
N-methyl Benzimidazole Tethered Cholic Acid Amphiphiles Can Eradicate S. aureus-Mediated Biofilms and Wound Infections

Abstract
1. Introduction
2. Results and Discussion
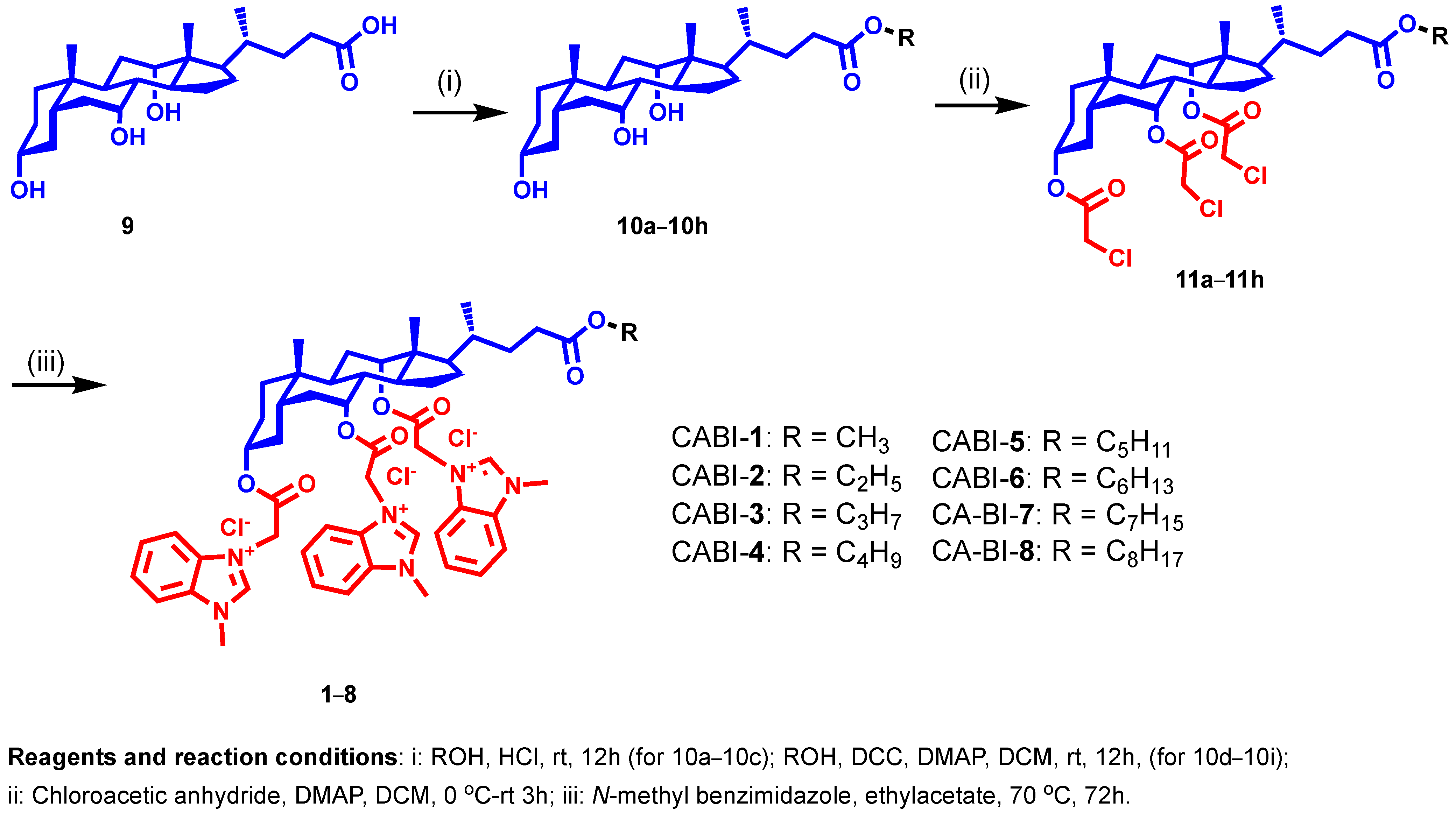
2.1. Design and Synthesis of CA-Benzimidazole Amphiphiles
2.2. Antibacterial Activities of CA-Benzimidazole Amphiphiles
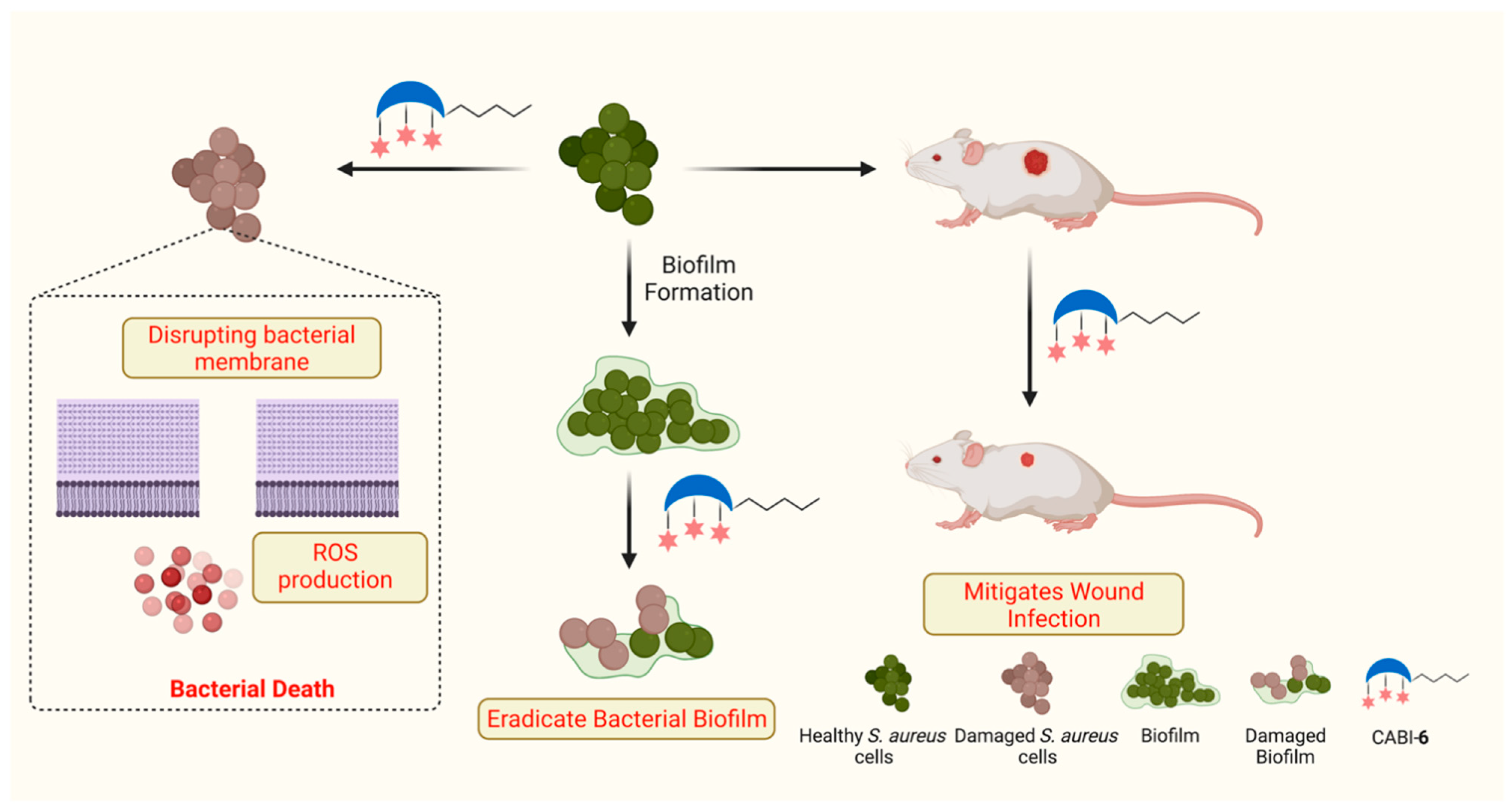
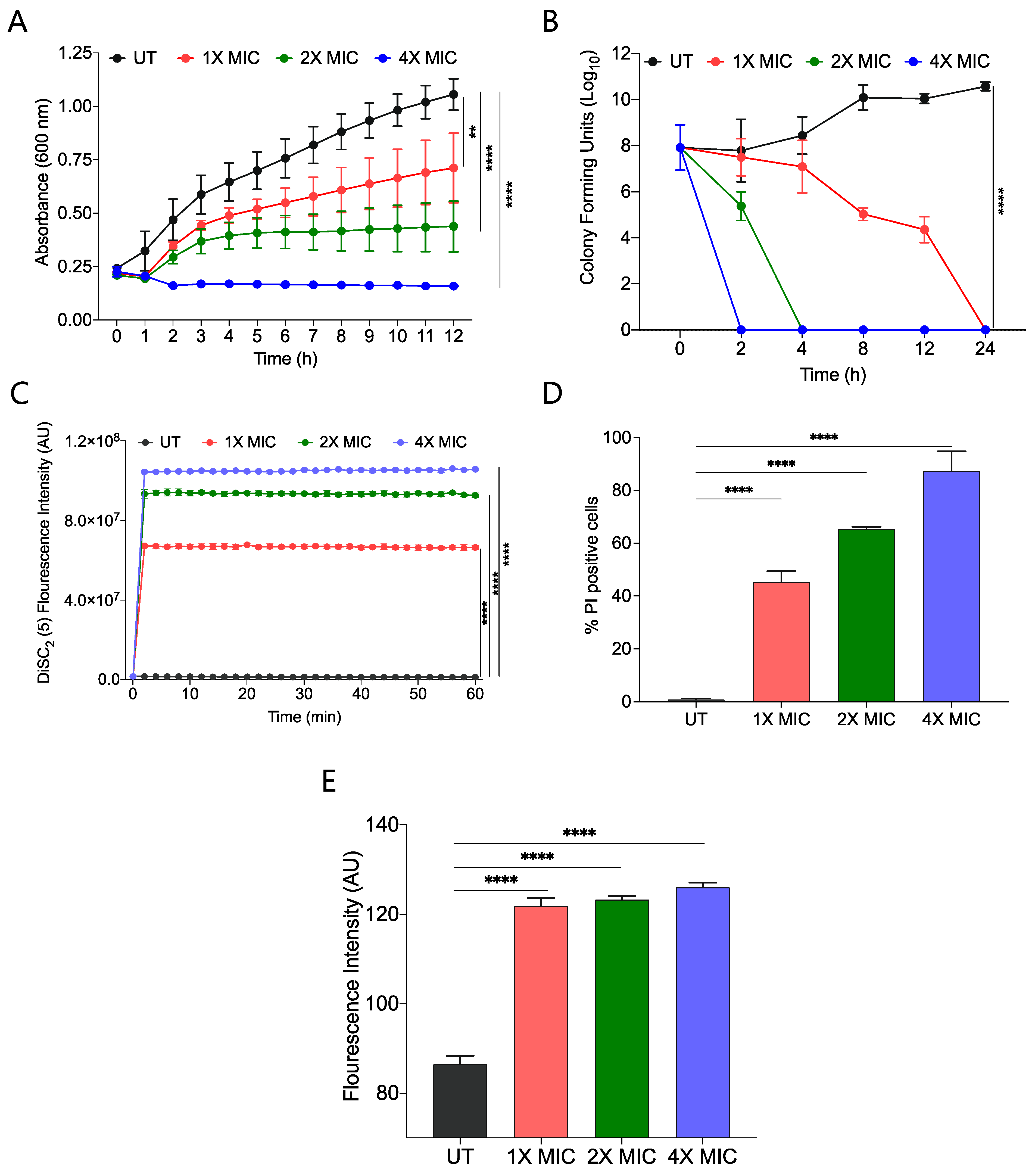
2.3. CABI-6 Amphiphile Is Bactericidal
2.4. CABI-6 Amphiphile Can Disrupt the Bacterial Membranes
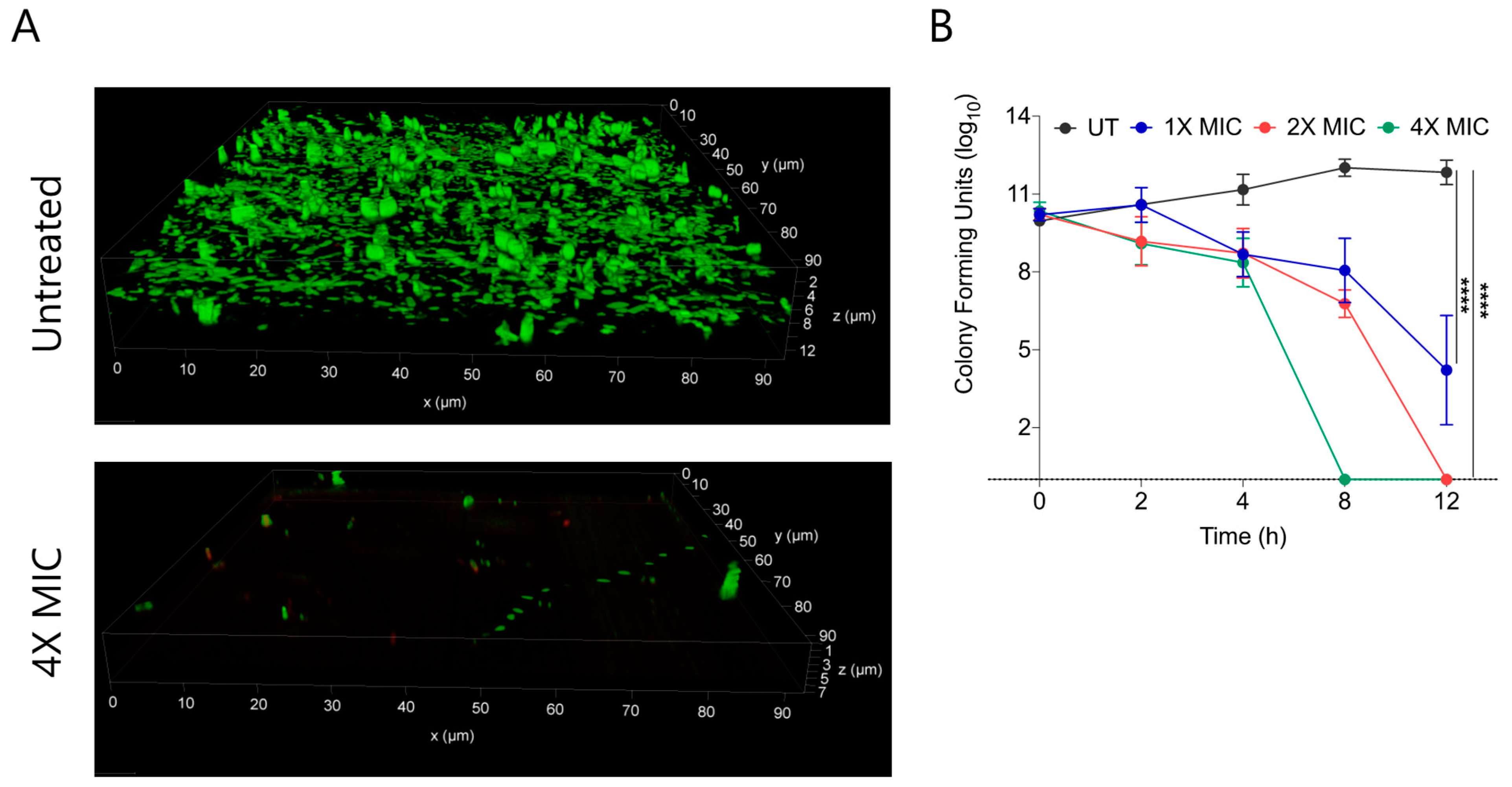
2.5. CABI-6 Can Eradicate the Preformed S. aureus Biofilms
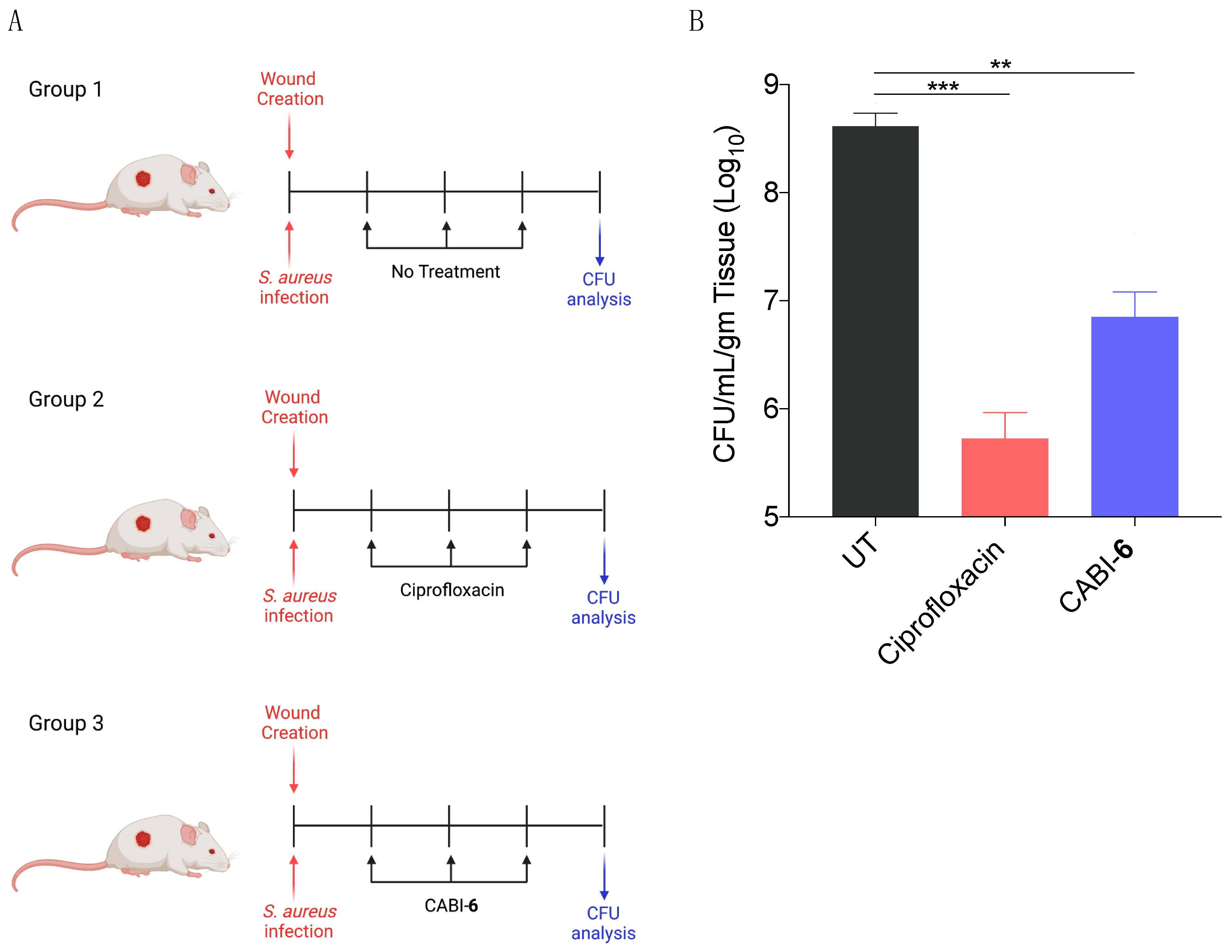
2.6. CABI-6 can Mitigate the S. aureus-Mediated Murine Wound Infection
3. Experimental Section
3.1. Materials
3.2. Synthesis of Cholic Acid Esters (10a–10h)
3.3. Synthesis of Chloroacetylated Cholic Acid Esters (11a–11h)
3.4. Synthesis of CABI Amphiphiles
3.5. Material and Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-Resistant Staphylococcus aureus: An Overview of Basic and Clinical Research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Coates, A.; Hu, Y.; Bax, R.; Page, C. The Future Challenges Facing the Development of New Antimicrobial Drugs. Nat. Rev. Drug Discov. 2002, 1, 895–910. [Google Scholar] [CrossRef] [PubMed]
- Toprak, E.; Veres, A.; Michel, J.B.; Chait, R.; Hartl, D.L.; Kishony, R. Evolutionary Paths to Antibiotic Resistance Under Dynamically Sustained Drug Selection. Nat. Genet. 2012, 44, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Idrees, M.; Sawant, S.; Karodia, N.; Rahman, A. Staphylococcus aureus Biofilm: Morphology, Genetics, Pathogenesis and Treatment Strategies. Int. J. Environ. Res. Public Health 2021, 18, 7602. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Shockman, G.D.; Barren, J.F. Structure, function, and assembly of cell walls of gram-positive bacteria. Annu. Rev. Microbiol. 1983, 37, 501–527. [Google Scholar] [CrossRef]
- Van der Es, D.; Hogendorf, W.F.; Overkleeft, H.S.; van der Marel, G.A.; Codée, J.D. Teichoic acids: Synthesis and applications. Chem. Soc. Rev. 2017, 46, 1464–1482. [Google Scholar] [CrossRef]
- Brown, S.; Santa Maria, J.P., Jr.; Walker, S. Wall teichoic acids of gram-positive bacteria. Annu. Rev. Microbiol. 2013, 67, 313–336. [Google Scholar] [CrossRef]
- Dias, C.; Rauter, A.P. Membrane-Targeting Antibiotics: Recent Developments Outside the Peptide Space. Future Med. Chem. 2019, 11, 211–228. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Sahl, H.G. Antimicrobial and Host-Defense Peptides as New Anti-Infective Therapeutic Strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Mehta, D.; Saini, V.; Aggarwal, B.; Khan, A.; Bajaj, A. Unlocking the bacterial membrane as a therapeutic target for next-generation antimicrobial amphiphiles. Mol. Asp. Med. 2021, 81, 100999. [Google Scholar] [CrossRef] [PubMed]
- Blanchet, M.; Brunel, J.M. Bile acid derivatives: From old molecules to a new potent therapeutic use: An overview. Curr. Med. Chem. 2018, 25, 3613–3636. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.Z.; Feng, Y.; Pollard, J.; Chin, J.N.; Rybak, M.J.; Bucki, R.; Epand, R.F.; Epand, R.M.; Savage, P.B. Ceragenins: Cholic acid-based mimics of antimicrobial peptides. Accou. Chem. Res. 2008, 41, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Wang, Y.; Le, M.; Chen, K.F.; Jia, Y.G. Recent progress in bile acid-based antimicrobials. Bioconjug. Chem. 2021, 32, 395–410. [Google Scholar] [CrossRef]
- Singh, M.; Bansal, S.; Kundu, S.; Bhargava, P.; Singh, A.; Motiani, R.K.; Shyam, R.; Sreekanth, V.; Sengupta, S.; Bajaj, A. Synthesis, structure–activity relationship, and mechanistic investigation of lithocholic acid amphiphiles for colon cancer therapy. Med. Chem. Comm. 2015, 6, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Mishra, D.; Khan, M.Z.; Saini, V.; Mehta, D.; Kumar, S.; Yadav, A.; Mitra, M.; Rani, P.; Singh, M.; et al. Development of a Highly Specific, Selective, and Sensitive Fluorescent Probe for Detection of Mycobacteria in Human Tissues. Adv. Healthc. Mater. 2022, 11, 2102640. [Google Scholar] [CrossRef]
- Li, C.H.; Peters, A.S.; Meredith, E.L.; Allman, G.W.; Savage, P.B. Design and synthesis of potent sensitizers of Gram-negative bacteria based on a cholic acid scaffolding. J. Am. Chem. Soc. 1998, 120, 2961–2962. [Google Scholar] [CrossRef]
- Li, C.; Budge, L.P.; Driscoll, C.D.; Willardson, B.M.; Allman, G.W.; Savage, P.B. Incremental conversion of outer-membrane permeabilizers into potent antibiotics for Gram-negative bacteria. J. Am. Chem. Soc. 1999, 121, 931–940. [Google Scholar] [CrossRef]
- Guan, Q.; Li, C.; Schmidt, E.J.; Boswell, J.S.; Walsh, J.P.; Allman, G.W.; Savage, P.B. Preparation and characterization of cholic acid-derived antimicrobial agents with controlled stabilities. Org. Lett. 2000, 2, 2837–2840. [Google Scholar] [CrossRef]
- Ding, B.; Yin, N.; Liu, Y.; Cardenas-Garcia, J.; Evanson, R.; Orsak, T.; Fan, M.; Turin, G.; Savage, P.B. Origins of cell selectivity of cationic steroid antibiotics. J. Am. Chem. Soc. 2004, 126, 13642–13648. [Google Scholar] [CrossRef]
- Rahman, M.A.; Bam, M.; Luat, E.; Jui, M.S.; Ganewatta, M.S.; Shokfai, T.; Nagarkatti, M.; Decho, A.W.; Tang, C. Macromolecular-clustered facial amphiphilic antimicrobials. Nat. Commun. 2018, 9, 5231. [Google Scholar] [CrossRef] [PubMed]
- Singla, P.; Dalal, P.; Kaur, M.; Arya, G.; Nimesh, S.; Singh, R.; Salunke, D.B. Bile Acid Oligomers and Their Combination with Antibiotics to Combat Bacterial Infections. J. Med. Chem. 2018, 61, 10265–10275. [Google Scholar] [CrossRef] [PubMed]
- Tahlan, S.; Kumar, S.; Kakkar, S.; Narasimhan, B. Benzimidazole scaffolds as promising antiproliferative agents: A review. BMC Chem. 2019, 13, 66. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Ma, S. Recent development of benzimidazole-containing antibacterial agents. ChemMedChem 2016, 11, 646–659. [Google Scholar] [CrossRef]
- Gaba, M.; Singh, S.; Mohan, C. Benzimidazole: An emerging scaffold for analgesic and anti-inflammatory agents. Eur. J. Med. Chem. 2014, 76, 494–505. [Google Scholar] [CrossRef]
- Zhang, L.; Addla, D.; Ponmani, J.; Wang, A.; Xie, D.; Wang, Y.N.; Zhang, S.L.; Geng, R.X.; Cai, G.X.; Li, S.; et al. Discovery of membrane active benzimidazole quinolones-based topoisomerase inhibitors as potential DNA-binding antimicrobial agents. Eur. J. Med. Chem. 2016, 111, 160–182. [Google Scholar] [CrossRef]
- Dokla, E.M.; Abutaleb, N.S.; Milik, S.N.; Li, D.; El-Baz, K.; Shalaby, M.A.W.; Al-Karaki, R.; Nasr, M.; Klein, C.D.; Abouzid, K.A.; et al. Development of benzimidazole-based derivatives as antimicrobial agents and their synergistic effect with colistin against gram-negative bacteria. Eur. J. Med. Chem. 2020, 186, 111850. [Google Scholar] [CrossRef]
- Yadav, K.; Yavvari, P.S.; Pal, S.; Kumar, S.; Mishra, D.; Gupta, S.; Mitra, M.; Soni, V.; Khare, N.; Sharma, P.; et al. Oral delivery of cholic acid-derived amphiphile helps in combating Salmonella-mediated gut infection and inflammation. Bioconjug. Chem. 2019, 30, 721–732. [Google Scholar] [CrossRef]
- Kumar, S.; Thakur, J.; Yadav, K.; Mitra, M.; Pal, S.; Ray, A.; Gupta, S.; Medatwal, N.; Gupta, R.; Mishra, D.; et al. Cholic Acid-Derived Amphiphile which Combats Gram-Positive Bacteria-Mediated Infections via Disintegration of Lipid Clusters. ACS Biomater. Sci. Eng. 2019, 5, 4764–4775. [Google Scholar] [CrossRef]
- Gupta, R.; Thakur, J.; Pal, S.; Mishra, D.; Rani, P.; Kumar, S.; Saini, A.; Singh, A.; Yadav, K.; Srivastava, A.; et al. Cholic-Acid-Derived amphiphiles can prevent and degrade fungal biofilms. ACS Appl. Bio Mater. 2020, 4, 7332–7341. [Google Scholar] [CrossRef]
- Pal, G.; Mehta, D.; Singh, S.; Magal, K.; Gupta, S.; Jha, G.; Bajaj, A.; Ramu, V.S. Foliar Application or Seed Priming of Cholic Acid-Glycine Conjugates can Mitigate/Prevent the Rice Bacterial Leaf Blight Disease via Activating Plant Defense Genes. Front. Plant Sci. 2021, 12, 2033. [Google Scholar] [CrossRef] [PubMed]
- Yadav, K.; Kumar, S.; Mishra, D.; Asad, M.; Mitra, M.; Yavvari, P.S.; Gupta, S.; Vedantham, M.; Ranga, P.; Komalla, V.; et al. Deciphering the Role of Intramolecular Networking in Cholic Acid–Peptide Conjugates on the Lipopolysaccharide Surface in Combating Gram-Negative Bacterial Infections. J. Med. Chem. 2019, 62, 1875–1886. [Google Scholar] [CrossRef] [PubMed]
- Mitra, M.; Asad, M.; Kumar, S.; Yadav, K.; Chaudhary, S.; Bhavesh, N.S.; Khalid, S.; Thukral, L.; Bajaj, A. Distinct intramolecular hydrogen bonding dictates antimicrobial action of membrane-targeting amphiphiles. J. Phys. Chem. Lett. 2019, 10, 754–760. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Thakur, J.; Pal, S.; Gupta, R.; Mishra, D.; Kumar, S.; Yadav, K.; Saini, A.; Yavvari, P.S.; Vedantham, M.; et al. Cholic acid-peptide conjugates as potent antimicrobials against interkingdom polymicrobial biofilms. Antimicrob. Agents Chemother. 2019, 63, e00520-19. [Google Scholar] [CrossRef]
- Hudson, M.A.; Siegele, D.A.; Lockless, S.W. Use of a fluorescence-based assay to measure Escherichia coli membrane potential changes in high throughput. Antimicrob. Agents Chemother. 2020, 64, e00910-20. [Google Scholar] [CrossRef]
- Davey, H.; Guyot, S. Estimation of microbial viability using flow cytometry. Curr. Protoc. Cytom. 2020, 93, e72. [Google Scholar]
- Su, H.L.; Chou, C.C.; Hung, D.J.; Lin, S.H.; Pao, I.C.; Lin, J.H.; Huang, F.L.; Dong, R.X.; Lin, J.J. The disruption of bacterial membrane integrity through ROS generation induced by nanohybrids of silver and clay. Biomaterials 2009, 30, 5979–5987. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Minimum Inhibitory Concentration (MIC99) (μg/mL) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Gram-Positive Strains | Gram-Negative Strains | |||||||
| S. aureus | S. pneumoniae | B. subtilis | S. oralis | E. Coli | P. aeruginosa | K. pneumoniae | S. typhimurium | |
| CABI-1 | >128 | >128 | >128 | 8 | >128 | >128 | >128 | >128 |
| CABI-2 | 128 | 128 | 128 | 4 | >128 | >128 | >128 | >128 |
| CABI-3 | 128 | 128 | 128 | 4 | >128 | 128 | >128 | >128 |
| CABI-4 | 32 | 32 | 64 | 2 | >128 | 64 | >128 | >128 |
| CABI-5 | 32 | 32 | 128 | 2 | >128 | 32 | >128 | >128 |
| CABI-6 | 16 | 32 | 128 | 2 | >128 | 32 | >128 | >128 |
| CABI-7 | 16 | 32 | 64 | 2 | >128 | 16 | >128 | 128 |
| CABI-8 | 32 | 32 | 32 | 2 | >128 | 32 | >128 | >128 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kakkar, H.; Chaudhary, N.; Mehta, D.; Saini, V.; Maheshwari, S.; Singh, J.; Walia, P.; Bajaj, A. N-methyl Benzimidazole Tethered Cholic Acid Amphiphiles Can Eradicate S. aureus-Mediated Biofilms and Wound Infections. Molecules 2022, 27, 3501. https://doi.org/10.3390/molecules27113501
Kakkar H, Chaudhary N, Mehta D, Saini V, Maheshwari S, Singh J, Walia P, Bajaj A. N-methyl Benzimidazole Tethered Cholic Acid Amphiphiles Can Eradicate S. aureus-Mediated Biofilms and Wound Infections. Molecules. 2022; 27(11):3501. https://doi.org/10.3390/molecules27113501
Chicago/Turabian StyleKakkar, Himanshu, Nalini Chaudhary, Devashish Mehta, Varsha Saini, Shallu Maheshwari, Jitender Singh, Preeti Walia, and Avinash Bajaj. 2022. "N-methyl Benzimidazole Tethered Cholic Acid Amphiphiles Can Eradicate S. aureus-Mediated Biofilms and Wound Infections" Molecules 27, no. 11: 3501. https://doi.org/10.3390/molecules27113501
APA StyleKakkar, H., Chaudhary, N., Mehta, D., Saini, V., Maheshwari, S., Singh, J., Walia, P., & Bajaj, A. (2022). N-methyl Benzimidazole Tethered Cholic Acid Amphiphiles Can Eradicate S. aureus-Mediated Biofilms and Wound Infections. Molecules, 27(11), 3501. https://doi.org/10.3390/molecules27113501