
A Novel Potent Crystalline Chitin Decomposer: Chitin Deacetylase from Acinetobacter schindleri MCDA01

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
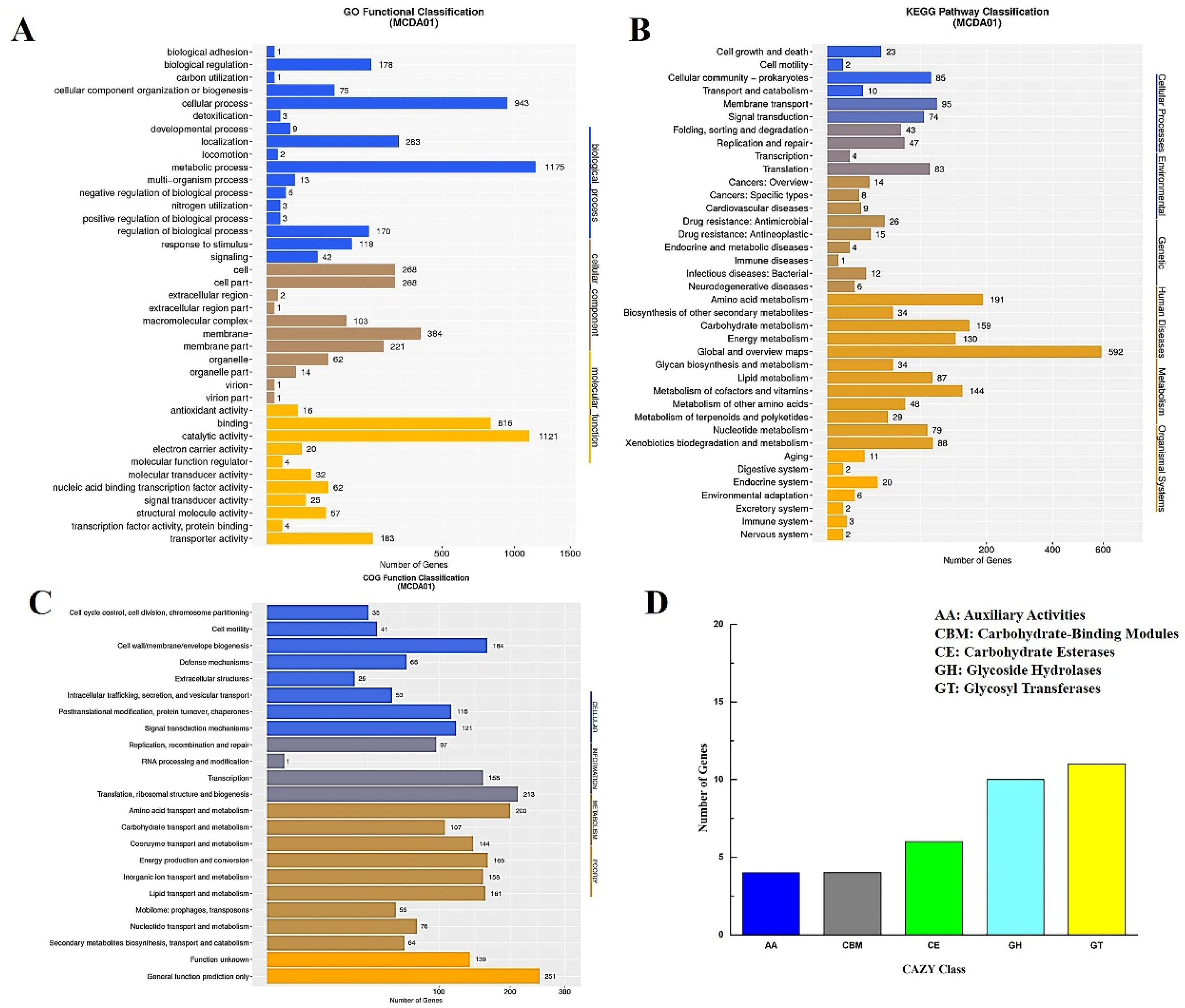
2.1. General Genomic Features of the A. schindleri MCDA01 Strain
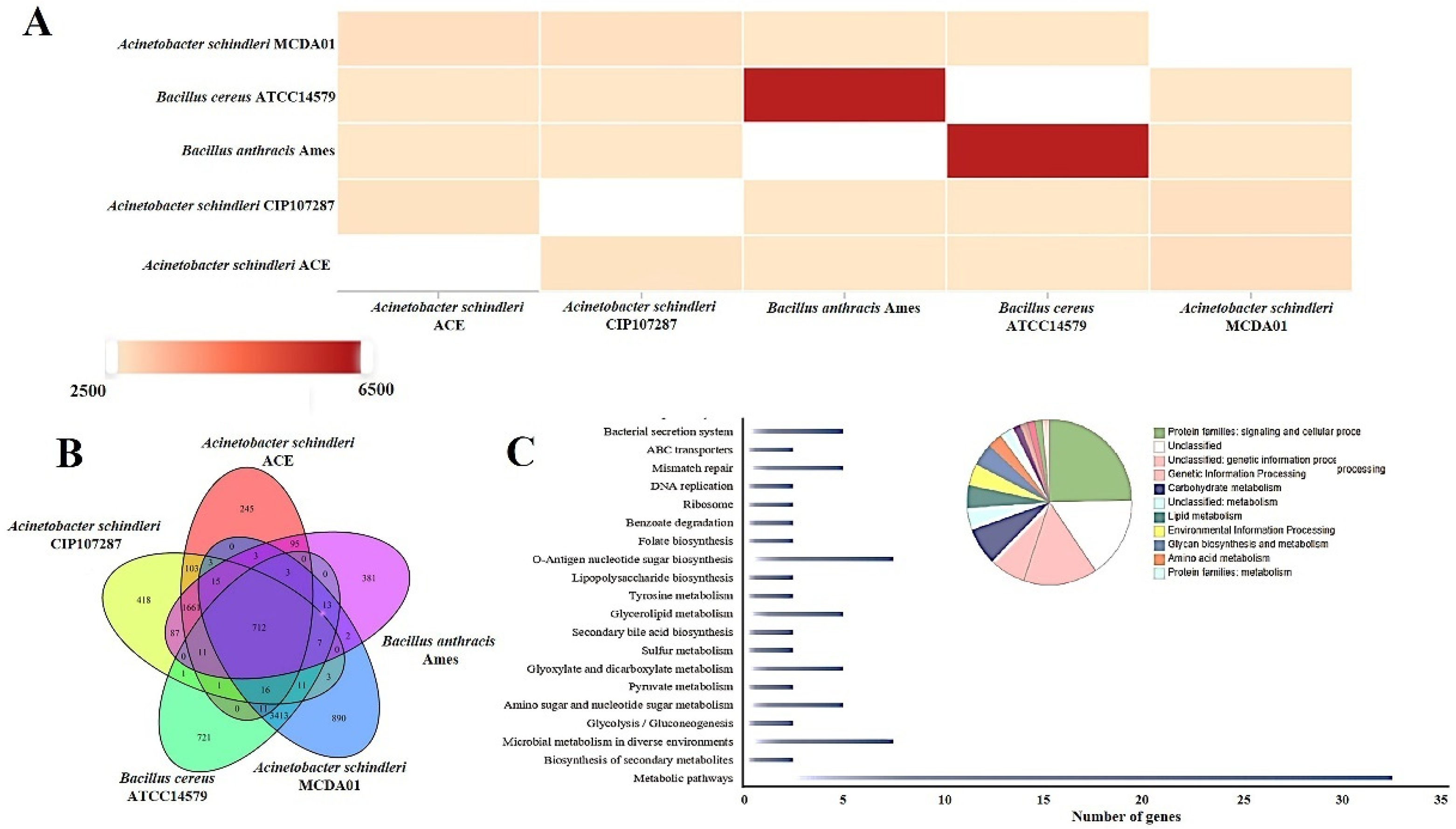
2.2. Comparative Genome Analysis of Acinetobacter schindleri MCDA01
2.3. Genome Mining and Bioinformatics Analysis of AsCDA
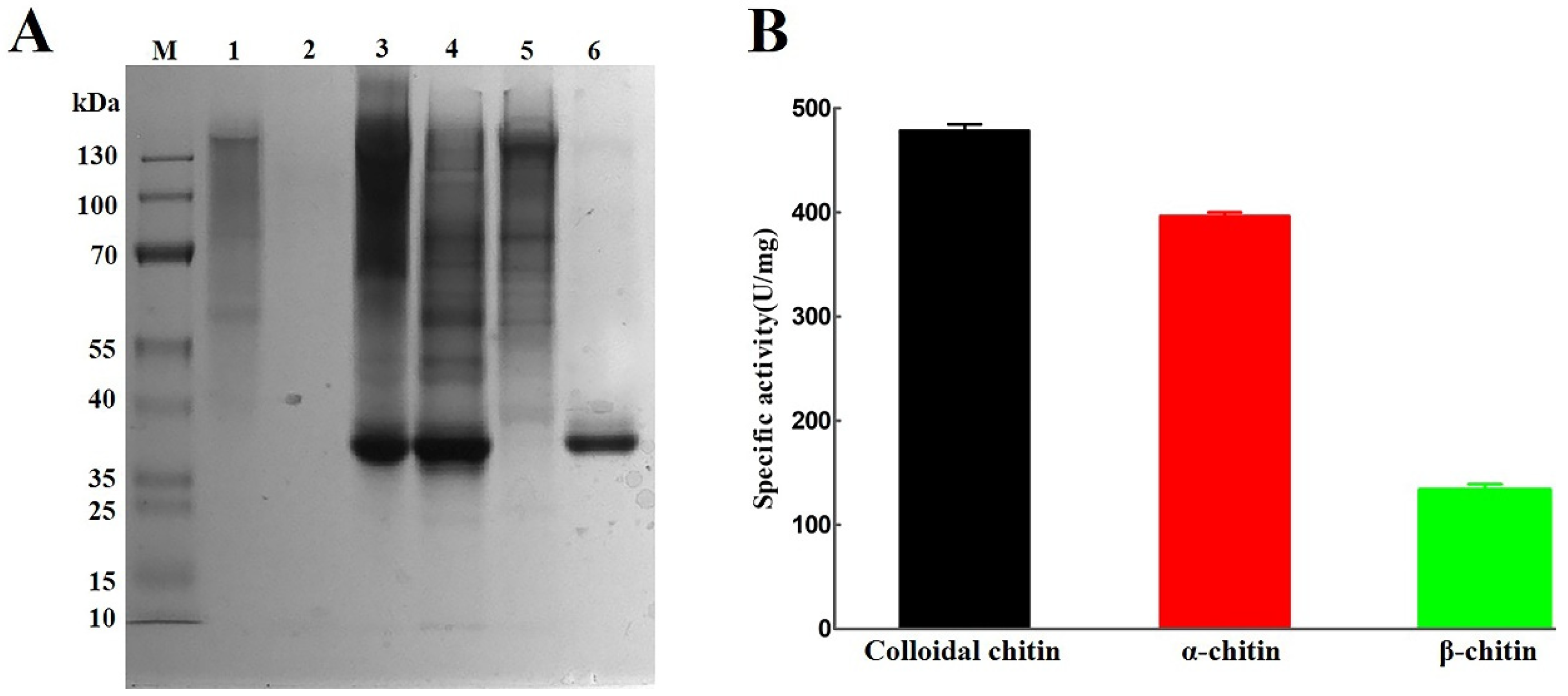
2.4. Heterologous Expression of AsCDA in Escherichia coli
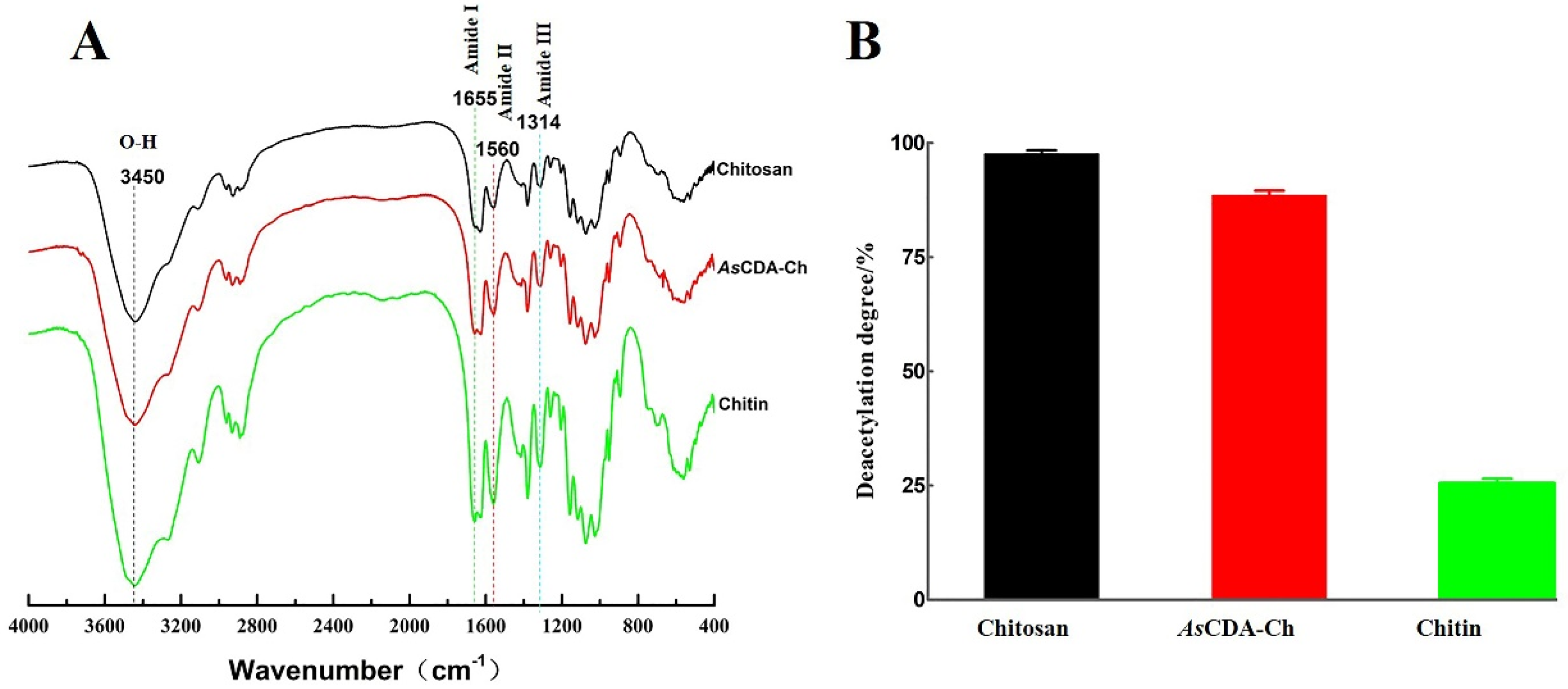
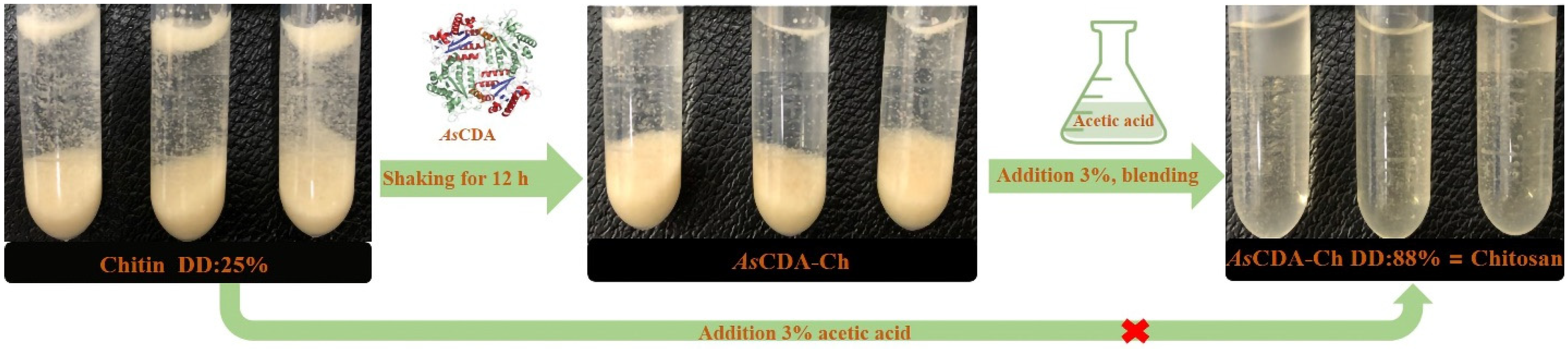
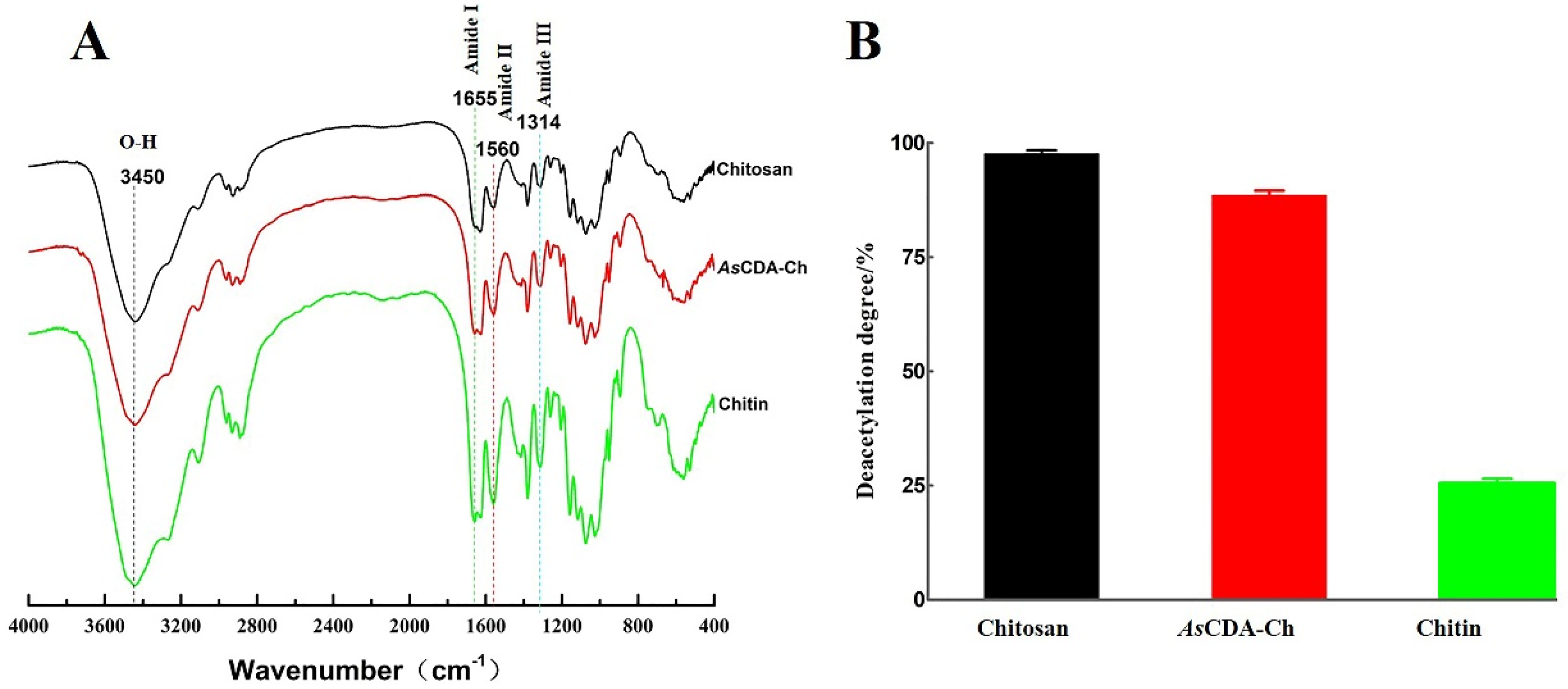
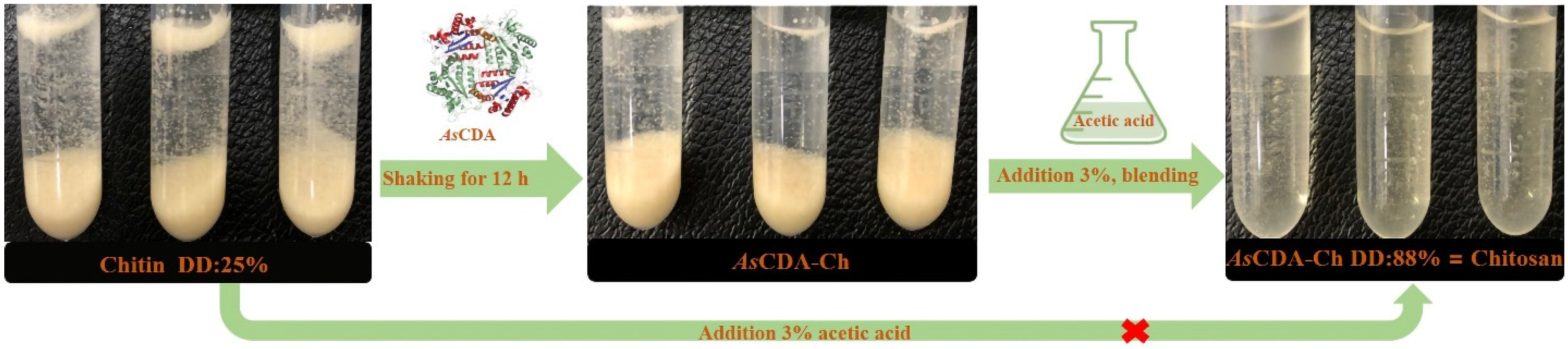
2.5. Deacetylation of Crystalline Chitin Catalyzed by Chitin Deacetylase AsCDA
3. Materials and Methods
3.1. Materials
3.2. Isolation, Sequencing, and De Novo Assembly of the Acinetobacter schindleri MCDA01 Genome
3.3. Gene Prediction and Annotation
3.4. Comparative Genome Analysis and Gene Mining of Chitin Deacetylase
3.5. Heterologous Expression and Purification of AsCDA in Escherichia coli BL21
3.6. Enzyme Assay of AsCDA
3.7. Scanning Electron Microscope Observation of Deacetylation Effect
3.8. Deacetylation Was Measured by Infrared Spectroscopy
3.9. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yan, N.; Chen, X. Sustainability: Don’t waste seafood waste. Nature 2015, 524, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Arnold, N.D.; Bruck, W.M.; Garbe, D.; Bruck, T.B. Enzymatic modification of native chitin and conversion to specialty chemical products. Mar. Drugs 2020, 18, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karrer, P.; Hofmann, A. Polysaccharide xxxix. Über den enzymatischen abbau von chitin und chitosan I. Helv. Chim. Acta 1929, 12, 616–637. [Google Scholar] [CrossRef]
- Aspras, I.; Jaworska, M.M.; Gorak, A. Kinetics of chitin deacetylase activation by the ionic liquid [Bmim][Br]. J. Biotechnol. 2017, 251, 94–98. [Google Scholar] [CrossRef]
- Deringer, V.L.; Englert, U.; Dronskowski, R. Nature, strength, and cooperativity of the hydrogen-bonding network in alpha-chitin. Biomacromolecules 2016, 17, 996–1003. [Google Scholar] [CrossRef] [Green Version]
- Tsigos, I.; Martinou, A.; Kafetzopoulos, D.; Bouriotis, V. Chitin deacetylases: New, versatile tools in biotechnology. Trends Biotechnol. 2000, 18, 305–312. [Google Scholar] [CrossRef]
- Kaczmarek, M.B.; Struszczyk-Swita, K.; Xiao, M.; Szczesna-Antczak, M.; Antczak, T.; Gierszewska, M.; Steinbuchel, A.; Daroch, M. Polycistronic expression system for Pichia pastoris composed of chitino- and chitosanolytic enzymes. Front Bioeng. Biotechnol. 2021, 9, 710922. [Google Scholar] [CrossRef]
- Casadidio, C.; Peregrina, D.V.; Gigliobianco, M.R.; Deng, S.; Censi, R.; Di Martino, P. Chitin and chitosans: Characteristics, eco-friendly processes, and applications in cosmetic science. Mar. Drugs 2019, 17, 369. [Google Scholar] [CrossRef] [Green Version]
- Muxika, A.; Etxabide, A.; Uranga, J.; Guerrero, P.; de la Caba, K. Chitosan as a bioactive polymer: Processing, properties and applications. Int. J. Biol. Macromol. 2017, 105, 1358–1368. [Google Scholar] [CrossRef]
- Park, B.K.; Kim, M.M. Applications of chitin and its derivatives in biological medicine. Int. J. Mol. Sci. 2010, 11, 5152–5164. [Google Scholar] [CrossRef] [Green Version]
- Ye, W.; Ma, H.; Liu, L.; Yu, J.; Lai, J.; Fang, Y.; Fan, Y. Biocatalyzed route for the preparation of surface-deacetylated chitin nanofibers. Green Chem. 2019, 21, 3143–3151. [Google Scholar] [CrossRef]
- Yuan, X.; Zheng, J.; Jiao, S.; Cheng, G.; Feng, C.; Du, Y.; Liu, H. A review on the preparation of chitosan oligosaccharides and application to human health, animal husbandry and agricultural production. Carbohydr. Polym. 2019, 220, 60–70. [Google Scholar] [CrossRef]
- Blair, D.E.; Schuttelkopf, A.W.; Macrae, J.I.; van Aalten, D.M. Structure and metal-dependent mechanism of peptidoglycan deacetylase, a streptococcal virulence factor. Proc. Natl. Acad. Sci. USA 2005, 102, 15429–15434. [Google Scholar] [CrossRef] [Green Version]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.J.; Xu, K.K.; Yan, Y.; Li, C.; Jin, D.C. Role of chitin deacetylase 1 in the molting and metamorphosis of the cigarette beetle Lasioderma serricorne. Int. J. Mol. Sci. 2020, 21, 2449. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, J.; Zhu, K.Y. Chitin in arthropods: Biosynthesis, modification, and metabolism. Adv. Exp. Med. Biol. 2019, 1142, 169–207. [Google Scholar]
- Araki, Y.; Ito, E. A pathway of chitosan formation in Mucor rouxii. Enzymatic deacetylation of chitin. Eur. J. Biochem. 1975, 55, 71–78. [Google Scholar] [CrossRef]
- Yang, G.; Hou, X.; Lu, J.; Wang, M.; Wang, Y.; Huang, Y.; Liu, Q.; Liu, S.; Fang, Y. Enzymatic modification of native chitin and chitin oligosaccharides by an alkaline chitin deacetylase from Microbacterium esteraromaticum MCDA02. Int. J. Biol. Macromol. 2022, 203, 671–678. [Google Scholar] [CrossRef]
- Shrestha, B.; Blondeau, K.; Stevens, W.F.; Hegarat, F.L. Expression of chitin deacetylase from Colletotrichum lindemuthianum in Pichia pastoris: Purification and characterization. Protein Expr. Purif. 2004, 38, 196–204. [Google Scholar] [CrossRef]
- Zhu, X.Y.; Zhao, Y.; Zhang, H.D.; Wang, W.X.; Cong, H.H.; Yin, H. Characterization of the specific mode of action of a chitin deacetylase and separation of the partially acetylated chitosan oligosaccharides. Mar. Drugs 2019, 17, 74. [Google Scholar] [CrossRef] [Green Version]
- Win, N.N.; Stevens, W.F. Shrimp chitin as substrate for fungal chitin deacetylase. Appl. Microbiol. Biotechnol. 2001, 57, 334–341. [Google Scholar] [PubMed]
- Yamada, M.; Kurano, M.; Inatomi, S.; Taguchi, G.; Okazaki, M.; Shimosaka, M. Isolation and characterization of a gene coding for chitin deacetylase specifically expressed during fruiting body development in the basidiomycete Flammulina velutipes and its expression in the yeast Pichia pastoris. FEMS Microbiol. Lett. 2008, 289, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Gay, L.M.; Tuveng, T.R.; Agger, J.W.; Westereng, B.; Mathiesen, G.; Horn, S.J.; Vaaje-Kolstad, G.; van Aalten, D.; Eijsink, V. Structure and function of a broad-specificity chitin deacetylase from Aspergillus nidulans FGSC A4. Sci. Rep. 2017, 7, 1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Q.; Gao, X.; Tu, L.; Han, Q.; Zhang, X.; Guo, Y.; Yan, W.; Shen, Y.; Wang, M. Enhanced chitin deacetylase production ability of Rhodococcus equi CGMCC14861 by co-culture fermentation with Staphylococcus sp. MC7. Front. Microbiol. 2020, 11, 592477. [Google Scholar] [CrossRef]
- Wang, Y.; Song, J.Z.; Yang, Q.; Liu, Z.H.; Huang, X.M.; Chen, Y. Cloning of a heat-stable chitin deacetylase gene from Aspergillus nidulans and its functional expression in Escherichia coli. Appl. Biochem. Biotechnol. 2010, 162, 843–854. [Google Scholar] [CrossRef]
- Kang, L.; Chen, X.; Zhai, C.; Ma, L. Synthesis and high expression of chitin deacetylase from Colletotrichum lindemuthianum in Pichia pastoris GS115. J. Microbiol. Biotechnol. 2012, 22, 1202–1207. [Google Scholar] [CrossRef] [Green Version]
- Blair, D.E.; Hekmat, O.; Schuttelkopf, A.W.; Shrestha, B.; Tokuyasu, K.; Withers, S.G.; van Aalten, D.M. Structure and mechanism of chitin deacetylase from the fungal pathogen Colletotrichum lindemuthianum. BMC Biochem. 2006, 45, 9416–9426. [Google Scholar] [CrossRef] [Green Version]
- Dsouza, M.; Taylor, M.W.; Turner, S.J.; Aislabie, J. Genomic and phenotypic insights into the ecology of Arthrobacter from antarctic soils. BMC Genom. 2015, 16, 36. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Dubey, S.K. Cellulose degradation potential of Paenibacillus lautus strain BHU3 and its whole genome sequence. Bioresour. Technol. 2018, 262, 124–131. [Google Scholar] [CrossRef]
- Ma, Q.; Gao, X.; Bi, X.; Tu, L.; Xia, M.; Shen, Y.; Wang, M. Isolation, characterisation, and genome sequencing of Rhodococcus equi: A novel strain producing chitin deacetylase. Sci. Rep. 2020, 10, 4329. [Google Scholar] [CrossRef] [Green Version]
- Ramazzina, I.; Cendron, L.; Folli, C.; Berni, R.; Monteverdi, D.; Zanotti, G.; Percudani, R. Logical identification of an allantoinase analog (puue) recruited from polysaccharide deacetylases. J. Biol. Chem. 2008, 283, 23295–23304. [Google Scholar] [CrossRef] [Green Version]
- Grifoll-Romero, L.; Pascual, S.; Aragunde, H.; Biarnes, X.; Planas, A. Chitin deacetylases: Structures, specificities, and biotech applications. Polymers 2018, 10, 352. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Park, R.D.; Muzzarelli, R.A. Chitin deacetylases: Properties and applications. Mar. Drugs 2010, 8, 24–46. [Google Scholar] [CrossRef]
- Alfonso, C.; Nuero, O.M.; Santamaria, F.; Reyes, F. Purification of a heat-stable chitin deacetylase from Aspergillus nidulans and its role in cell wall degradation. Curr. Microbiol. 1995, 30, 49–54. [Google Scholar] [CrossRef]
- Chai, J.; Hang, J.; Zhang, C.; Yang, J.; Wang, S.; Liu, S.; Fang, Y. Purification and characterization of chitin deacetylase active on insoluble chitin from Nitratireductor aquimarinus MCDA3-3. Int. J. Biol. Macromol. 2020, 152, 922–929. [Google Scholar] [CrossRef]
- Pareek, N.; Vivekanand, V.; Agarwal, P.; Saroj, S.; Singh, R.P. Bioconversion to chitosan: A two stage process employing chitin deacetylase from Penicillium oxalicum SAEM-51. Carbohydr. Polym. 2013, 96, 417–425. [Google Scholar] [CrossRef]
- Li, J.; Huang, W.C.; Gao, L.; Sun, J.; Liu, Z.; Mao, X. Efficient enzymatic hydrolysis of ionic liquid pretreated chitin and its dissolution mechanism. Carbohydr. Polym. 2019, 211, 329–335. [Google Scholar] [CrossRef]
- Ma, Q.; Gao, X.; Bi, X.; Han, Q.; Tu, L.; Yang, Y.; Shen, Y.; Wang, M. Dissolution and deacetylation of chitin in ionic liquid tetrabutylammonium hydroxide and its cascade reaction in enzyme treatment for chitin recycling. Carbohydr. Polym. 2020, 230, 115605. [Google Scholar] [CrossRef]
- Velde, K.K.V.D.; Kiekens, P. Structure analysis and degree of substitution of chitin, chitosan and dibutyrylchitin by FT-IR spectroscopy and solid state 13C NMR. Carbohyd. Polym. 2004, 58, 409–416. [Google Scholar] [CrossRef]
- Peh, K.K.; Khan, T.A.; Ch’Ng, H.S. Mechanical, bioadhesive strength and biological evaluations of chitosan films for wound dressing. J. Pharm. Pharmacent. 2000, 3, 303–311. [Google Scholar]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. Soap: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Zhu, H.; Ruan, J.; Qian, W.; Fang, X.; Shi, Z.; Li, Y.; Li, S.; Shan, G.; Kristiansen, K.; et al. De novo assembly of human genomes with massively parallel short read sequencing. Genome Res. 2010, 20, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. Erratum: Soapdenovo2: An empirically improved memory-efficient short-read de novo assembler. Gigascience 2015, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. Genemarks: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. Dbcan2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [Green Version]
- Jungo, F.; Bougueleret, L.; Xenarios, I.; Poux, S. The uniprotkb/swiss-prot tox-prot program: A central hub of integrated venom protein data. Toxicon 2012, 60, 551–557. [Google Scholar] [CrossRef] [Green Version]
- Magrane, M. Uniprot knowledgebase: A hub of integrated protein data. Database 2011, 2011, r9. [Google Scholar] [CrossRef] [Green Version]
- Koonin, E.V.; Fedorova, N.D.; Jackson, J.D.; Jacobs, A.R.; Krylov, D.M.; Makarova, K.S.; Mazumder, R.; Mekhedov, S.L.; Nikolskaya, A.N.; Rao, B.S.; et al. A comprehensive evolutionary classification of proteins encoded in complete eukaryotic genomes. Genome Biol. 2004, 5, R7. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Cui, X.; Liu, S.; Lu, J.; Hou, X.; Meng, W.; Wu, B.; Su, Y.; Zhang, H.; Zheng, W.; et al. Effects of dietary Lactobacillus helveticus on the growth rate, disease resistance and intestinal health of pond loach (Misgurnus anguillicaudatus). Aquaculture 2021, 544, 737038. [Google Scholar] [CrossRef]
- Vinuesa, P.; Contreras-Moreira, B. Robust identification of orthologues and paralogues for microbial pan-genomics using get_homologues: A case study of pinca/c plasmids. Methods Mol. Biol. 2015, 1231, 203–232. [Google Scholar]
- Wang, Y.; Coleman-Derr, D.; Chen, G.; Gu, Y.Q. Orthovenn: A web server for genome wide comparison and annotation of orthologous clusters across multiple species. Nucleic Acids Res. 2015, 43, W78–W84. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tang, Y.; Fang, X.; Qiao, T.; Han, S.; Zhu, T. Whole-genome sequence of Arthrinium phaeospermum, a globally distributed pathogenic fungus. Genomics 2020, 112, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Sigala, J.C.; Suarez, B.P.; Lara, A.R.; Borgne, S.L.; Bustos, P.; Santamaria, R.I.; Gonzalez, V.; Martinez, A. Genomic and physiological characterization of a laboratory-isolated Acinetobacter schindleri ACE strain that quickly and efficiently catabolizes acetate. Microbiology 2017, 163, 1052–1064. [Google Scholar] [CrossRef] [PubMed]
- Sigala, J.C.; Quiroz, L.; Arteaga, E.; Olivares, R.; Lara, A.R.; Martinez, A. Physiological and transcriptional comparison of acetate catabolism between Acinetobacter schindleri ACE and Escherichia coli JM101. FEMS Microbiol. Lett. 2019, 366, fnz151. [Google Scholar] [CrossRef]
- Read, T.D.; Peterson, S.N.; Tourasse, N.; Baillie, L.W.; Paulsen, I.T.; Nelson, K.E.; Tettelin, H.; Fouts, D.E.; Eisen, J.A.; Gill, S.R.; et al. The genome sequence of Bacillus anthracis Ames and comparison to closely related bacteria. Nature 2003, 423, 81–86. [Google Scholar] [CrossRef]
- Andreou, A.; Giastas, P.; Christoforides, E.; Eliopoulos, E.E. Structural and evolutionary insights within the polysaccharide deacetylase gene family of Bacillus anthracis and Bacillus cereus. Genes 2018, 9, 386. [Google Scholar] [CrossRef] [Green Version]
- Khatri, I.; Tomar, R.; Ganesan, K.; Prasad, G.S.; Subramanian, S. Complete genome sequence and comparative genomics of the probiotic yeast Saccharomyces boulardii. Sci. Rep. 2017, 7, 371. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Xiu, Y.; Chen, Y.; Bai, L.; Sha, Z. Identification and expression of complement component c8α, c8β and c8γ gene in half-smooth tongue sole (Cynoglossus semilaevis) and c8α recombinant protein antibacterial activity analysis. Fish Shellfish Immunol. 2017, 72, 658–669. [Google Scholar] [CrossRef]
- Bergmeyer, H.-U. Methods of Enzymatic Analysis; Wiley: Hoboken, NJ, USA, 1974; Volume 4. [Google Scholar]
- Hou, X.; Wang, Z.; Sun, J.; Li, M.; Wang, S.; Chen, K.; Gao, Z. A microwave-assisted aqueous ionic liquid pretreatment to enhance enzymatic hydrolysis of eucalyptus and its mechanism. Bioresour. Technol. 2019, 272, 99–104. [Google Scholar] [CrossRef]
- Ngo, T.; Ngo, D.N. Effects of low-frequency ultrasound on heterogenous deacetylation of chitin. Int. J. Biol. Macromol. 2017, 104, 1604–1610. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| General Features | A. schindleri MCDA01 |
|---|---|
| Genome size (bp) | 3,332,387 |
| GC content (%) | 43 |
| Gene number | 3107 |
| Gene length | 2,809,674 |
| % of Genome (genes) | 84.31 |
| Gene average length | 904.3 |
| rRNAs | 21 |
| tRNAs | 88 |
| sRNAs | 9 |
| CAZy assignment | 35 |
| COG assignment | 2267 |
| GO assignment | 1902 |
| KEGG alignment | 1881 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Wang, Y.; Fang, Y.; An, J.; Hou, X.; Lu, J.; Zhu, R.; Liu, S. A Novel Potent Crystalline Chitin Decomposer: Chitin Deacetylase from Acinetobacter schindleri MCDA01. Molecules 2022, 27, 5345. https://doi.org/10.3390/molecules27165345
Yang G, Wang Y, Fang Y, An J, Hou X, Lu J, Zhu R, Liu S. A Novel Potent Crystalline Chitin Decomposer: Chitin Deacetylase from Acinetobacter schindleri MCDA01. Molecules. 2022; 27(16):5345. https://doi.org/10.3390/molecules27165345
Chicago/Turabian StyleYang, Guang, Yuhan Wang, Yaowei Fang, Jia An, Xiaoyue Hou, Jing Lu, Rongjun Zhu, and Shu Liu. 2022. "A Novel Potent Crystalline Chitin Decomposer: Chitin Deacetylase from Acinetobacter schindleri MCDA01" Molecules 27, no. 16: 5345. https://doi.org/10.3390/molecules27165345
APA StyleYang, G., Wang, Y., Fang, Y., An, J., Hou, X., Lu, J., Zhu, R., & Liu, S. (2022). A Novel Potent Crystalline Chitin Decomposer: Chitin Deacetylase from Acinetobacter schindleri MCDA01. Molecules, 27(16), 5345. https://doi.org/10.3390/molecules27165345