
Flavone-Rich Fractions and Extracts from Oroxylum indicum and Their Antibacterial Activities against Clinically Isolated Zoonotic Bacteria and Free Radical Scavenging Effects


Abstract
1. Introduction
2. Results
2.1. Determination of In Vitro Antioxidant Activity of O. indicum Samples Using DPPH Scavenging Method
2.2. Determination of Total Phenolic Content in O. indicum Samples Using the Folin-Ciocalteu Method and Determination of Total Flavonoid Content in O. indicum Samples Using the Aluminium Chloride Method
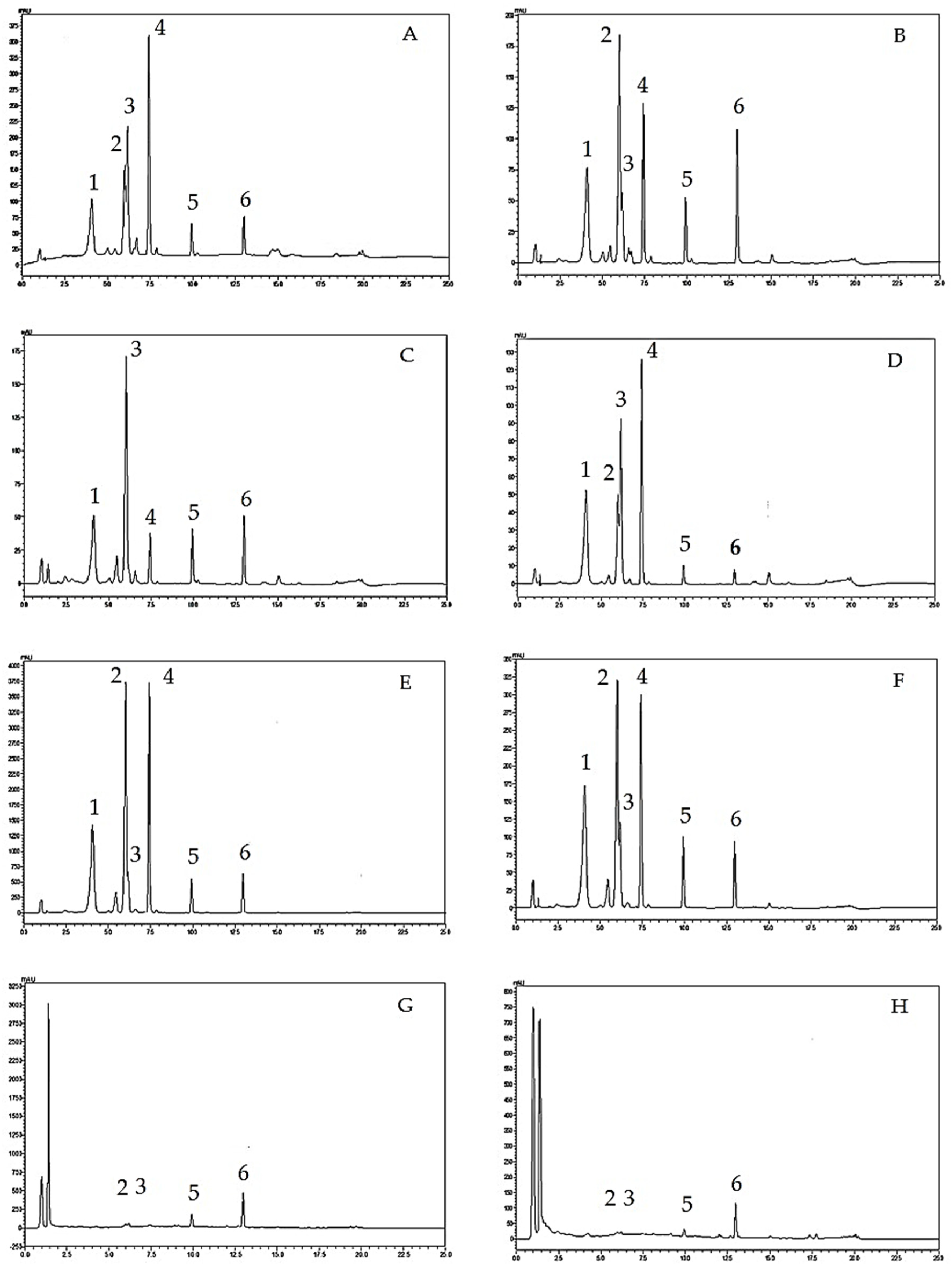
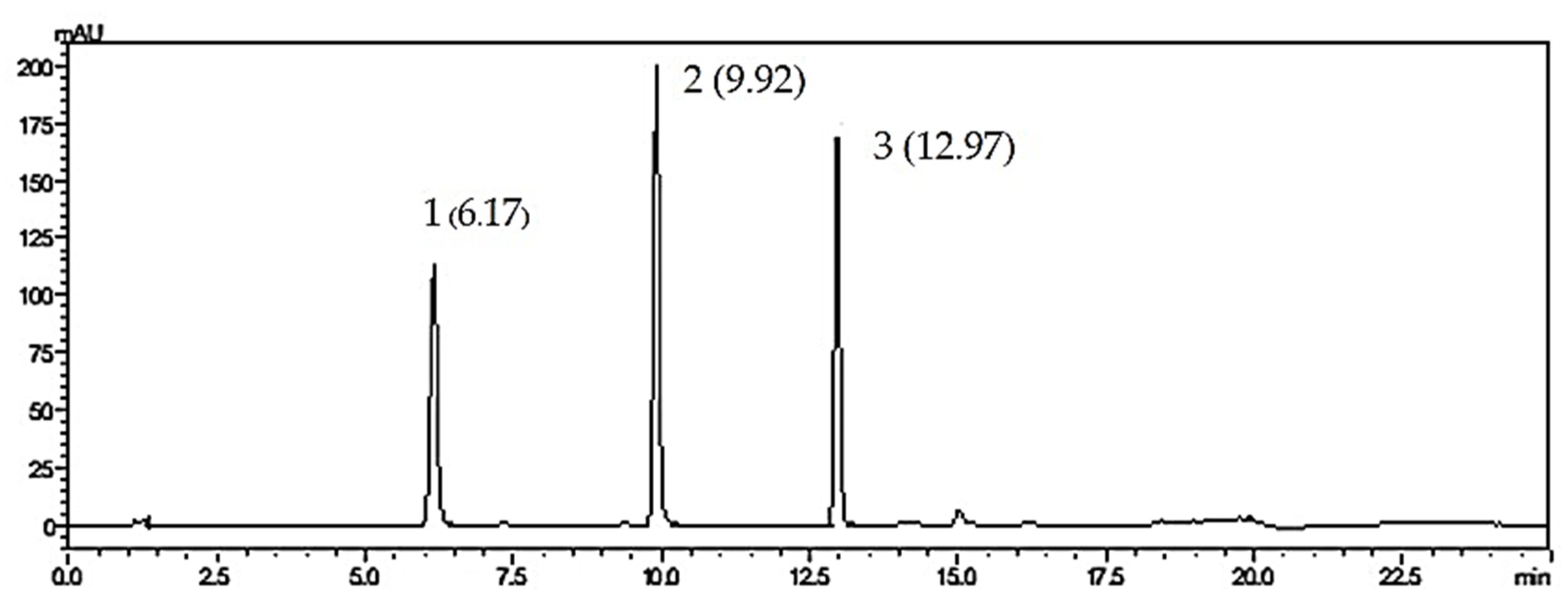
2.3. Analysis of Flavone Contents in O. indicum Samples by HPLC
2.4. Determination of In Vitro Antibacterial Activity of O. indicum Samples Using Disc Diffusion Assay
2.5. Determination of the Half-Maximal Inhibitory Concentrations (IC50) and the Minimum Inhibitory Concentrations (MICs)
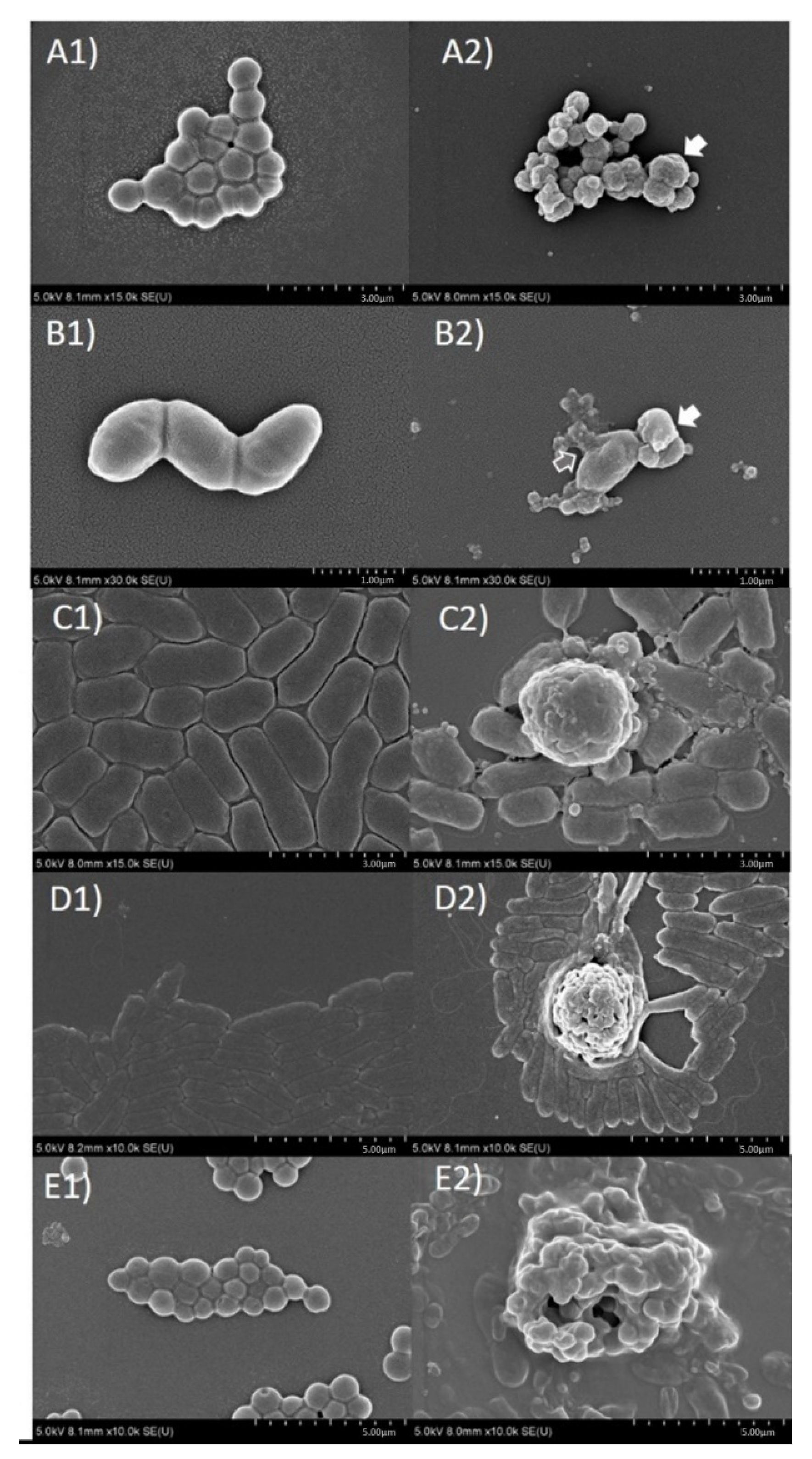
2.6. Evaluation of Antibacterial Mechanism of Action of O. indicum by Scanning Electron Microscopic Technique
2.7. Evaluation of Correlations between Antioxidant and Antibacterial Activities and Phytochemical Contents
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Materials
4.3. Plant Extract Preparations
4.4. Separation of Yellow Precipitate and Orange-Red Crystal from the Seed Extracts of O. indicum
4.5. Determination of In Vitro Antioxidant Activity of O. indicum Samples Using DPPH Scavenging Method
4.6. Determination of Total Phenolic Content in O. indicum Samples Using the Folin-Ciocalteu Method
4.7. Determination of Total Flavonoid Content in O. indicum Samples Using the Aluminium Chloride Method
4.8. Analysis of Flavone Contents in O. indicum Samples by HPLC
4.9. Clinical Isolated Bacterial, Bacterial Culture, and Culture Media
4.10. Determination of In Vitro Antibacterial Activity of O. indicum Samples Using Disc Diffusion Susceptibility Test
4.11. Determination of the Half-Maximal Inhibitory Concentrations (IC50) and the Minimum Inhibitory Concentrations (MICs)
4.12. Drop Plate Technique for Bacterial Enumeration (DP)
4.13. Evaluation of Antibacterial Mechanism of Action of O. indicum by Scanning Electron Microscopic Technique
4.14. Evaluation of Correlations between Antioxidant and Antibacterial Activities and Phytochemical Contents
4.15. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Harminder, S.V.; Chaudhary, A.K. A review on the taxonomy, ethnobotany, chemistry and pharmacology of Oroxylum indicum Vent. Indian J. Pharm. Sci. 2011, 73, 483–490. [Google Scholar] [PubMed]
- Warrier, P.K.; Nambiar, V.P.K.; Ramankutty, C. Indian Medicinal Plant: A Compendium of 500 Species (4); Orient Longman: Hyderabad, India, 1993; pp. 186–190. [Google Scholar]
- Subramaniam, S.S.; Nair, A.G. Flavonoids of the stem bark of Oroxylum indicum. Curr. Sci. 1972, 41, 62–63. [Google Scholar]
- Jayaram, K.; Prasad, M.N. Genetic diversity in Oroxylum indicum (L.) Vent. (Bignoniaceae), a vulnerable medicinal plant by random amplified polymorphic DNA marker. Afr. J. Biotechnol. 2008, 7, 254–262. [Google Scholar]
- Bhattacharje, A.K.; Das, A.K. Phytochemical screening of some Indian plants. J. Crude Drug Res. 1969, 9, 1408–1412. [Google Scholar] [CrossRef]
- Chen, L.J.; Games, D.E.; Jones, J. Isolation and identification of four flavonoid constituents from the seeds of Oroxylum indicum by high-speed counter-current chromatography. J. Chromatogr. A. 2003, 988, 95–105. [Google Scholar] [CrossRef]
- Chen, L.J.; Song, H.; Lan, X.Q.; Games, D.E.; Sutherland, I.A. Comparison of high-speed counter-current chromatography instruments for the separation of the extracts of the seeds of Oroxylum indicum. J. Chromatogr. A 2005, 1063, 241–245. [Google Scholar] [CrossRef]
- Zaveri, M.; Khandhar, A.; Jain, S. Quantification of baicalein, chrysin, biochanin-A and ellagic acid in root bark of Oroxylum indicum by RP-HPLC with UV detection. Eurasian J. Anal. Chem. 2008, 3, 245–257. [Google Scholar]
- Tomimori, T.; Imoto, Y.; Ishida, M.; Kizu, H.; Namba, T. Studies on the Nepalese crude drug. VIII. on the flavonoid constituents of the seed of Oroxylum indicum. Shoyakugaku Zasshi. 1988, 42, 98–101. [Google Scholar]
- Yan, R.Y.; Cao, Y.Y.; Chen, C.Y.; Dai, H.Q.; Yu, S.X.; Wei, J.L.; Li, H.; Yang, B. Antioxidant flavonoids from the seed of Oroxylum indicum. Fitoterapia 2011, 82, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Krüger, A.; Ganzera, M. Oroxylum indicum seeds—Analysis of flavonoids by HPLC-MS. J. Pharm. Biomed. Anal. 2012, 70, 553–556. [Google Scholar] [CrossRef]
- Sithisarn, P.; Rojsanga, P.; Sithisarn, P. Inhibitory effects on clinical isolated bacteria and simultaneous HPLC quantitative analysis of flavone contents in extracts from Oroxylum indicum. Molecules 2019, 24, 1937. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Shang, X.; Zhu, C.; Qin, S.; Zhou, Y.; Liao, Q.; Zhang, R.; Zhao, Z.; Zhang, L. Qualitative and quantitative evaluation of Oroxylum indicum (L.) Kurz by HPLC and LC/qTOF/MS/MS. Biomed. Chromatogr. 2019, 33, e4657. [Google Scholar] [CrossRef] [PubMed]
- Praserdmek, K.; Palanuvej, C.; Ruangrungsi, N. Pharmacognostic specification and chrysin content of Oroxylum indicum seeds in Thailand. J. Sci. Tech. UBU 2017, 60–68. [Google Scholar]
- Roy, M.K.; Nakahara, K.; Na, T.V.; Trakoontivakorn, G.; Takenaka, M.; Isobe, S.; Tsushida, T. Baicalein, a flavonoid extracted from a methanolic extract of Oroxylum indicum inhibits proliferation of a cancer cell line in vitro via induction of apoptosis. Pharmazie 2007, 62, 149–153. [Google Scholar]
- Subramanian, S.S.; Nair, A.G.R. Flavanoids of the leaves of Oroxylum indicum and Pajanelia longifolia. Phytochemistry 1972, 11, 439–440. [Google Scholar] [CrossRef]
- Dey, A.K.; Mukherjee, A.; Das, P.C.; Chatterjee, A. Occurrence of aloe-emodin in the leaves of Oroxylum indicum Vent. Indian J. Chem. 1978, 16, 1042. [Google Scholar]
- Yuan, Y.; Hou, W.; Tang, M.; Luo, H.; Chen, L.J.; Guan, Y.H.; Sutherland, I.A. Separation of flavonoids from the leaves of Oroxylum indicum by HSCCC. Chromatographia 2008, 68, 885–892. [Google Scholar] [CrossRef]
- Buranrat, B.; Noiwetch, S.; Suksar, T.; Ta-ut, A. Inhibition of cell proliferation and migration by Oroxylum indicum extracts on breast cancer cells via Rac1 modulation. J. Pharm. Anal. 2020, 10, 187–193. [Google Scholar] [CrossRef]
- Dunkhunthod, B.; Talabnin, C.; Murphy, M.; Thumanu, K.; Sittisart, P.; Hengpratom, T.; Eumkeb, G. Intracellular ROS scavenging and anti-inflammatory activities of Oroxylum indicum Kurz (L.) extract in LPS plus IFN-γ-Activated RAW264.7 macrophages. Evid. Based Complement. Altern. Med. 2020, 2020, 7436920. [Google Scholar] [CrossRef]
- Rai, B.; Bhutia, S.; Pal, P.; Kakoti, B.B. Phytochemical analysis and antibacterial evaluation against selected gram strains by Oroxylum indicum (L.) Kurz stem bark extract, a folklore medicine of Sikkim Himalaya. J. Pharmacogn. Phytochem. 2020, 9, 11–16. [Google Scholar]
- Kim, G.; Gan, R.Y.; Zhang, D.; Farha, A.K.; Habimana, O.; Mavumengwana, V.; Li, H.B.; Wang, X.H.; Corke, H. Large-scale screening of 239 traditional Chinese medicinal plant extracts for their antibacterial activities against multidrug-resistant Staphylococcus aureus and cytotoxic activities. Pathogens 2020, 9, 185. [Google Scholar] [CrossRef]
- Sithisarn, P.; Nantateerapong, P.; Rojsanga, P.; Sithisarn, P. Screening for antibacterial and antioxidant activities and phytochemical analysis of Oroxylum indicum fruit extracts. Molecules 2016, 21, 446. [Google Scholar] [CrossRef]
- Rojsanga, P.; Bunsupa, S.; Brantner, A.H.; Sithisarn, P. Comparative phytochemical profiling and in vitro antioxidant activity of extracts from raw materials, tissue-cultured plants, and callus of Oroxylum indicum (L.) Vent. Evid. Based Complement. Alternat. Med. 2017, 2017, 6853212. [Google Scholar] [CrossRef] [PubMed]
- Rojsanga, P.; Bunsupa, S.; Sithisarn, P. Flavones contents in extracts from Oroxylum indicum seeds and plant tissue cultures. Molecules 2020, 25, 1545. [Google Scholar] [CrossRef]
- Lin, L.Z.; Harnly, J.M. A screening method for the identification of glycosylated flavonoids and other phenolic compounds using a standard analytical approach for all plant materials. J. Agric. Food Chem. 2007, 2007. 21, 1084–1096. [Google Scholar] [CrossRef]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef]
- Chen, H.; He, G.; Li, C.; Dong, L.; Xie, X.; Wu, J.; Gao, Y.; Zhou, J. Development of a concise synthetic approach to access oroxin A. RSC Adv. 2014, 4, 45151–45154. [Google Scholar] [CrossRef]
- Sun, W.; Zhang, B.; Yu, X.; Zhuang, C. Oroxin A from Oroxylum indicum prevents the progression from prediabetes to diabetes in streptozotocin and high-fat diet induced mice. Phytomedicine 2017, 38, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Fu, S.; Cao, Z.; Liao, H.; Huo, Z.; Pan, Y.; Zhang, G.; Gao, A.; Zhou, Q. Oroxin B selectively induces tumor-suppressive ER stress and concurrently inhibits tumor-adaptive ER stress in B-lymphoma cells for effective anti-lymphoma therapy. Toxicol. Appl. Pharmacol. 2015, 288, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.Z.; Wang, D.C.; Zhang, Y.; Dong, J.; Wang, J.F.; Niu, X.D. Molecular modeling reveals the novel inhibition mechanism and binding mode of three natural compounds to Staphylococcal α-hemolysin. PLoS ONE 2013, 8, e80197. [Google Scholar] [CrossRef] [PubMed]
- Li, N.N.; Meng, X.S.; Bao, Y.R.; Wang, S.; Li, T.J. Evidence for the involvement of COX-2/VEGF and PTEN/Pl3K/AKT pathway the mechanism of oroxin B treated liver cancer. Pharmacogn. Mag. 2018, 14, 207–213. [Google Scholar] [PubMed]
- Li, N.; Men, W.; Zheng, Y.; Wang, H.; Meng, X. Oroxin B induces apoptosis by down-regulating microRNA-221 resulting in the inactivation of the PTEN/PI3K/AKT pathway in liver cancer. Molecules 2019, 24, 4384. [Google Scholar] [CrossRef]
- Li, K.; Yao, F.; Xue, Q.; Fan, H.; Yang, L.; Li, X. Inhibitory effects against α-glucosidase and α-amylase of the flavonoids-rich extract from Scutellaria baicalensis shoots and interpretation of structure–activity relationship of its eight flavonoids by a refined assign-score method. Chem. Cent. J. 2018, 12, 82. [Google Scholar] [CrossRef]
- Shan, N.; Toda, F.; Jones, W. Mechanochemistry and co-crystal formation: Effect of solvent on reaction kinetics. Chem. Comm. 2002, 20, 2372–2373. [Google Scholar] [CrossRef] [PubMed]
- Khandavilli, U.B.R.; Skořepová, E.; Sinha, A.S.; Bhogala, B.R.; Maguire, N.M.; Maguire, A.R.; Lawrence, S.E. Cocrystals and a salt of the bioactive flavonoid: Naringenin. Cryst. Growth Des. 2018, 18, 4571–4577. [Google Scholar] [CrossRef]
- Smith, A.J.; Kavuru, P.; Wojtas, L.; Zaworotko, M.J.; Shytle, R.D. Cocrystals of quercetin with improved solubility and oral Bioavailability. Mol. Pharm. 2011, 8, 1867–1876. [Google Scholar] [CrossRef]
- Vasisht, K.; Chadha, K.; Karan, M.; Bhalla, Y.; Jena, A.K.; Chadha, R. Enhancing biopharmaceutical parameters of bioflavonoid quercetin by cocrystallization. Cryst. Eng. Comm. 2016, 18, 1403–1415. [Google Scholar] [CrossRef]
- Zaini, E.; Azhari, D.; Fitriani, L. Identification and characterization of solid binary system of quercetin-nicotinamide. Orient. J. Chem. 2016, 32, 1545–1550. [Google Scholar] [CrossRef]
- De Grano, R.V.R.; Vashchenko, E.V.; Nisar, M.; Sung, H.H.Y.; Vashchenko, V.V.; Williams, I.D. Crystal structures of the flavonoid oroxylin A and the regioisomers negletein and wogonin. Acta Cryst. 2020, 76, 490–499. [Google Scholar] [CrossRef]
- Zhu, N.; Li, J.C.; Zhu, J.X.; Wang, X.; Zhang, J. Characterization and bioavailability of wogonin by different administration routes in beagles. Med. Sci. Monit. 2016, 22, 373–3745. [Google Scholar] [CrossRef]
- Smith, G.; Bartley, J.P.; Wang, E.; Bott, R.C. Two crystal polymorphs of a flavonoid from Melicope ellyrana. Acta Cryst. 2001, 57, 1336–1337. [Google Scholar]
- Tsuchiya, H. Membrane interactions of phytochemicals as their molecular mechanism applicable to the discovery of drug leads from plants. Molecules 2015, 20, 18923–18966. [Google Scholar] [CrossRef]
- Arora, A.; Byrem, T.M.; Nair, M.G.; Strasburg, G.M. Modulation of liposomal membrane fluidity by flavonoids and isoflavonoids. Arch. Biochem. Biophys. 2000, 373, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Ollila, F.; Halling, K.; Vuorela, P.; Vuorela, H.; Slotte, J.P. Characterization of flavonoid—Biomembrane interactions. Arch. Biochem. Biophys. 2002, 399, 103–108. [Google Scholar] [CrossRef]
- Oteiza, P.I.; Erlejman, A.G.; Verstraeten, S.V.; Keen, C.L.; Fraga, C.G. Flavonoid-membrane Interactions: A protective role of flavonoids at the membrane surface? Clin. Dev. Immunol. 2005, 12, 19–25. [Google Scholar] [CrossRef]
- Chabot, S.; Bel-Rhlid, R.; Chenevert, R.; Piche, Y. Hyphal growth promotion in vitro of the VA mycorrhizal fungus, Gigaspora margarita Becker and Hall, by the activity of structurally specific flavonoid compounds under CO2-enriched conditions. New Phytol. 1992, 122, 461–467. [Google Scholar] [CrossRef]
- Matijasˇevic´, D.; Pantic´, M.; Rasˇkovic´, B.; Pavlovic´, V.; Duvnjak, D.; Sknepnek, A.; Nikšić, M. The antibacterial activity of coriolus versicolor methanol extract and its effect on ultrastructural changes of Staphylococcus aureus and Salmonella enteritidis. Front. Microbiol. 2016, 7, 1–15. [Google Scholar]
- Mishra, A.K.; Mishra, A.; Kehri, H.; Sharma, B.; Pandey, A.K. Inhibitory activity of Indian spice plant Cinnamomum zeylanicum extracts against Alternaria solani and Curvularia lunata, the pathogenic dematiaceous moulds. Ann. Clin. Microbiol. Antimicrob. 2009, 8, 9. [Google Scholar] [CrossRef]
- Sato, M.; Fujiwara, S.; Tsuchiya, H.; Fujii, T.; Iinuma, M.; Tosa, H.; Ohkawa, Y. Flavones with antibacterial activity against cariogenic bacteria. J. Ethnopharmacol. 1996, 54, 171–176. [Google Scholar] [CrossRef]
- Lee, J.H.; Regmi, S.C.; Kim, J.A.; Cho, M.H.; Yun, H.; Lee, C.S.; Lee, J. Apple flavonoid phloretin inhibits Escherichia coli O157:H7 biofilm formation and ameliorates colon inflammation in rats. Infect. Immun. 2011, 79, 4819–4827. [Google Scholar] [CrossRef] [PubMed]
- Elmasri, W.A.; Zhu, R.; Peng, W.; Al-Hariri, M.; Kobeissy, F.; Tran, P.; Hamood, A.N.; Hegazy, M.F.; Pare, P.W.; Mechref, Y. Multitargeted flavonoid inhibition of the pathogenic bacterium Staphylococcus aureus: A proteomic characterization. J. Proteome Res. 2017, 16, 2579–2586. [Google Scholar] [CrossRef]
- Fujita, M.; Shiota, S.; Kuroda, T.; Hatano, T.; Yoshida, T.; Mizushima, T.; Tsuchiya, T. Remarkable synergies between baicalein and tetracycline, and baicalein and β-lactams against methicillin-resistant Staphylococcus aureus. Microbiol. Immunol. 2005, 49, 391–396. [Google Scholar] [CrossRef]
- Wu, T.; Zang, X.; He, M.; Pan, S.; Xu, X. Structure-activity relationship of flavonoids on their anti-Escherichia coli activity and inhibition of DNA gyrase. J. Agric. Food Chem. 2013, 61, 8185–8190. [Google Scholar] [CrossRef]
- Fang, Y.; Lu, Y.; Zang, X.; Wu, T.; Qi, X.; Pan, S.; Xu, X. 3DQSAR and docking studies of flavonoids as potent Escherichia coli inhibitors. Sci. Rep. 2016, 6, 23634. [Google Scholar] [CrossRef]
- Plaper, A.; Golob, M.; Hafner, I.; Oblak, M.; Solmajer, T.; Jerala, R. Characterization of quercetin binding site on DNA gyrase. Biochem. Biophys. Res. Commun. 2003, 306, 530–536. [Google Scholar] [CrossRef]
- Xu, H.; Ziegelin, G.; Schroder, W.; Frank, J.; Ayora, S.; Alonso, J.C.; Lanka, E.; Saenger, W. Flavones inhibit the hexameric replicative helicase RepA. Nucleic Acids Res. 2001, 29, 5058–5066. [Google Scholar] [CrossRef]
- Chinnam, N.; Dadi, P.K.; Sabri, S.A.; Ahmad, M.; Kabir, M.A.; Ahmad, Z. Dietary bioflavonoids inhibit Escherichia coli ATP synthase in a differential manner. Int. J. Biol. Macromol. 2010, 46, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Abreu, A.C.; McBain, A.J.; Simoes, M. Plants as sources of new antimicrobials and resistance-modifying agents. Nat. Prod. Rep. 2012, 29, 1007–1021. [Google Scholar] [CrossRef] [PubMed]
- Qian, M.; Tang, S.; Wu, C.; Wang, Y.; He, T.; Chen, T.; Xiao, X. Synergy between baicalein and penicillins against penicillinase-producing Staphylococcus aureus. Int. J. Med. Microbiol. 2015, 305, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Eumkeb, G.; Sakdarat, S.; Siriwong, S. Reversing β-lactam antibiotic resistance of Staphylococcus aureus with galangin from Alpinia officinarum Hance and synergism with ceftazidime. Phytomedicine 2010, 18, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.D.; Gibbons, S.; Bucar, F. Plant phenolic compounds as ethidium bromide efflux inhibitors in mycobacterium smegmatis. J. Antimicrob. Chemother. 2008, 62, 345–348. [Google Scholar]
- Cao, Y.Y.; Dai, B.D.; Sun, X.R.; Zhu, Z.Y.; Cao, Y.B.; Wang, Y.; Gao, P.H.; Jiang, Y.Y. In vitro synergism of fluconazole and baicalein against clinical isolates of Candida albicans resistant to fluconazole. Biol. Pharm. Bull. 2008, 31, 2234–2236. [Google Scholar]
- Chan, B.C.L.; Ip, M.; Lau, C.B.S.; Lui, S.L.; Jolivalt, C.; Ganem-Elbaz, C.; Litaudon, M.; Reiner, N.E.; Gong, H.; See, R.H.; et al. Synergistic effects of baicalein with ciprofloxacin against NorA over-expressed methicillin-resistant Staphylococcus aureus (MRSA) and inhibition of MRSA pyruvate kinase. J. Ethnopharmacol. 2011, 137, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Dzoyem, J.P.; Hamamoto, H.; Ngameni, B.; Ngadjui, B.T.; Sekimizu, K. Antimicrobial action mechanism of flavonoids from Dorstenia species. Drug Discov. Ther. 2013, 7, 66–72. [Google Scholar] [PubMed]
- Xin, X.; Zhang, M.; Li, X.F.; Zhao, G. Biocatalytic synthesis of lipophilic baicalin derivatives as antimicrobial agents. J. Agric. Food Chem. 2019, 67, 11684–11693. [Google Scholar] [CrossRef]
- Baron, E.J.; Pererson, L.R.; Finegold, S.M. Bailey and Scott’s Diagnostic Microbiology, 9th ed.; Baron, E.J., Pererson, L.R., Finegold, S.M., Eds.; CVMosby: St. Louis, MO, USA, 1997. [Google Scholar]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 36, 493–496. [Google Scholar] [CrossRef]
- Herigstad, B.; Hamilton, M.; Heersink, J. How to optimize the drop plate method for enumerating bacteria. J. Microbiol. Methods 2011, 44, 121–129. [Google Scholar] [CrossRef]
- Koch, A.L. Growth measurement. In Methods for General and Molecular Bacteriology; Gerhardt, P., Murray, R.G.E., Wood, W.A., Eds.; American Society for Microbiology: Colombia, WT, USA, 1994. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sample | DPPH Assay (EC50, μg/mL) | TPC (g% GAE) | TFC (g% QE) | Content (g% in the Extract) * | |||
|---|---|---|---|---|---|---|---|
| Baicalin + B0 | Baicalin | Baicalein | Chrysin | ||||
| OIRL | 25.99 ± 3.30 a | 8.20 ± 0.31 a | 6.01 ± 0.41 a | 12.12 ± 0.02 a | 6.82 ± 0.04 a | 1.12 ± 0.01 a | 1.49 ± 0.01 a |
| OIYL | 54.57 ± 5.76 b | 6.94 ± 0.34 b | 6.56 ± 0.49 b | 7.93 ± 0.06 b | 1.62 ± 0.04 b | 1.06 ± 0.01b | 2.34 ± 0.03 b |
| OISL | 122.63 ± 4.88 c | 5.95 ± 0.76 b,c,d | 4.04 ± 0.72 c,d | 6.87 ± 0.11 c | 6.87 ± 0.11 a | 0.98 ± 0.00 c | 1.31 ± 0.01 c |
| OIRP | 70.52 ± 0.80 d | 1.94 ± 0.00 e | 4.78 ± 0.18 c | 4.70 ± 0.00 d | 2.97 ± 0.03 c | 0.32 ± 0.01 d | 0.25 ± 0.00 d |
| OIYP | 29.16 ± 0.07 a | 7.88 ± 0.17 a | 8.74 ± 0.19 e | 15.29 ± 0.02 e | 1.83 ± 0.04 d | 1.06 ± 0.01 b | 1.48 ± 0.00 a |
| OISP | 47.87 ± 1.10 b | 4.53 ± 0.31 c | 3.98 ± 0.21 d | 16.24 ± 0.05 f | 3.66 ± 0.01 e | 2.10 ± 0.01 e | 2.24 ± 0.01 b |
| OIYF | 164.41 ± 9.87 e | 4.68 ± 0.37 c, d | 5.16 ± 0.16 c | 0.40 ± 0.00 g | 0.19 ± 0.00 f | 0.36 ± 0.00 d | 1.12 ± 0.00 e |
| OIFL | 180.24 ± 2.20 e | 3.78 ± 0.29 d | 0.58 ± 0.14 f | 0.09 ± 0.00 h | 0.04 ± 0.00 g | 0.06 ± 0.00 f | 0.26 ± 0.00 d |
| Ascorbic acid | 4.04 ± 0.20 | - | - | - | - | - | - |
| Peak No. | Rt (min.) | UV Spectra (λmax, nm) | Previous Reported MS Data # (m/z) | Compound | Chemical Structure |
|---|---|---|---|---|---|
| 1 | 4.09 | 215, 276, 315 | compound Rt 4 | ||
| 214.9, 277.4, 316.6 d | 593, 269 b | oroxin B * (baicalein-7-O-diglucoside) |  | ||
| 2 | 6.00 | 214, 276, 316 | compound B0 | ||
| 214.5, 277, 316.6 d | 431, 269, 251, 241, 223, 195 b | oroxin A * (baicalein7-O-glucoside) |  | ||
| 3 | 6.17 | 214, 276, 316 | 269, 445, 891 a | Baicalin (baicalein-7-O-glucuronide) |  |
| 4 | 7.42 | 217, 266, 303 | compound Rt 7 | ||
| 267.9, 305.9 d | 429, 253, 209, 175, 113 b | Chrysin-7-O-glucuronide * |  | ||
| 5 | 9.92 | 214, 273, 321 | 223, 251, 269 a | baicalein |  |
| 6 | 12.97 | 214, 261, 311 | 180, 181, 209, 253, 254 a | chrysin |  |
| Sample | Zone of Inhibition at the Concentration of 50 mg/mL (mm) | ||||
|---|---|---|---|---|---|
| S. intermedius | S. suis | P. aeruginosa | β-Hemolytic E. coli | S. aureus | |
| OIRL | 9.50 ± 0.70 | 0 | 0 | 20.00 ± 0.00 | 0 |
| OIYL | 0 | 0 | 0 | 0 | 0 |
| OISL | 17.00 ± 2.78 | 11.83 ± 2.25 | 11.50 ± 0.70 | 15.75 ± 1.77 | 10.50 ± 1.32 |
| OIYF | 12.83 ± 0.76 | 10.42 ± 2.13 | 10.25 ± 1.06 | 13.00 ± 1.41 | 11.33 ± 1.26 |
| AMC30 | 31.00 ± 1.73 | 25.60 ± 0.96 | 0 | 22.00 ± 1.73 | 25.60 ± 0.96 |
| DO30 | 15.67 ± 2.31 | 16.83 ± 1.29 | 14.25 ± 0.35 | 25.00 ± 1.41 | 16.83 ± 1.29 |
| SXT25 | 0 | 0 | 14.50 ± 0.70 | 25.67 ± 0.58 | 0 |
| Bacterial Species | IC50 (mg/mL) | MIC (mg/mL) |
|---|---|---|
| S. intermedius | 0.81 ± 0.04 | 5.17 ± 1.23 |
| S. suis | 0.96 ± 0.46 | 10.54 ± 1.00 |
| P. aeruginosa | 3.59 ± 1.01 | 46.72 ± 1.94 |
| β-hemolytic E. coli | 6.57 ± 0.69 | 19.11 ± 0.62 |
| S. aureus | 0.40 ± 0.00 | 4.67 ± 0.04 |
| 1/DPPH (2,2-Diphenyl-1-Picrylhydrazyl) | S. intermedius | S. suis | P. aeruginosa | β-Hemolytic E. coli | S. aureus | |
|---|---|---|---|---|---|---|
| TPC | 0.698 | 0.441 | 0.838 | 0.842 | 0.116 | 0.890 |
| TFC | 0.689 | 0.925 | 0.920 | 0.917 | 0.505 | 0.862 |
| B1 | 0.665 | 0.545 | 0.055 | 0.049 | 0.774 | 0.043 |
| B2 | 0.455 | 0.208 | 0.439 | 0.446 | 0.022 | 0.562 |
| C | 0.196 | 0.164 | 0.433 | 0.441 | 0.050 | 0.557 |
| B1 + B2 + C | 0.593 | 0.023 | 0.064 | 0.072 | 0.066 | 0.203 |
| B1 + B0 | 0.573 | 0.406 | 0.827 | 0.830 | 0.171 | 0.874 |
| B0 | 0.556 | 0.940 | 0.957 | 0.956 | 0.516 | 0.920 |
| Rt 4 | 0.574 | 0.488 | 0.785 | 0.790 | 0.063 | 0.863 |
| Rt 7 | 0.598 | 0.316 | 0.790 | 0.791 | 0.344 | 0.793 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sithisarn, P.; Rojsanga, P.; Sithisarn, P. Flavone-Rich Fractions and Extracts from Oroxylum indicum and Their Antibacterial Activities against Clinically Isolated Zoonotic Bacteria and Free Radical Scavenging Effects. Molecules 2021, 26, 1773. https://doi.org/10.3390/molecules26061773
Sithisarn P, Rojsanga P, Sithisarn P. Flavone-Rich Fractions and Extracts from Oroxylum indicum and Their Antibacterial Activities against Clinically Isolated Zoonotic Bacteria and Free Radical Scavenging Effects. Molecules. 2021; 26(6):1773. https://doi.org/10.3390/molecules26061773
Chicago/Turabian StyleSithisarn, Patchima, Piyanuch Rojsanga, and Pongtip Sithisarn. 2021. "Flavone-Rich Fractions and Extracts from Oroxylum indicum and Their Antibacterial Activities against Clinically Isolated Zoonotic Bacteria and Free Radical Scavenging Effects" Molecules 26, no. 6: 1773. https://doi.org/10.3390/molecules26061773
APA StyleSithisarn, P., Rojsanga, P., & Sithisarn, P. (2021). Flavone-Rich Fractions and Extracts from Oroxylum indicum and Their Antibacterial Activities against Clinically Isolated Zoonotic Bacteria and Free Radical Scavenging Effects. Molecules, 26(6), 1773. https://doi.org/10.3390/molecules26061773