
Isolation of Chalcomoracin as a Potential α-Glycosidase Inhibitor from Mulberry Leaves and Its Binding Mechanism
Abstract
1. Introduction
2. Result and Discussion
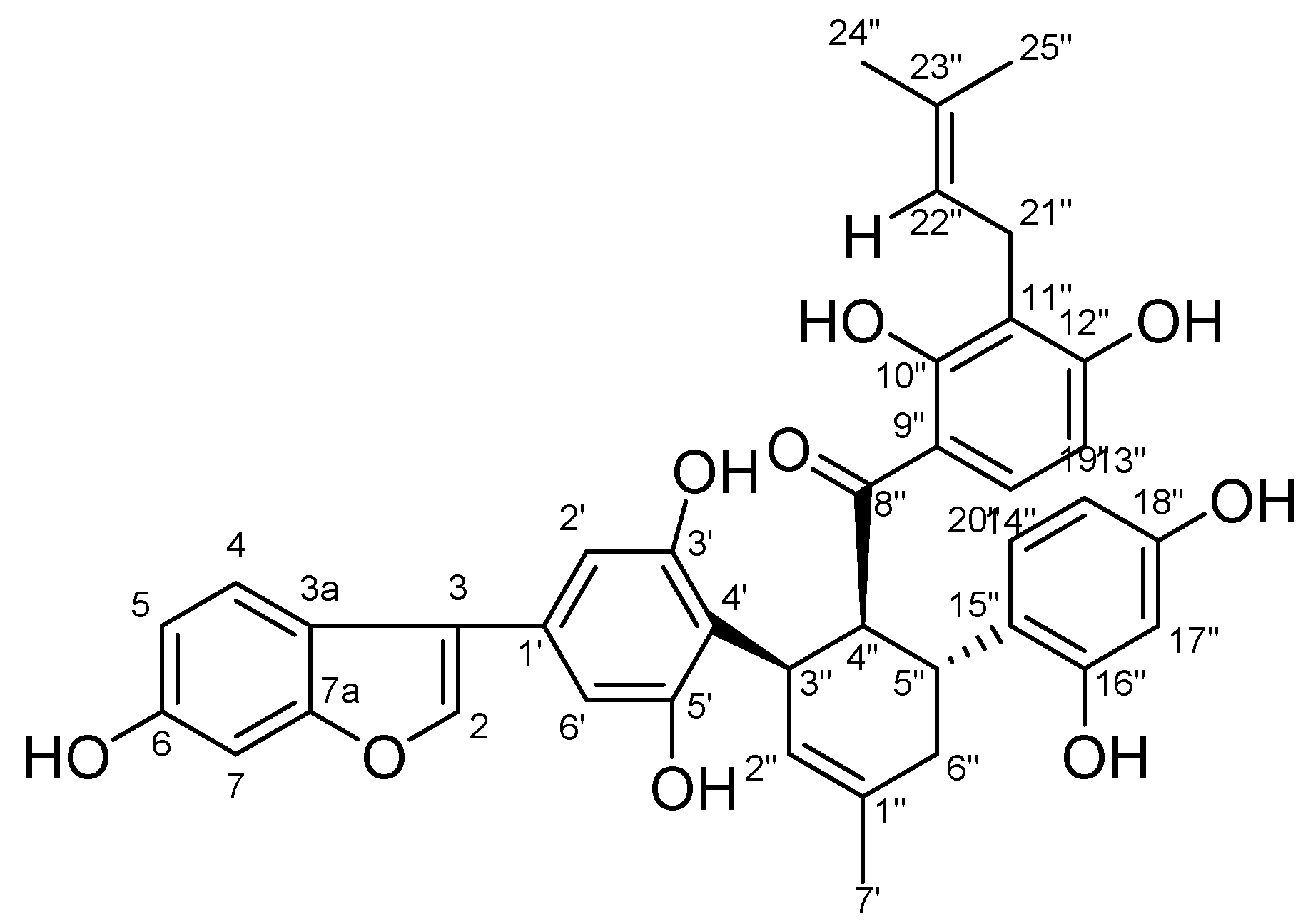
2.1. Structure Identification of Chalcomoracin
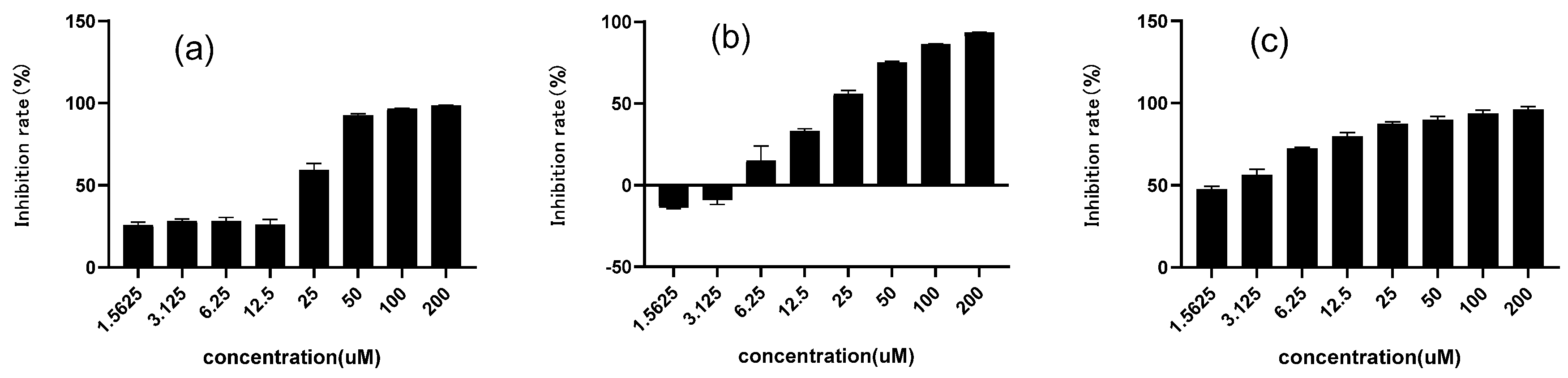
2.2. Inhibitory Effect on α-Glucosidase of Chalcomoracin
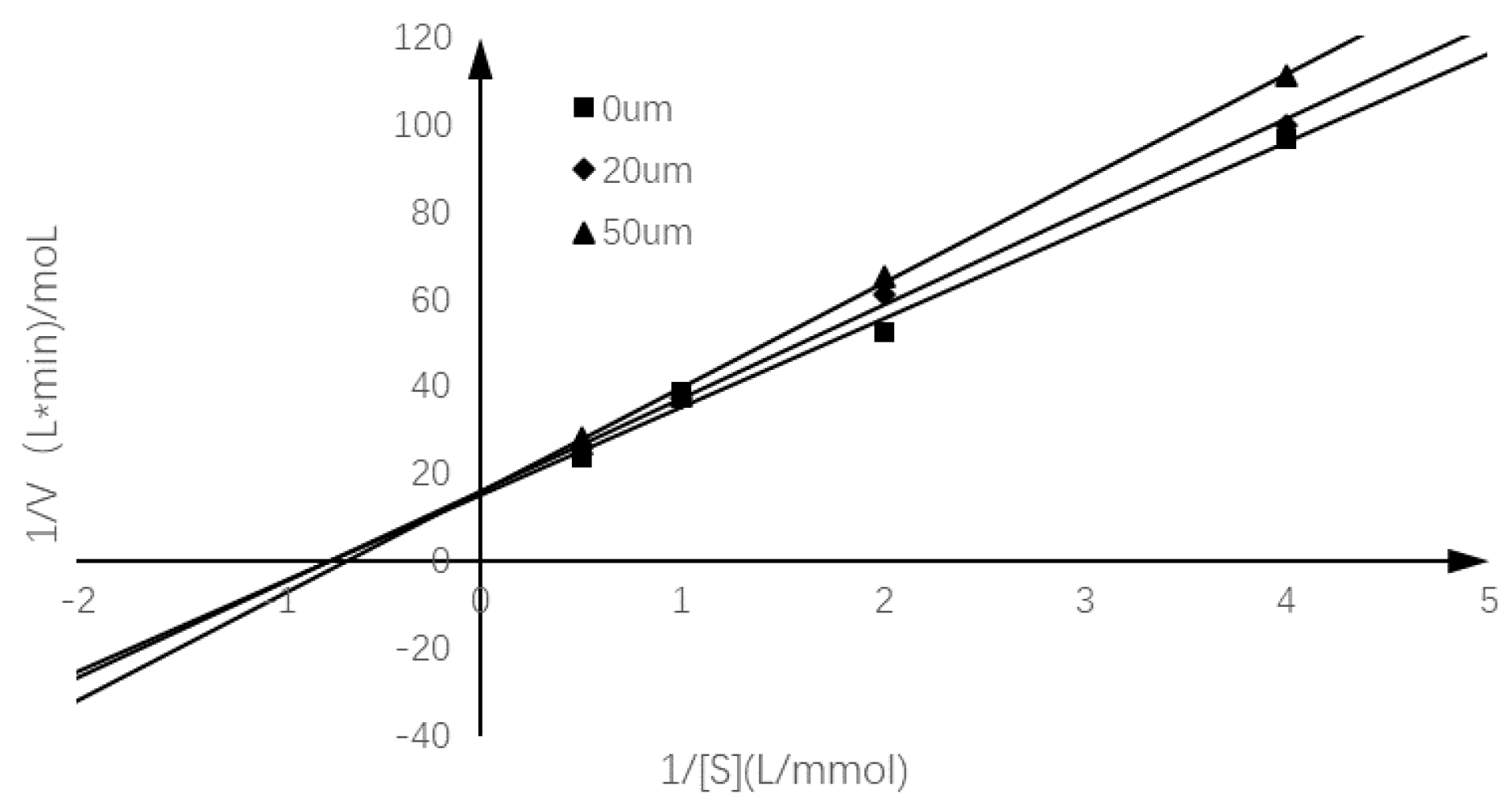
2.3. Inhibition Mode of Chalcomoracin on α-Glucosidase
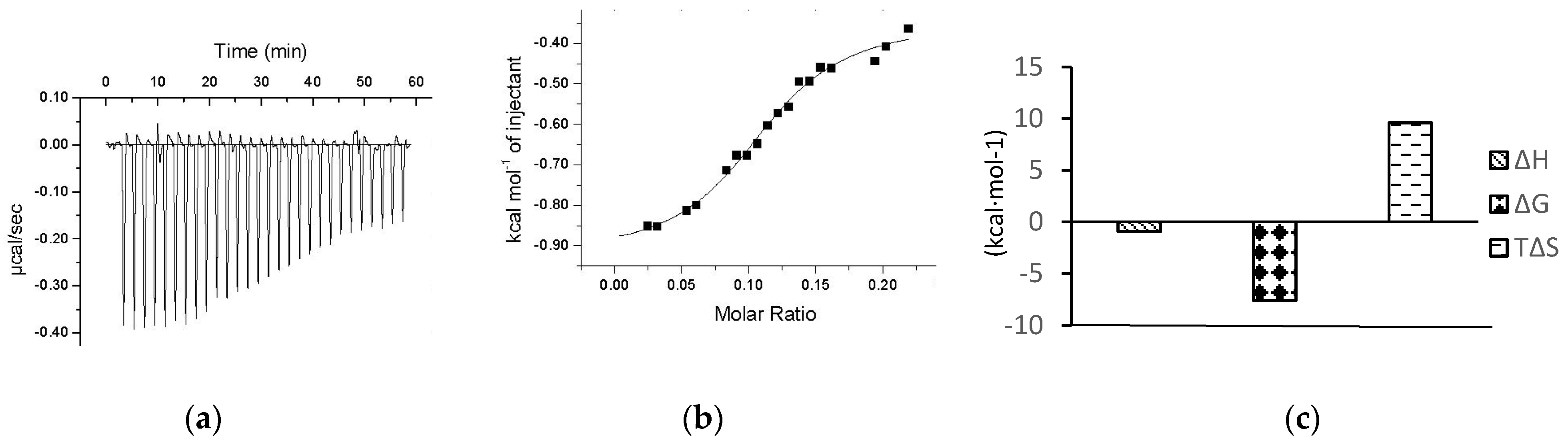
2.4. ITC Determination of Interaction between Chalcomoracin and α-Glucosidase
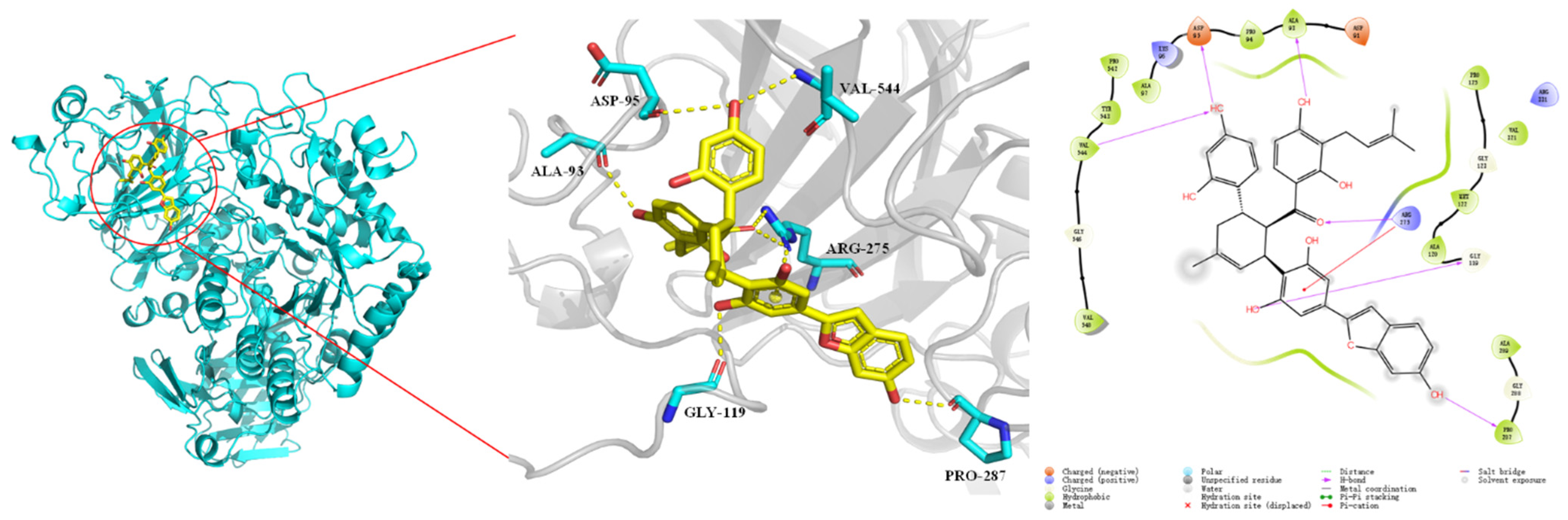
2.5. Binding Prediction by Molecular Docking Simulation
3. Materials and Methods
3.1. Materials and Instruments
3.2. Isolation of Chalcomoracin and Structure Identification
3.3. α-Glucosidase Inhibition Assay
3.4. Interaction Kinetics of α-Glucosidase Inhibition
3.5. Determination of Thermodynamic Parameters
3.6. Molecular Docking Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Adeghate, E.; Schattner, P.; Dunn, E. An Update on the Etiology and Epidemiology of Diabetes Mellitus. Ann. N. Y. Acad. Sci. 2006, 1084, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, M.; Rahmaninan, M.; Mirzaei, M.; Nadjarzadeh, A.; Dehghani tafti, A.A. Epidemiology of diabetes mellitus, pre-diabetes, undiagnosed and uncontrolled diabetes in Central Iran: Results from Yazd health study. BMC Public Health 2020, 20, 166. [Google Scholar] [CrossRef] [PubMed]
- Gujral, U.P.; Kanaya, A.M. Epidemiology of diabetes among South Asians in the United States: Lessons from the MASALA study. Ann. N. Y. Acad. Sci. 2021, 1495, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Trellu, M.; Lacombe, S.; Morin, D.; Dalla-Vale, F. Registre épidémiologique du diabète chez l’enfant en Languedoc-Roussillon. Arch. De Pédiatrie 2015, 22, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Koye, D.N.; Magliano, D.J.; Nelson, R.G.; Pavkov, M.E. The Global Epidemiology of Diabetes and Kidney Disease. Adv. Chronic Kidney Dis. 2018, 25, 121–132. [Google Scholar] [CrossRef]
- Everett, E.; Mathioudakis, N. Update on management of diabetic foot ulcers. Ann. N. Y. Acad. Sci. 2018, 1411, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Uricoechea, H.; Casas-Figueroa, L.Á. Epidemiología de la diabetes mellitus en Sudamérica: La experiencia de Colombia. Clínica E Investig. En Arterioscler. 2016, 28, 245–256. [Google Scholar] [CrossRef]
- Gary-Webb, T.L.; Suglia, S.F.; Tehranifar, P. Social Epidemiology of Diabetes and Associated Conditions. Curr. Diabetes Rep. 2013, 13, 850–859. [Google Scholar] [CrossRef]
- Cheng, H.-T.; Xu, X.; Lim, P.S.; Hung, K.-Y. Worldwide Epidemiology of Diabetes-Related End-Stage Renal Disease, 2000–2015. Diabetes Care 2020, 44, 89–97. [Google Scholar] [CrossRef]
- Ma, R.C.W. Epidemiology of diabetes and diabetic complications in China. Diabetologia 2018, 61, 1249–1260. [Google Scholar] [CrossRef]
- Juang, P.S.; Henry, R.R. Treatment for Type 2 Diabetes. In Endotext [Internet]; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., et al., Eds.; MDText.com, Inc.: Dartmouth, MA, USA, 2013. [Google Scholar]
- Aschner, P. Insulin Therapy in Type 2 Diabetes. Am. J. Ther. 2020, 27, e79–e90. [Google Scholar] [CrossRef] [PubMed]
- Lau, D.C.W. Current and Emerging Pharmacotherapies for Type 2 Diabetes. Can. J. Diabetes 2015, 39, S127–S128. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Padhi, S.; Nayak, A.K.; Behera, A. Type II diabetes mellitus: A review on recent drug based therapeutics. Biomed. Pharmacother. 2020, 131, 110708. [Google Scholar] [CrossRef] [PubMed]
- Ríos, J.L.; Francini, F.; Schinella, G.R. Natural Products for the Treatment of Type 2 Diabetes Mellitus. Planta Med. 2015, 81, 975–994. [Google Scholar] [CrossRef]
- Scheen, A. Pathophysiology of type 2 diabetes. Acta Clin. Belg. 2003, 58, 335. [Google Scholar] [CrossRef]
- Governa, P.; Baini, G.; Borgonetti, V.; Cettolin, G.; Giachetti, D.; Magnano, A.R.; Miraldi, E.; Biagi, M. Phytotherapy in the Management of Diabetes: A Review. Molecules 2018, 23, 105. [Google Scholar] [CrossRef]
- Kumar, S.; Mittal, A.; Babu, D.; Mittal, A. Herbal Medicines for Diabetes Management and its Secondary Complications. Curr. Diabetes Rev. 2021, 17, 437–456. [Google Scholar] [CrossRef]
- Kang, J.; Chen, R.-Y.; Yu, D.-Q. Five New Diels-Alder Type Adducts from the Stem and Root Bark of Morus mongolica. Planta Med. 2006, 72, 52–59. [Google Scholar] [CrossRef]
- Tan, Y.-X.; Yan, R.-Y.; Wang, H.-Q.; Chen, R.-y.; Yu, D.-Q. Wittiorumins A–F, Antioxidant Diels-Alder-Type Adducts from Morus wittiorum. Planta Med. 2009, 75, 249–255. [Google Scholar] [CrossRef]
- Zhang, S.-R.; Zhang, X.-C.; Liang, J.-F.; Fang, H.-M.; Huang, H.-X.; Zhao, Y.-Y.; Chen, X.-Q.; Ma, S.-L. Chalcomoracin inhibits cell proliferation and increases sensitivity to radiotherapy in human non-small cell lung cancer cells via inducing endoplasmic reticulum stress-mediated paraptosis. Acta Pharmacol. Sin. 2020, 41, 825–834. [Google Scholar] [CrossRef]
- Lyu, K.; Yue, W.; Ran, J.; Liu, Y.; Zhu, X. In vivo therapeutic exploring for Mori folium extract against type 2 diabetes mellitus in rats. Biosci. Rep. 2021, 41, BSR20210977. [Google Scholar] [CrossRef] [PubMed]
- Tam, N.H.D.; Nam, H.N.; Elhady, T.M.; Tran, L.; Hassan, G.O.; Sadik, M.; Tien, T.M.P.; Elshafei, A.G.; Huy, T.N. Effects of Mulberry on The Central Nervous System: A Literature Review. Curr. Neuropharmacol. 2021, 19, 193–219. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Zhou, T.; Jiang, Y.; Gong, L.; Yang, B. Identification of prenylated phenolics in mulberry leaf and their neuroprotective activity. Phytomedicine 2021, 90, 153641. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Hu, Y. Systematic Evaluation of the Mechanisms of Mulberry Leaf (Morus alba Linne) Acting on Diabetes Based on Network Pharmacology and Molecular Docking. Comb. Chem. High Throughput Screen. 2021, 24, 668–682. [Google Scholar] [CrossRef]
- Marchetti, L.; Saviane, A.; Montà, A.d.; Paglia, G.; Pellati, F.; Benvenuti, S.; Bertelli, D.; Cappellozza, S. Determination of 1-Deoxynojirimycin (1-DNJ) in Leaves of Italian or Italy-Adapted Cultivars of Mulberry (Morus sp.pl.) by HPLC-MS. Plants 2021, 10, 1553. [Google Scholar] [CrossRef]
- Han, H.; Chou, C.-C.; Li, R.; Liu, J.; Zhang, L.; Zhu, W.; Hu, J.; Yang, B.; Tian, J. Chalcomoracin is a potent anticancer agent acting through triggering Oxidative stress via a mitophagy- and paraptosis-dependent mechanism. Sci. Rep. 2018, 8, 9566. [Google Scholar] [CrossRef]
- Alaei, L.; Izadi, Z.; Jafari, S.; Jahanshahi, F.; Jaymand, M.; Mohammadi, P.; Paray, B.A.; Hasan, A.; Falahati, M.; Varnamkhasti, B.S.; et al. Irreversible thermal inactivation and conformational lock of alpha glucosidase. J. Biomol. Struct. Dyn. 2021, 39, 3256–3262. [Google Scholar] [CrossRef]
- Bach, T.J.; Lichtenthaler, H.K. Application of modified Lineweaver-Burk plots to studies of kinetics and regulation of radish 3-hydroxy-3-methylglutaryl-CoA reductase. Biochim. Et Biophys. Acta (BBA)-Lipids Lipid Metab. 1984, 794, 152–161. [Google Scholar] [CrossRef]
- Abbasi, M.A.; Zia ur, R.; Aziz ur, R.; Siddiqui, S.Z.; Nazir, M.; Hassan, M.; Raza, H.; Shah, S.A.A.; Seo, S.Y. Synthesis of Bi-Heterocyclic Sulfonamides as Tyrosinase Inhibitors: Lineweaver-Burk Plot Evaluation and Computational Ascriptions. Acta Chim. Slov. 2020, 67, 403–414. [Google Scholar] [CrossRef]
- Vajravijayan, S.; Nandhagopal, N.; Krishnan, D.A.; Gunasekaran, K. Isolation and characterization of an iridoid, Arbortristoside-C from Nyctanthes arbor-tristis Linn., a potential drug candidate for diabetes targeting alpha-glucosidase. J. Biomol. Struct. Dyn. 2022, 40, 337–347. [Google Scholar] [CrossRef]
- Jia, Y.; Gao, X.; Xue, Z.; Wang, Y.; Lu, Y.; Zhang, M.; Panichayupakaranant, P.; Chen, H. Characterization, antioxidant activities, and inhibition on α-glucosidase activity of corn silk polysaccharides obtained by different extraction methods. Int. J. Biol. Macromol. 2020, 163, 1640–1648. [Google Scholar] [CrossRef] [PubMed]
- Jelesarov, I.; Bosshard, H.R. Isothermal titration calorimetry and differential scanning calorimetry as complementary tools to investigate the energetics of biomolecular recognition. J. Mol. Recognit. 1999, 12, 3–18. [Google Scholar] [CrossRef]
- Freyer, M.W.; Lewis, E.A. Isothermal Titration Calorimetry: Experimental Design, Data Analysis, and Probing Macromolecule/Ligand Binding and Kinetic Interactions. In Methods in Cell Biology; Academic Press: Cambridge, MA, USA, 2008; Volume 84, pp. 79–113. [Google Scholar]
- Yang, Z.Z.; Wang, Y.C.; Wang, Y.; Zhang, Y.F. Bioassay-guided screening and isolation of alpha-glucosidase and tyrosinase inhibitors from leaves of Morus alba. Food Chem. 2012, 131, 617–625. [Google Scholar] [CrossRef]
- Sheng, Z.; Dai, H.; Pan, S.; Wang, H.; Hu, Y.; Ma, W. Isolation and Characterization of an α-Glucosidase Inhibitor from Musa spp. (Baxijiao) Flowers. Molecules 2014, 19, 10563–10573. [Google Scholar] [CrossRef] [PubMed]
- Zambelli, B. Characterization of Enzymatic Reactions Using ITC. Microcalorim. Biol. Mol. Methods Protoc. 2019, 1964, 251–266. [Google Scholar] [CrossRef]
- Roig-Zamboni, V.; Cobucci-Ponzano, B.; Iacono, R.; Ferrara, M.C.; Germany, S.; Bourne, Y.; Parenti, G.; Moracci, M.; Sulzenbacher, G. Structure of human lysosomal acid α-glucosidase–a guide for the treatment of Pompe disease. Nat. Commun. 2017, 8, 1111. [Google Scholar] [CrossRef]
- Rahman, N.; Muhammad, I.; Gul-E-Nayab, K.H.; Aschner, M.; Filosa, R.; Daglia, M. Molecular Docking of Isolated Alkaloids for Possible α-Glucosidase Inhibition. Biomolecules 2019, 9, 544. [Google Scholar] [CrossRef]
- He, C.; Liu, X.; Jiang, Z.; Geng, S.; Ma, H.; Liu, B. Interaction Mechanism of Flavonoids and α-Glucosidase: Experimental and Molecular Modelling Studies. Foods 2019, 8, 355. [Google Scholar] [CrossRef]
- Dong, Q.; Hu, N.; Yue, H.; Wang, H. Inhibitory Activity and Mechanism Investigation of Hypericin as a Novel α-Glucosidase Inhibitor. Molecules 2021, 26, 4566. [Google Scholar] [CrossRef]
- Jia, Y.; Xue, Z.; Wang, Y.; Lu, Y.; Li, R.; Li, N.; Wang, Q.; Zhang, M.; Chen, H. Chemical structure and inhibition on α-glucosidase of polysaccharides from corn silk by fractional precipitation. Carbohydr. Polym. 2021, 252, 117185. [Google Scholar] [CrossRef]
- Bastos, M.; Velazquez-Campoy, A. Isothermal titration calorimetry (ITC): A standard operating procedure (SOP). Eur. Biophys. J. 2021, 50, 363–371. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chalcomoracin Concentration | Lineweaver–Burk Equation | R2 | −1/Km | Km | 1/Vmax | Vmax |
|---|---|---|---|---|---|---|
| 0 μM | y = 20.192x + 15.231 | 0.992 | −0.754 | 1.326 | 15.231 | 0.066 |
| 20 μM | y = 21.322x + 15.901 | 0.9964 | −0.746 | 1.341 | 15.901 | 0.063 |
| 50 μM | y = 23.896x + 15.914 | 0.9985 | −0.666 | 1.502 | 15.914 | 0.063 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Zhou, X.; Zhou, D.; Jian, Y.; Jia, J.; Ge, F. Isolation of Chalcomoracin as a Potential α-Glycosidase Inhibitor from Mulberry Leaves and Its Binding Mechanism. Molecules 2022, 27, 5742. https://doi.org/10.3390/molecules27185742
Liu Y, Zhou X, Zhou D, Jian Y, Jia J, Ge F. Isolation of Chalcomoracin as a Potential α-Glycosidase Inhibitor from Mulberry Leaves and Its Binding Mechanism. Molecules. 2022; 27(18):5742. https://doi.org/10.3390/molecules27185742
Chicago/Turabian StyleLiu, Yang, Xue Zhou, Dan Zhou, Yongxing Jian, Jingfu Jia, and Fahuan Ge. 2022. "Isolation of Chalcomoracin as a Potential α-Glycosidase Inhibitor from Mulberry Leaves and Its Binding Mechanism" Molecules 27, no. 18: 5742. https://doi.org/10.3390/molecules27185742
APA StyleLiu, Y., Zhou, X., Zhou, D., Jian, Y., Jia, J., & Ge, F. (2022). Isolation of Chalcomoracin as a Potential α-Glycosidase Inhibitor from Mulberry Leaves and Its Binding Mechanism. Molecules, 27(18), 5742. https://doi.org/10.3390/molecules27185742