
Cytotoxic Activity of Polyphenol Extracts from Three Oregano Species: Hedeoma patens, Lippia graveolens and Lippia palmeri, and Antiproliferative Potential of Lippia graveolens against Two Types of Breast Cancer Cell Lines (MDA-MB-231 and MCF-7)

 , ,
, ,  ,
,  ,
,  , ,
, , 
Abstract
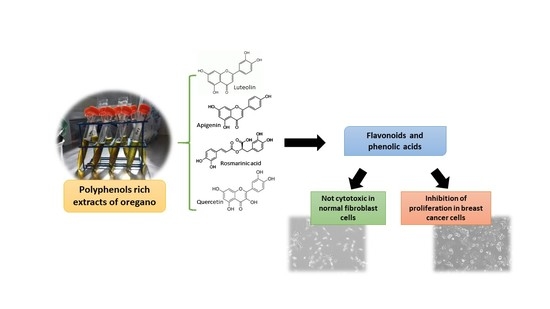
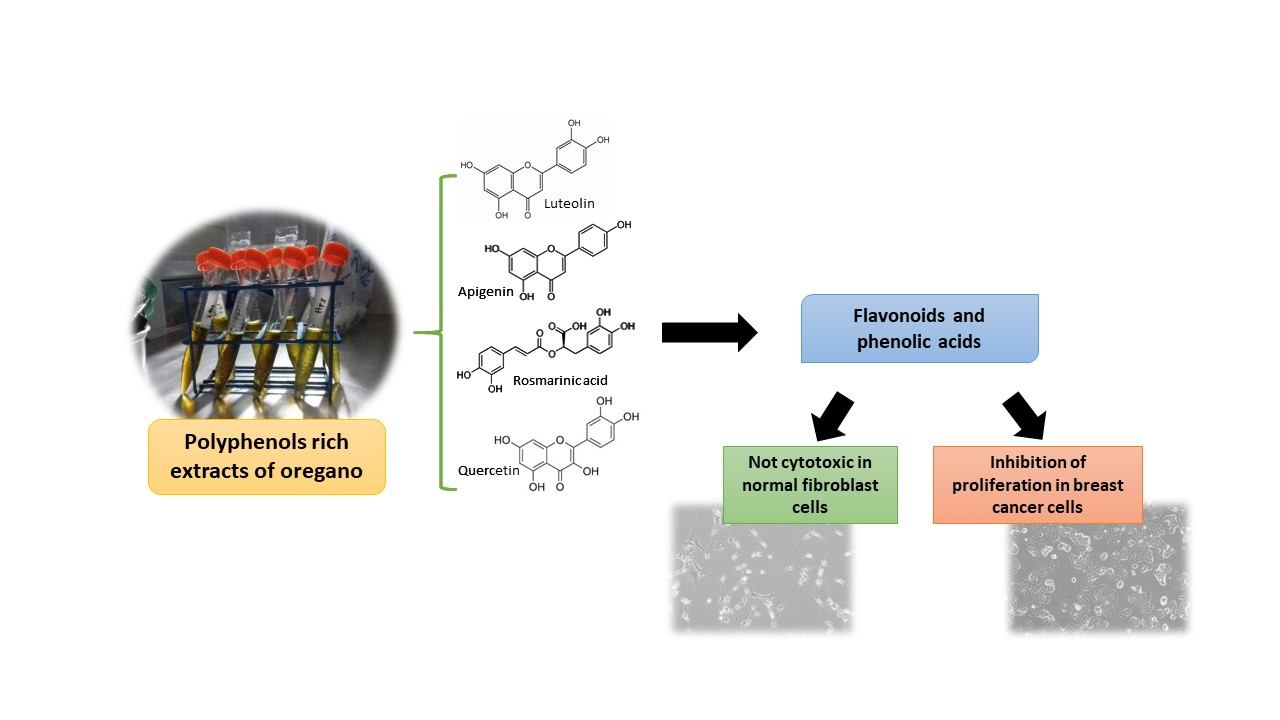
:
1. Introduction
2. Results and Discussion
2.1. Antioxidant Activity of PRE of Three Oregano Species
2.2. Total Phenolic Content of PRE of Three Oregano Species
2.3. Phenolic Compounds Profile Identified in PRE of Three Oregano Species
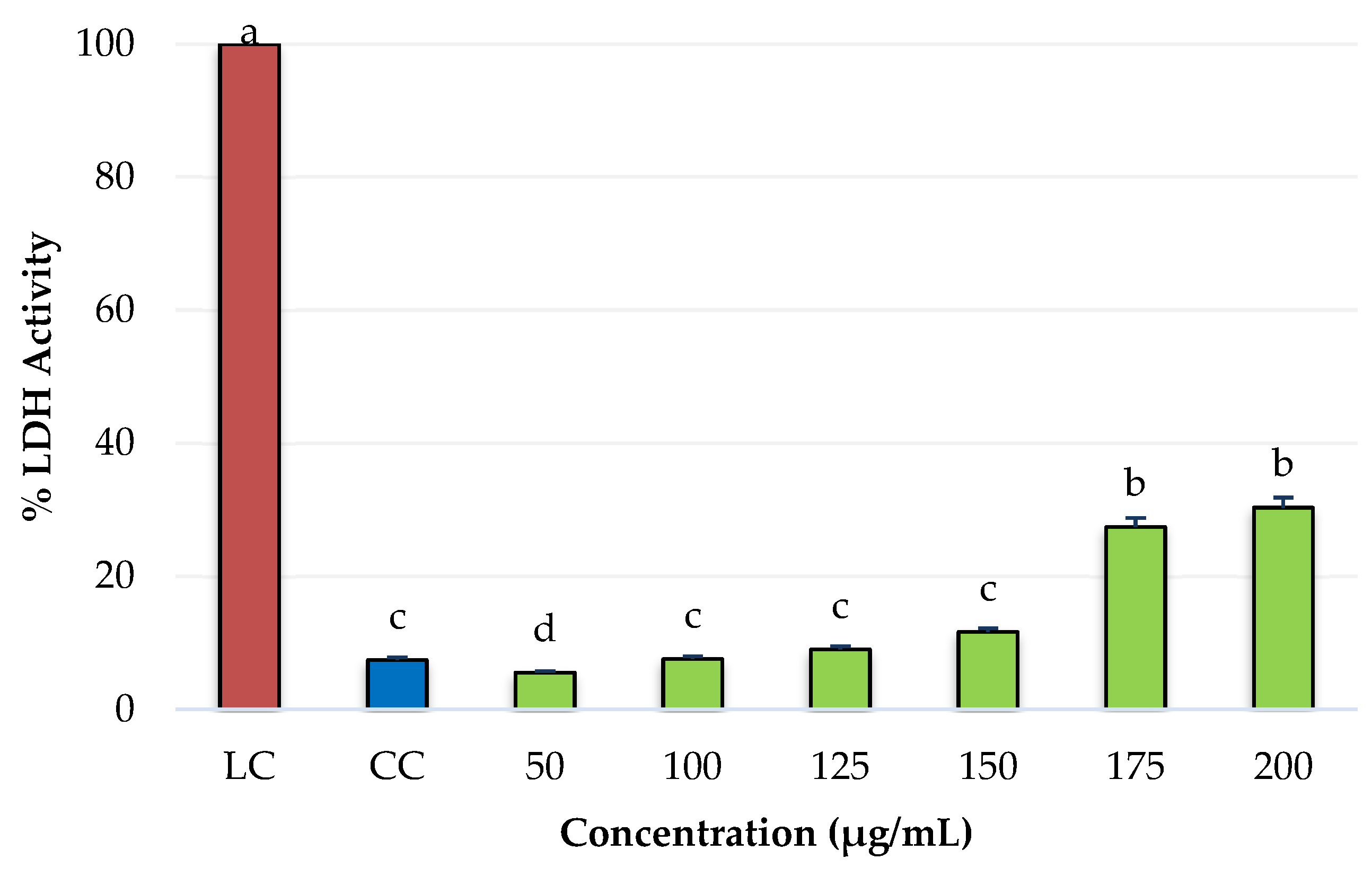
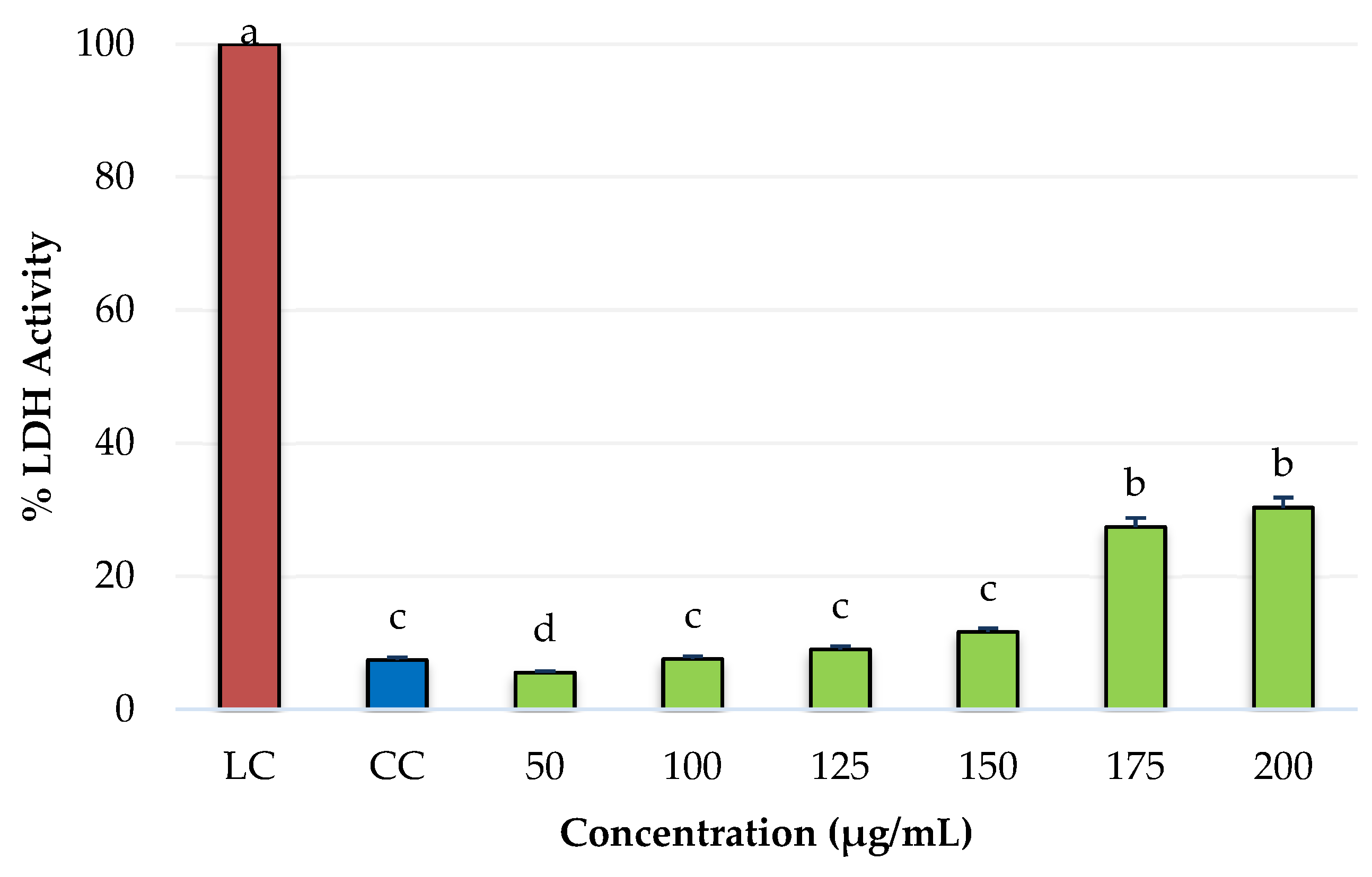
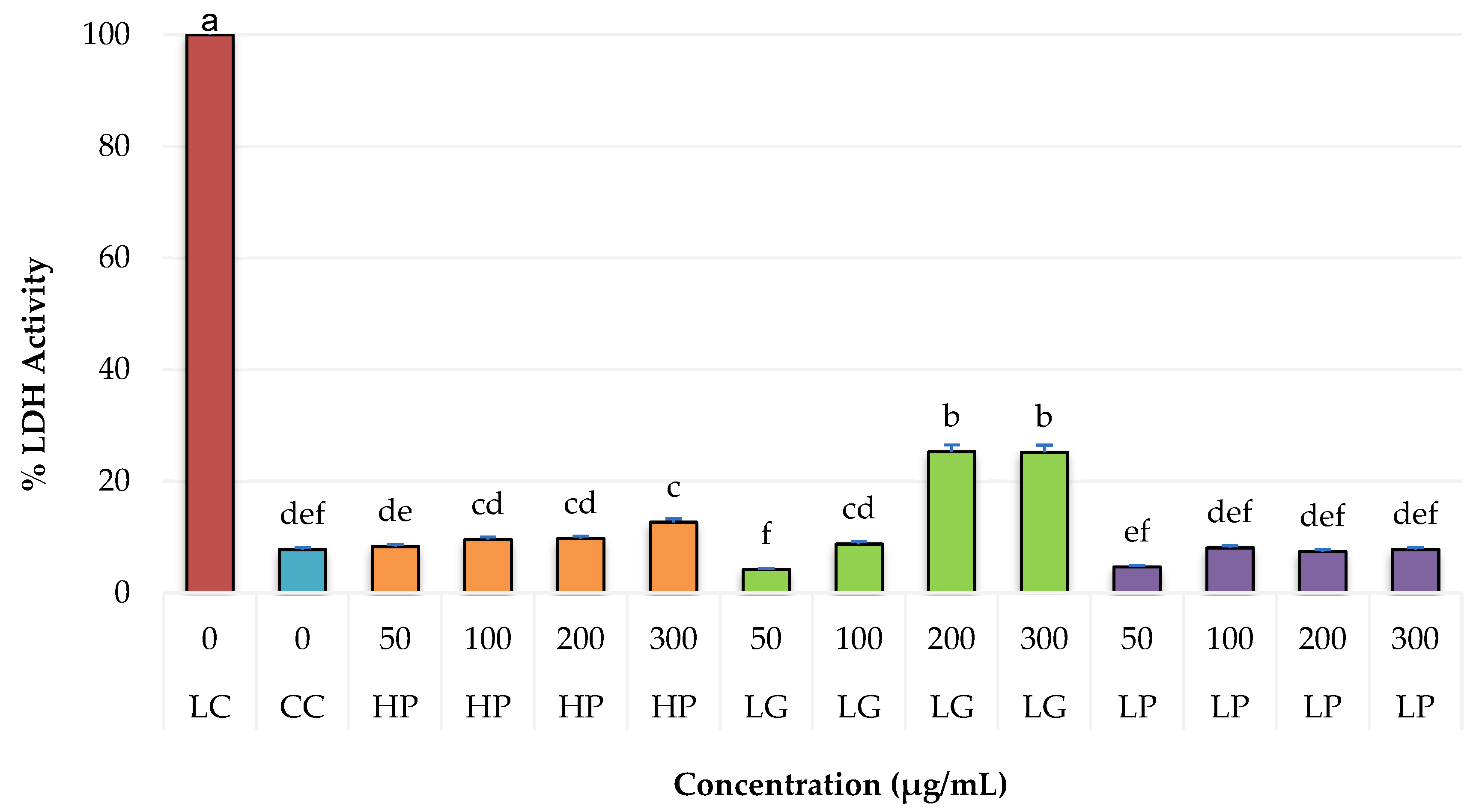
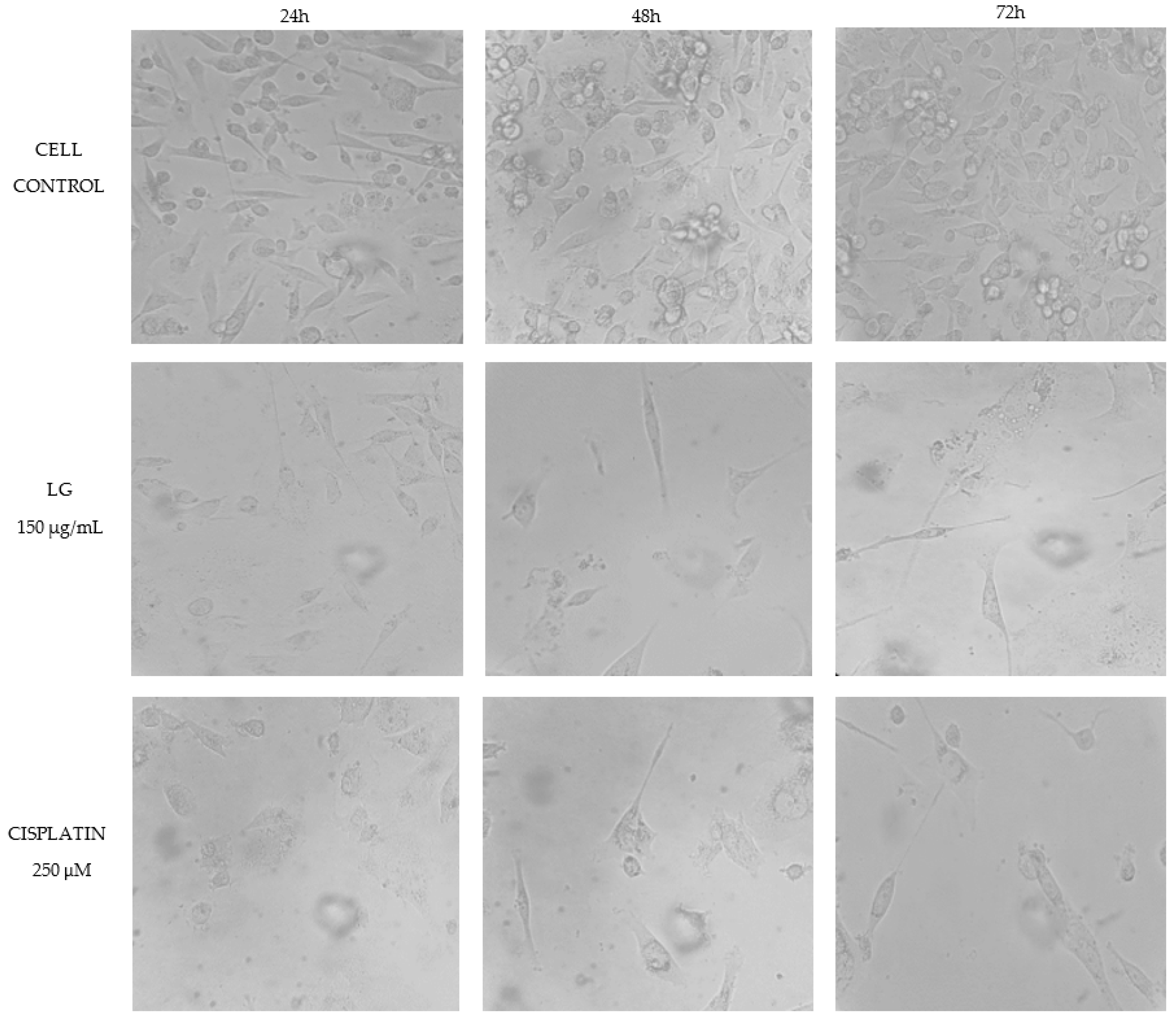
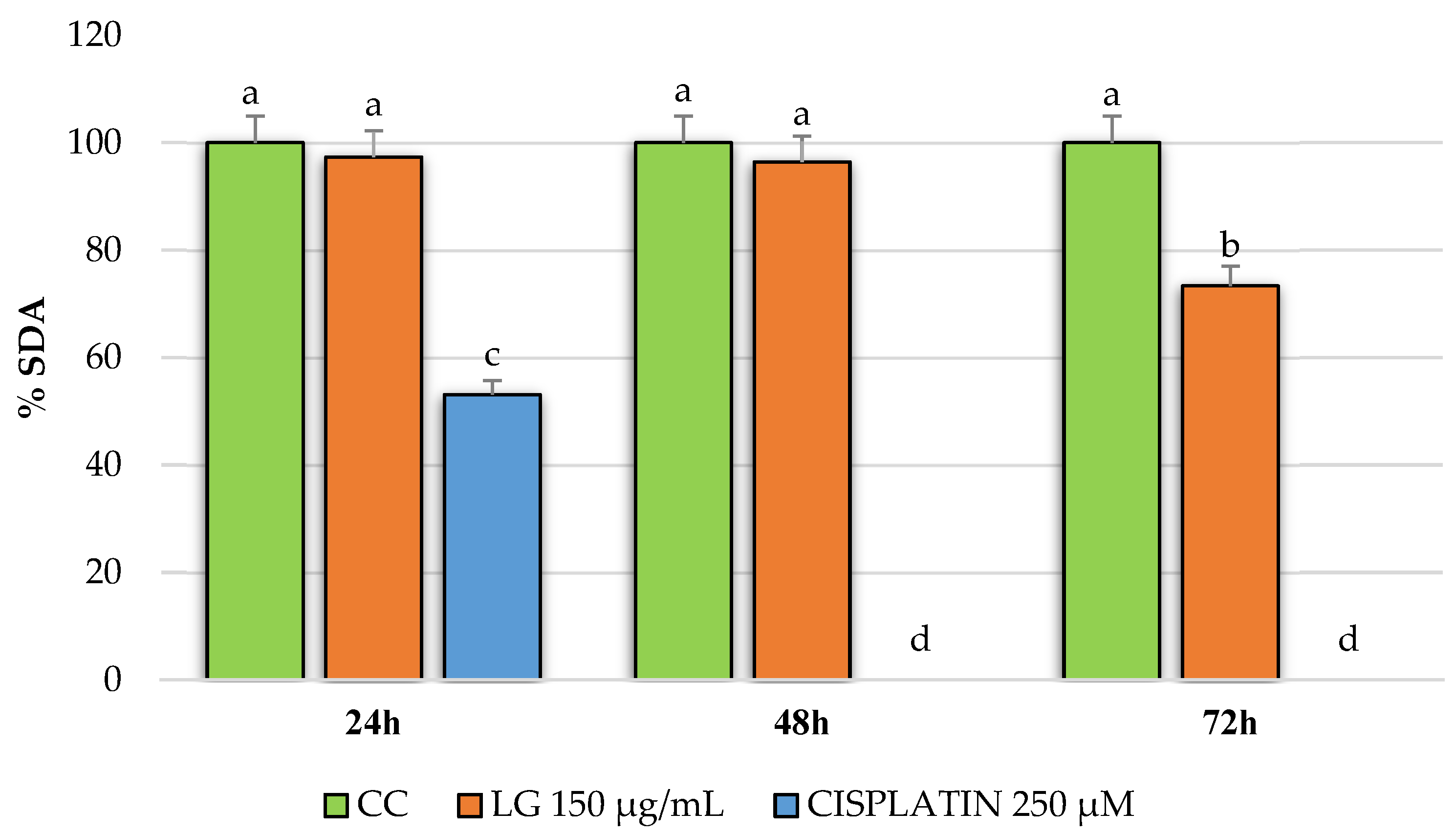
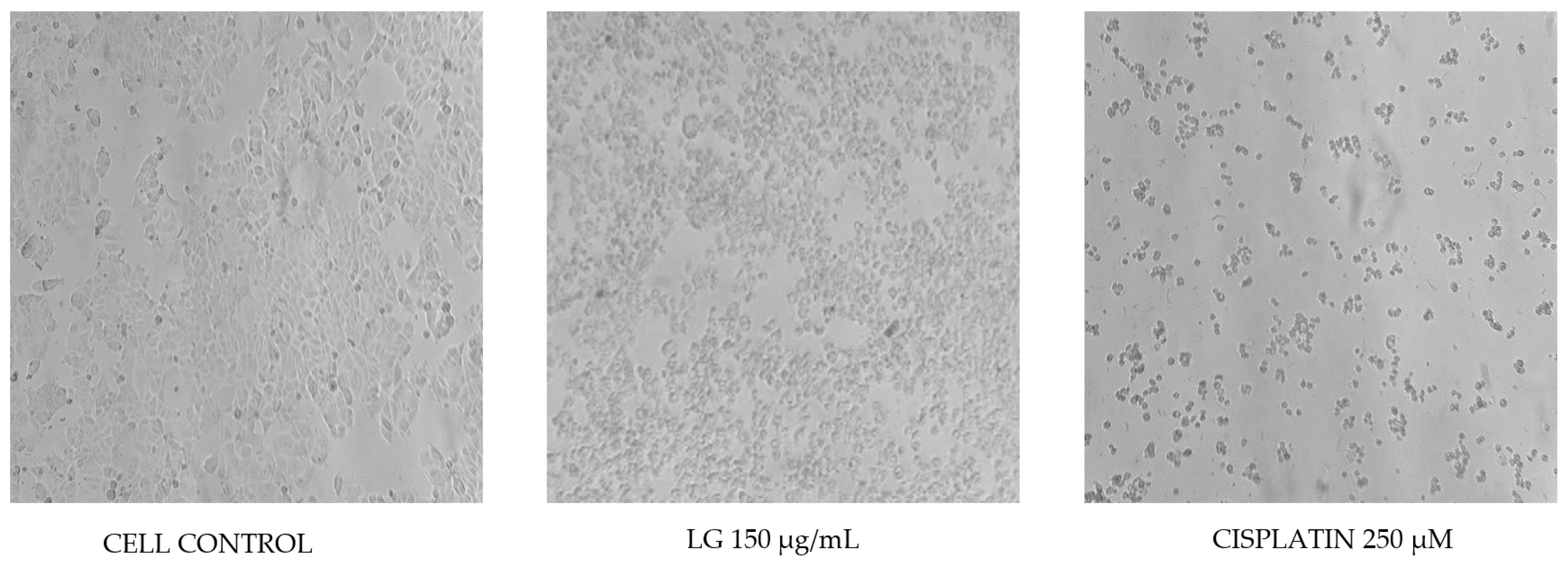
2.4. Cytotoxicity of PRE of Three Oregano Species on Non-Cancer Fibroblast Cells and Antiproliferative Activity of the Extracts on Breast Cancer Cells
3. Materials and Methods
3.1. Plant Material
3.2. Preparation of the Oregano Polyphenols Rich-Extracts (PRE)
3.3. Antioxidant Activity
3.3.1. ORAC
3.3.2. DPPH
3.4. Determination of Total Phenolic Content
3.5. Identification of Phenolic Compounds by LC-ESI-QTOF-MS/MS
3.6. Cell Culture
3.7. Cytotoxicity Assay
3.8. Antiproliferative Activity
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Fleisher, A.; Sneer, N. Oregano spices and Origanum chemotypes. J. Sci. Food Agric. 1982, 33, 441–446. [Google Scholar] [CrossRef]
- Kintzios, S.E. 21-Oregano. In Handbook of Herbs and Spices, 2nd ed.; Peter, K.V., Ed.; Woodhead Publishing: Cambridge, UK, 2012; pp. 417–436. [Google Scholar]
- Rivero-Cruz, I.; Duarte, G.; Navarrete, A.; Bye, R.; Linares, E.; Mata, R. Chemical composition and antimicrobial and spasmolytic properties of Poliomintha longiflora and Lippia graveolens essential oils. J. Food Sci. 2011, 76, C309–C317. [Google Scholar] [CrossRef]
- Arcila, C.; Loarca, G.; Lecona, S.; González, E. El orégano: Propiedades, composición y actividad biológica de sus componentes. Arch. Latinoam. Nutr. 2004, 54, 100–111. [Google Scholar]
- Baser, K. The Turkish Origanum species. Med. Aromat. Plants 2002, 26, 109. [Google Scholar]
- Laferrière, J.E.; Weber, C.; Kohlhepp, E.A. Use and nutritional composition of some traditional mountain pima plant foods. J. Ethnobiol. 1991, 11, 93–114. [Google Scholar]
- Gutiérrez-Grijalva, E.; Picos-Salas, M.; Leyva-López, N.; Criollo-Mendoza, M.; Vazquez-Olivo, G.; Heredia, J. Flavonoids and phenolic acids from oregano: Occurrence, biological activity and health benefits. Plants 2018, 7, 2. [Google Scholar] [CrossRef]
- Leyva-López, N.; Nair, V.; Bang, W.Y.; Cisneros-Zevallos, L.; Heredia, J.B. Protective role of terpenes and polyphenols from three species of Oregano (Lippia graveolens, Lippia palmeri and Hedeoma patens) on the suppression of lipopolysaccharide-induced inflammation in RAW 264.7 macrophage cells. J. Ethnopharmacol. 2016, 187, 302–312. [Google Scholar] [CrossRef]
- Kogiannou, D.A.; Kalogeropoulos, N.; Kefalas, P.; Polissiou, M.G.; Kaliora, A.C. Herbal infusions; their phenolic profile, antioxidant and anti-inflammatory effects in HT29 and PC3 cells. Food Chem. Toxicol. 2013, 61, 152–159. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, S.Y. Antioxidant activity and phenolic compounds in selected herbs. J. Agric. Food Chem. 2001, 49, 5165–5170. [Google Scholar] [CrossRef]
- Yoshino, K.; Higashi, N.; Koga, K. Antioxidant and anti-inflammatory activities of oregano extract. J. Health Sci. 2006, 52, 169–173. [Google Scholar] [CrossRef]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Chaouki, W.; Leger, D.Y.; Eljastimi, J.; Beneytout, J.L.; Hmamouchi, M. Antiproliferative effect of extracts from Aristolochia baetica and Origanum compactum on human breast cancer cell line MCF-7. Pharm. Biol. 2010, 48, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Berdowska, I.; Zielinski, B.; Fecka, I.; Kulbacka, J.; Saczko, J.; Gamian, A. Cytotoxic impact of phenolics from Lamiaceae species on human breast cancer cells. Food Chem. 2013, 141, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- El Babili, F.; Bouajila, J.; Souchard, J.P.; Bertrand, C.; Bellvert, F.; Fouraste, I.; Moulis, C.; Valentin, A. Oregano: Chemical analysis and evaluation of its antimalarial, antioxidant, and cytotoxic activities. J. Food Sci. 2011, 76, C512–C518. [Google Scholar] [CrossRef]
- Kaliora, A.C.; Kogiannou, D.A.; Kefalas, P.; Papassideri, I.S.; Kalogeropoulos, N. Phenolic profiles and antioxidant and anticarcinogenic activities of Greek herbal infusions; balancing delight and chemoprevention? Food Chem. 2014, 142, 233–241. [Google Scholar] [CrossRef]
- Marrelli, M.; Cristaldi, B.; Menichini, F.; Conforti, F. Inhibitory effects of wild dietary plants on lipid peroxidation and on the proliferation of human cancer cells. Food Chem. Toxicol. 2015, 86, 16–24. [Google Scholar] [CrossRef]
- Al-Kalaldeh, J.Z.; Abu-Dahab, R.; Afifi, F.U. Volatile oil composition and antiproliferative activity of Laurus nobilis, Origanum syriacum, Origanum vulgare, and Salvia triloba against human breast adenocarcinoma cells. Nutr. Res. 2010, 30, 271–278. [Google Scholar] [CrossRef]
- WHO Cancer Fact Sheet No. 296. Available online: http://www.who.int/mediacentre/factsheets/fs297/es/ (accessed on 20 January 2022).
- Ferlay, J.; Héry, C.; Autier, P.; Sankaranarayanan, R. Global Burden of Breast Cancer; Springer: New York, NY, USA, 2010. [Google Scholar]
- Ullah, M.F. Cancer multidrug resistance (MDR): A major impediment to effective chemotherapy. Asian Pac. J. Cancer Prev. 2008, 9, 1–6. [Google Scholar]
- Henderson, J.W.; Donatelle, R.J. Complementary and alternative medicine use by women after completion of allopathic treatment for breast cancer. Altern. Ther. Health Med. 2004, 10, 52–57. [Google Scholar]
- Boon, H.S.; Olatunde, F.; Zick, S.M. Trends in complementary/alternative medicine use by breast cancer survivors: Comparing survey data from 1998 and 2005. BMC Women Health 2007, 7, 4. [Google Scholar] [CrossRef]
- Bower, A.M.; Real Hernandez, L.M.; Berhow, M.A.; de Mejia, E.G. Bioactive compounds from culinary herbs inhibit a molecular target for type 2 diabetes management, dipeptidyl peptidase IV. J. Agric. Food Chem. 2014, 62, 6147–6158. [Google Scholar] [CrossRef] [PubMed]
- Timoteo, P.; Karioti, A.; Leitao, S.G.; Vincieri, F.F.; Bilia, A.R. A validated HPLC method for the analysis of herbal teas from three chemotypes of Brazilian Lippia alba. Food Chem. 2015, 175, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Grijalva, E.P.; Angulo-Escalante, M.A.; Leon-Felix, J.; Heredia, J.B. Effect of in vitro digestion on the total antioxidant capacity and phenolic content of 3 species of oregano (Hedeoma patens, Lippia graveolens, Lippia palmeri). J. Food Sci. 2017, 82, 2832–2839. [Google Scholar] [CrossRef] [PubMed]
- García-Mier, L.; Guevara-González, R.G.; Mondragón-Olguín, V.M.; Verduzco-Cuellar, B.d.R.; Torres-Pacheco, I. Agriculture and bioactives: Achieving both crop yield and phytochemicals. Int. J. Mol. Sci. 2013, 14, 4203–4222. [Google Scholar] [CrossRef]
- Yin, H.; Frette, X.C.; Christensen, L.P.; Grevsen, K. Chitosan oligosaccharides promote the content of polyphenols in Greek oregano (Origanum vulgare ssp. hirtum). J. Agric. Food Chem. 2012, 60, 136–143. [Google Scholar] [CrossRef]
- Yan, F.; Azizi, A.; Janke, S.; Schwarz, M.; Zeller, S.; Honermeier, B. Antioxidant capacity variation in the oregano (Origanum vulgare L.) collection of the German National Genebank. Ind. Crop. Prod. 2016, 92, 19–25. [Google Scholar] [CrossRef]
- Hennebelle, T.; Sahpaz, S.; Gressier, B.; Joseph, H.; Bailleul, F. Antioxidant and neurosedative properties of polyphenols and iridoids from Lippia alba. Phytother. Res. 2008, 22, 256–258. [Google Scholar] [CrossRef]
- Proestos, C.; Komaitis, M. Analysis of naturally occurring phenolic compounds in aromatic plants by RP-HPLC coupled to diode array detector (DAD) and GC-MS after silylation. Foods 2013, 2, 90–99. [Google Scholar] [CrossRef]
- Taamalli, A.; Arraez-Roman, D.; Abaza, L.; Iswaldi, I.; Fernandez-Gutierrez, A.; Zarrouk, M.; Segura-Carretero, A. LC-MS-based metabolite profiling of methanolic extracts from the medicinal and aromatic species Mentha pulegium and Origanum majorana. Phytochem. Anal. 2015, 26, 320–330. [Google Scholar] [CrossRef]
- Lin, L.-Z.; Mukhopadhyay, S.; Robbins, R.J.; Harnly, J.M. Identification and quantification of flavonoids of Mexican oregano (Lippia graveolens) by LC-DAD-ESI/MS analysis. J. Food Compos. Anal. 2007, 20, 361–369. [Google Scholar] [CrossRef]
- Koldas, S.; Demirtas, I.; Ozen, T.; Demirci, M.A.; Behcet, L. Phytochemical screening, anticancer and antioxidant activities of Origanum vulgare L. ssp. viride (Boiss.) Hayek, a plant of traditional usage. J. Sci. Food Agric. 2015, 95, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Stashenko, E.E.; Martinez, J.R.; Cala, M.P.; Duran, D.C.; Caballero, D. Chromatographic and mass spectrometric characterization of essential oils and extracts from Lippia (Verbenaceae) aromatic plants. J. Sep. Sci. 2013, 36, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Veciana Galindo, C.; Cortés Castell, E.; Torro Montell, L.; Sirvent Segura, E.; Rizo-Baeza, M.; Gil Guillén, V. Evaluación de la citotoxicidad y bioseguridad de un extracto de polifenoles de huesos de aceitunas. Nutr. Hosp. 2014, 29, 1388–1393. [Google Scholar] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Rad. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Chao, J.I.; Su, W.C.; Liu, H.F. Baicalein induces cancer cell death and proliferation retardation by the inhibition of CDC2 kinase and survivin associated with opposite role of p38 mitogen-activated protein kinase and AKT. Mol. Cancer Ther. 2007, 6, 3039–3048. [Google Scholar] [CrossRef]
- Kapinova, A.; Kubatka, P.; Golubnitschaja, O.; Kello, M.; Zubor, P.; Solar, P.; Pec, M. Dietary phytochemicals in breast cancer research: Anticancer effects and potential utility for effective chemoprevention. Environ. Health Prev. Med. 2018, 23, 36. [Google Scholar] [CrossRef]
- Li, T.; Zhu, J.; Guo, L.; Shi, X.; Liu, Y.; Yang, X. Differential effects of polyphenols-enriched extracts from hawthorn fruit peels and fleshes on cell cycle and apoptosis in human MCF-7 breast carcinoma cells. Food Chem. 2013, 141, 1008–1018. [Google Scholar] [CrossRef]
- Hilbig, J.; Policarpi, P.B.; Grinevicius, V.; Mota, N.; Toaldo, I.M.; Luiz, M.T.B.; Pedrosa, R.C.; Block, J.M. Aqueous extract from pecan nut [Carya illinoinensis (Wangenh) C. Koch] shell show activity against breast cancer cell line MCF-7 and Ehrlich ascites tumor in Balb-C mice. J. Ethnopharmacol. 2018, 211, 256–266. [Google Scholar] [CrossRef]
- Vukovic, N.L.; Obradovic, A.D.; Vukic, M.D.; Jovanovic, D.; Djurdjevic, P.M. Cytotoxic, proapoptotic and antioxidative potential of flavonoids isolated from propolis against colon (HCT-116) and breast (MDA-MB-231) cancer cell lines. Food Res. Int. 2018, 106, 71–80. [Google Scholar] [CrossRef]
- Kawaii, S.; Tomono, Y.; Katase, E.; Ogawa, K.; Yano, M. Antiproliferative activity of flavonoids on several cancer cell lines. Biosci. Biotechnol. Biochem. 1999, 63, 896–899. [Google Scholar] [CrossRef]
- Kandaswami, C.; Lee, L.T.; Lee, P.P.; Hwang, J.J.; Ke, F.C.; Huang, Y.T.; Lee, M.T. The antitumor activities of flavonoids. In Vivo 2005, 19, 895–909. [Google Scholar]
- Ravishankar, D.; Rajora, A.K.; Greco, F.; Osborn, H.M. Flavonoids as prospective compounds for anti-cancer therapy. Int. J. Biochem. Cell Biol. 2013, 45, 2821–2831. [Google Scholar] [CrossRef] [PubMed]
- Mojzis, J.; Varinska, L.; Mojzisova, G.; Kostova, I.; Mirossay, L. Antiangiogenic effects of flavonoids and chalcones. Pharmacol. Res. 2008, 57, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Xu, W.; Shi, X.; Zhang, Z. Dietary flavonoids as cancer prevention agents. J. Environ. Sci. Health. C 2011, 29, 1–31. [Google Scholar] [CrossRef]
- Link, A.; Balaguer, F.; Goel, A. Cancer chemoprevention by dietary polyphenols: Promising role for epigenetics. Biochem. Pharmacol. 2010, 80, 1771–1792. [Google Scholar] [CrossRef]
- Cavalcante, F.P.; Millen, E.C.; Zerwes, F.P.; Novita, G.G. Progress in Local Treatment of Breast Cancer: A Narrative Review. Rev. Bras. Ginecol. Obstet. 2020, 42, 356–364. [Google Scholar] [CrossRef]
- Szostakowska, M.; Trębińska-Stryjewska, A.; Grzybowska, E.A.; Fabisiewicz, A. Resistance to endocrine therapy in breast cancer: Molecular mechanisms and future goals. Breast Cancer Res. Treat. 2019, 173, 489–497. [Google Scholar] [CrossRef]
- Cao, L.; Niu, Y. Triple negative breast cancer: Special histological types and emerging therapeutic methods. Cancer Biol. Med. 2020, 17, 293–306. [Google Scholar] [CrossRef]
- Boice, A.; Bouchier-Hayes, L. Targeting apoptotic caspases in cancer. Biochim. Biophys. Acta 2020, 1867, 118688. [Google Scholar] [CrossRef]
- Leri, M.; Scuto, M.; Ontario, M.L.; Calabrese, V.; Calabrese, E.J.; Bucciantini, M.; Stefani, M. Healthy Effects of Plant Polyphenols: Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 1250. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef] [PubMed]
- Karadag, A.; Ozcelik, B.; Saner, S. Review of Methods to Determine Antioxidant Capacities. Food Anal. Met. 2009, 2, 41–60. [Google Scholar] [CrossRef]
- Swain, T.; Hillis, W.E. The phenolic constituents of Prunus domestica I.-The quantitative analysis of phenolic constituents. J. Sci. Food Agric. 1959, 10, 63–68. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oregano Species | TPC (mg of GAE/g) | DPPH (mMol TE/g) | ORAC (µMol TE/g) |
|---|---|---|---|
| H. patens | 99.58 ± 0.42 c | 2140.89 ± 0.26 c | 575.42 ± 33.23 c |
| L. graveolens | 143.87 ± 1.29 a | 2523.67 ± 0.36 a | 3870.01 ± 27.41 a |
| L. palmeri | 114.30 ± 4.48 b | 2301.93 ± 0.21 b | 1365.54 ± 106.46 b |
| Oregano Species | |||||||
|---|---|---|---|---|---|---|---|
| MS | [M-H]- | Fragmentation Pattern | Tentative Identification | Classification | H. pattens | L. graveolens | L. palmeri |
| 170.02 | 169.01 | 125.02 *, 168.83, 170.83 | Gallic acid | Phenolic acids (hydroxybenzoic acid derivative) | + | + | + |
| 354.09 | 353.09 | 179.03, 191.05, 354.09 | Chlorogenic acid | Phenolic acids (hydroxycinnamic acid derivative) | + | ||
| 168.04 | 167.03 | 108.02, 123.04, 152.01 | Vanillic acid | Phenolic acids (hydroxybenzoic acid derivative) | + | + | + |
| 180.04 | 179.03 | 134.03, 135.04, 178.84 | Caffeic acid | Phenolic acids (hydroxycinnamic acid derivative) | + | + | + |
| 288.06 | 287.05 | 151.00, 288.06 | Eriodictyol | Flavanone | + | + | + |
| 506.01 | 505.09 | 463.08, 506.09 | Quercetin-3-O-glucose-6′′-acetate | Flavonols | + | + | |
| 448.1 | 447.09 | 448.09, 449.10 | Luteolin-7-glucoside | Flavone | + | + | + |
| 462.08 | 461.07 | 285.04, 462.07 | Kempferol-3-glucuronide | Flavonols | + | + | |
| 464.08 | 463.08 | 300.02, 302.02, 464.09 | Quercetin-3-O-hexoside | Flavonols | + | ||
| 578.16 | 577.15 | 269.04, 270.05, 578.15 | Apigenin-7-neohesperidoside | Flavone | + | ||
| 446.08 | 445.07 | 175.02, 269.04, 446.08 | Baicalin | Flavone | + | + | + |
| 302.04 | 301.03 | 302.03, 303.04 | Quercetin | Flavonols | + | + | |
| 436.13 | 435.13 | 273.07, 436.13 | Phloridzin | Dihydrochalcone | + | + | |
| 286.04 | 285.04 | 151.00, 286.04 | Luteolin | Flavone | + | + | + |
| 270.05 | 269.04 | 151.00, 270.05 | Apigenin | Flavone | + | + | |
| 272.06 | 271.06 | 151.00, 177.02, 269.04 | Naringenin | Flavanone | + | + | |
| 360.08 | 359.08 | 161.02, 197.05, 360.23 | Rosmarinic acid | Phenolic acids (hydroxycinnamic acid derivative) | + | + | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Criollo-Mendoza, M.S.; Ramos-Payán, R.; Contreras-Angulo, L.A.; Gutiérrez-Grijalva, E.P.; León-Félix, J.; Villicaña, C.; Angulo-Escalante, M.A.; Heredia, J.B. Cytotoxic Activity of Polyphenol Extracts from Three Oregano Species: Hedeoma patens, Lippia graveolens and Lippia palmeri, and Antiproliferative Potential of Lippia graveolens against Two Types of Breast Cancer Cell Lines (MDA-MB-231 and MCF-7). Molecules 2022, 27, 5240. https://doi.org/10.3390/molecules27165240
Criollo-Mendoza MS, Ramos-Payán R, Contreras-Angulo LA, Gutiérrez-Grijalva EP, León-Félix J, Villicaña C, Angulo-Escalante MA, Heredia JB. Cytotoxic Activity of Polyphenol Extracts from Three Oregano Species: Hedeoma patens, Lippia graveolens and Lippia palmeri, and Antiproliferative Potential of Lippia graveolens against Two Types of Breast Cancer Cell Lines (MDA-MB-231 and MCF-7). Molecules. 2022; 27(16):5240. https://doi.org/10.3390/molecules27165240
Chicago/Turabian StyleCriollo-Mendoza, Marilyn S., Rosalío Ramos-Payán, Laura A. Contreras-Angulo, Erick P. Gutiérrez-Grijalva, Josefina León-Félix, Claudia Villicaña, Miguel A. Angulo-Escalante, and J. Basilio Heredia. 2022. "Cytotoxic Activity of Polyphenol Extracts from Three Oregano Species: Hedeoma patens, Lippia graveolens and Lippia palmeri, and Antiproliferative Potential of Lippia graveolens against Two Types of Breast Cancer Cell Lines (MDA-MB-231 and MCF-7)" Molecules 27, no. 16: 5240. https://doi.org/10.3390/molecules27165240
APA StyleCriollo-Mendoza, M. S., Ramos-Payán, R., Contreras-Angulo, L. A., Gutiérrez-Grijalva, E. P., León-Félix, J., Villicaña, C., Angulo-Escalante, M. A., & Heredia, J. B. (2022). Cytotoxic Activity of Polyphenol Extracts from Three Oregano Species: Hedeoma patens, Lippia graveolens and Lippia palmeri, and Antiproliferative Potential of Lippia graveolens against Two Types of Breast Cancer Cell Lines (MDA-MB-231 and MCF-7). Molecules, 27(16), 5240. https://doi.org/10.3390/molecules27165240