
Metabolic Profile of Agropyron repens (L.) P. Beauv. Rhizome Herbal Tea by HPLC-PDA-ESI-MS/MS Analysis






Abstract
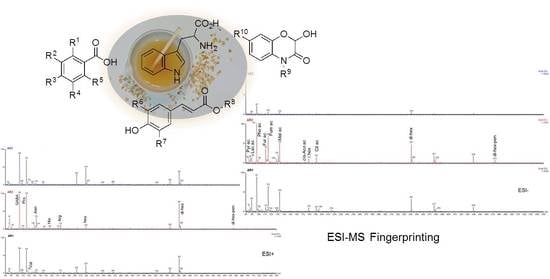
1. Introduction
2. Results and Discussion
2.1. Untargeted ESI-MS/MS Profile
2.1.1. Saccharides
2.1.2. Small Organic Acids
2.1.3. Amino Acids
2.2. HPLC-PDA-ESI-MS/MS Targeted Analysis
3. Materials and Methods
3.1. Chemicals and Solvents
3.2. Plant Material Characterization
3.3. Agropyron repens (L.) P. Beauv. Rhizome Extraction Procedure
3.4. HPLC-PDA-ESI-MS/MS Analysis
3.5. Tryptophan Calibration Curve by HPLC-ESI-MS/MS
3.6. Untargeted ESI-MS/MS Profile
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Touchette, R.; Leroux, G.D.; Deschênes, J.M. Allelopathic activity of quackgrass (Agropyron repens) extracts and residues on alfalafa (Medicago sativa). Can. J. Plant. Sci. 1988, 68, 785–792. [Google Scholar] [CrossRef]
- Alosy, B.D.M.; Thakir, M.; Khalaf, S.A. Role of Agropyron repens extract in treatment renal calculus in pediatric group. Indian J. Forensic Med. Toxicol. 2019, 13, 405–409. [Google Scholar] [CrossRef]
- Tsubanova, N.A.; Barska, A.V.; Cherniavski, E.S. Clinical efficiency of preparations based on medical plant raw materials in the treatment of urolithiasis. Fam. Med. 2019, 81, 80–87. [Google Scholar] [CrossRef][Green Version]
- Kasote, D.M.; Jagtap, S.D.; Thapa, D.; Khyade, M.S.; Russell, W.R. Herbal remedies for urinary stones used in India and China: A review. J. Ethnopharmacol. 2017, 203, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Zimdahl, R.L. Allelopathy: Detrimental Effects on Other Plants Growth Due to the Production of Chemical Inhibitors, Fundamentals of Weed Science, 5th ed.; Academic Press, Elsevier: Amsterdam, The Netherlands, 2018; pp. 253–270. [Google Scholar]
- Al-Snafi, A.E. Chemical constituent and pharmacological importance of Agropyron repens, A review. Res. J. Pharmacol. Toxicol. 2015, 1, 37–41. [Google Scholar]
- Eddouks, M.; Maghrani, M.; Michel, J.B. Hypoglycaemic effect of Triticum repens P. Beauv. in normal and diabetic rats. J. Ethnopharmacol. 2005, 102, 228–232. [Google Scholar] [CrossRef]
- European Medicines Agency. Assessment Report on Agropyron Repens (L.) P. Beauv., Rhizoma; EMA/HMPC/563395/2010; European Medicines Agency: London, UK, 2011. [Google Scholar]
- Mascolo, N. Biological screening of Italian medicinal plants for anti-inflammatory activity. Phytother. Res. 1987, 1, 28–29. [Google Scholar] [CrossRef]
- Petrova, A.P.; Krasnov, E.A.; Saprykina, E.V.; Subbotina, Y.A.; Ermilova, E.V. Chemical composition of couch grass and studies of its antioxidant activity in allergic contact dermatitis. Pharm. Chem. J. 2009, 43, 30–32. [Google Scholar] [CrossRef]
- Neagu, E.; Păun, G.; Moroeanu, V.; Ungureanu, O.; Radu, G.L. Antioxidant ant antidiabetic properties of polyphenolic-rich extracts of Apium graveolens and Agropyrum repens. Rev. Roum. Chim. 2019, 64, 909–913. [Google Scholar] [CrossRef]
- Kiesewetter, R.; Möller, M. Zur Frage der “sedativen” Wirkung von Radix Valerianae. Pharmazie 1958, 13, 777–781. [Google Scholar]
- Hautmann, C.; Scheithe, K. Fluid extract of Agropyron repens for the treatment of urinary tract infections or irritable bladder. Results of multicentric post-marketing surveillance. Z. Phytother. 2000, 21, 252–255. [Google Scholar]
- Minář, J.; Mannsbartová, E.; Tichý, V. Izmyenyeniya biologi-chyeskoi, aktivnosti agropiryena priyego prim yenyenii s gumusoymi vye shchyestvami na kul-turuvodorosli Scenedesmus obliquus (Turp.). Biol. Plant 1964, 6, 265–272. [Google Scholar] [CrossRef]
- Schilcher, H.; Boesel, R.; Effenberger, S.; Segebrecht, S. Recent results of investigations concerning medicinal plants with “aquaretic”, antibacterial and prostatic effects. Z. Phytother. 1989, 10, 77–82. [Google Scholar]
- Boesel, R.; Schilcher, H. Composition of the essential oil of Agropyron repens rhizome. Planta Med. 1989, 55, 399–400. [Google Scholar] [CrossRef]
- Gabor, W.E.; Veatch, C. Isolation of a Phytotoxin from Quackgrass (Agropyron repens) Rhizomes. Weed Sci. 1981, 29, 155–159. [Google Scholar] [CrossRef]
- Weston, L.A.; Putnam, A.R. Inhibition of Legume Seedling Growth by Residues and Extracts of Quackgrass (Agropyron repens). Weed Sci. 1986, 34, 366–372. [Google Scholar] [CrossRef]
- Weston, L.A.; Burke, B.A.; Putnam, A.R. Isolation, characterization and activity of phytotoxic compounds from quack grass. J. Chem. Ecol. 1987, 13, 403–421. [Google Scholar] [CrossRef]
- Hagin, R.D.; Bobnick, S.J. Isolation and identification of a slug-specific molluscicide from quackgrass (Agropyron repens L. Beauv.). J. Agric. Food Chem. 1991, 39, 192–196. [Google Scholar] [CrossRef]
- Koetter, U.; Kaloga, M.; Schilcher, H. Isolation Isolation and structure elucidation of p-hydroxycinnamic acid esters from the rhizon of Agropyron repens, Part I. Planta Med. 1993, 59, 279–280. [Google Scholar] [CrossRef]
- Koetter, U.; Kaloga, M.; Schilcher, H. Isolation and structure elucidation of p-hydroxycinnamic acid esters from the rhizom of Agropyron repens, Part II. Planta Med. 1994, 60, 488–489. [Google Scholar] [CrossRef]
- Beydokthi, S.S.; Sendker, J.; Brandt, S.; Hensel, A. Traditionally used medicinal plants against uncomplicated urinary tract infections: Hexadecyl coumaric acid ester from the rhizomes of Agropyron repens (L.) P. Beauv. with antiadhesive activity against uropathogenic E. coli. Fitoterapia 2017, 117, 22–27. [Google Scholar] [CrossRef]
- Kubus, G.; Tłuścik, F. Alkyl resorcinols in grains from plants from the family Gramineae. Acta Soc. Bot. Pol. 1983, 52, 223–230. [Google Scholar] [CrossRef]
- Korhammer, S.A.; Haslinger, E. Isolation of a biologically active substance from rhizomes of quackgrass, Elymus repens (L.) Gould. J. Agric. Food Chem. 1994, 42, 2048–2050. [Google Scholar] [CrossRef]
- Schulz, M.; Friebe, A.; Kück, P.; Seipel, M.; Schnabl, H. Allelopathic effects of living quackgrass (Agropyron repens L.). Identification of inhibitory allelochemicals exuded from rhizome borne troots. Angew. Bot. 1994, 68, 195–200. [Google Scholar]
- Friebe, A.; Schulz, M.; Kück, P.; Schnabl, H. Phytotoxins from shoot extract and root exudates of Agropyron repens seedlings. Phytochemistry 1995, 38, 1157–1159. [Google Scholar] [CrossRef]
- Hagin, R.D. Isolation and Identification of 5-Hydroxyindole-3-acetic Acid and 5-Hydroxytryptophan, Major Allelopathic Aglyconsi in Quackgrass (Agropyron repens L. Beauv.). J. Agric. Food Chem. 1989, 37, 1143–1149. [Google Scholar] [CrossRef]
- Maffei, M.E. 5-Hydroxytryptophan (5-HTP): Natural occurrence, analysis, biosynthesis, biotechnology, physiology and toxicology. Int. J. Mol. Sci. 2021, 22, 181. [Google Scholar] [CrossRef]
- Palego, L.; Betti, L.; Rossi, A.; Giannaccini, G. Tryptophan Biochemistry: Structural, Nutritional, Metabolic, and Medical Aspects in Humans. J. Amino Acids 2016, 2016, 8952520. [Google Scholar] [CrossRef]
- Barik, S. The Uniqueness of Tryptophan in Biology: Properties, Metabolism, Interactions and Localization in Proteins. Int. J. Mol. Sci. 2020, 21, 8776. [Google Scholar] [CrossRef]
- Friedman, M. Analysis, Nutrition, and Health Benefits of Tryptophan. Int. J. Tryptophan Res. 2018, 11, 1–12. [Google Scholar] [CrossRef]
- European Medicines Agency. Assessment Report on Agropyron Repens (L.) P. Beauv., Rhizoma; EMA/HMPC/113793/2021; European Medicines Agency: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Graminis Rhizoma (Couch Grass Rhizome)- ESCOP Herbal Monograph 2016, The Scientific Foundation for Herbal Medicinal Products, Ed. Available online: www.escop.com (accessed on 11 February 2022).
- Arni, P.C.; Percival, E.G.V. Studies on Fructosans. Part II. Triticin from the Rhizomes of Couch Grass (Triticum repens L.). J. Chem. Soc. 1951, 399, 1822–1830. [Google Scholar] [CrossRef]
- Stoop, J.M.H.; Williamson, J.D.; Pharr, D.M. Mannitol metabolism in plants: A method for coping with stress. Trends Plant. Sci. 1996, 1, 139–144. [Google Scholar] [CrossRef]
- Whitehead, D.C.; Dibb, H.; Hartley, R.D. Bound phenolic compounds in water extracts of soils, plant roots and leaf litter. Soil Biol. Biochem. 1983, 15, 133–136. [Google Scholar] [CrossRef]
- Stanić, G.; Gavrić, D.; Šimić, I. Phytochemical study of Elymus repens Gould and Cynodon dactylon (L.) pers. Farm. Glasnik. 2000, 56, 1–9. [Google Scholar]
- Mueller, S.O.; Schmitt, M.; Dekant, W.; Stopper, H.; Schlatter, J.; Schreier, P.; Lutz, W.K. Occurrence of emodin, chrysophanol and physcion in vegetables, herbs and liquors. Genotoxicity and anti-genotoxicity of the anthraquinones and of the whole plants. Food Chem. Toxicol. 1999, 37, 481–491. [Google Scholar] [CrossRef]
- Paslawska, S.; Piekos, R. Studies on the optimum conditions of extraction of silicon species from plants with water IV. Agropyron repens. Planta Med. 1976, 30, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.M.; Hall, K.C.; Anderson, H.A.; Hepburn, A. Organic acids from the anaerobic decomposition of Agropyron repens rhizomes. Phytochemistry 1980, 19, 1846–1847. [Google Scholar] [CrossRef]
- Araújo, A.S.; da Rocha, L.L.; Tomazela, D.M.; Sawaya, A.C.H.F.; Almeida, R.R.; Catharino, R.R.; Eberlin, M.N. Electrospray ionization mass spectrometry fingerprinting of beer. Analyst 2005, 130, 884–889. [Google Scholar] [CrossRef]
- Petrucci, R.; Di Matteo, P.; De Francesco, G.; Mattiello, L.; Perretti, G.; Russo, P. Novel fast identification and determination of free polyphenols in untreated craft beers by HPLC-PDA-ESI-MS/MS in SIR mode. J. Agric. Food Chem. 2020, 68, 7984–7994. [Google Scholar] [CrossRef]
- Pinto, G.; Aurilia, M.; Illiano, A.; Fontanarosa, C.; Sannia, G.; Trifuoggi, M.; Lettera, V.; Sperandeo, R.; Pucci, P.; Amoresano, A. From untargeted metabolomics to the multiple reaction monitoring-based quantification of polyphenols in chocolates from different geographical areas. J. Mass Spectrom. 2021, 56, e4651. [Google Scholar] [CrossRef]
- Bald, I.; Flosadóttir, H.D.; Kopyra, J.; Illenberger, E.; Ingólfsson, O. Fragmentation of deprotonated d-ribose and d-fructose in MALDI—Comparison with dissociative electron attachment. Int. J. Mass Spectrom. 2009, 280, 190–197. [Google Scholar] [CrossRef]
- Taylor, V.F.; March, R.E.; Longerich, H.P.; Stadey, C.J. A mass spectrometric study of glucose, sucrose, and fructose using an inductively coupled plasma and electrospray ionization. Int. J. Mass Spectrom. 2005, 243, 71–84. [Google Scholar] [CrossRef]
- Beneito-Cambra, M.; Bernabé-Zafón, V.; Herrero-Martínez, J.M.; Ramis-Ramos, G. Study of the Fragmentation of D-Glucose and Alkylmonoglycosides in the Presence of Sodium Ions in an Ion-Trap Mass Spectrometer. Anal. Lett. 2009, 42, 907–921. [Google Scholar] [CrossRef]
- European Tea Committee (ETC) and European Herbal Infusions Association (EHIA). Compendium of Guidelines for Herbal and Fruit Infusions (Former EHIA Document), Issue 6, 22 June 2018. Available online: https://thie-online.eu/files/thie/docs/thie-statutes_new.pdf (accessed on 11 February 2022).
- Farouk, M.; El-Ganiny, A.; Abdelmonem, M.; Serry, F. Evaluation of microbial quality of herbal tea pharmaceutical products found in the Egyptian market. Zagazig J. Pharm. Sci. 2017, 26, 32–38. [Google Scholar]
- Abualhasan, M.N.; Jaradat, N.; Hawash, M.; Khayat, R.; Khatatbeh, E.; Ehmidan, M.; Al-Atrash, M. Evaluation of Heavy Metal and Microbial Contamination in Green Tea and Herbal Tea Used for Weight Loss in the Palestinian Market. Evid.-Based Complementary Altern. Med. 2020, 2020, 7631562. [Google Scholar] [CrossRef] [PubMed]
- Bantle, J.P. Is fructose the optimal low glycemic index sweetener? Nestle Nutr. Workshop Ser. Clin. Perform Programme 2006, 11, 83–95. [Google Scholar] [PubMed]
- Bauer, A.; Luetjohann, J.; Rohn, S.; Kuballa, J.; Jantzen, E. Determination of Fosetyl and Phosphonic Acid at 0.010 mg/kg Level by Ion Chromatography Tandem Mass Spectrometry. J. Agric. Food Chem. 2018, 66, 346–350. [Google Scholar] [CrossRef]
- Zawadzki, M.; Luxford, T.F.M.; Kočišek, J. Carboxylation Enhances Fragmentation of Furan upon Resonant Electron Attachment. J. Phys. Chem. A 2020, 124, 9427–9435. [Google Scholar] [CrossRef]
- Dong, R.; Yu, Q.; Liao, W.; Liu, S.; He, Z.; Hu, X.; Chen, Y.; Xie, J.; Nie, S.; Xie, M. Composition of bound polyphenols from carrot dietary fiber and its in vivo and in vitro antioxidant activity. Food Chem. 2021, 339, 127879. [Google Scholar] [CrossRef] [PubMed]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef]
- Chuang, C.K.; Wang, T.J.; Yeung, C.Y.; Lin, D.S.; Lin, H.Y.; Liu, H.L.; Ho, H.T.; Hsieh, W.S.; Lin, S.P. A method for lactate and pyruvate determination in filter-paper dried blood spots. J. Chromatogr. A 2009, 1216, 8947–8952. [Google Scholar] [CrossRef]
- Tan, B.; Malu, S.; Roth, K.D. Development of ion pairing LC-MS/MS method for itaconate and cisaconitate in cell extract and cell media. J. Chromatogr. B 2020, 1146, 122120. [Google Scholar] [CrossRef]
- Carvalho, E.; Punyasiri, P.A.N.; Somasiri, H.P.P.S.; Abeysinghe, S.B.; Martens, S. Quantification of γ-Aminobutyric Acid in Sri Lankan Tea by Means of Ultra Performance Tandem Mass Spectrometry. Nat. Prod. Commun. 2014, 9, 525–528. [Google Scholar] [CrossRef]
- Thiele, B.; Füllner, K.; Stein, N.; Oldiges, M.; Kuhn, A.J.; Hofmann, D. Analysis of amino acids without derivatization in barley extracts by LC-MS-MS. Anal. Bioanal. Chem. 2008, 391, 2663–2672. [Google Scholar] [CrossRef]
- Nigam, S.N.; McIntyre, G.I. Apical dominance in the rhizome of Agropyron repens. The relation of amino acids composition to bud activity. Can. J. Bot. 1977, 55, 2001–2010. [Google Scholar] [CrossRef]
- Petrucci, R.; Di Matteo, P.; Sobolev, A.P.; Liguori, L.; Albanese, D.; Proietti, N.; Bortolami, M.; Russo, P. Impact of dealcoholisation by osmotic distillation on metabolic profile, phenolic content and antioxidant capacity of low alcoholic craft beers with different malt composition. J. Agric. Food Chem. 2021, 69, 4816–4826. [Google Scholar] [CrossRef]
- Di Matteo, P.; Bortolami, M.; Di Virgilio, L.; Petrucci, R. Targeted phenolic profile of radler beers by HPLC-ESI-MS/MS: The added value of hesperidin to beer antioxidants. J. Food Sci. Technol. 2022. [Google Scholar] [CrossRef]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical scheme for LC-MSn identification of chlorogenic acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Taniguchi, H.; Yamada, M.; Matsukura, Y.; Koizumi, H.; Furihata, K.; Shindo, K. Analysis of the Components of Hard Resin in Hops (Humulus lupulus L.) and Structural Elucidation of Their Transformation Products Formed during the Brewing Process. J. Agric. Food Chem. 2014, 62, 11602–11612. [Google Scholar] [CrossRef]
- Trani, A.; Petrucci, R.; Marrosu, G.; Curulli, A. Determination of Caffeine Gold Nanoparticles Modified Gold (Au) Electrode: A Preliminary Study, in Lecture Notes in Electrical Engineering, Sensors; Springer International Publishing: Cham, Switzerland, 2015; Chapter 26; Volume 319, pp. 147–151. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Compounds | Quantitation | Extraction Solvent | Ref. |
|---|---|---|---|---|
| Polysaccharides | fructosan (triticin), inulin | ~12% | water | [10,35] |
| Disaccharides | not identified | water | [35] | |
| Monosaccharides | fructose, glucose, rhamnose | (17.1, 3.4, nd%) | water | [10,35] |
| Sugar alcohols | mannitol, inositol | nd | nr | [6,36] |
| Phenolic acids, free (and bound) | chlorogenic acid | nd | nr | [10] |
| p-coumaric acid | 87 (24) μg/g | water | [37] | |
| p-hydroxybenzoic acid | 10 (55) μg/g | water | [37] | |
| vanillic acid | 27 (11) μg/g | water | [37] | |
| ferulic acid | 38 (12) μg/g | water | [37] | |
| vanillin | 4 (17) μg/g | water | [37] | |
| p-hydroxybenzaldehyde | tr (4) μg/g | water | [37] | |
| Flavonoids | tricin | nd | nr | [10,19,38] |
| rutin | ||||
| hyperoside | ||||
| baicalein | ||||
| quercetin | ||||
| luteolin | ||||
| Tannins | 5% | water | [10] | |
| Antraquinones | emodin | 0.06–0.2 μg/g | acetonitrile/water | [39] |
| chrysophanol | 0.05–0.2 μg/g | |||
| physcion | 0.08–0.3 μg/g | |||
| Si | silicon species | 0.4% | water | [40] |
| Main Volatiles | palmitic acid | 0.05% | steam distillation | [6,16] |
| carvacrol | ||||
| trans-anethole | ||||
| carvone | ||||
| thymol | ||||
| menthol | ||||
| menthone | ||||
| Vitamins | ascorbic acid | nd | nr | [10] |
| β-carotene | ||||
| Organic acids | acetic acid | nd | water | [10,41] |
| propionic acid | ||||
| butyric acid | ||||
| hexanoic acid | ||||
| phenylacetic acid | ||||
| succinic acid | ||||
| cinnamic acid | ||||
| p-hydroxyphenylpropionic acid | ||||
| p-coumaric acid | ||||
| 3,4-dihydroxyphenylpropionic acid | ||||
| Total polyphenols contents | 257.95 μgGAE/mL | ethanol/water | [11] | |
| Total flavonoid contents | 70.89 μgRE/mL | ethanol/water | [11] | |
| Antioxidant activity | ~35% (DPPH) | ethanol/water | [11] | |
| ~60% (reducing power) | ethanol/water | [11] | ||
| Sample | Abs. H (g/m3) | aw (21.5 °C) | TAC (CFU/g) | Y&M (CFU/g) | LAB (CFU/g) | EB (CFU/g) |
|---|---|---|---|---|---|---|
| AR1 | 9.53 | 0.495 ± 0.001 | 1.41 ± 0.15 × 103 | 18.0 ± 0.10 | <10 | <10 |
| AR2 | 10.60 | 0.545 ± 0.001 | 3.05 ± 0.20 × 103 | 10.0 ± 0.20 | <10 | <10 |
| AR3 | 8.63 | 0.485 ± 0.001 | 3.83 ± 0.50 × 103 | 22.00 ± 0.14 | <10 | <10 |
| Organic Acid | Precursor [M-H]− (m/z) | Fragments (m/z) | Ref. |
|---|---|---|---|
| Pyr ac | 87 | 43 [M-H-CO2]− | [52] |
| Lac ac | 89 | 71 [M-H-H2O]−; 43 [M-H-HCO2H]− | [52] |
| Pho ac | 97 | 79 [M-H2O]− | [53] |
| Fur ac | 111 | 67 [M-H-CO2]−; 41 [M-H-CO2-C2H2]− | [54] |
| Fum ac | 115 | 71 [M-H-CO2]− | [55] |
| Mal ac | 133 | 115 [M-H-H2O]−; 89 [M-H-CO2]−; 71 [M-H-CO2-H2O]− | [55] |
| cis-Aco ac | 173 | 111 [M-CO2-H2O]− | [56] |
| Cit ac | 191 | 129 [M-H-CO2-H2O]−; 111 [M-H-CO2-2H2O]− | [55] |
| Amino Acid | Precursor [M+H]+ (m/z) | Fragments (m/z) | Ref. |
|---|---|---|---|
| GABA | 104 | 87 | [58] |
| 69 | [59] | ||
| Pro | 116 | 70 [C4H8N]+ | [60] |
| Val | 118 | 72 [M+H-HCO2H]+ | [60] |
| Asn | 133 | 87 [M+H-HCO2H]+; 74 [NH2=CHCO2H]+ | [60] |
| His | 156 | 110 [M+H-HCO2H]+ | [60] |
| Arg | 175 | 116 [M+H-NH2C(NH)=NH2]+; 70 [C4H8N]+ | [60] |
| Compound | tR (min) | AR1-IN (μg/g ± σ) | AR1-50 (μg/g ± σ) | AR1-25 (μg/g ± σ) | AR2-IN (μg/g ± σ) | AR2-50 (μg/g ± σ) | AR2-25 (μg/g ± σ) | AR3-IN (μg/g ± σ) | AR3-50 (μg/g ± σ) | AR3-25 (μg/g ± σ) | [M-H]− (m/z) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| GA | 3.15 | 0.70 ± 0.08 a,A | 2.78 ± 0.08 b | 0.63 ± 0.04 a | 0.67 ± 0.09 a,A | 0.60 ± 0.08 a | 0.61 ± 0.07 a | 2.80 ± 0.13 a,B | 1.90 ± 0.25 a | 5.10 ± 0.00 b | 169 |
| 3,5-DHBA | 4.61 | nd | nd | nd | nd | nd | nd | nd | nd | nd | 153 |
| PCA | 5.15 | 1.49 ± 0.19 a,A | 6.64 ± 0.25 b | 2.99 ± 0.12 c | 1.21 ± 0.09 a,A | 1.85 ± 0.08 a | 2.03 ± 0.05 b | 7.16 ± 0.56 a,B | 6.19 ± 0.19 a | 7.39 ± 0.92 a | 153 |
| TP | 5.51 | 19.92 ± 1.19 a,A | 20.52 ± 1.92 a | 12.61 ± 0.41 b | 203.66 ± 18.70 a,B | 116.20 ± 18.64 b | 196.05 ± 3.30 c | 13.01 ± 0.63 a,C | 20.76 ± 3.89 b | 20.99 ± 2.13 b | 203 |
| 5-CQA | 6.90 | 57.40 ± 4.40 a,A | 59.02 ± 5.08 a | 20.06 ± 0.60 b | 107.56 ± 7.04 a,B | 101.23 ± 0.59 b | 105.10 ± 6.02 b | 112.22 ± 5.96 a,C | 220.06 ± 22.42 b | 202.31 ± 26.06 b | 353 |
| GeA | 7.14 | 0.86 ± 0.02 a,A | 2.38 ± 0.03 b | 0.87 ± 0.06 a | 1.23 ± 0.06 a,B | 1.10 ± 0.09 a | 1.63 ± 0.19 b | 2.07 ± 0.27 a,B | 4.19 ± 0.34 a | 3.46 ± 0.34 b | 153 |
| Ca | 7.32 | nd | nd | nd | nd | nd | nd | nd | nd | nd | 289 |
| 4-CQA | 7.55 | 32.43 ± 0.65 a,A | 35.69 ± 0.64 b | 17.45 ± 1.75 c | 65.66 ± 3.92 a,B | 47.09 ± 1.56 b | 54.34 ± 6.24 b | 46.39 ± 4.75 a,C | 85.84 ± 2.55 b | 74.88 ± 8.81 c | 353 |
| p-HBA | 7.73 | 5.32 ± 0.47 a,A | 3.01 ± 0.78 b | 4.82 ± 0.59 a | 3.64 ± 0.47 a,B | 6.04 ± 0.57 b | 4.69 ± 0.82 a | 9.52 ± 0.94 a,C | 8.73 ± 0.55 b | 10.67 ± 1.00 b | 137 |
| 3-FQA | 7.77 | 9.99 ± 0.83 a,A | 53.70 ± 3.42 b | 13.03 ± 0.04 c | 53.99 ± 0.18 a,B | 33.43 ± 0.79 b | 65.19 ± 4.89 c | 16.08 ± 0.25 a,C | 25.01 ± 3.84 b | 56.21 ± 2.75 c | 367 |
| VA | 9.13 | 3.18 ± 0.24 a,A | 6.37 ± 0.19 b | 2.60 ± 0.21 c | 3.36 ± 0.24 a,A | 4.07 ± 0.36 b | 4.39 ± 0.42 b | 5.22 ± 0.54 a,B | 6.91 ± 0.59 b | 8.87 ± 0.38 c | 167 |
| CA | 9.45 | 2.55 ± 0.14 a,A | 34.88 ± 0.30 b | 16.57 ± 2.54 c | 6.98 ± 0.34 a,B | 25.90 ± 1.69 b | 12.13 ± 1.29 c | 6.91 ± 0.59 a,B | 26.86 ± 2.21 b | 86.52 ± 2.09 c | 179 |
| SyA | 9.56 | 1.89 ± 0.26 a,A | 3.25 ± 0.40 b | 1.76 ± 0.08 a | 1.95 ± 0.20 a,A | 1.77 ± 0.06 b | 2.20 ± 0.11 a | 3.69 ± 0.20 a,B | 4.59 ± 0.29 b | 4.80 ± 0.44 b | 197 |
| 2,6-DHBA | 10.48 | <LOD | 1.60 ± 0.06 | <LOD | 0.85 ± 0.11 a,A | 1.58 ± 0.12 b | 1.35 ± 0.06 b | 1.02 ± 0.08 a,A | 2.61 ± 0.05 b | 2.42 ± 0.16 b | 153 |
| m-HBA | 10.66 | nd | nd | nd | nd | nd | nd | nd | nd | nd | 137 |
| 4-FQA | 12.12 | 3.97 ± 0.35 a,A | 26.50 ± 1.37 b | 5.14 ± 0.38 c | 27.14 ± 2.82 a,B | 17.59 ± 0.06 b | 30.30 ± 2.97 a | 7.97 ± 0.18 a,C | 23.25 ± 3.52 b | 28.84 ± 3.23 c | 367 |
| CouA | 14.19 | 5.83 ± 0.56 a,A | 10.21 ± 1.41 b | 4.62 ± 0.03 c | 8.53 ± 0.27 a,B | 14.58 ± 0.40 b | 10.24 ± 0.67 c | 7.60 ± 0.08 a,C | 11.92 ± 1.38 b | 20.74 ± 1.94 c | 163 |
| EA | 15.74 | <LOD | <LOD | <LOD | <LOD | 0.18 ± 0.02 a | 1.24 ± 0.08 b | <LOD | nd | nd | 301 |
| SiA | 15.92 | 0.40 ± 0.06 a,A | 0.73 ± 0.11 b | 0.41 ± 0.11 a | 0.50 ± 0.03 a,A | 0.54 ± 0.04 a | 0.69 ± 0.10 a | 0.78 ± 0.03 a,B | 1.18 ± 0.10 b | 0.93 ± 0.09 b | 223 |
| FA | 16.13 | 1.97 ± 0.08 a,A | 7.55 ± 0.56 b | 2.36 ± 0.99 c | 3.58 ± 0.47 a,B | 11.13 ± 0.01 b | 6.29 ± 0.18 c | 4.05 ± 0.20 a,B | 8.28 ± 0.87 b | 13.98 ± 0.79 c | 193 |
| My | 16.74 | nd | nd | nd | nd | nd | 0.73 ± 0.07 | nd | nd | nd | 463 |
| Ru | 16.83 | nd | nd | nd | nd | nd | nd | nd | nd | nd | 609 |
| Q-3-G | 17.75 | nd | <LOD | <LOD | <LOD | 0.59 ± 0.03 a | 0.74 ± 0.06 b | nd | nd | 0.73 ± 0.03 | 463 |
| K-3-R | 20.07 | nd | nd | nd | <LOD | <LOD | 0.16 ± 0.02 | nd | <LOD | nd | 593 |
| SaA | 20.33 | 0.30 ± 0.04 a,A | 0.43 ± 0.03 a | 0.34 ± 0.05 a | 0.29 ± 0.52 a,A | 0.55 ± 0.08 b | 1.08 ± 0.11 c | 1.37 ± 0.15 a,B | 1.94 ± 0.19 b | 3.44 ± 0.17 c | 137 |
| He | 23.26 | 1.37 ± 0.22 a,A | 1.01 ± 0.02 a | 1.05 ± 0.04 a | 1.71 ± 0.28 a,B | 1.30 ± 0.07 b | 1.17 ± 0.12 b | 4.40 ± 0.58 a,C | 1.11 ± 0.11 b | 6.91 ± 0.58 c | 609 |
| Q | 27.76 | <LOD | 0.19 ± 0.01 | <LOD | <LOD | 0.16 ± 0.06 a | 1.66 ± 0.15 b | <LOD | nd | <LOD | 301 |
| K | 30.27 | <LOD | <LOD | <LOD | <LOD | <LOD | 0.42 ± 0.12 | <LOD | <LOD | <LOD | 285 |
| i-XH | 32.70 | 0.22 ± 0.005 a,A | 0.23 ± 0.002 a | 0.25 ± 0.01 a | 0.24 ± 0.005 a,A | 0.28 ± 0.004 a | 0.49 ± 0.12 b | nd | nd | nd | 353 |
| XH | 38.04 | <LOD | <LOD | <LOD | <LOD | 0.24 ± 0.02 a | 4.14 ± 0.21 b | nd | nd | nd | 353 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bortolami, M.; Di Matteo, P.; Rocco, D.; Feroci, M.; Petrucci, R. Metabolic Profile of Agropyron repens (L.) P. Beauv. Rhizome Herbal Tea by HPLC-PDA-ESI-MS/MS Analysis. Molecules 2022, 27, 4962. https://doi.org/10.3390/molecules27154962
Bortolami M, Di Matteo P, Rocco D, Feroci M, Petrucci R. Metabolic Profile of Agropyron repens (L.) P. Beauv. Rhizome Herbal Tea by HPLC-PDA-ESI-MS/MS Analysis. Molecules. 2022; 27(15):4962. https://doi.org/10.3390/molecules27154962
Chicago/Turabian StyleBortolami, Martina, Paola Di Matteo, Daniele Rocco, Marta Feroci, and Rita Petrucci. 2022. "Metabolic Profile of Agropyron repens (L.) P. Beauv. Rhizome Herbal Tea by HPLC-PDA-ESI-MS/MS Analysis" Molecules 27, no. 15: 4962. https://doi.org/10.3390/molecules27154962
APA StyleBortolami, M., Di Matteo, P., Rocco, D., Feroci, M., & Petrucci, R. (2022). Metabolic Profile of Agropyron repens (L.) P. Beauv. Rhizome Herbal Tea by HPLC-PDA-ESI-MS/MS Analysis. Molecules, 27(15), 4962. https://doi.org/10.3390/molecules27154962