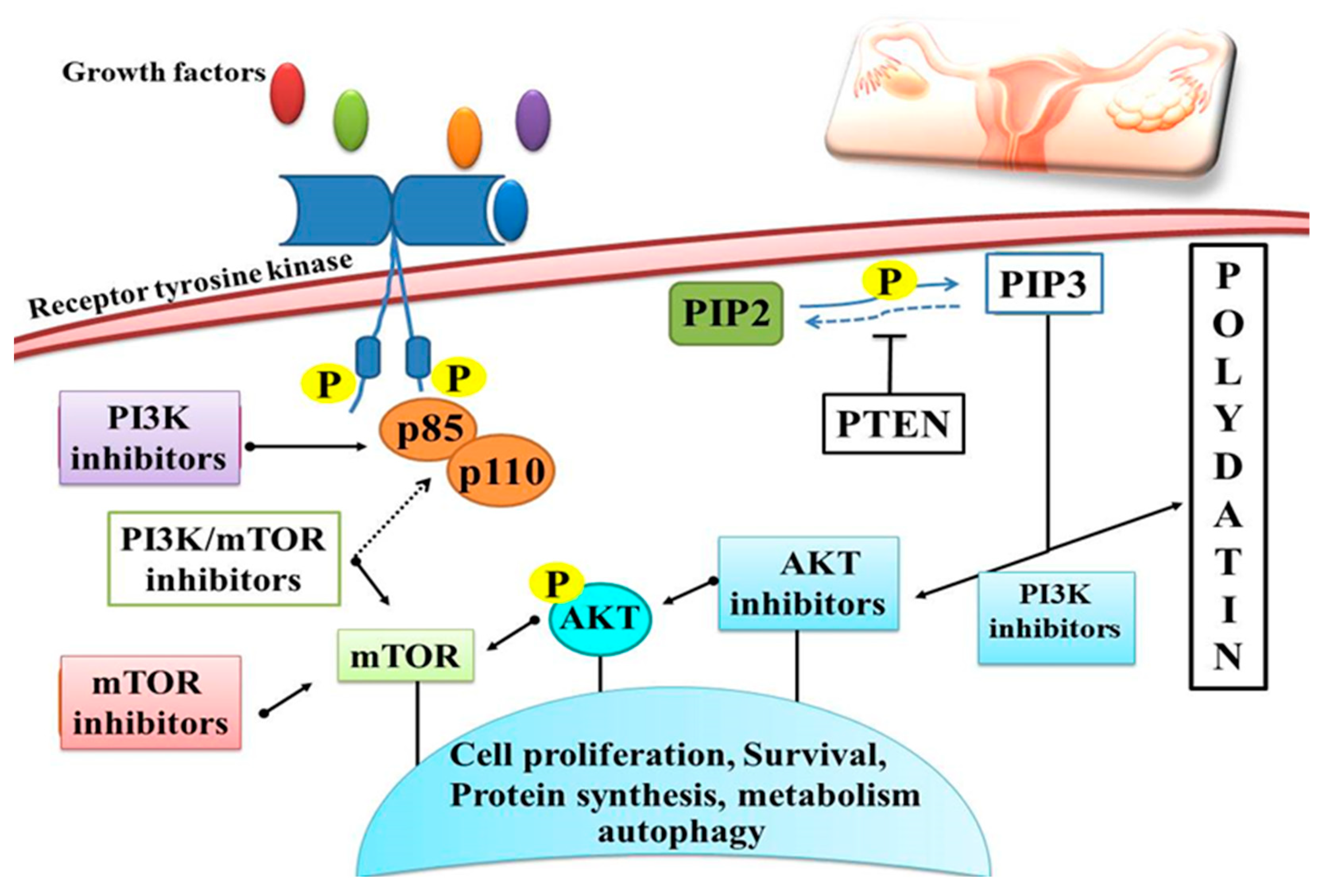
| Ovarian cancer | OVCAR-3, A2780, and HO-8910 | In vitro | 50 μM | ↑ P13K ↑ AKT | AKT signaling | [124] |
| SKOV-3 and OVCAR-8 | In vitro | 5, 10, 50, 100 μM | ↓ Her-2 ↓ EGFR ↓ VEGF ↑ ERK ↑ PARP-1 | Down/upregulation of various cell signaling molecules | [125] | |
| Liver cancer | HCC cells | In vitro | 100 μM 150 μM | ↓G2/M Phase ↓ STAT3 ↓ AKT ↓ JAK1 | Cell cycle arrest JAK1/STAT3 and P13K/AKT signaling | [53] |
| HepG2 SMMC-7721 | In vitro | 1, 3, 10, 30, and 100 µM | ↓ β-catenin ↓ Bcl 2 ↑ Bax ↑ Caspase-3 ↑ Caspase-9 | Apoptotic pathway | [80] | |
| HepG2 | In Vitro | (10, 30, and 100 μM) | ↓ Bcl 2 ↑ Bax ↓ Wnt | Wnt signaling Apoptotic pathway | [54] | |
| Colon carcinoma | CaCo-2 | In vitro | 1–50 μM | ↓ DNA synthesis ↓G0/G1 | Cell cycle arrest | [56] |
| Caco-2 | In vitro | 100 240 μM | ↓ AKT ↑ PARP ↓ Erk-1 ↓ Erk-2 | Regulation of Akt/PKB signaling | [55] | |
| Human myeloma cells | RPMI 8226 | In vitro | 50, 100, 200 μmol/L | ↑ Caspase-3 ↑ Caspase-9 ↑ Bax ↓ mTOR/p70s6k | Apoptotic pathway | [58] |
| Osteosarcoma cells | 143B MG63 | In vitro | 10, 30, 100 μM | ↑ Caspase -3 ↓ Bcl 2 ↑ Bax ↓ β-catenin | Regulation of Apoptotic pathway | [63] |
| Lukemia cells | MOLT-4 | In vitro | 1, 4 or 20 µM | ↓ Cyclin D1 ↓ CYCLIN B1 ↓ Bcl2 | Cell cycle arrest and apoptotic pathway | [126] |
| Nasal carcinoma | CNE | In vitro | 5, 10, 20 µM | ↓ AKT ↑ Endoplasmic ↑ Reticulum stress ↑ Caspase 3 ↑ Caspase 4 ↑ Caspase 9 | Regulation of apoptotic pathway molecules | [127] |
| Laryngeal cancer | AMC-HN-8 cells | In vitro | 2, 4, 6 µM | ↓ PDGF-B ↓ Ki67 ↓ Bcl 2 ↑ Bax ↓ Akt | Regulation of apoptotic pathway and Akt signaling molecules | [128] |
4.6. Lung Cancer
4.7. Ovarian Cancer
5. Underlying Polydatin Anticancer Mechanisms of Action
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- National Cancer Institute. What Is Cancer. Available online: https://www.cancer.gov/about-cancer/understanding/what-is-cancer (accessed on 5 May 2021).
- WHO Cancer Prevalence. Available online: https://www.who.int/news-room/fact-sheets/detail/breast-cancer#:~:text=In%202020%2C%20there%20were%202.3,the%20world's%20most%20prevalent%20cancer (accessed on 26 March 2021).
- WHO, Cervical Cancer Prevalence. Available online: https://www.who.int/news-room/fact-sheets/detail/cervical-cancer (accessed on 22 February 2022).
- Medscape, Non Small Lung Cancer. Available online: https://emedicine.medscape.com/article/279960-overview (accessed on 19 August 2022).
- SEER Cancer Stat Facts: Ovarian Cancer. National Cancer Institute. Bethesda, MD, USA. Available online: https://seer.cancer.gov/statfacts/html/ovary.html (accessed on 3 August 2022).
- Breast Cancer: Types of Treatment. Available online: https://www.cancer.net/cancer-types/breast-cancer/types-treatment (accessed on 19 October 2021).
- Meegan, M.J.; O’Boyle, N.M. Special Issue “Anticancer Drugs”. Pharmaceuticals 2019, 12, 134. [Google Scholar] [CrossRef] [PubMed]
- Sarfraz, A.; Javeed, M.; Shah, M.A.; Hussain, G.; Shafiq, N.; Sarfraz, I.; Riaz, A.; Sadiqa, A.; Zara, R.; Zafar, S. Biochanin A: A novel bioactive multifunctional compound from nature. Sci. Total Environ. 2020, 722, 137907. [Google Scholar] [CrossRef] [PubMed]
- Veeresham, C. Natural products derived from plants as a source of drugs. J. Adv. Pharm. Technol. Res. 2012, 3, 200. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.-H.; Peng, C.; Zhang, H. Polydatin: A review of pharmacology and pharmacokinetics. Pharm Biol 2013, 51, 1347–1354. [Google Scholar] [CrossRef]
- Jeandet, P.; Delaunois, B.; Conreux, A.; Donnez, D.; Nuzzo, V.; Cordelier, S.; Clément, C.; Courot, E. Biosynthesis, metabolism, molecular engineering, and biological functions of stilbene phytoalexins in plants. Biofactors 2010, 36, 331–341. [Google Scholar] [CrossRef]
- Jeandet, P.; Vannozzi, A.; Sobarzo-Sánchez, E.; Uddin, M.S.; Bru, R.; Martínez-Márquez, A.; Clément, C.; Cordelier, S.; Manayi, A.; Nabavi, S.F. Phytostilbenes as agrochemicals: Biosynthesis, bioactivity, metabolic engineering and biotechnology. Nat. Prod. Rep. 2021, 38, 1282–1329. [Google Scholar] [CrossRef]
- Jeandet, P.; Sobarzo-Sánchez, E.; Uddin, M.S.; Bru, R.; Clément, C.; Jacquard, C.; Nabavi, S.F.; Khayatkashani, M.; Batiha, G.E.-S.; Khan, H. Resveratrol and cyclodextrins, an easy alliance: Applications in nanomedicine, green chemistry and biotechnology. Biotechnol Adv. 2021, 53, 107844. [Google Scholar] [CrossRef]
- Quan, Z.; Gu, J.; Dong, P.; Lu, J.; Wu, X.; Wu, W.; Fei, X.; Li, S.; Wang, Y.; Wang, J. Reactive oxygen species-mediated endoplasmic reticulum stress and mitochondrial dysfunction contribute to cirsimaritin-induced apoptosis in human gallbladder carcinoma GBC-SD cells. Cancer Lett. 2010, 295, 252–259. [Google Scholar] [CrossRef]
- Pan, J.-H.; Wang, H.-B.; Du, X.-F.; Liu, J.-Y.; Zhang, D.-J. Polydatin induces human cervical cancer cell apoptosis via PI3K/AKT/mTOR signaling pathway. Zhongguo Zhongyao Zazhi—China J. Chin. Mater. Med. 2017, 42, 2345–2349. [Google Scholar]
- DeSalvo, J.; Kuznetsov, J.N.; Du, J.; Leclerc, G.M.; Leclerc, G.J.; Lampidis, T.J.; Barredo, J.C. Inhibition of Akt potentiates 2-DG–Induced apoptosis via downregulation of UPR in acute lymphoblastic leukemia. Mol. Cancer Res. 2012, 10, 969–978. [Google Scholar] [CrossRef]
- Estañ, M.C.; Calviño, E.; de Blas, E.; del Carmen Boyano-Adánez, M.; Mena, M.L.; Gómez-Gómez, M.; Rial, E.; Aller, P. 2-Deoxy-D-glucose cooperates with arsenic trioxide to induce apoptosis in leukemia cells: Involvement of IGF-1R-regulated Akt/mTOR, MEK/ERK and LKB-1/AMPK signaling pathways. Biochem. Pharmacol. 2012, 84, 1604–1616. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Peng, J.; Huang, K.; Huang, J.; Shen, X.; Liu, P.; Huang, H. Polydatin ameliorates experimental diabetes-induced fibronectin through inhibiting the activation of NF-κB signaling pathway in rat glomerular mesangial cells. Mol. Cell. Endocrinol. 2012, 362, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Mikulski, D.; Molski, M. Quantitative structure–antioxidant activity relationship of trans-resveratrol oligomers, trans-4, 4′-dihydroxystilbene dimer, trans-resveratrol-3-O-glucuronide, glucosides: Trans-piceid, cis-piceid, trans-astringin and trans-resveratrol-4′-O-β-D-glucopyranoside. Eur. J. Med. Chem. 2010, 45, 2366–2380. [Google Scholar]
- Zou, J.; Yang, Y.; Yang, Y.; Liu, X. Polydatin suppresses proliferation and metastasis of non-small cell lung cancer cells by inhibiting NLRP3 inflammasome activation via NF-κB pathway. Biomed. Pharm. 2018, 108, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Xing, W.-W.; Wu, J.-Z.; Jia, M.; Du, J.; Zhang, H.; Qin, L.-P. Effects of polydatin from Polygonum cuspidatum on lipid profile in hyperlipidemic rabbits. Biomed. Pharm. 2009, 63, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Choi, O.; Lee, J.K.; Kang, S.-Y.; Pandey, R.P.; Sohng, J.-K.; Ahn, J.S.; Hong, Y.-S. Construction of artificial biosynthetic pathways for resveratrol glucoside derivatives. J. Microbiol. Biotechnol. 2014, 24, 614–618. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Grigorchuk, V.P.; Ogneva, Z.V.; Suprun, A.; Dubrovina, A. Stilbene biosynthesis in the needles of spruce Picea jezoensis. Phytochemistry 2016, 131, 57–67. [Google Scholar] [CrossRef]
- Cichewicz, R.H.; Kouzi, S.A. Biotransformation of resveratrol to piceid by Bacillus cereus. J. Nat. Prod. 1998, 61, 1313–1314. [Google Scholar] [CrossRef]
- Ozaki, S.-i.; Imai, H.; Iwakiri, T.; Sato, T.; Shimoda, K.; Nakayama, T.; Hamada, H. Regioselective glucosidation of trans-resveratrol in Escherichia coli expressing glucosyltransferase from Phytolacca americana. Biotechnol. Lett. 2012, 34, 475–481. [Google Scholar] [CrossRef]
- Choi, O.; Wu, C.-Z.; Kang, S.Y.; Ahn, J.S.; Uhm, T.-B.; Hong, Y.-S. Biosynthesis of plant-specific phenylpropanoids by construction of an artificial biosynthetic pathway in Escherichia coli. J. Ind. Microbiol. Biotechnol. 2011, 38, 1657–1665. [Google Scholar] [CrossRef]
- Regev-Shoshani, G.; Shoseyov, O.; Bilkis, I.; Kerem, Z. Glycosylation of resveratrol protects it from enzymic oxidation. Biochem. J. 2003, 374, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Jiang, K.; Wu, H.; Qiu, C.; Deng, G.; Peng, X. Polydatin reduces Staphylococcus aureus lipoteichoic acid-induced injury by attenuating reactive oxygen species generation and TLR 2-NF κB signalling. J. Cell. Mol. Med. 2017, 21, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, X.; Liu, G.; Li, J.; Zhang, J.; Cao, Y.; Miao, J. Antioxidant activity and mechanism of resveratrol and polydatin isolated from mulberry (Morus alba L.). Molecules 2021, 26, 7574. [Google Scholar] [CrossRef]
- Galeano-Díaz, T.; Durán-Merás, I.; Airado-Rodríguez, D. Isocratic chromatography of resveratrol and piceid after previous generation of fluorescent photoproducts: Wine analysis without sample preparation. J. Sep. Sci. 2007, 30, 3110–3119. [Google Scholar] [CrossRef] [PubMed]
- Kammerer, D.; Claus, A.; Carle, R.; Schieber, A. Polyphenol screening of pomace from red and white grape varieties (Vitis vinifera L.) by HPLC-DAD-MS/MS. J. Agric. Food Chem. 2004, 52, 4360–4367. [Google Scholar] [CrossRef] [PubMed]
- Romero-Perez, A.I.; Ibern-Gomez, M.; Lamuela-Raventos, R.M.; de La Torre-Boronat, M.C. Piceid, the major resveratrol derivative in grape juices. J. Agric. Food Chem. 1999, 47, 1533–1536. [Google Scholar] [CrossRef]
- Peng, X.L.; Qu, W.; Wang, L.Z.; Huang, B.Q.; Ying, C.J.; Sun, X.F.; Hao, L.P. Resveratrol ameliorates high glucose and high-fat/sucrose diet-induced vascular hyperpermeability involving Cav-1/eNOS regulation. PLoS ONE 2014, 9, e113716. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Andres-Lacueva, C.; Lamuela-Raventós, R.M.; Berenguer, T.; Jakszyn, P.; Martínez, C.; Sanchez, M.J.; Navarro, C.; Chirlaque, M.D.; Tormo, M.-J. Concentrations of resveratrol and derivatives in foods and estimation of dietary intake in a Spanish population: European Prospective Investigation into Cancer and Nutrition (EPIC)-Spain cohort. Br. J. Nutr. 2008, 100, 188–196. [Google Scholar] [CrossRef]
- Peng, X.L.; Xu, J.; Sun, X.F.; Ying, C.J.; Hao, L.P. Analysis of trans-resveratrol and trans-piceid in vegetable foods using high-performance liquid chromatography. Int. J. Food Sci. Nutr. 2015, 66, 729–735. [Google Scholar] [CrossRef]
- Chiva-Blanch, G.; Urpi-Sarda, M.; Rotchés-Ribalta, M.; Zamora-Ros, R.; Llorach, R.; Lamuela-Raventós, R.M.; Estruch, R.; Andrés-Lacueva, C. Determination of resveratrol and piceid in beer matrices by solid-phase extraction and liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2011, 1218, 698–705. [Google Scholar] [CrossRef]
- Jensen, J.S.; Wertz, C.F.; O’Neill, V.A. Preformulation stability of trans-resveratrol and trans-resveratrol glucoside (piceid). J. Agric. Food Chem. 2010, 58, 1685–1690. [Google Scholar] [CrossRef] [PubMed]
- Şöhretoğlu, D.; Baran, M.Y.; Arroo, R.; Kuruüzüm-Uz, A. Recent advances in chemistry, therapeutic properties and sources of polydatin. Phytochem. Rev. 2018, 17, 973–1005. [Google Scholar] [CrossRef]
- Ibern-Gomez, M.; Roig-Perez, S.; Lamuela-Raventos, R.M.; de la Torre-Boronat, M.C. Resveratrol and piceid levels in natural and blended peanut butters. J. Agric. Food Chem. 2000, 48, 6352–6354. [Google Scholar] [CrossRef] [PubMed]
- Hurst, W.J.; Glinski, J.A.; Miller, K.B.; Apgar, J.; Davey, M.H.; Stuart, D.A. Survey of the trans-resveratrol and trans-piceid content of cocoa-containing and chocolate products. J. Agric. Food Chem. 2008, 56, 8374–8378. [Google Scholar] [CrossRef]
- Xu, L.-Q.; Xie, Y.-L.; Gui, S.-H.; Zhang, X.; Mo, Z.-Z.; Sun, C.-Y.; Li, C.-L.; Luo, D.-D.; Zhang, Z.-B.; Su, Z.-R. Polydatin attenuates d-galactose-induced liver and brain damage through its anti-oxidative, anti-inflammatory and anti-apoptotic effects in mice. Food Funct. 2016, 7, 4545–4555. [Google Scholar] [CrossRef]
- Su, D.; Cheng, Y.; Liu, M.; Liu, D.; Cui, H.; Zhang, B.; Zhou, S.; Yang, T.; Mei, Q. Comparision of piceid and resveratrol in antioxidation and antiproliferation activities in vitro. PLoS ONE 2013, 8, e54505. [Google Scholar] [CrossRef]
- Yousef, A.I.; Shawki, H.H.; El-Shahawy, A.A.; El-Twab, S.M.A.; Abdel-Moneim, A.; Oishi, H. Polydatin mitigates pancreatic β-cell damage through its antioxidant activity. Biomed. Pharmacother. 2021, 133, 111027. [Google Scholar] [CrossRef]
- Aggarwal, S. Targeted cancer therapies. Nat. Rev. Drug Discov. 2010, 9, 427. [Google Scholar] [CrossRef]
- Zhang, H.; Park, S.; Huang, H.; Kim, E.; Yi, J.; Choi, S.-K.; Ryoo, Z.; Kim, M. Anticancer effects and potential mechanisms of ginsenoside Rh2 in various cancer types. Oncol. Rep. 2021, 45, 1–10. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Y.; Mao, Z.; Yu, D.; Gao, C. Toxicity of ZnO nanoparticles to macrophages due to cell uptake and intracellular release of zinc ions. J. Nanosci. Nanotechnol. 2014, 14, 5688–5696. [Google Scholar] [CrossRef]
- Hong, C.; Wang, D.; Liang, J.; Guo, Y.; Zhu, Y.; Xia, J.; Qin, J.; Zhan, H.; Wang, J. Novel ginsenoside-based multifunctional liposomal delivery system for combination therapy of gastric cancer. Theranostics 2019, 9, 4437. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Wan, J.Y.; Zeng, J.; Huang, W.H.; Sava-Segal, C.; Li, L.; Niu, X.; Wang, Q.; Wang, C.Z.; Yuan, C.S. Effects of compound K, an enteric microbiome metabolite of ginseng, in the treatment of inflammation associated colon cancer. Oncol. Lett. 2018, 15, 8339–8348. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.-S.; Jia, M.; Chen, L.; Zhu, B.; Dong, H.-X.; Si, J.-P.; Peng, W.; Han, T. Cytotoxic and antifungal constituents isolated from the metabolites of endophytic fungus DO14 from Dendrobium officinale. Molecules 2015, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Trautwein, C.; Friedman, S.L.; Schuppan, D.; Pinzani, M. Hepatic fibrosis: Concept to treatment. J. Hepatol. 2015, 62, S15–S24. [Google Scholar] [CrossRef]
- Nakamoto, Y. Promising new strategies for hepatocellular carcinoma. Hepatol. Res. 2017, 47, 251–265. [Google Scholar] [CrossRef]
- Jiang, J.; Chen, Y.; Dong, T.; Yue, M.; Zhang, Y.; An, T.; Zhang, J.; Liu, P.; Yang, X. Polydatin inhibits hepatocellular carcinoma via the AKT/STAT3-FOXO1 signaling pathway. Oncol. Lett. 2019, 17, 4505–4513. [Google Scholar] [CrossRef]
- Jiao, Y.; Wu, Y.; Du, D. Polydatin inhibits cell proliferation, invasion and migration, and induces cell apoptosis in hepatocellular carcinoma. Braz. J. Med. Biol. Res. 2018, 51. [Google Scholar] [CrossRef]
- De Maria, S.; Scognamiglio, I.; Lombardi, A.; Amodio, N.; Caraglia, M.; Cartenì, M.; Ravagnan, G.; Stiuso, P. Polydatin, a natural precursor of resveratrol, induces cell cycle arrest and differentiation of human colorectal Caco-2 cell. J. Transl. Med. 2013, 11, 1–11. [Google Scholar] [CrossRef]
- Storniolo, C.E.; Quifer-Rada, P.; Lamuela-Raventos, R.M.; Moreno, J.J. Piceid presents antiproliferative effects in intestinal epithelial Caco-2 cells, effects unrelated to resveratrol release. Food Funct. 2014, 5, 2137–2144. [Google Scholar] [CrossRef]
- Bae, H.; Lee, W.; Song, J.; Hong, T.; Kim, M.H.; Ham, J.; Song, G.; Lim, W. Polydatin Counteracts 5-Fluorouracil Resistance by Enhancing Apoptosis via Calcium Influx in Colon Cancer. Antioxidants 2021, 10, 1477. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Zhao, S. Polydatin regulates proliferation, apoptosis and autophagy in multiple myeloma cells through mTOR/p70s6k pathway. Onco Targets. 2017, 10, 935. [Google Scholar] [CrossRef] [PubMed]
- Taran, S.J.; Taran, R.; Malipatil, N.B. Pediatric osteosarcoma: An updated review. Indian J. Med. Paediatr. Oncol. 2017, 38, 33–43. [Google Scholar] [CrossRef]
- Misaghi, A.; Goldin, A.; Awad, M.; Kulidjian, A.A. Osteosarcoma: A comprehensive review. Sicot-J 2018, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Wang, W.; He, J.; Jiang, L.; Li, X. Osteopontin as a biomarker for osteosarcoma therapy and prognosis. Oncol. Lett. 2019, 17, 2592–2598. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, C.P.; Kukekov, V.G.; Reith, J.D.; Tchigrinova, O.; Suslov, O.N.; Scott, E.W.; Ghivizzani, S.C.; Ignatova, T.N.; Steindler, D.A. Stem-like cells in bone sarcomas: Implications for tumorigenesis. Neoplasia 2005, 7, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Kuang, G.; Jiang, W.; Jiang, R.; Jiang, D. Polydatin promotes apoptosis through upregulation the ratio of Bax/Bcl-2 and inhibits proliferation by attenuating the β-catenin signaling in human osteosarcoma cells. Am. J. Transl. Res. 2016, 8, 922. [Google Scholar]
- Luce, A.; Lama, S.; Millan, P.C.; Itro, A.; Sangiovanni, A.; Caputo, C.; Ferranti, P.; Cappabianca, S.; Caraglia, M.; Stiuso, P. Polydatin Induces Differentiation and Radiation Sensitivity in Human Osteosarcoma Cells and Parallel Secretion through Lipid Metabolite Secretion. Oxid. Med. Cell. Longev. 2021, 2021, 3337013. [Google Scholar] [CrossRef]
- United States Cancer Statistics (USCS). Prevention, Global Cancer Statistics. 2012. 2019. Available online: https://www.cdc.gov/cancer/uscs/index.htm (accessed on 3 August 2022).
- Ginsburg, O.; Yip, C.H.; Brooks, A.; Cabanes, A.; Caleffi, M.; Dunstan Yataco, J.A.; Gyawali, B.; McCormack, V.; McLaughlin de Anderson, M.; Mehrotra, R. Breast cancer early detection: A phased approach to implementation. Cancer 2020, 126, 2379–2393. [Google Scholar] [CrossRef]
- Schwartz, G.K.; Shah, M.A. Targeting the cell cycle: A new approach to cancer therapy. J. Clin. Oncol. 2005, 23, 9408–9421. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Cao, J.; Lin, W.; Chen, H.; Xiong, X.; Ao, H.; Yu, M.; Lin, J.; Cui, Q. The roles of cyclin-dependent kinases in cell-cycle progression and therapeutic strategies in human breast cancer. Int. J. Mol. Sci. 2020, 21, 1960. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.P.; Zhu, Y.-L.; Ratner, E.S. Targeting cyclin-dependent kinases for treatment of gynecologic cancers. Front. Oncol. 2018, 8, 303. [Google Scholar] [CrossRef] [PubMed]
- Kuo, S.-H.; Wei, M.-F.; Lee, Y.-H.; Yang, W.-C.; Yang, S.-Y.; Lin, J.-C.; Huang, C.-S. MAP3K1 Expression Is Associated with Progression and Poor Prognosis of Hormone Receptor-Positive, HER2-Negative Early-Stage Breast Cancer. 2020. Available online: https://assets.researchsquare.com/files/rs-53956/v1/20512c99-6481-480b-9d9e-37881a993bb9.pdf?c=1631849978 (accessed on 3 August 2022).
- Mayr, B.; Montminy, M. Transcriptional regulation by the phosphorylation-dependent factor CREB. Nat. Rev. Mol. Cell Biol. 2001, 2, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, A.; Fernando, H.; Watkins, G.; Mansel, R.E.; Jiang, W.G. Expression of transcription factor CREB1 in human breast cancer and its correlation with prognosis. Oncol. Rep. 2007, 18, 953–958. [Google Scholar] [CrossRef]
- Zhang, X.; Odom, D.T.; Koo, S.-H.; Conkright, M.D.; Canettieri, G.; Best, J.; Chen, H.; Jenner, R.; Herbolsheimer, E.; Jacobsen, E. Genome-wide analysis of cAMP-response element binding protein occupancy, phosphorylation, and target gene activation in human tissues. Proc. Natl. Acad. Sci. 2005, 102, 4459–4464. [Google Scholar] [CrossRef]
- Alao, J.P. The regulation of cyclin D1 degradation: Roles in cancer development and the potential for therapeutic invention. Mol. Cancer 2007, 6, 1–16. [Google Scholar] [CrossRef]
- Hartmann, N.B.; Rist, S.; Bodin, J.; Jensen, L.H.; Schmidt, S.N.; Mayer, P.; Meibom, A.; Baun, A. Microplastics as vectors for environmental contaminants: Exploring sorption, desorption, and transfer to biota. Integr. Environ. Assess. Manag. 2017, 13, 488–493. [Google Scholar] [CrossRef]
- Jin, Y.; Huynh, D.T.N.; Nguyen, T.L.L.; Jeon, H.; Heo, K.S. Therapeutic effects of ginsenosides on breast cancer growth and metastasis. Arch. Pharmacal Res. 2020, 43, 773–787. [Google Scholar] [CrossRef]
- Orgaz, J.L.; Pandya, P.; Dalmeida, R.; Karagiannis, P.; Sanchez-Laorden, B.; Viros, A.; Albrengues, J.; Nestle, F.O.; Ridley, A.J.; Gaggioli, C. Diverse matrix metalloproteinase functions regulate cancer amoeboid migration. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef]
- Chabottaux, V.; Noel, A. Breast cancer progression: Insights into multifaceted matrix metalloproteinases. Clin. Exp. Metastasis 2007, 24, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.V.; Chang, J.P.; Parachoniak, C.A.; Pandika, M.M.; Aghi, M.K.; Meyronet, D.; Isachenko, N.; Fouse, S.D.; Phillips, J.J.; Cheresh, D.A. VEGF inhibits tumor cell invasion and mesenchymal transition through a MET/VEGFR2 complex. Cancer Cell 2012, 22, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhu, X.; Wu, H.; Jiang, K.; Zhao, G.; Shaukat, A.; Deng, G.; Qiu, C. Targeting the ROS/PI3K/AKT/HIF-1alpha/HK2 axis of breast cancer cells: Combined administration of Polydatin and 2-Deoxy-d-glucose. J. Cell. Mol. Med. 2019, 23, 3711–3723. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Song, D.Y.; Hwang, J.J.; Park, H.J.; Lee, J.S.; Song, S.Y.; Jeong, S.-Y.; Choi, E.K. Induction of p53-mediated senescence is essential for the eventual anticancer therapeutic effect of RH1. Arch. Pharm Res. 2019, 42, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-Salinas, F.L.; Delgado-Magallón, A.; Montes-Alvarado, J.B.; Ramírez-Ramírez, D.; Flores-Alonso, J.C.; Cortés-Hernández, P.; Reyes-Leyva, J.; Herrera-Camacho, I.; Anaya-Ruiz, M.; Pelayo, R. Breast cancer subtypes present a differential production of reactive oxygen species (ROS) and susceptibility to antioxidant treatment. Front. Oncol. 2019, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- Morgan, M.J.; Liu, Z.-G. Crosstalk of reactive oxygen species and NF-κB signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef]
- de Sá Junior, P.L.; Câmara, D.A.D.; Porcacchia, A.S.; Fonseca, P.M.M.; Jorge, S.D.; Araldi, R.P.; Ferreira, A.K. The roles of ROS in cancer heterogeneity and therapy. Oxidative Med. Cell. Longev. 2017, 2017, 2467940. [Google Scholar] [CrossRef]
- Kubli, S.P.; Bassi, C.; Roux, C.; Wakeham, A.; Göbl, C.; Zhou, W.; Jafari, S.M.; Snow, B.; Jones, L.; Palomero, L. AhR controls redox homeostasis and shapes the tumor microenvironment in BRCA1-associated breast cancer. Proc. Natl. Acad. Sci. USA 2019, 116, 3604–3613. [Google Scholar] [CrossRef]
- Han, B.; Liu, W.; Li, J.; Wang, J.; Zhao, D.; Xu, R.; Lin, Z. Catalytic hydrodechlorination of triclosan using a new class of anion-exchange-resin supported palladium catalysts. Water Res. 2017, 120, 199–210. [Google Scholar] [CrossRef]
- Chen, L.; Lan, Z.; Lin, Q.; Mi, X.; He, Y.; Wei, L.; Lin, Y.; Zhang, Y.; Deng, X. Polydatin ameliorates renal injury by attenuating oxidative stress-related inflammatory responses in fructose-induced urate nephropathic mice. Food Chem. Toxicol. 2013, 52, 28–35. [Google Scholar] [CrossRef]
- Checker, R.; Gambhir, L.; Sharma, D.; Kumar, M.; Sandur, S.K. Plumbagin induces apoptosis in lymphoma cells via oxidative stress mediated glutathionylation and inhibition of mitogen-activated protein kinase phosphatases (MKP1/2). Cancer Lett. 2015, 357, 265–278. [Google Scholar] [CrossRef]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Dwarakanath, B. Cytotoxicity, radiosensitization, and chemosensitization of tumor cells by 2-deoxy-D-glucose in vitro. J. Cancer Res. Ther. 2009, 5, 27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, J.; Wang, F.; Hu, J.; Wang, S.; Sun, Y. 2-Deoxy-D-glucose targeting of glucose metabolism in cancer cells as a potential therapy. Cancer Lett. 2014, 355, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A.; Agnihotri, S.; Micallef, J.; Mukherjee, J.; Sabha, N.; Cairns, R.; Hawkins, C.; Guha, A. Hexokinase 2 is a key mediator of aerobic glycolysis and promotes tumor growth in human glioblastoma multiforme. J. Exp. Med. 2011, 208, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Shafaee, A.; Dastyar, D.Z.; Islamian, J.P.; Hatamian, M. Inhibition of tumor energy pathways for targeted esophagus cancer therapy. Metabolism 2015, 64, 1193–1198. [Google Scholar] [CrossRef]
- Palazon, A.; Tyrakis, P.A.; Macias, D.; Veliça, P.; Rundqvist, H.; Fitzpatrick, S.; Vojnovic, N.; Phan, A.T.; Loman, N.; Hedenfalk, I. An HIF-1α/VEGF-A axis in cytotoxic T cells regulates tumor progression. Cancer Cell 2017, 32, 669–683.e5. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Dai, W.; Zhang, Q.; Feng, J.; Wu, L.; Liu, T.; Yu, Q.; Xu, S.; Wang, W. Genistein suppresses aerobic glycolysis and induces hepatocellular carcinoma cell death. Br. J. Cancer 2017, 117, 1518–1528. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA A Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef]
- Vaccarella, S.; Laversanne, M.; Ferlay, J.; Bray, F. Cervical cancer in a frica, L atin a merica and the C aribbean and a sia: Regional inequalities and changing trends. Int. J. Can. 2017, 141, 1997–2001. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Ma, D. The precision prevention and therapy of HPV-related cervical cancer: New concepts and clinical implications. Can. Med. 2018, 7, 5217–5236. [Google Scholar] [CrossRef] [PubMed]
- Den Boon, J.A.; Pyeon, D.; Wang, S.S.; Horswill, M.; Schiffman, M.; Sherman, M.; Zuna, R.E.; Wang, Z.; Hewitt, S.M.; Pearson, R. Molecular transitions from papillomavirus infection to cervical precancer and cancer: Role of stromal estrogen receptor signaling. Proc. Natl. Acad. Sci. USA 2015, 112, E3255–E3264. [Google Scholar] [CrossRef] [PubMed]
- Marquina, G.; Manzano, A.; Casado, A. Targeted agents in cervical cancer: Beyond bevacizumab. Curr. Oncol. Rep. 2018, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Lowe, G.; Roberts, C.M.; Finlay, J.; Han, E.S.; Glackin, C.A.; Dellinger, T.H. Pterostilbene suppresses ovarian cancer growth via induction of apoptosis and blockade of cell cycle progression involving inhibition of the STAT3 pathway. Int J. Mol. Sci. 2018, 19, 1983. [Google Scholar] [CrossRef]
- Yu, H.; Pan, C.; Zhao, S.; Wang, Z.; Zhang, H.; Wu, W. Resveratrol inhibits tumor necrosis factor-α-mediated matrix metalloproteinase-9 expression and invasion of human hepatocellular carcinoma cells. Biomed. Pharmacother. 2008, 62, 366–372. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, Y.; Di, S.; Feng, X.; Liu, D.; Jiang, S.; Hu, W.; Qin, Z.; Li, Y.; Lv, J. Pterostilbene exerts anticancer activity on non-small-cell lung cancer via activating endoplasmic reticulum stress. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Bai, L.; Ma, Y.; Wang, X.; Feng, Q.; Zhang, Z.; Wang, S.; Zhang, H.; Lu, X.; Xu, Y.; Zhao, E. Polydatin inhibits cell viability, migration, and invasion through suppressing the c-Myc expression in human cervical cancer. Front. Cell Dev. Biol. 2021, 9, 587218. [Google Scholar] [CrossRef]
- Nishioka, R.; Itoh, S.; Gui, T.; Gai, Z.; Oikawa, K.; Kawai, M.; Tani, M.; Yamaue, H.; Muragaki, Y. SNAIL induces epithelial-to-mesenchymal transition in a human pancreatic cancer cell line (BxPC3) and promotes distant metastasis and invasiveness in vivo. Exp. Mol. Pathol. 2010, 89, 149–157. [Google Scholar] [CrossRef]
- Cercelaru, L.; Stepan, A.E.; Mărgăritescu, C.; Osman, A.; Popa, I.-C.; Florescu, M.M.; Simionescu, C.E.; Mărgăritescu, O.C. E-cadherin, β-catenin and Snail immunoexpression in laryngeal squamous cell carcinoma. Rom. J. Morphol. Embryol. 2017, 58, 761–766. [Google Scholar]
- Dhasarathy, A.; Phadke, D.; Mav, D.; Shah, R.R.; Wade, P.A. The transcription factors Snail and Slug activate the transforming growth factor-beta signaling pathway in breast cancer. PLoS ONE 2011, 6, e26514. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Guo, W.-J.; Yao, R.-X. Advances research on C-MYC proto-oncogene in multiple myeloma-review. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2016, 24, 1248–1251. [Google Scholar] [PubMed]
- Wu, S.-H.; Zeng, X.-F.; Wang, P.; Zhou, Y.; Lin, W. The Expression and Significance of c-myc and bcat1 in Cervical Cancer. Djournal Sichuan Univ. 2018, 49, 725–730. [Google Scholar]
- Gao, K.; Eurasian, M.; Zhang, J.; Wei, Y.; Zheng, Q.; Ye, H.; Li, L. Can genomic amplification of human telomerase gene and C-MYC in liquid-based cytological specimens be used as a method for opportunistic cervical cancer screening? Gynecol. Obstet. Investig. 2015, 80, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Zhao, E.; Ding, J.; Xia, Y.; Liu, M.; Ye, B.; Choi, J.-H.; Yan, C.; Dong, Z.; Huang, S.; Zha, Y. KDM4C and ATF4 cooperate in transcriptional control of amino acid metabolism. Cell Rep. 2016, 14, 506–519. [Google Scholar] [CrossRef] [PubMed]
- Nowak, D.G.; Cho, H.; Herzka, T.; Watrud, K.; DeMarco, D.V.; Wang, V.M.; Senturk, S.; Fellmann, C.; Ding, D.; Beinortas, T. MYC drives Pten/Trp53-deficient proliferation and metastasis due to IL6 secretion and AKT suppression via PHLPP2. Cancer Discov. 2015, 5, 636–651. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, H.; Qing, G. Targeting oncogenic Myc as a strategy for cancer treatment. Signal. Transduct. Target. Ther. 2018, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Musgrove, E.A.; Lee, C.; Sutherland, R.L. Progestins both stimulate and inhibit breast cancer cell cycle progression while increasing expression of transforming growth factor alpha, epidermal growth factor receptor, c-fos, and c-myc genes. Mol. Cell. Biol. 1991, 11, 5032–5043. [Google Scholar]
- García-Gutiérrez, L.; Delgado, M.D.; León, J. MYC oncogene contributions to release of cell cycle brakes. Genes 2019, 10, 244. [Google Scholar] [CrossRef]
- Finn, R.S.; Aleshin, A.; Slamon, D.J. Targeting the cyclin-dependent kinases (CDK) 4/6 in estrogen receptor-positive breast cancers. Breast Cancer Res. 2016, 18, 1–11. [Google Scholar] [CrossRef]
- Thomasova, D.; Anders, H.-J. Cell cycle control in the kidney. Nephrol. Dial. Transplant. 2015, 30, 1622–1630. [Google Scholar] [CrossRef] [PubMed]
- Karimian, A.; Ahmadi, Y.; Yousefi, B. Multiple functions of p21 in cell cycle, apoptosis and transcriptional regulation after DNA damage. Dna Repair 2016, 42, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Aleem, E.; Kiyokawa, H.; Kaldis, P. Cdc2–cyclin E complexes regulate the G1/S phase transition. Nat. Cell Biol. 2005, 7, 831–836. [Google Scholar] [CrossRef]
- Hsiao, Y.-H.; Lin, C.-W.; Wang, P.-H.; Hsin, M.-C.; Yang, S.-F. The potential of Chinese herbal medicines in the treatment of cervical cancer. Integr. Cancer Ther. 2019, 18, 1534735419861693. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Tao, J.; Zhong, F.; Jiao, Y.; Xu, J.; Shen, Q.; Wang, H.; Fan, S.; Zhang, Y. Polydatin down-regulates the phosphorylation level of Creb and induces apoptosis in human breast cancer cell. PLoS ONE 2017, 12, e0176501. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhuang, Z.; Meng, Q.; Jiao, Y.; Xu, J.; Fan, S. Polydatin inhibits growth of lung cancer cells by inducing apoptosis and causing cell cycle arrest. Oncol Lett 2014, 7, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. Effects of polydatin on the proliferation, migration, and invasion of ovarian cancer. Biocell 2019, 43, 313. [Google Scholar] [CrossRef]
- Hogg, S.J.; Chitcholtan, K.; Hassan, W.; Sykes, P.H.; Garrill, A. Resveratrol, acetyl-resveratrol, and polydatin exhibit antigrowth activity against 3D cell aggregates of the SKOV-3 and OVCAR-8 ovarian cancer cell lines. Obs. Gynecol. Int. 2015, 2015. [Google Scholar] [CrossRef]
- Cao, W.J.; Wu, K.; Wang, C.; Wan, D.M. Polydatin-induced cell apoptosis and cell cycle arrest are potentiated by Janus kinase 2 inhibition in leukemia cells. Mol. Med. Rep. 2016, 13, 3297–3302. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, S.; Zhang, Y.; Wu, J.; Peng, H.; Fan, J.; Liao, J. Reactive oxygen species-mediated endoplasmic reticulum stress and mitochondrial dysfunction contribute to polydatin-induced apoptosis in human nasopharyngeal carcinoma CNE cells. J. Cell. Biochem. 2011, 112, 3695–3703. [Google Scholar] [CrossRef]
- Li, H.; Shi, B.; Li, Y.; Yin, F. Polydatin inhibits cell proliferation and induces apoptosis in laryngeal cancer and HeLa cells via suppression of the PDGF/AKT signaling pathway. J. Biochem. Mol. Toxicol. 2017, 31, e21900. [Google Scholar] [CrossRef] [PubMed]
- Spiro, S.G.; Tanner, N.T.; Silvestri, G.A.; Janes, S.M.; Lim, E.; Vansteenkiste, J.F.; Pirker, R. Lung cancer: Progress in diagnosis, staging and therapy. Respirology 2010, 15, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Lemjabbar-Alaoui, H.; Hassan, O.U.; Yang, Y.-W.; Buchanan, P. Lung cancer: Biology and treatment options. Biochim. Biophys. Acta (Bba)—Rev. Cancer 2015, 1856, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Luan, J.; Duan, H.; Liu, Q.; Yagasaki, K.; Zhang, G. Inhibitory effects of norcantharidin against human lung cancer cell growth and migration. Cytotechnology 2010, 62, 349–355. [Google Scholar] [CrossRef]
- Jiang, T.; Tian, F.; Zheng, H.; Whitman, S.A.; Lin, Y.; Zhang, Z.; Zhang, N.; Zhang, D.D. Nrf2 suppresses lupus nephritis through inhibition of oxidative injury and the NF-κB-mediated inflammatory response. Kidney Int. 2014, 85, 333–343. [Google Scholar] [CrossRef]
- Neto, C.C.; Amoroso, J.W.; Liberty, A.M. Anticancer activities of cranberry phytochemicals: An update. Mol. Nutr. Food Res. 2008, 52, S18–S27. [Google Scholar] [CrossRef]
- Verma, N.; Tiku, A.B. Polydatin-induced direct and bystander effects in a549 lung cancer cell line. Nutr. Cancer 2022, 74, 237–249. [Google Scholar] [CrossRef]
- Yang, J.; Liu, X.; Bhalla, K.; Kim, C.N.; Ibrado, A.M.; Cai, J.; Peng, T.-I.; Jones, D.P.; Wang, X. Prevention of apoptosis by Bcl-2: Release of cytochrome c from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar] [CrossRef]
- Kluck, R.M.; Bossy-Wetzel, E.; Green, D.R.; Newmeyer, D.D. The release of cytochrome c from mitochondria: A primary site for Bcl-2 regulation of apoptosis. Science 1997, 275, 1132–1136. [Google Scholar] [CrossRef]
- Graña, X.; Reddy, E.P. Cell cycle control in mammalian cells: Role of cyclins, cyclin dependent kinases (CDKs), growth suppressor genes and cyclin-dependent kinase inhibitors (CKIs). Oncogene 1995, 11, 211–220. [Google Scholar]
- Pavletich, N.P. Mechanisms of cyclin-dependent kinase regulation: Structures of Cdks, their cyclin activators, and Cip and INK4 inhibitors. J. Mol. Biol. 1999, 287, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Hitomi, M.; Stacey, D.W. Variations in cyclin D1 levels through the cell cycle determine the proliferative fate of a cell. Cell Div. 2006, 1, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, J.J.; Ebus, M.E.; Koster, J.; van Sluis, P.; van Noesel, C.J.; Versteeg, R.; Caron, H.N. Cyclin D1 and CDK4 activity contribute to the undifferentiated phenotype in neuroblastoma. Cancer Res. 2008, 68, 2599–2609. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.; Peters, G. Genetic alterations of cyclins, cyclin-dependent kinases, and Cdk inhibitors in human cancer. Adv. Cancer Res. 1996, 68, 67–108. [Google Scholar]
- Liu, Y.-L.; Chen, B.-Y.; Nie, J.; Zhao, G.-H.; Zhuo, J.-Y.; Yuan, J.; Li, Y.-C.; Wang, L.-L.; Chen, Z.-W. Polydatin prevents bleomycin-induced pulmonary fibrosis by inhibiting the TGF-β/Smad/ERK signaling pathway. Exp. Ther. Med. 2020, 20, 1. [Google Scholar] [CrossRef]
- Wang, Y.; Kong, H.; Zeng, X.; Liu, W.; Wang, Z.; Yan, X.; Wang, H.; Xie, W. Activation of NLRP3 inflammasome enhances the proliferation and migration of A549 lung cancer cells. Oncol. Rep. 2016, 35, 2053–2064. [Google Scholar] [CrossRef]
- Howlader, N.; Noone, A.-M.; Krapcho, M.; Garshell, J.; Neyman, N.; Altekruse, S.; Kosary, C.; Yu, M.; Ruhl, J.; Tatalovich, Z. SEER cancer statistics review, 1975–2010. Natl. Cancer Inst. 2014. [Google Scholar]
- Bast, R.C.; Hennessy, B.; Mills, G.B. The biology of ovarian cancer: New opportunities for translation. Nat. Rev. Cancer 2009, 9, 415–428. [Google Scholar] [CrossRef]
- Cheaib, B.; Auguste, A.; Leary, A. The PI3K/Akt/mTOR pathway in ovarian cancer: Therapeutic opportunities and challenges. Chin. J. Cancer 2015, 34, 4–16. [Google Scholar] [CrossRef]
- Leary, A.; Auclin, E.; Pautier, P.; Lhommé, C. The PI3K/Akt/mTOR pathway in ovarian cancer: Biological rationale and therapeutic opportunities. Ovarian Cancer—A Clin. Transl. Update 2013, 275–302. [Google Scholar]
- Mabuchi, S.; Kuroda, H.; Takahashi, R.; Sasano, T. The PI3K/AKT/mTOR pathway as a therapeutic target in ovarian cancer. Gynecol. Oncol. 2015, 137, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Ye, P.; Wu, H.; Jiang, Y.; Xiao, X.; Song, D.; Xu, N.; Ma, X.; Zeng, J.; Guo, Y. Old dog, new tricks: Polydatin as a multitarget agent for current diseases. Phytother. Res. 2022, 36, 214–230. [Google Scholar] [CrossRef] [PubMed]
- Varoni, E.M.; Lo Faro, A.F.; Sharifi-Rad, J.; Iriti, M. Anticancer molecular mechanisms of resveratrol. Front. Nutr. 2016, 3, 8. [Google Scholar] [CrossRef]
- Diaz-Gerevini, G.T.; Repossi, G.; Dain, A.; Tarres, M.C.; Das, U.N.; Eynard, A.R. Beneficial action of resveratrol: How and why? Nutrition 2016, 32, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Kubota, T.; Uemura, Y.; Kobayashi, M.; Taguchi, H. Combined effects of resveratrol and paclitaxel on lung cancer cells. Anticancer Res. 2003, 23, 4039–4046. [Google Scholar]
- Shankar, S.; Chen, Q.; Siddiqui, I.; Sarva, K.; Srivastava, R.K. Sensitization of TRAIL-resistant LNCaP cells by resveratrol (3,4′,5-tri-hydroxystilbene): Molecular mechanisms and therapeutic potential. J. Mol. Signal. 2007, 2, 1–17. [Google Scholar] [CrossRef]








| Plant | Plant Organ | Solvent for Extraction | Polydatin Quantity (µg/100 g) | References |
|---|---|---|---|---|
| Banana | Fruit | Methanol | 1.70 µg/100g | [35] |
| Lychee | Fruit | Methanol | 1.00 µg/100g | [35] |
| Pomegranate | Fruit | Methanol | 7.56 µg/100g | [35] |
| Waterchestnut | Fruit | Methanol | 0.50 µg/100g | [35] |
| Waxapple | Fruit | Methanol | 1.58 µg/100g | [35] |
| Tomato | Fruit | Methanol | 4.22 µg/100g | [35] |
| Chili pepper | Fruit | Methanol | 14.47 µg/100g | [35] |
| Bell pepper | Fruit | Methanol | 36.22 µg/100g | [35] |
| Grape | Fruit | Methanol | 71.54 µg/100g | [35] |
| Gauva | Fruit | Methanol | 0.72 µg/100g | [35] |
| Lemon | Fruit | Methanol | 17.00 µg/100g | [35] |
| Strawberry | Fruit | Methanol | 100 µg/100g | [35] |
| Orange | Fruit | Methanol | 5.31 µg/100g | [35] |
| Mushroom | Fruit | Methanol | 2.16 µg/100g | [35] |
| Pear | Fruit | Methanol | 13.10 µg/100g | [35] |
| Soya bean | Fruit | Methanol | 42.58 µg/100g | [35] |
| Gallic bulb | Fruit | Methanol | 2.00 µg/100g | [35] |
| Chrysanthemum | Fruit | Methanol | 5.20 µg/100g | [35] |
| Coca | Fruit | Methanol | 7.56 µg/100g | [35] |
| White dammar | Leaves | Diethly ether | 0.22 mg/g | [38] |
| Peanut | Seeds | Ethanol | 0..128 µg/100g | [39] |
| Cocoa | Seeds | Hexane | 7.14 µg/g | [40] |
| Norway spruce | Phloem | Methanol | 16 mg/g | [23] |
| Norway spruce | Bark | Methanol | 1.3 mg/g | [23] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, M.A.; Hamid, A.; Faheem, H.I.; Rasul, A.; Baokbah, T.A.S.; Haris, M.; Yousaf, R.; Saleem, U.; Iqbal, S.; Alves, M.S.; et al. Uncovering the Anticancer Potential of Polydatin: A Mechanistic Insight. Molecules 2022, 27, 7175. https://doi.org/10.3390/molecules27217175
Shah MA, Hamid A, Faheem HI, Rasul A, Baokbah TAS, Haris M, Yousaf R, Saleem U, Iqbal S, Alves MS, et al. Uncovering the Anticancer Potential of Polydatin: A Mechanistic Insight. Molecules. 2022; 27(21):7175. https://doi.org/10.3390/molecules27217175
Chicago/Turabian StyleShah, Muhammad Ajmal, Ayesha Hamid, Hafiza Ishmal Faheem, Azhar Rasul, Tourki A. S. Baokbah, Muhammad Haris, Rimsha Yousaf, Uzma Saleem, Shabnoor Iqbal, Maria Silvana Alves, and et al. 2022. "Uncovering the Anticancer Potential of Polydatin: A Mechanistic Insight" Molecules 27, no. 21: 7175. https://doi.org/10.3390/molecules27217175
APA StyleShah, M. A., Hamid, A., Faheem, H. I., Rasul, A., Baokbah, T. A. S., Haris, M., Yousaf, R., Saleem, U., Iqbal, S., Alves, M. S., Khan, Z., Hussain, G., Alsharfi, I., Khan, H., & Jeandet, P. (2022). Uncovering the Anticancer Potential of Polydatin: A Mechanistic Insight. Molecules, 27(21), 7175. https://doi.org/10.3390/molecules27217175