
Functional Expression, Purification and Identification of Interaction Partners of PACRG

Abstract
1. Introduction
2. Results
2.1. Recombinant Expression of PACRG
2.2. Optimizations of Expression Conditions of pCold-PACRG and pCold-GST-PACRG
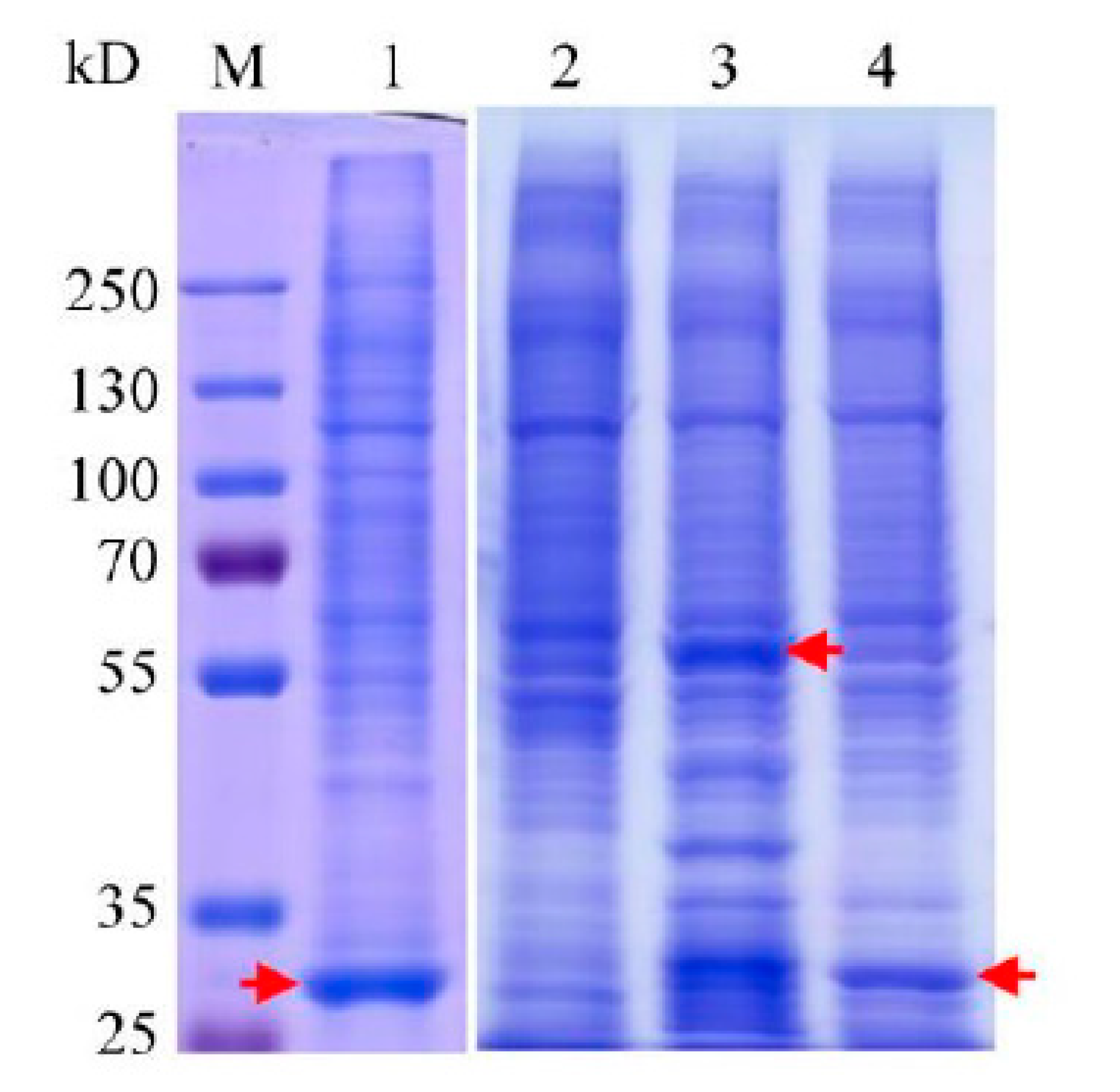
2.3. Purifications of the Recombinant Proteins
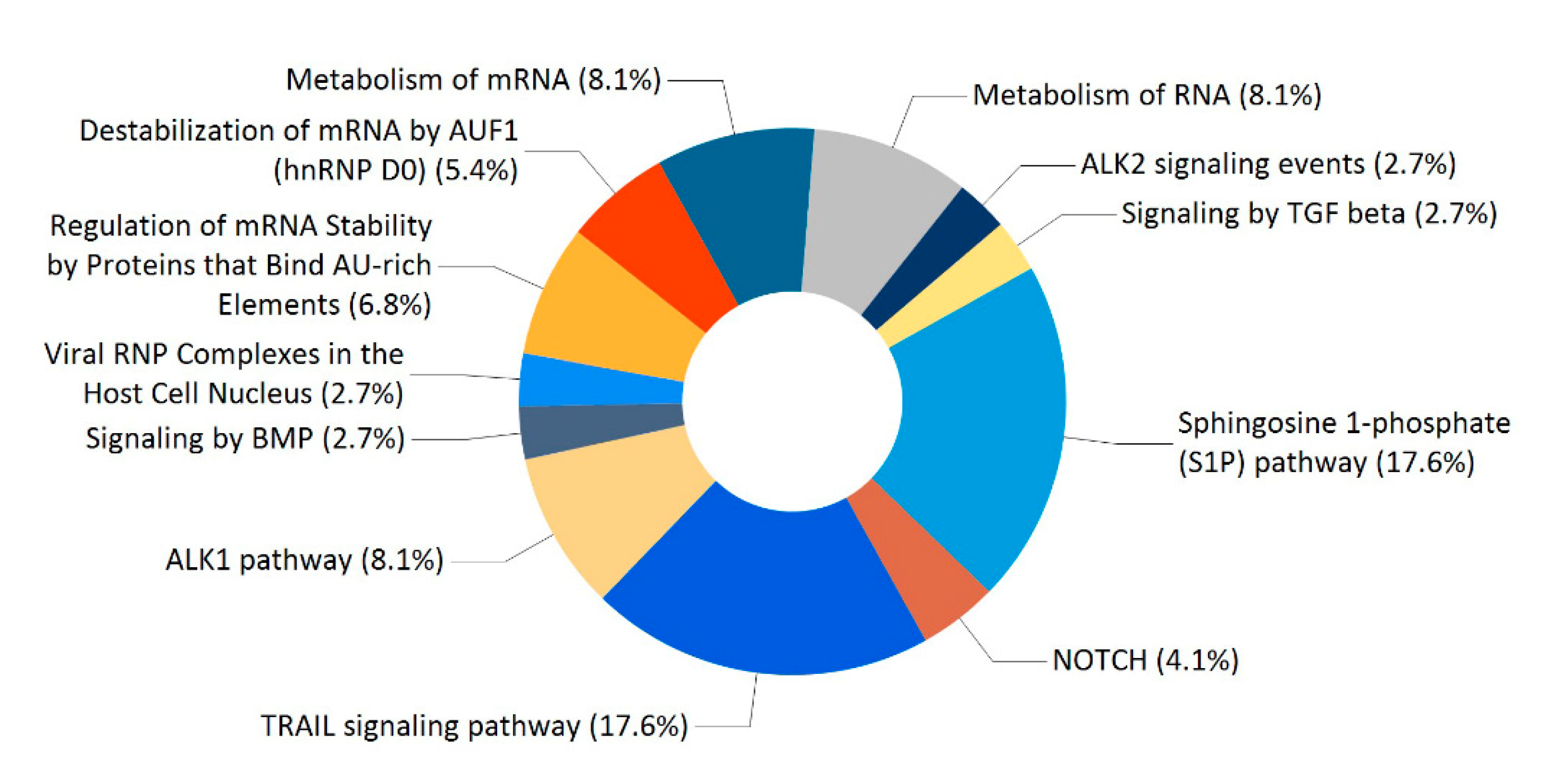
2.4. Enriched Analysis of the PACRG Interaction Partners
3. Discussion
4. Materials and Methods
4.1. Construction of Expression Plasmids pQE-PACRG, pCold-PACRG and pCold-GST-PACRG
4.2. Prokaryotic Expression of pQE-PACRG, pCold-PACRG and pCold-GST-PACRG
4.3. Purification of Recombinant Proteins
4.4. SDS-PAGE and Western Blot
4.5. His-Tag Pull Down Assay
4.6. LC-MS/MS Analysis
4.7. Cell Culture of HEK293T
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- West, A.B.; Lockhart, P.J.; Ofarell, C.; Farrer, M.J. Identification of a Novel Gene Linked to Parkin via a Bi-directional Promoter. J. Mol. Biol. 2003, 326, 11–19. [Google Scholar] [CrossRef]
- Kitada, T.; Asakawa, S.; Hattori, N.; Matsumine, H.; Yamamura, Y.; Minoshima, S.; Yokochi, M.; Mizuno, Y.; Shimizu, N. Mutations in the Parkin Gene Cause Autosomal Recessive Juvenile Parkinsonism. Nature 1998, 392, 605–608. [Google Scholar] [CrossRef]
- Ikeda, T. Parkin-co-regulated Gene (PACRG) Product Interacts with Tubulin and Microtubules. FEBS Lett. 2008, 582, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- Dawe, H.R.; Farr, H.; Portman, N.; Shaw, M.K.; Gull, K. The Parkin Co-regulated Gene Product, PACRG, is an Evolutionarily Conserved Axonemal Protein That Functions in Outer-doublet Microtubule Morphogenesis. J. Cell Sci. 2005, 118, 5421–5430. [Google Scholar] [CrossRef]
- Mizuno, K.; Dymek, E.E.; Smith, E.F. Microtubule Binding Protein PACRG Plays a Role in Regulating Specific ciliary Dyneins during Microtubule Sliding. Cytoskeleton 2016, 73, 703–711. [Google Scholar] [CrossRef]
- Lorenzetti, D.; Bishop, C.E.; Justice, M.J. Deletion of the Parkin Coregulated Gene Causes Male Sterility in the Quakingviable Mouse Mutant. Proc. Natl. Acad. Sci. USA 2004, 101, 8402–8407. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.Z.; Li, J.; Ma, Y.H.; Liu, W.H. Research Progress of Parkin Co-regulated Gene. Chin. Pharmacol. Bull. 2018, 34, 1333–1336. [Google Scholar]
- Imai, Y.; Soda, M.; Murakami, T.; Shoji, M.; Abe, K.; Takahashi, R. A Product of the Human Gene Adjacent to Parkin is a Component of Lewy Bodies and Suppresses Pael Receptor-induced Cell Death. J. Biol. Chem. 2003, 278, 51901–51910. [Google Scholar] [CrossRef]
- Taylor, J.M.; Song, Y.J.; Huang, Y.; Farrer, M.J.; Delatycki, M.B.; Halliday, G.M.; Lockhart, P.J. Parkin Co-regulated Gene (PACRG) is Regulated by the Ubiquitin–proteasomal System and is Present in the Pathological Features of Parkinsonian Diseases. Neurobio. Dis. 2007, 27, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Mira, M.T.; Alcaïs, A.; Nguyen, V.T.; Moraes, M.O.; Di Flumeri, C.; Vu, H.T.; Mai, C.P.; Nguyen, T.H.; Nguyen, N.B.; Pham, X.K.; et al. Susceptibility to Leprosy is Associated with PARK2 and PACRG. Nature 2004, 427, 636–640. [Google Scholar] [CrossRef]
- Bragina, E.Y.; Tiys, E.S.; Rudko, A.A.; Ivanisenko, V.A.; Freidin, M.B. Novel Tuberculosis Susceptibility Candidate Genes Revealed by the Reconstruction and Analysis of Associative Networks. Infect. Genet. Evol. 2016, 46, 118–123. [Google Scholar] [CrossRef]
- Ali, S.; Vollaard, A.M.; Widjaja, S.; Surjadi, C.; van de Vosse, E.; van Dissel, J.T. PARK2/PACRG Polymorphisms and Susceptibility to Typhoid and Paratyphoid Fever. Clin. Exp. Immunol. 2006, 144, 425–431. [Google Scholar] [CrossRef]
- Ichimura, K.; Mungall, A.J.; Fiegler, H.; Pearson, D.M.; Dunham, I.; Carter, N.P.; Peter Collins, V. Small Regions of Overlapping Deletions on 6q26 in Human Astrocytic Tumours Identified Using Chromosome 6 Tile Path Array-CGH. Oncogene 2006, 25, 1261–1271. [Google Scholar] [CrossRef]
- Meschede, J.; Šadić, M.; Furthmann, N.; Miedema, T.; Sehr, D.A.; Müller-Rischart, A.K.; Bader, V.; Berlemann, L.A.; Pilsl, A.; Schlierf, A.; et al. The Parkin-coregulated Gene Product PACRG Promotes TNF Signaling by Stabilizing LUBAC. Sci. Signal. 2020, 13, eaav1256. [Google Scholar] [CrossRef]
- Li, W.; Walavalkar, N.M.; Buchwald, W.A.; Teves, M.E.; Zhang, L.; Liu, H.; Bilinovich, S.; Peterson, D.L.; Strauss, J.F., 3rd; Williams, D.C., Jr.; et al. Dissecting the Structural Basis of MEIG1 Interaction with PACRG. Sci. Rep. 2016, 6, 18278. [Google Scholar] [CrossRef]
- Khan, N.; Pelletier, D.; McAlear, T.S.; Croteau, N.; Veyron, S.; Bayne, A.N.; Black, C.; Ichikawa, M.; Khalifa, A.A.Z.; Chaaban, S.; et al. Crystal Structure of Human PACRG in Complex with MEIG1 Reveals Roles in Axoneme Formation and Tubulin binding. Structure 2021, 29, 1–15. [Google Scholar]
- Qing, G.; Ma, L.C.; Khorchid, A.; Swapna, G.V.; Mal, T.K.; Takayama, M.M.; Xia, B.; Phadtare, S.; Ke, H.; Acton, T.; et al. Cold-shock Induced High-yield Protein Production in Escherichia coli. Nat. Biotechnol. 2004, 22, 877–882. [Google Scholar] [CrossRef]
- Sugiki, T.; Fujiwara, T.; Kojima, C. Cold-Shock Expression System in E. coli for Protein NMR Studies. Methods Mol. Biol. 2017, 1586, 345–357. [Google Scholar] [PubMed]
- Hayashi, K.; Kojima, C. pCold-GST Vector: A Novel Cold-shock Vector Containing GST Tag for Soluble Protein Production. Protein. Expr. Purif. 2008, 62, 120–127. [Google Scholar] [CrossRef]
- Lee, D.; Han, S.; Woo, S.; Lee, H.W.; Sun, H.; Kim, W. Enhanced Expression and Purification of Inositol 1,4,5-trisphosphate 3-kinase A through Use of the pCold1-GST Vector and a C-terminal Hexahistidine Tag in Escherichia coli. Protein. Expr. Purif. 2014, 97, 72–80. [Google Scholar] [CrossRef]
- Loucks, C.M.; Bialas, N.J.; Dekkers, M.P.; Walker, D.S.; Grundy, L.J.; Li, C.; Inglis, P.N.; Kida, K.; Schafer, W.R.; Blacque, O.E.; et al. PACRG, a Protein Linked to Ciliary Motility, Mediates Cellular Signaling. Mol. Biol. Cell. 2016, 27, 2133–2144. [Google Scholar] [CrossRef]
- Shires, S.E.; Quiles, J.M.; Najor, R.H.; Leon, L.J.; Cortez, M.Q.; Lampert, M.A.; Mark, A.; Gustafsson, Å.B. Nuclear Parkin Activates the ERRα Transcriptional Program and Drives Widespread Changes in Gene Expression Following Hypoxia. Sci. Rep. 2020, 10, 8499. [Google Scholar] [CrossRef] [PubMed]
- Nezich, C.L.; Wang, C.; Fogel, A.I.; Youle, R.J. MiT/TFE Transcription Factors are Activated during Mitophagy Downstream of Parkin and Atg5. J. Cell Biol. 2015, 210, 435–450. [Google Scholar] [CrossRef]
- Ren, Y.; Jiang, H.; Ma, D.; Nakaso, K.; Feng, J. Parkin Degrades Estrogen-related Receptors to Limit the Expression of Monoamine Oxidases. Hum. Mol. Genet. 2011, 20, 1074–1083. [Google Scholar] [CrossRef]
- Li, Y.Y.; Yu, H.; Guo, Z.M.; Guo, T.Q.; Tu, K.; Li, Y.X. Systematic Analysis of Head-to-head Gene Organization: Evolutionary Conservation and Potential Biological Relevance. PLoS Comput. Biol. 2006, 2, e74. [Google Scholar]
- Shimura, H.; Hattori, N.; Kubo, S.; Mizuno, Y.; Asakawa, S.; Minoshima, S.; Shimizu, N.; Iwai, K.; Chiba, T.; Tanaka, K.; et al. Familial Parkinson Disease Gene Product, Parkin, is a Ubiquitin-protein Ligase. Nat. Genet. 2000, 25, 302–305. [Google Scholar] [CrossRef]
- Tsai, H.C.; Han, M.H. Sphingosine-1-phosphate (S1P) and S1P Signaling Pathway: Therapeutic Targets in Autoimmunity and Inflammation. Drugs 2016, 11, 1067–1079. [Google Scholar] [CrossRef]
- Grbčić, P.; Sedić, M. Sphingosine 1-phosphate Signaling and Metabolism in Chemoprevention and Chemoresistance in Colon Cancer. Molecules 2020, 25, 2436. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Larrauri, A.; Presa, N.; Dominguez-Herrera, A.; Ouro, A.; Trueba, M.; Gomez-Muñoz, A. Role of Bioactive Sphingolipids in Physiology and Pathology. Essays Biochem. 2020, 64, 579–589. [Google Scholar] [PubMed]
- Woo, S.M.; Kwon, T.K. E3 Ubiquitin Ligases and Deubiquitinases as Modulators of TRAIL-mediated Extrinsic Apoptotic Signaling Pathway. BMB Rep. 2019, 52, 119–126. [Google Scholar] [CrossRef]
- Stöhr, D.; Jeltsch, A.; Rehm, M. TRAIL Receptor Signaling: From the Basics of Canonical Signal Transduction Toward Its Entanglement with ER Stress and the Unfolded Protein Response. Int. Rev. Cell Mol. Biol. 2020, 351, 57–99. [Google Scholar]
- Mazini, P.S.; Alves, H.V.; Reis, P.G.; Lopes, A.P.; Sell, A.M.; Santos-Rosa, M.; Visentainer, J.E.; Rodrigues-Santos, P. Gene Association with Leprosy: A Review of Published Data. Front. Immunol. 2016, 6, 658. [Google Scholar] [CrossRef]
- Leturiondo, A.L.; Noronha, A.B.; Mendonça, C.Y.R.; Ferreira, C.O.; Alvarado-Arnez, L.E.; Manta, F.S.N.; Bezerra, O.C.L.; Carvalho, E.F.; Moraes, M.O.; Rodrigues, F.D.C.; et al. Association of NOD2 and IFNG Single Nucleotide Polymorphisms with Leprosy in the Amazon Ethnic Admixed Population. PLoS Negl. Trop. Dis. 2020, 14, e0008247. [Google Scholar] [CrossRef]
- Agirre, X.; Román-Gómez, J.; Vázquez, I.; Jiménez-Velasco, A.; Garate, L.; Montiel-Duarte, C.; Artieda, P.; Cordeu, L.; Lahortiga, I.; Calasanz, M.J.; et al. Abnormal Methylation of the Common PARK2 and PACRG Promoter is Associated with Downregulation of Gene Expression in Acute Lymphoblastic Leukemia and Chronic Myeloid Leukemia. Int. J. Cancer 2006, 118, 1945–1953. [Google Scholar] [CrossRef]
- Toma, M.I.; Wuttig, D.; Kaiser, S.; Herr, A.; Weber, T.; Zastrow, S.; Koch, R.; Meinhardt, M.; Baretton, G.B.; Wirth, M.P.; et al. PARK2 and PACRG are Commonly Downregulated in Clear-cell Renal Cell Carcinoma and are Associated with Aggressive Disease and Poor Clinical Outcome. Genes Chromosomes Cancer 2013, 52, 265–273. [Google Scholar] [CrossRef]
- Han, B.; Yang, X.; Zhang, P.; Zhang, Y.; Tu, Y.; He, Z.; Li, Y.; Yuan, J.; Dong, Y.; Hosseini, D.K.; et al. DNA Methylation Biomarkers for Nasopharyngeal Carcinoma. PLoS ONE 2020, 15, e0230524. [Google Scholar] [CrossRef]
- Froes Jr, L.A.R.; Trindade, M.A.B.; Sotto, M.N. Immunology of Leprosy. Int. Rev. Immunol. 2020, 26, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Verheul, T.C.J.; van Hijfte, L.; Perenthaler, E.; Barakat, T.S. The Why of YY1: Mechanisms of Transcriptional Regulation by Yin Yang 1. Front. Cell Dev. Biol. 2020, 8, 592164. [Google Scholar] [CrossRef]
- Yao, Y.L.; Yang, W.M.; Seto, E. Regulation of Transcription Factor YY1 by Acetylation and Deacetylation. Mol. Cell. Biol. 2001, 21, 5979–5991. [Google Scholar] [CrossRef]
- Gordon, S.; Akopyan, G.; Garban, H.; Bonavida, B. Transcription Factor YY1: Structure, Function, and Therapeutic Implications in Cancer Biology. Oncogene 2006, 25, 1125–1142. [Google Scholar] [CrossRef]
- Khachigian, L.M. The Yin and Yang of YY1 in Tumor Growth and Suppression. Int. J. Cancer 2018, 143, 460–465. [Google Scholar] [CrossRef]
- Krajewska, J.; Arent, Z.; Zolkiewski, M.; Kędzierska-Mieszkowska, S. Isolation and Identification of Putative Protein Substrates of the AAA+ Molecular Chaperone ClpB from the Pathogenic Spirochaete Leptospira interrogans. Int. J. Mol. Sci. 2018, 19, 1234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.P.; Yan, H.; Yuan, Y.F.; Feng, Y.Q. Searching for Dysbindin-1 Interacting Proteins in Mouse Testis by GST Pull-down and Mass Spectrometry. Chin. J. Biochem. Mol. Biol. 2017, 33, 286–293. [Google Scholar]
- Pathan, M.; Keerthikumar, S.; Ang, C.S.; Gangoda, L.; Quek, C.Y.J.; Williamson, N.A.; Mouradov, D.; Sieber, O.M.; Simpson, R.J.; Salim, A.; et al. FunRich: An Open Access Standalone Functional Enrichment and Interaction Network Analysis Tool. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Molecular Function | Gene Names | No. of Genes | p-Value |
|---|---|---|---|---|
| 1 | Transcription factor activity | UBP1; SMAD4; IRF3; RSRC2; FOXC1; DCP1A; HEXIM1; YY1; BCLAF1; MNX1 | 10 | 0.0014 |
| 2 | Heat shock protein activity | HSPD1; HSPA8 | 2 | 0.0048 |
| 3 | Receptor signaling complex scaffold activity | STAM2; IRS4; SNTB2; PDLIM5; HOMER1 | 5 | 0.0080 |
| 4 | Chaperone activity | HSPA1A; HSPA5 | 2 | 0.013 |
| 5 | RNA binding | DHX15; IGF2BP2; CSTF2; RNPS1; SF1 | 5 | 0.013 |
| 6 | Transcription regulator activity | SMAD3; TRIP4; FUBP1; SNW1; MID1; ZNF24; POLR3C; SAP30BP | 8 | 0.014 |
| 7 | Intracellular transporter activity | ATG16L1 | 1 | 0.015 |
| 8 | Structural constituent of cytoskeleton | TUBB2B; TUBGCP4; CCDC6 | 3 | 0.015 |
| 9 | ATP binding | ABCD3 | 1 | 0.023 |
| 10 | GTP binding | SEPT9 | 1 | 0.030 |
| 11 | Exonuclease activity | EXD2 | 1 | 0.038 |
| 12 | Enzyme regulator activity | PPP2R1B | 1 | 0.049 |
| S. No. | Biological Process | Gene Names | No. of Genes | p-Value |
|---|---|---|---|---|
| 1 | Regulation of nucleotide metabolism | SMAD3; TRIP4; FUBP1; SNW1; MID1; ZNF24; POLR3C; UBP1; SMAD4; IRF3; FOXC1; DCP1A; HEXIM1; YY1; BCLAF1; MNX1; DHX15; IGF2BP2; CSTF2; RNPS1; SF1; EXD2; HIST2H2BF; ZNF787; ORC2; SETMAR | 26 | 9.86 × 10−6 |
| 2 | Nuclear organization and biogenesis | TMPO | 1 | 0.0039 |
| 3 | Cell proliferation | RSRC2; SEPT9 | 2 | 0.0099 |
| 4 | Neurotransmitter transport | STXBP1 | 1 | 0.015 |
| 5 | Amino acid and derivative metabolism | ALDH18A1 | 1 | 0.015 |
| 6 | Protein metabolism | HSPD1; HSPA8; HSPA1A; HSPA5; MID1; PSMD4; TRIM11; USP39; USP35 | 9 | 0.030 |
| 7 | Cell surface receptor linked signal transduction | SMAD1 | 1 | 0.030 |
| 8 | Regulation of immune response | HSPD1 | 1 | 0.030 |
| 9 | Regulation of translation | IGF2BP2 | 1 | 0.049 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, T.; Zhao, H.; Jian, S.; Gong, S.; Li, S.; Ma, Y.; Chen, J.; Liu, W. Functional Expression, Purification and Identification of Interaction Partners of PACRG. Molecules 2021, 26, 2308. https://doi.org/10.3390/molecules26082308
Liu T, Zhao H, Jian S, Gong S, Li S, Ma Y, Chen J, Liu W. Functional Expression, Purification and Identification of Interaction Partners of PACRG. Molecules. 2021; 26(8):2308. https://doi.org/10.3390/molecules26082308
Chicago/Turabian StyleLiu, Tiankai, Haizhou Zhao, Shaofen Jian, Shu Gong, Sainan Li, Yanhong Ma, Jun Chen, and Wenhua Liu. 2021. "Functional Expression, Purification and Identification of Interaction Partners of PACRG" Molecules 26, no. 8: 2308. https://doi.org/10.3390/molecules26082308
APA StyleLiu, T., Zhao, H., Jian, S., Gong, S., Li, S., Ma, Y., Chen, J., & Liu, W. (2021). Functional Expression, Purification and Identification of Interaction Partners of PACRG. Molecules, 26(8), 2308. https://doi.org/10.3390/molecules26082308