
Pulsed Power Applications for Protein Conformational Change and the Permeabilization of Agricultural Products



Abstract
:1. Introduction
2. Pulsed Power Generators for Food Processing
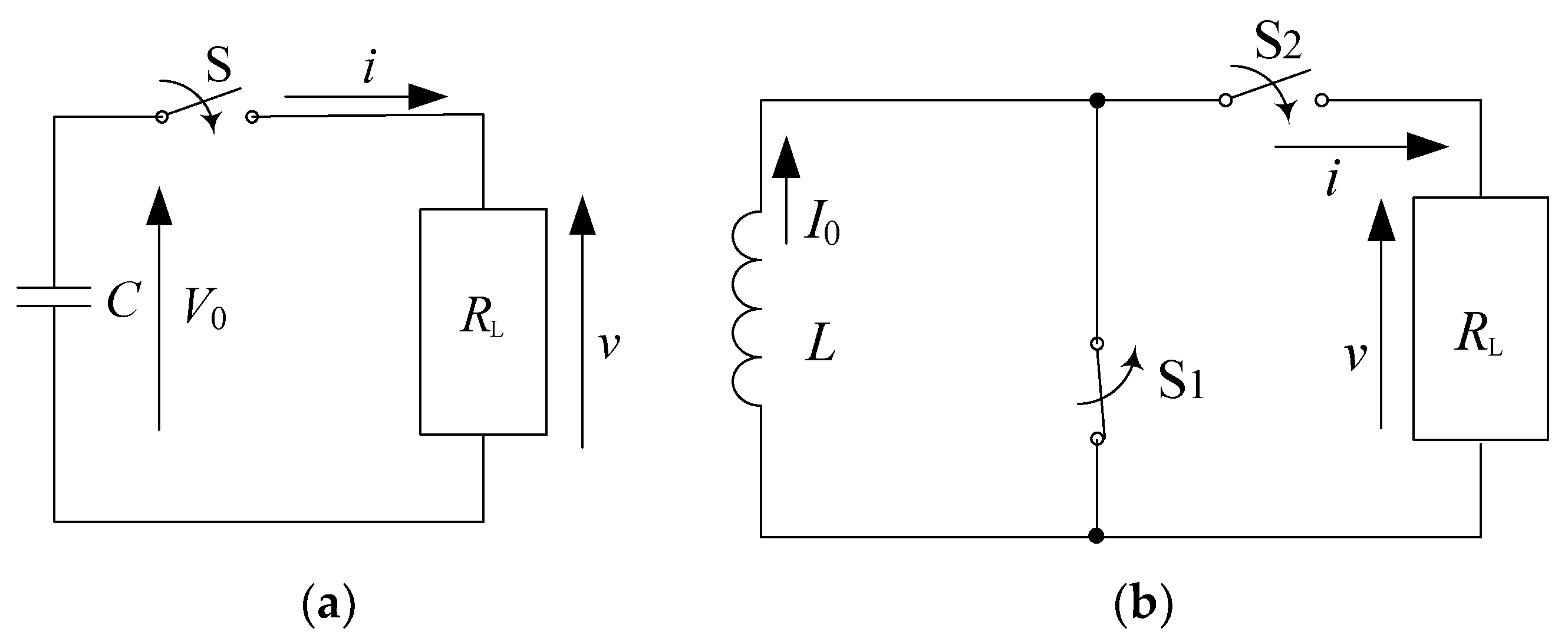
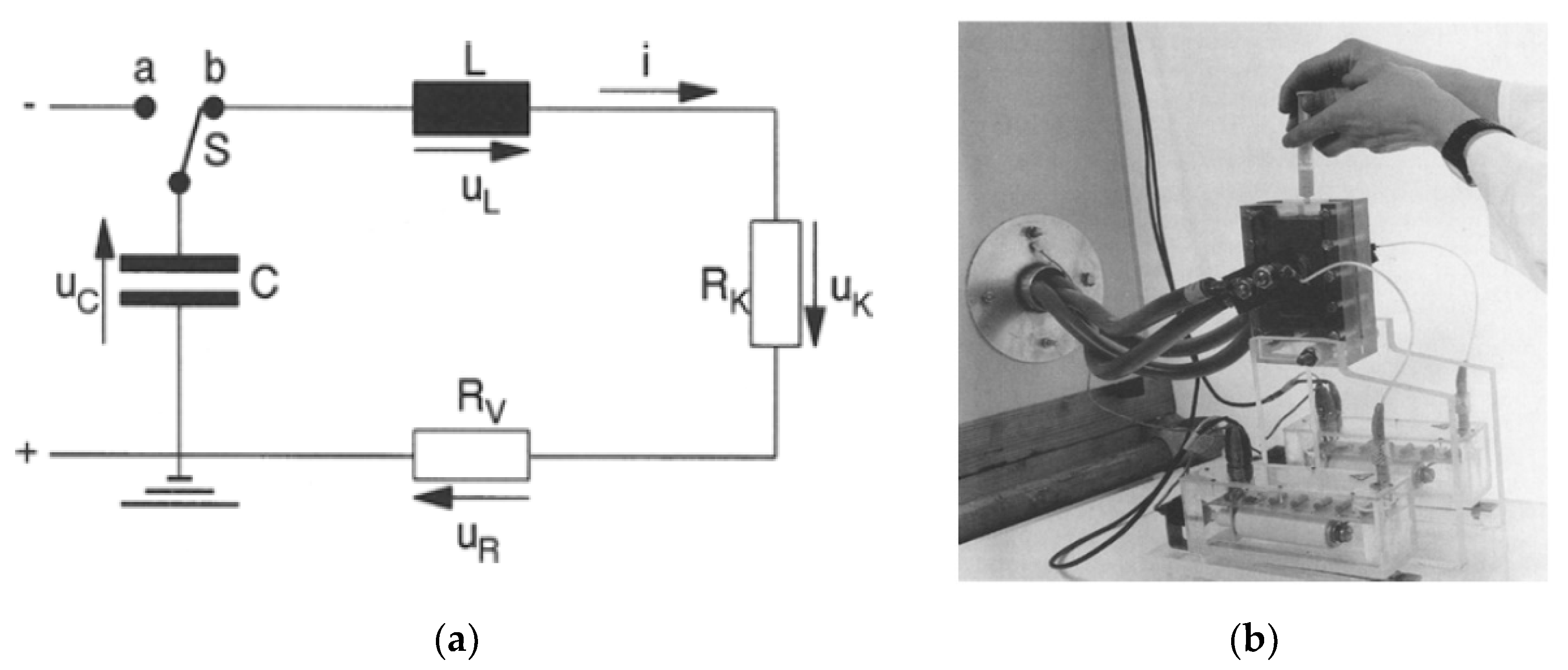
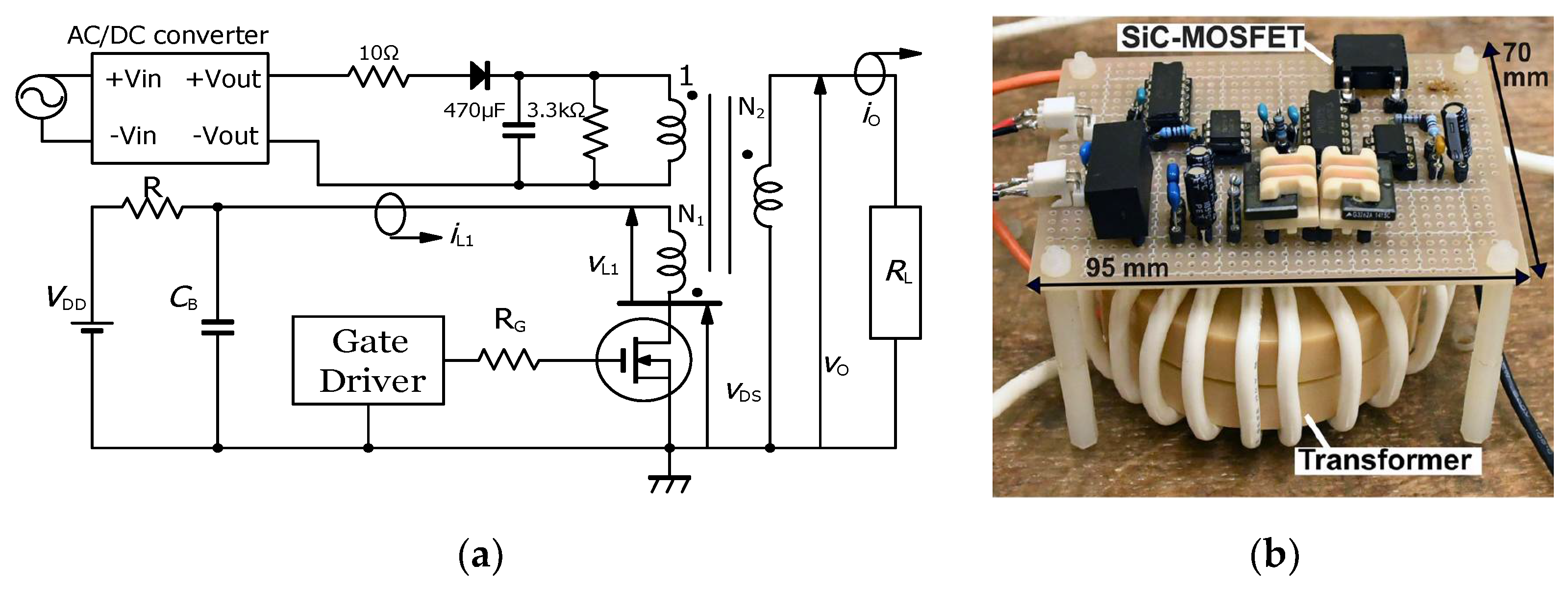
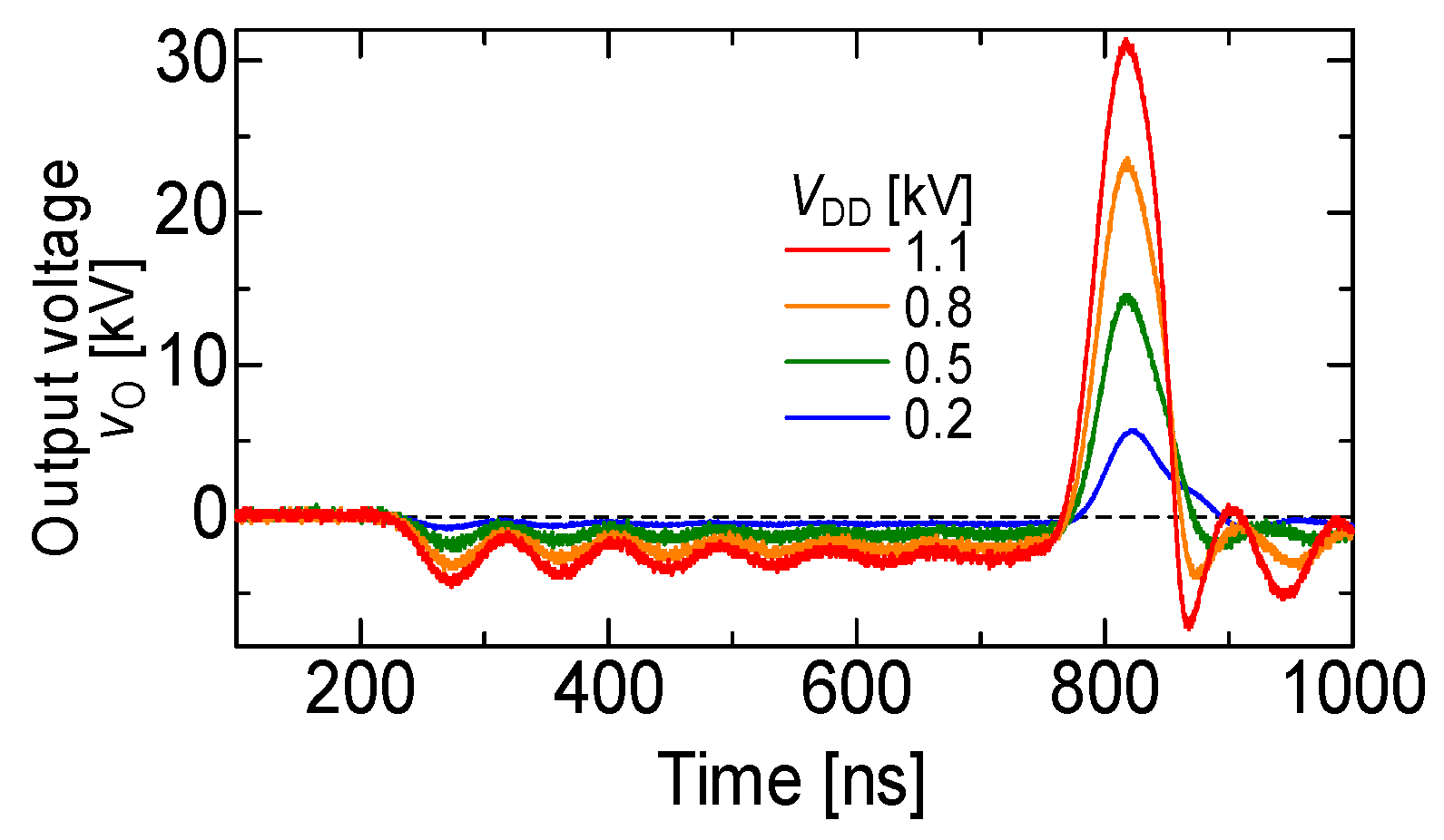
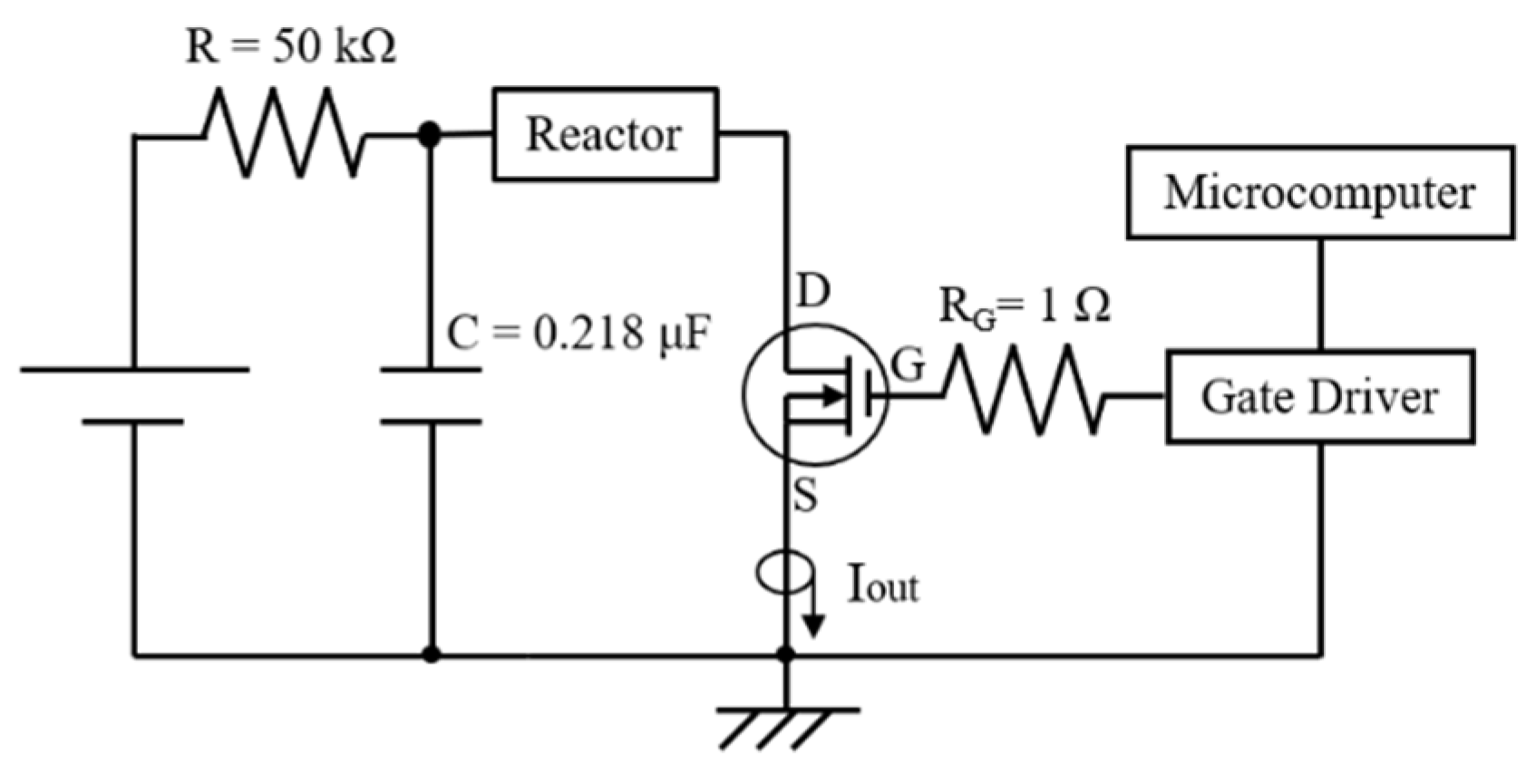
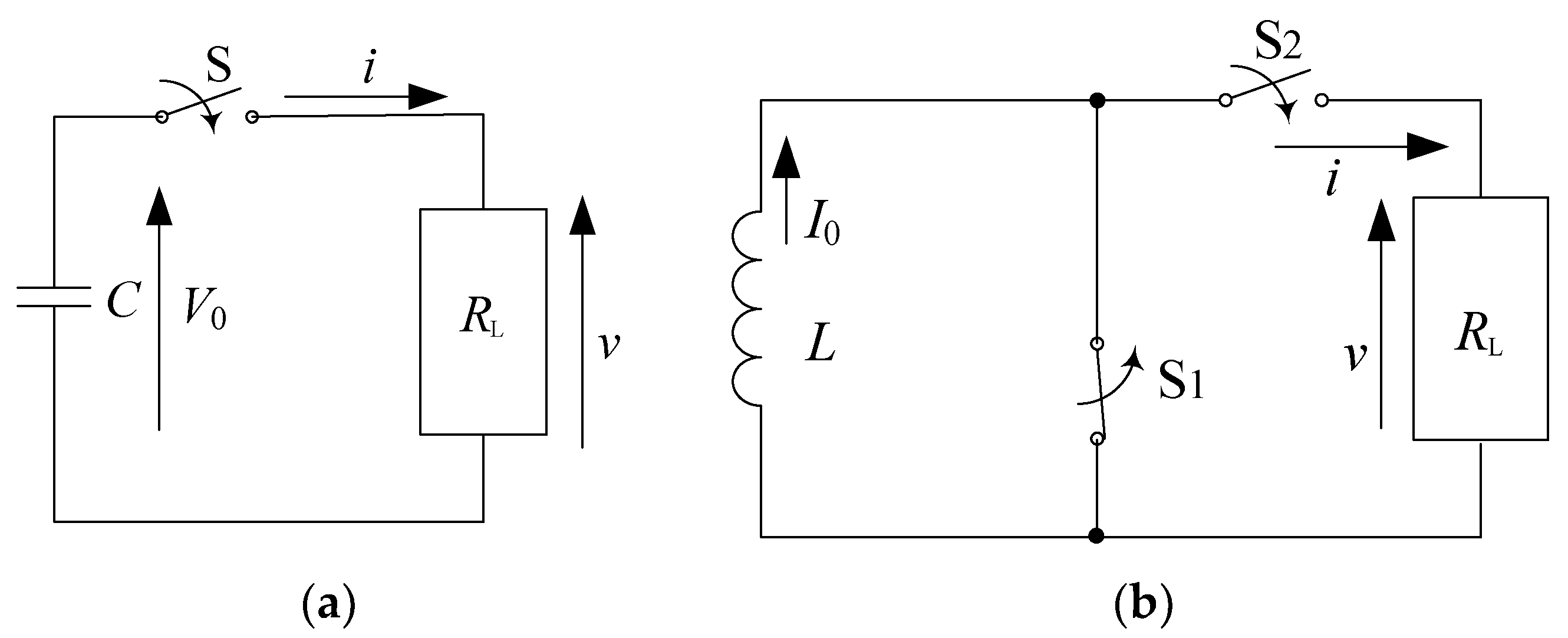
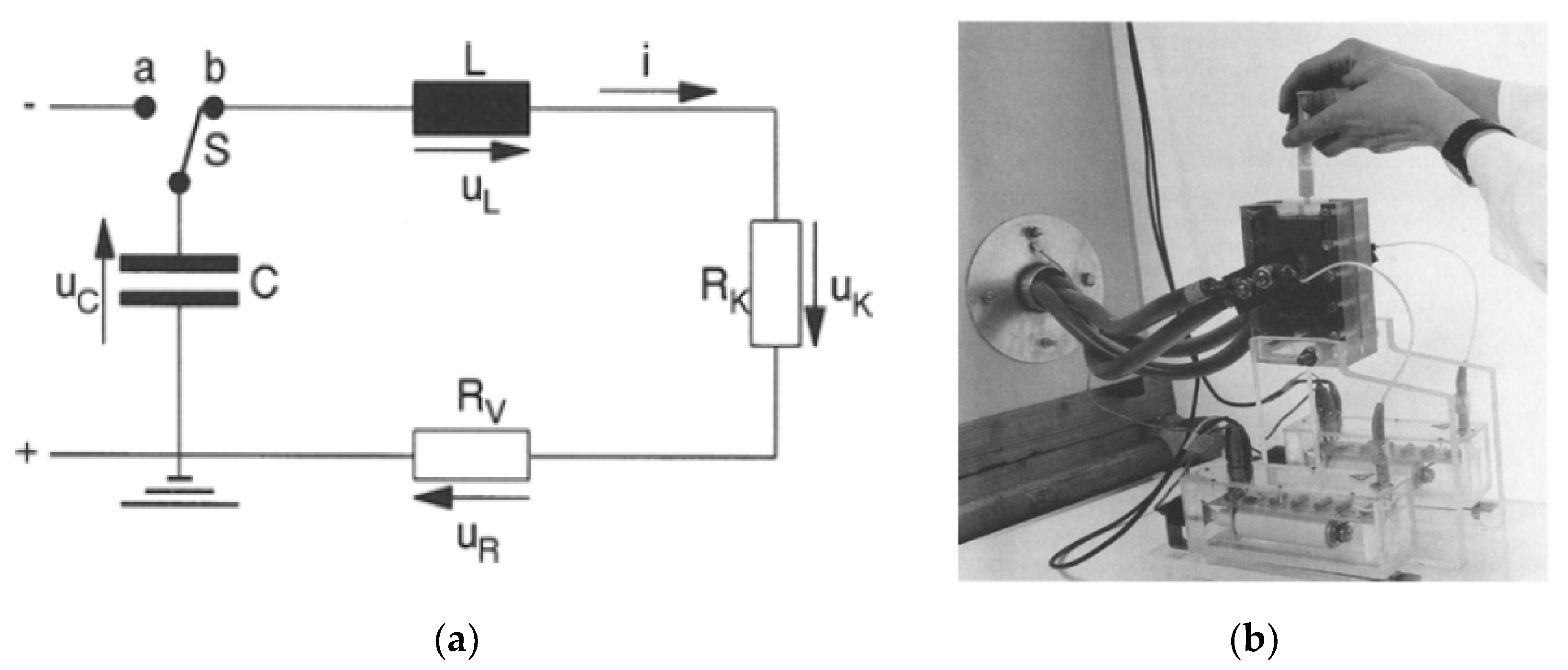
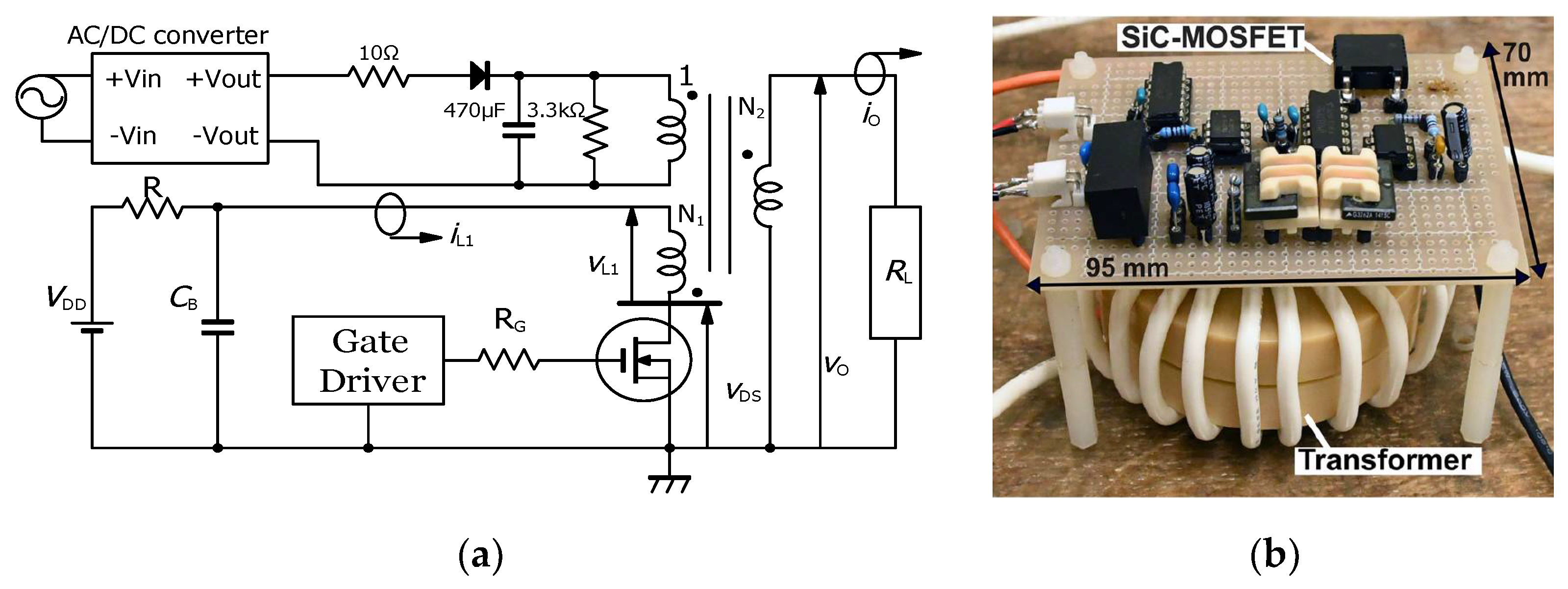
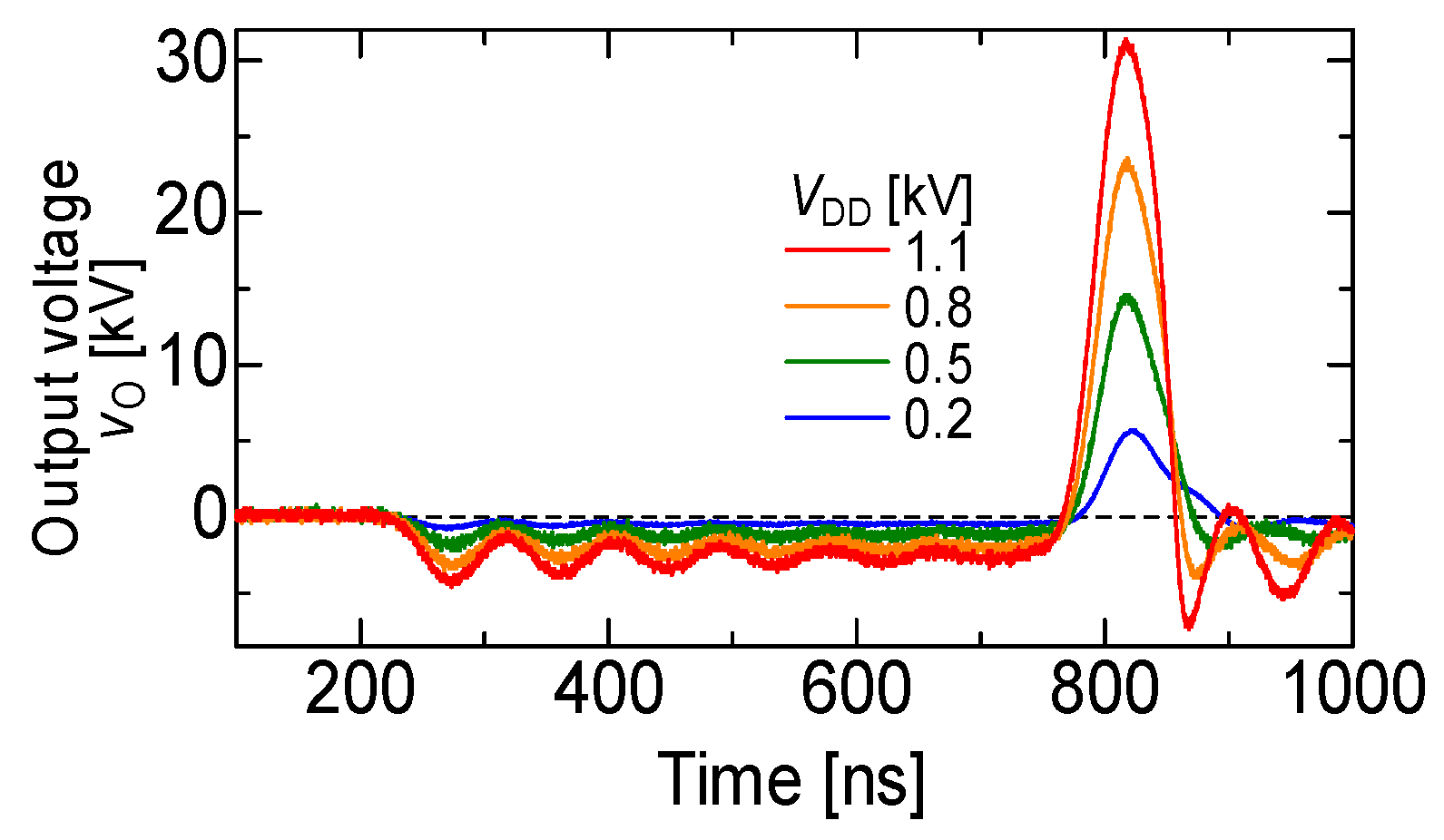
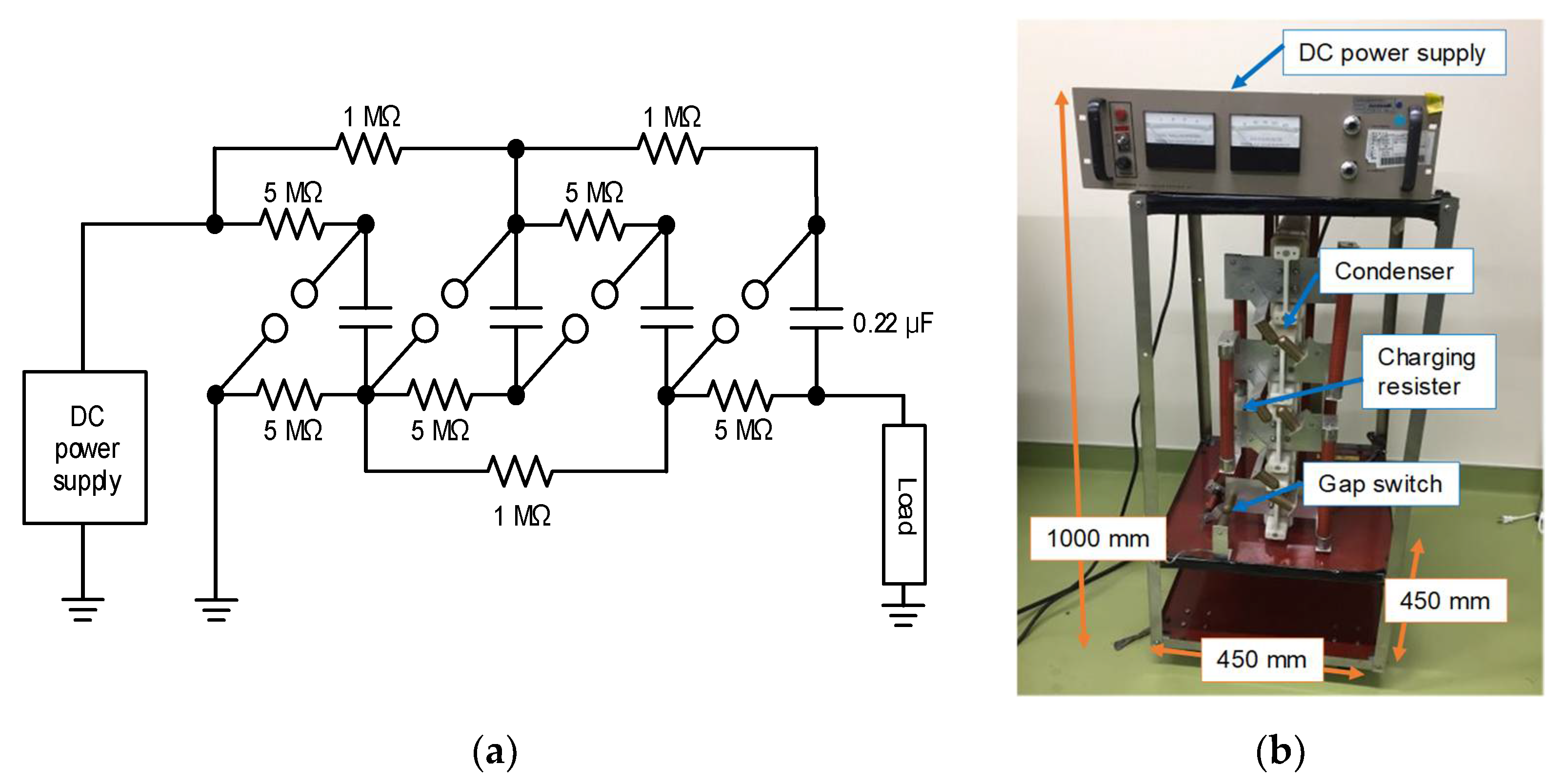
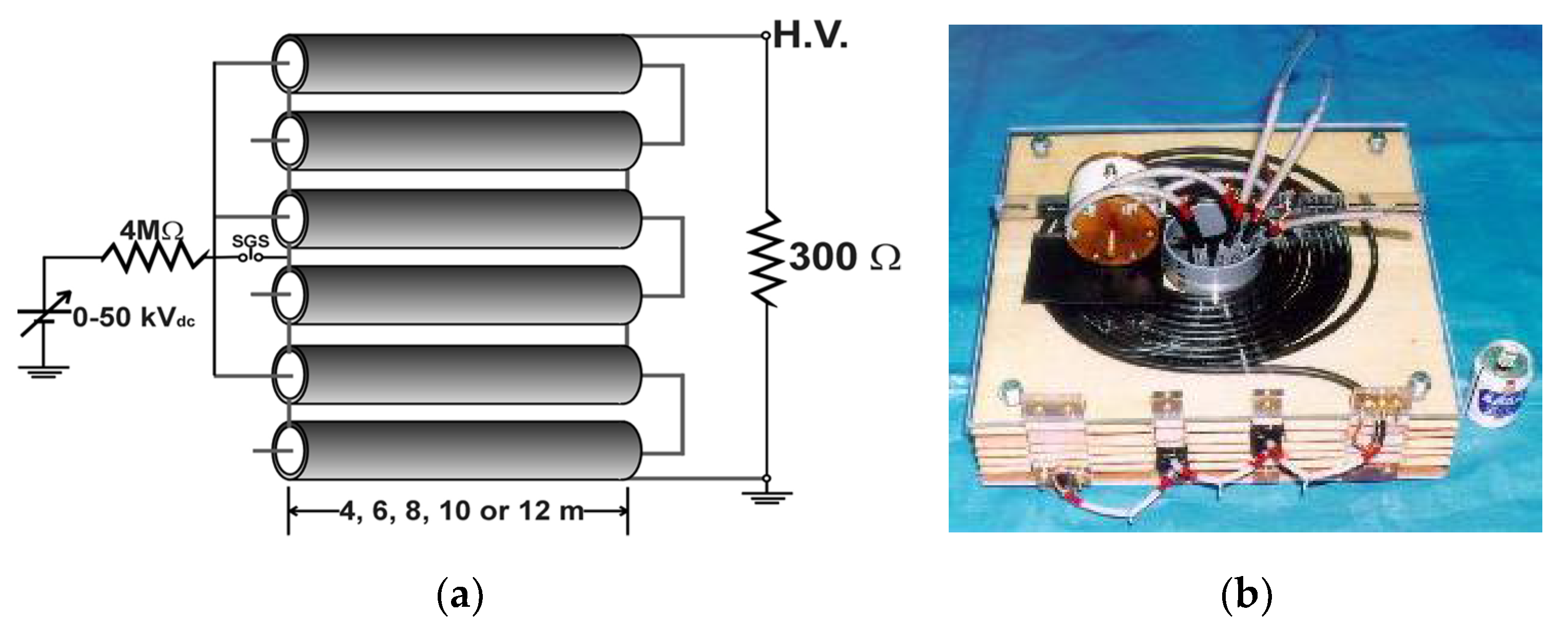
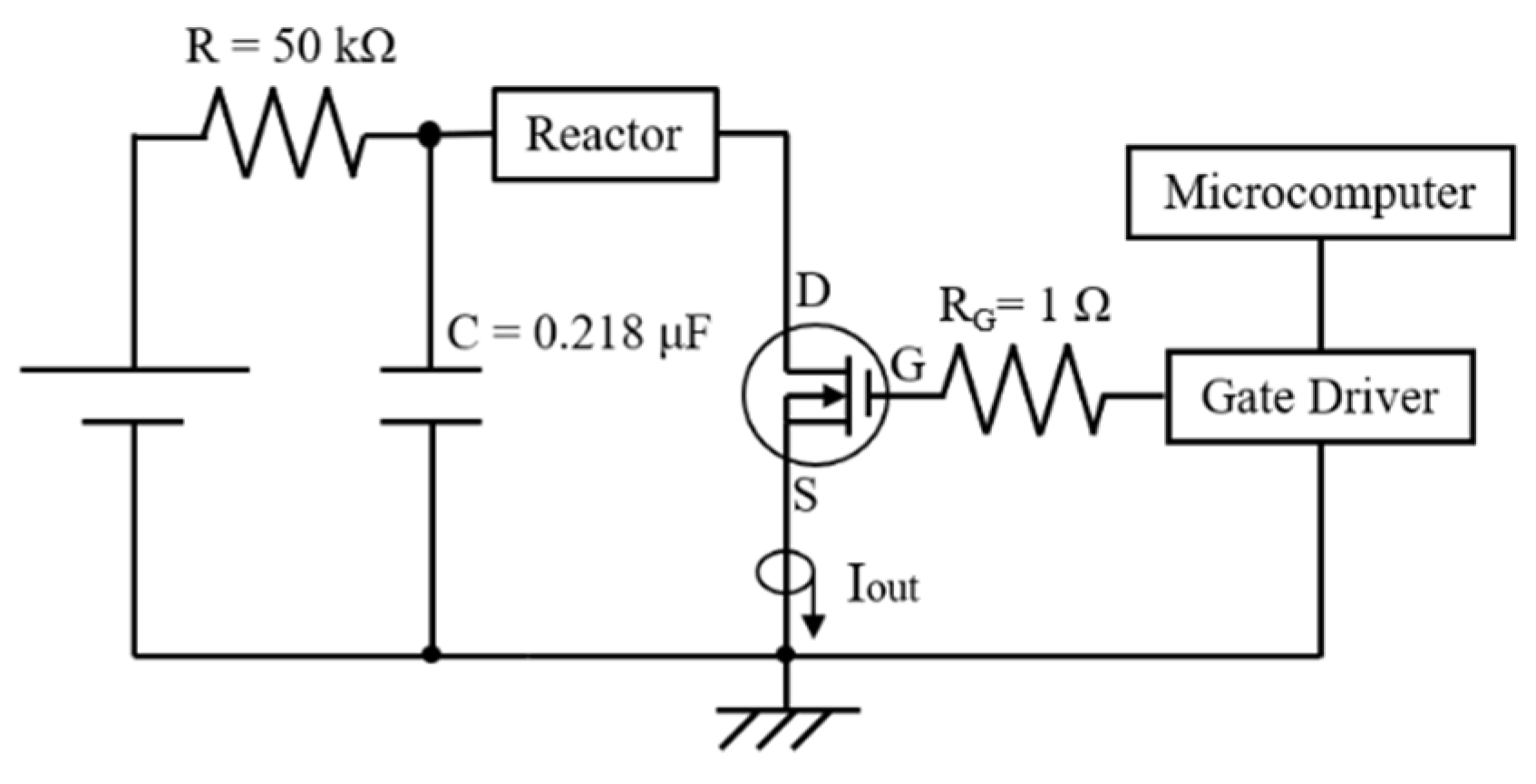
2.1. Basic Circuit for Pulsed Power
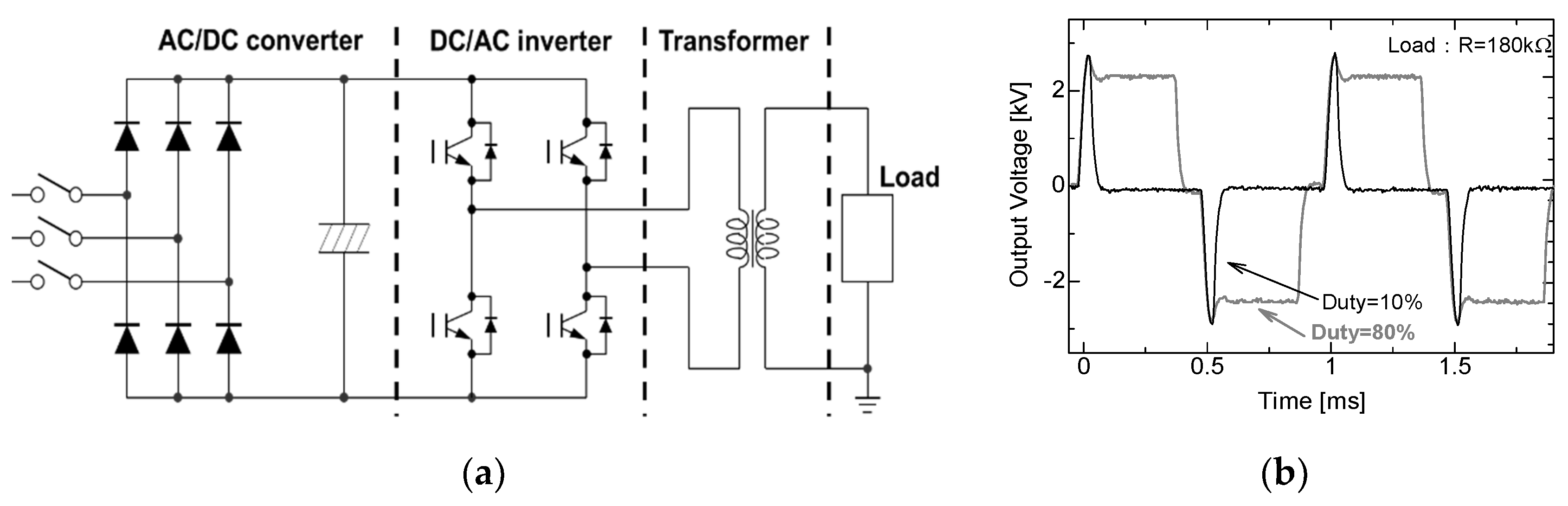
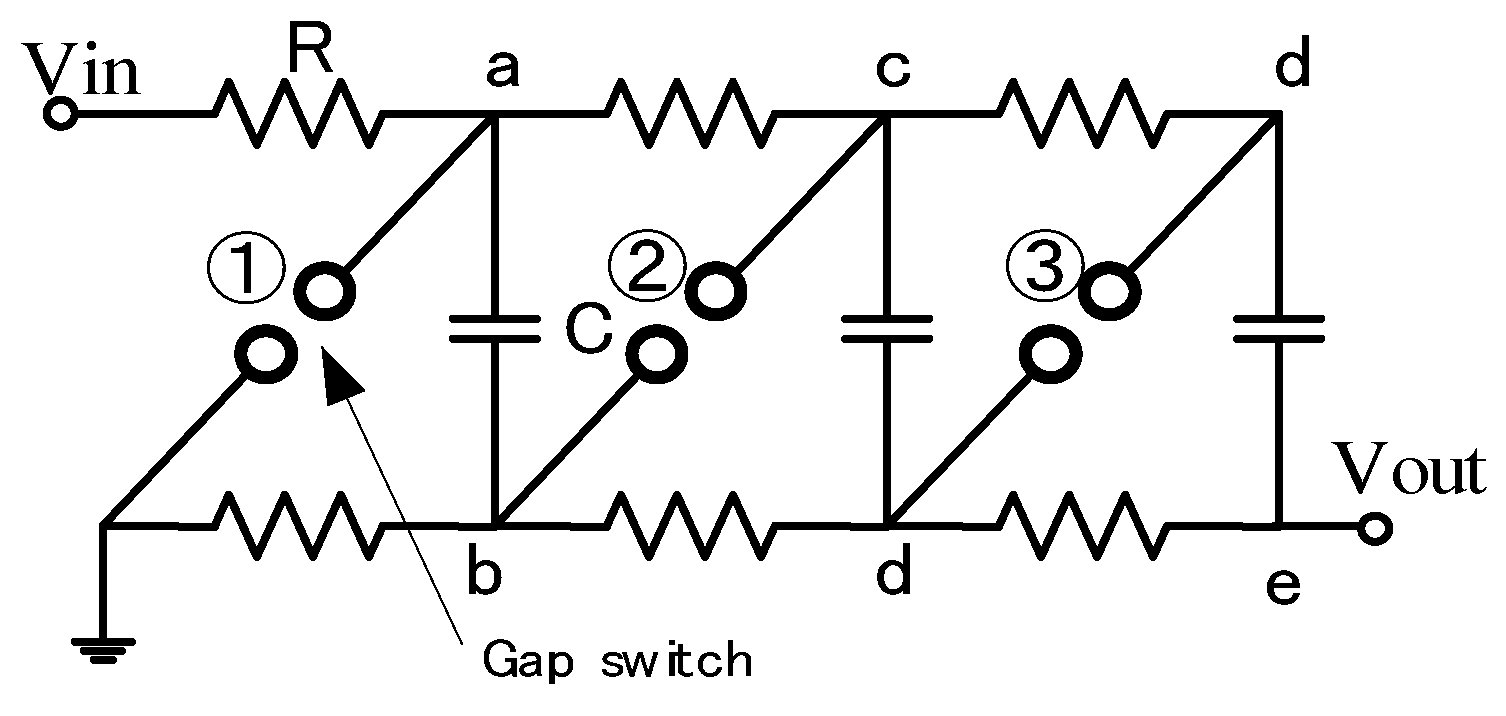
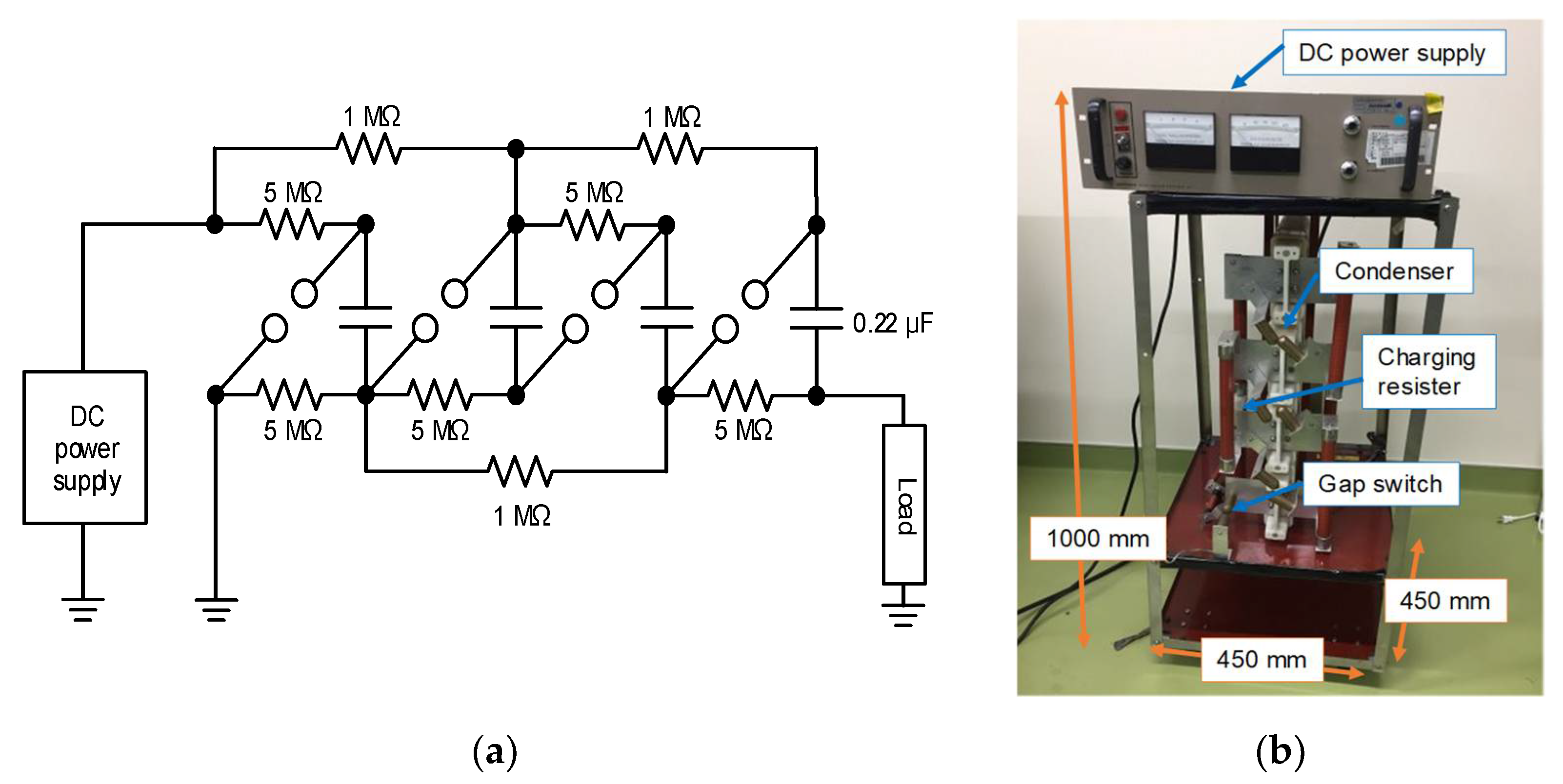
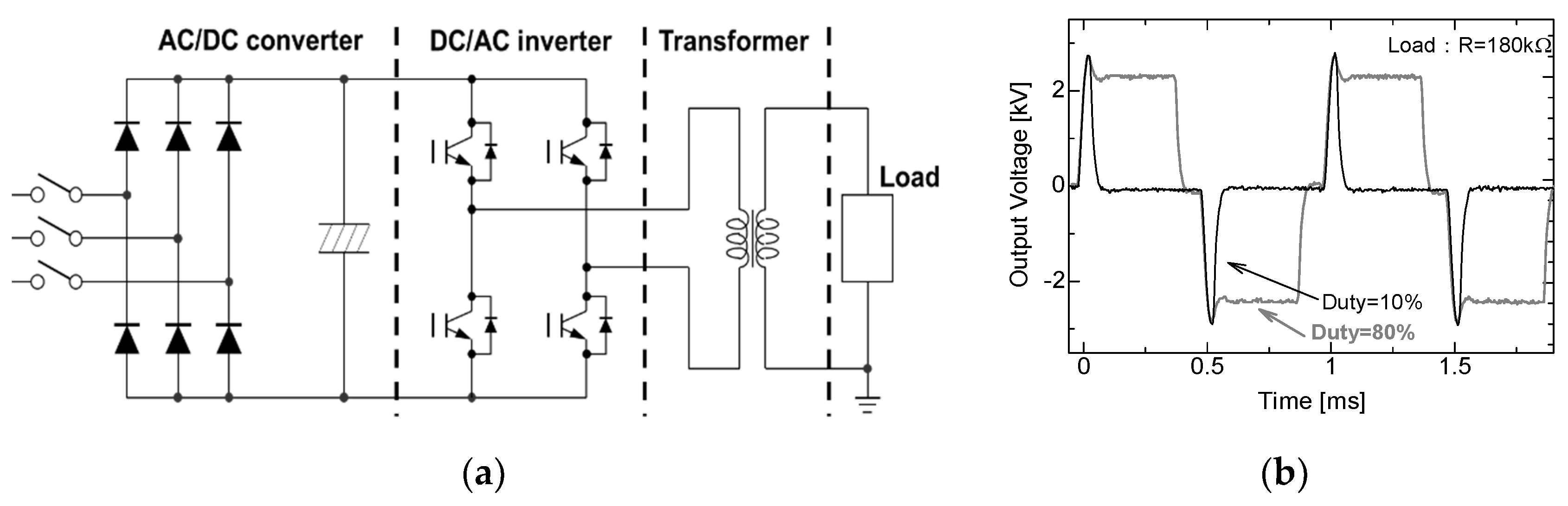
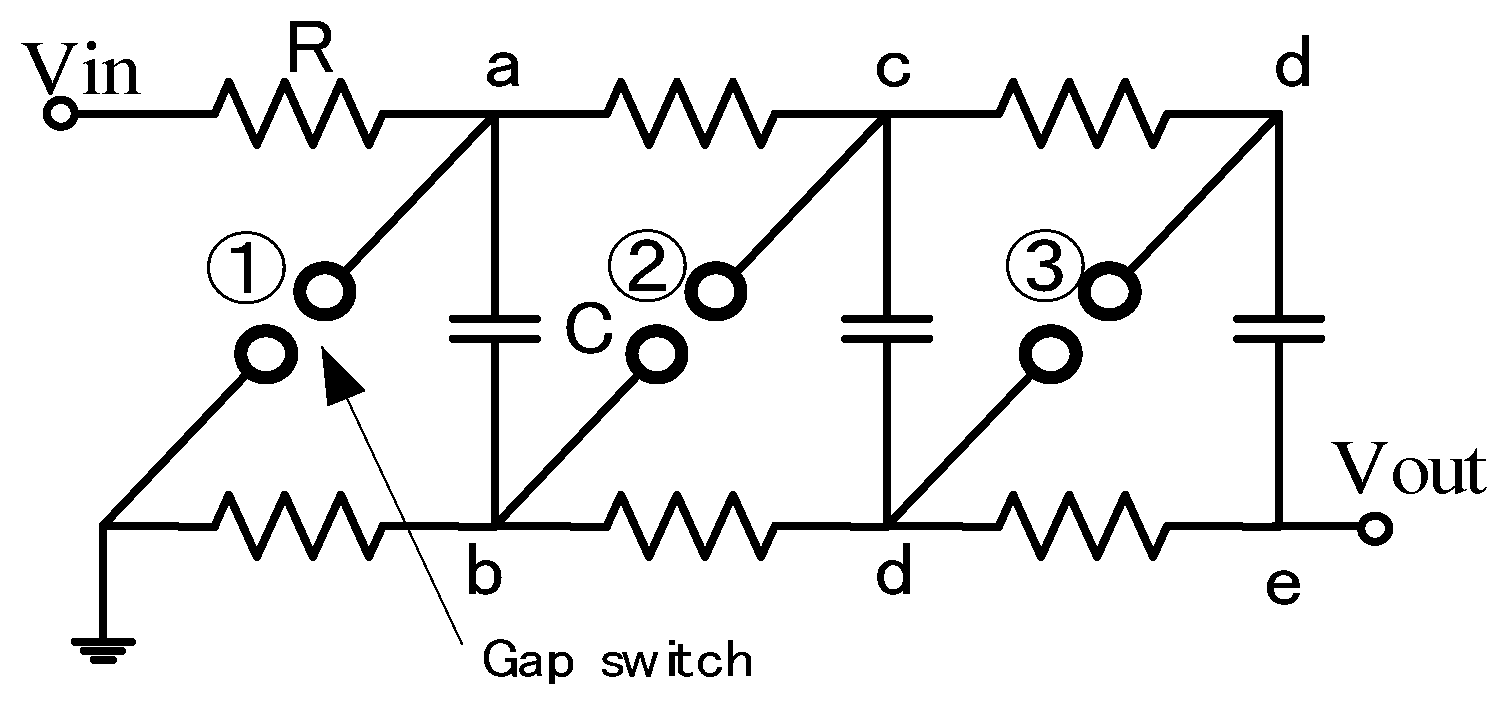
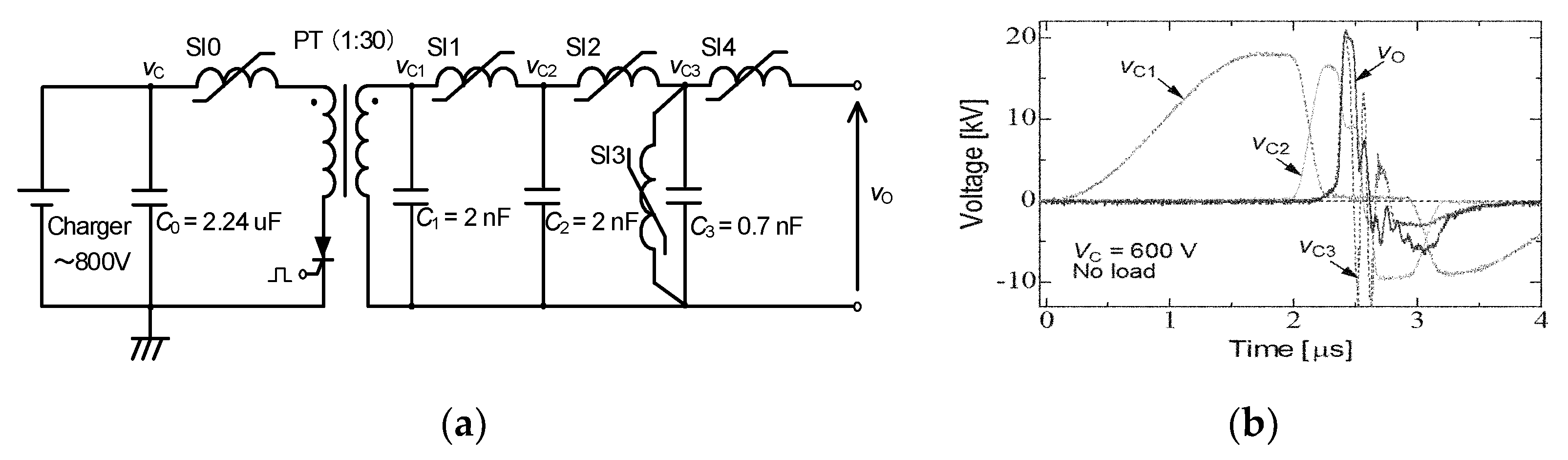
2.2. Cascade Connection for Voltage Multiplication
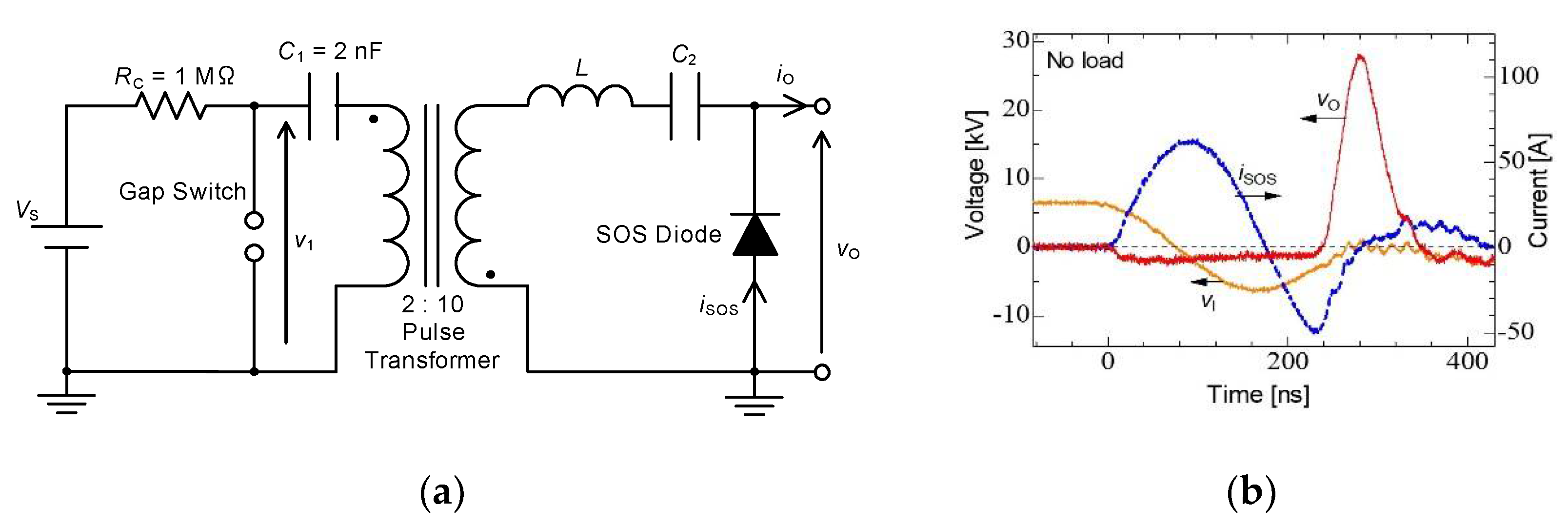
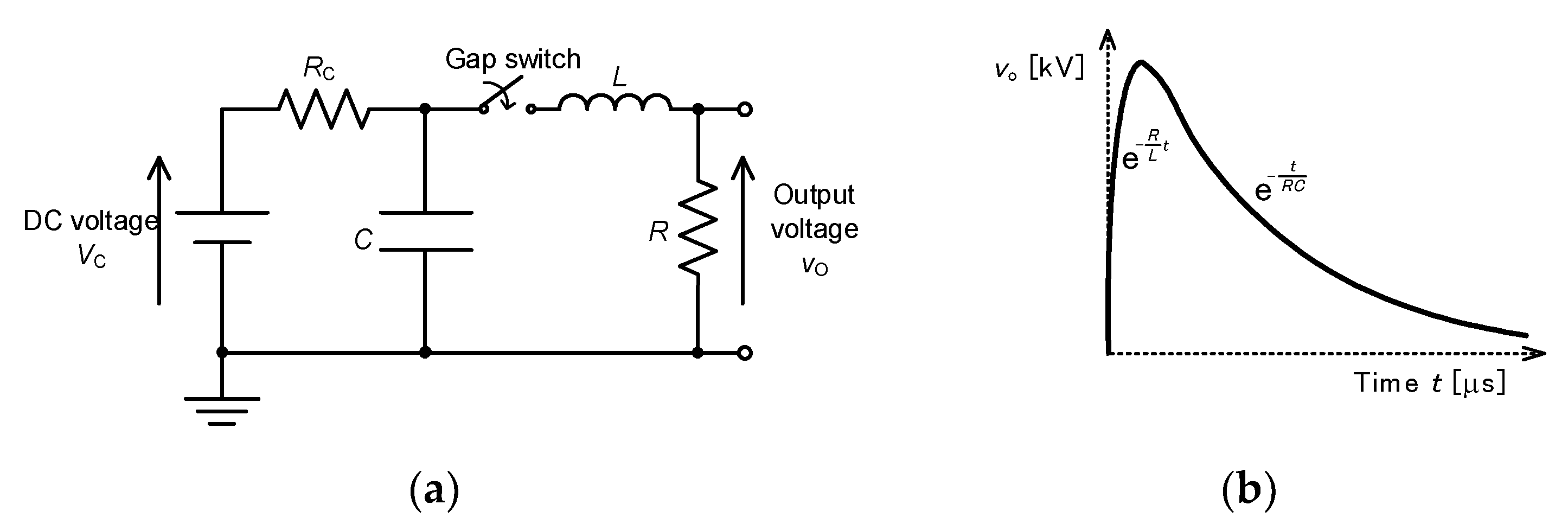
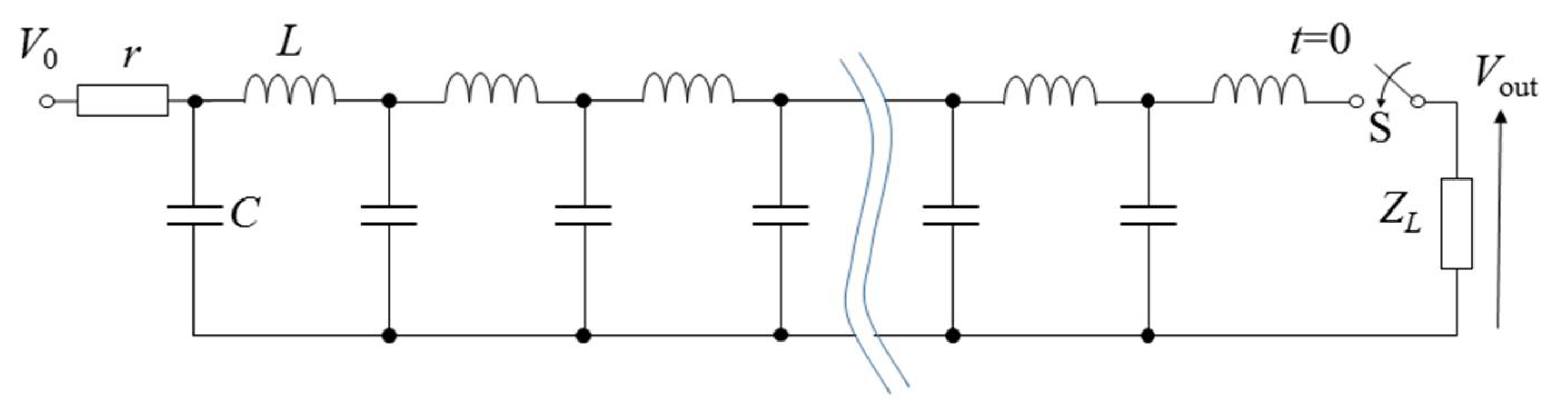
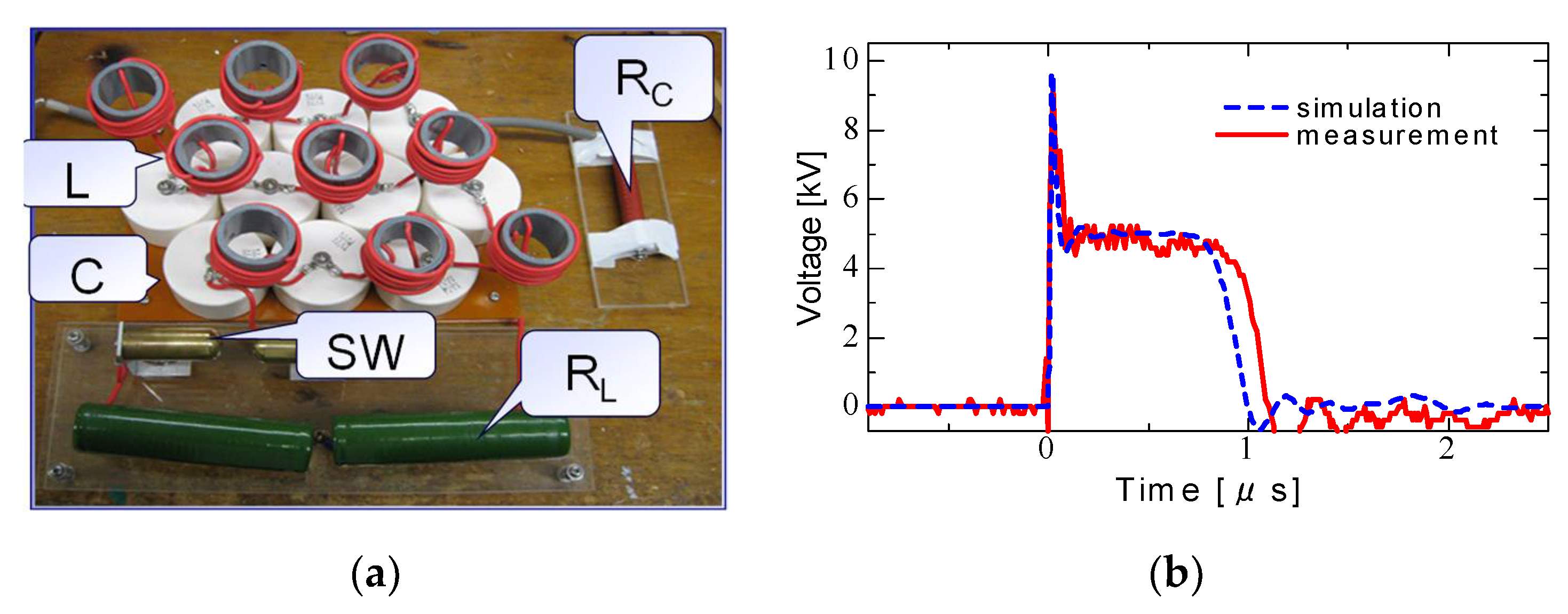
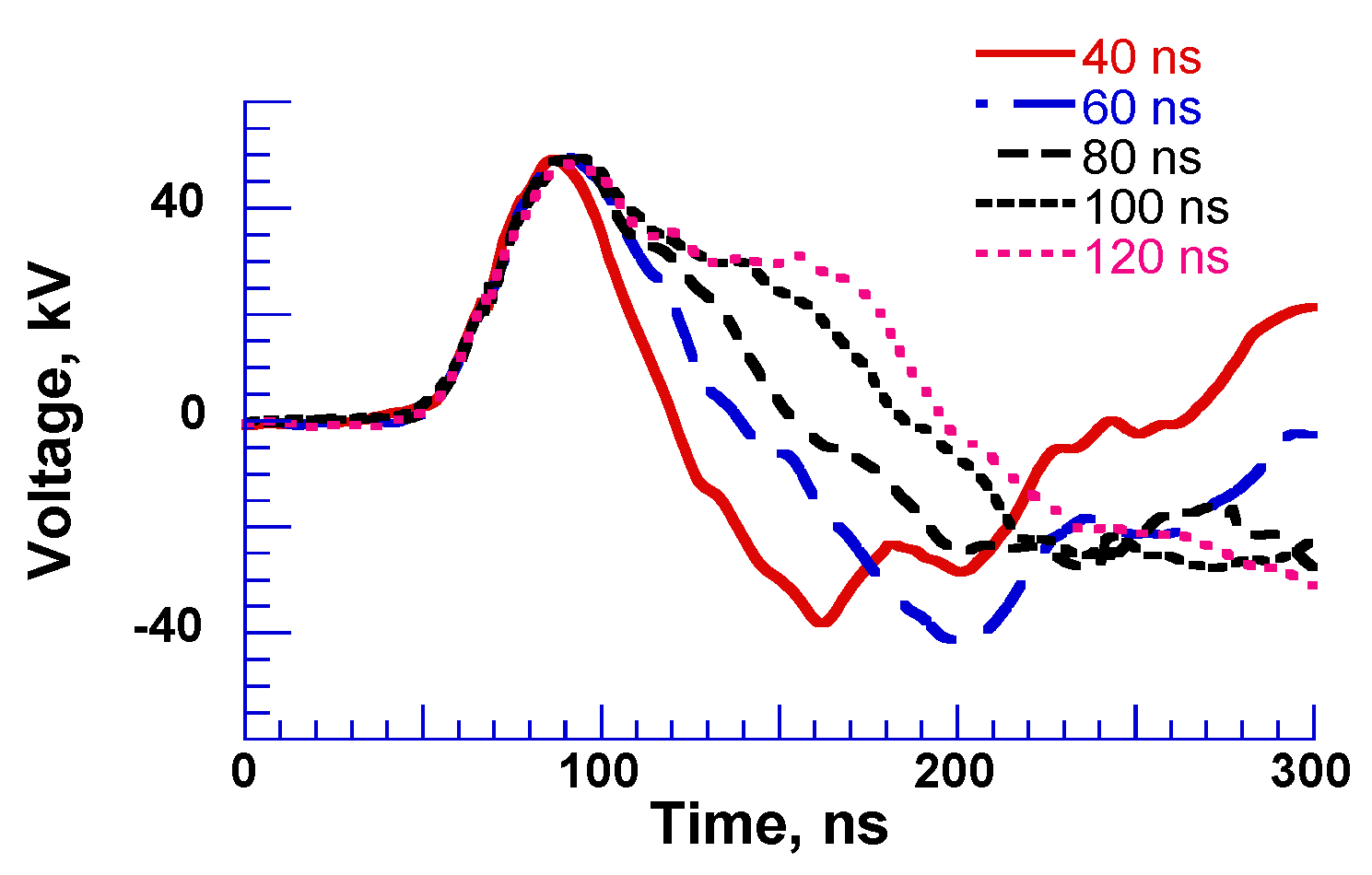
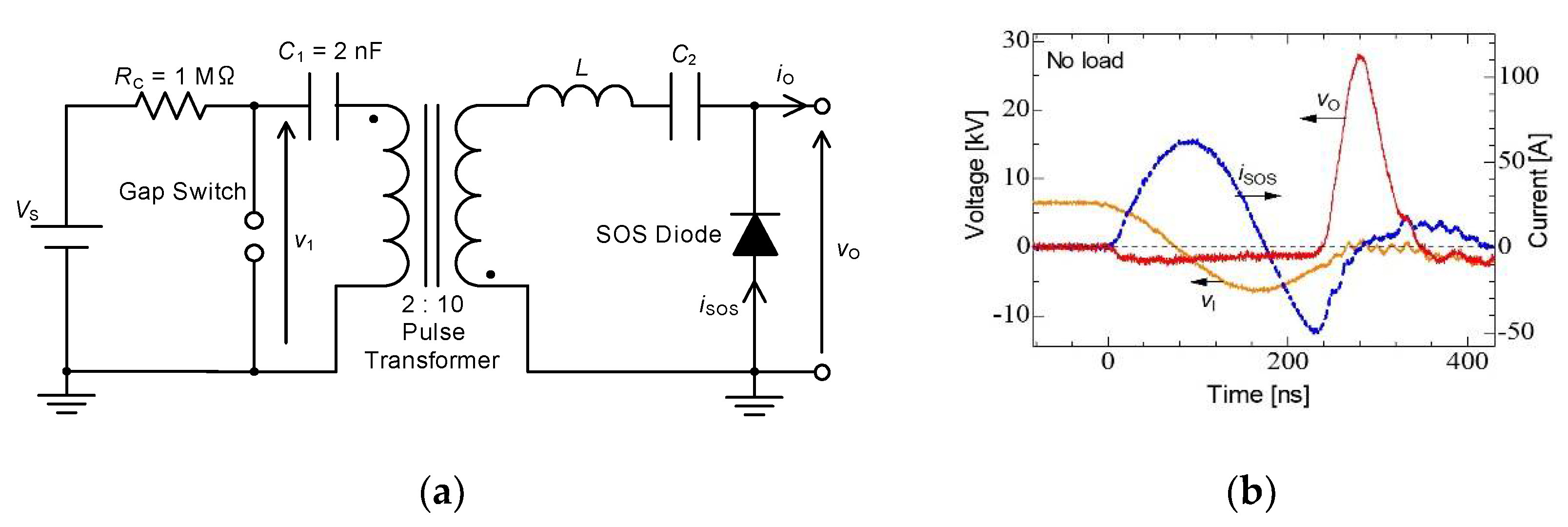
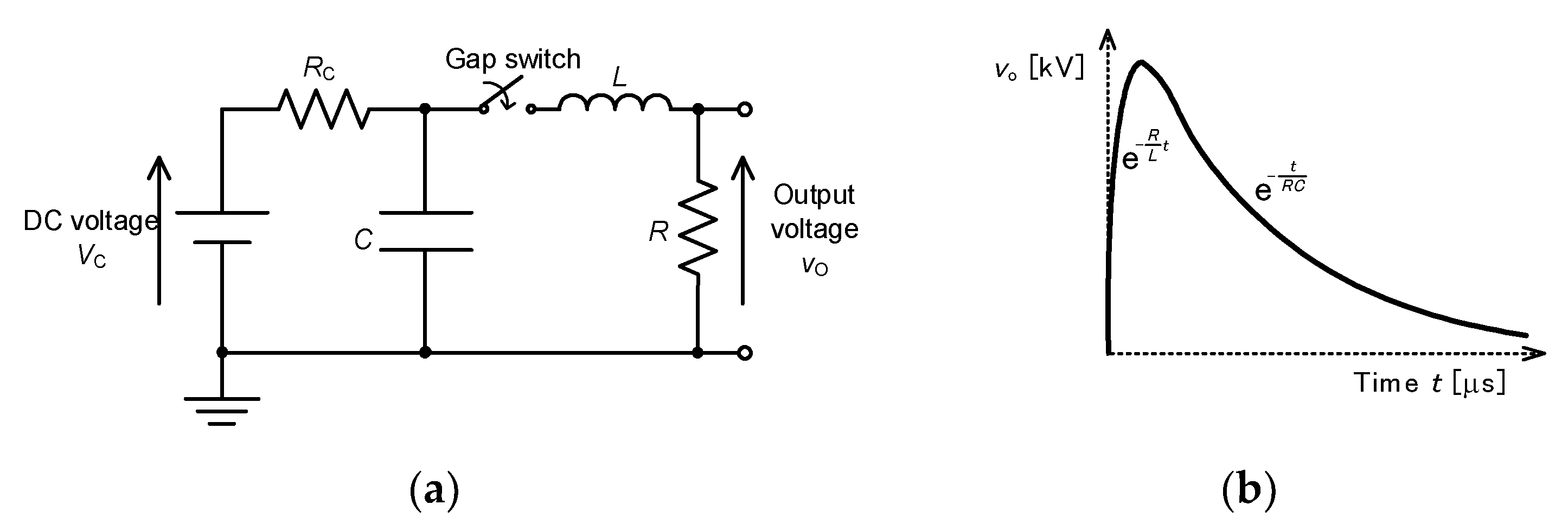
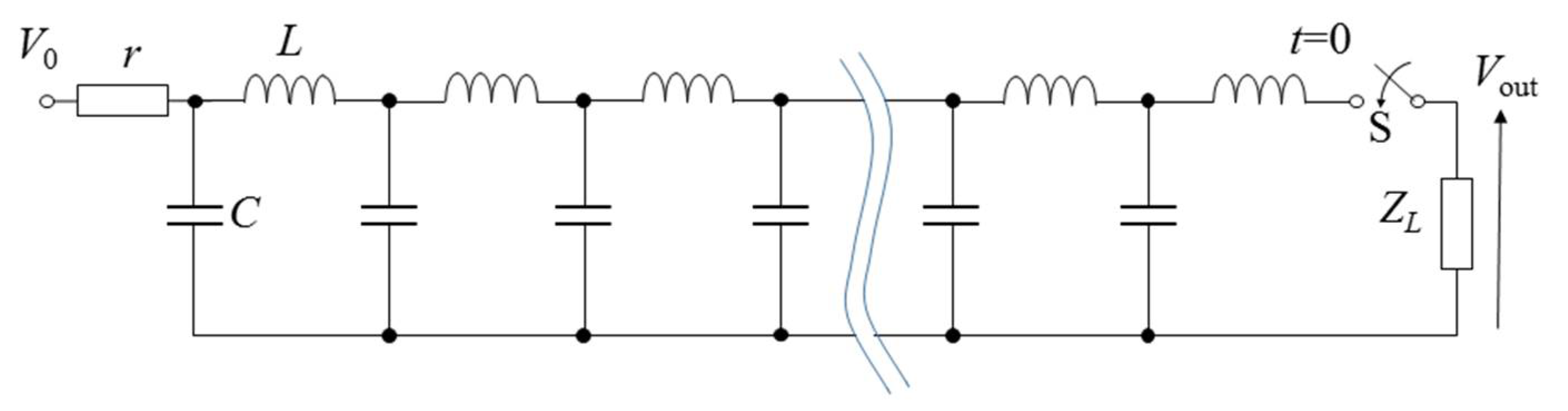
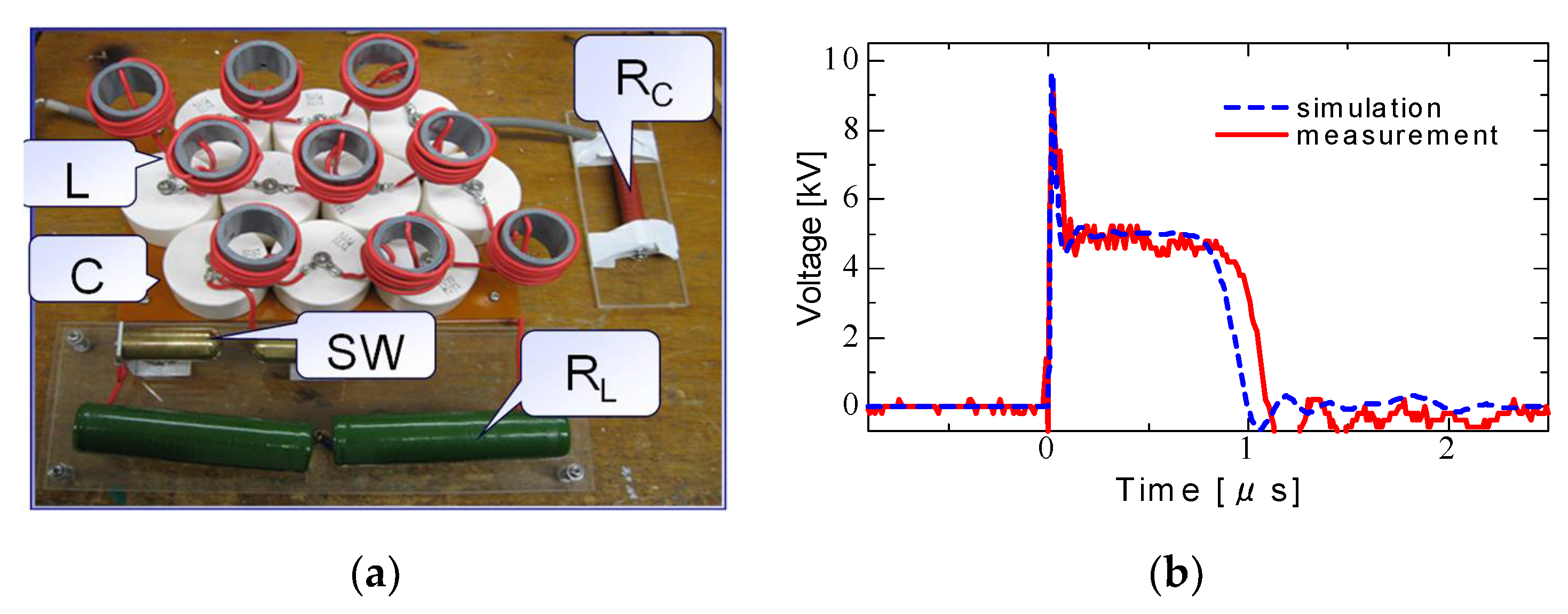
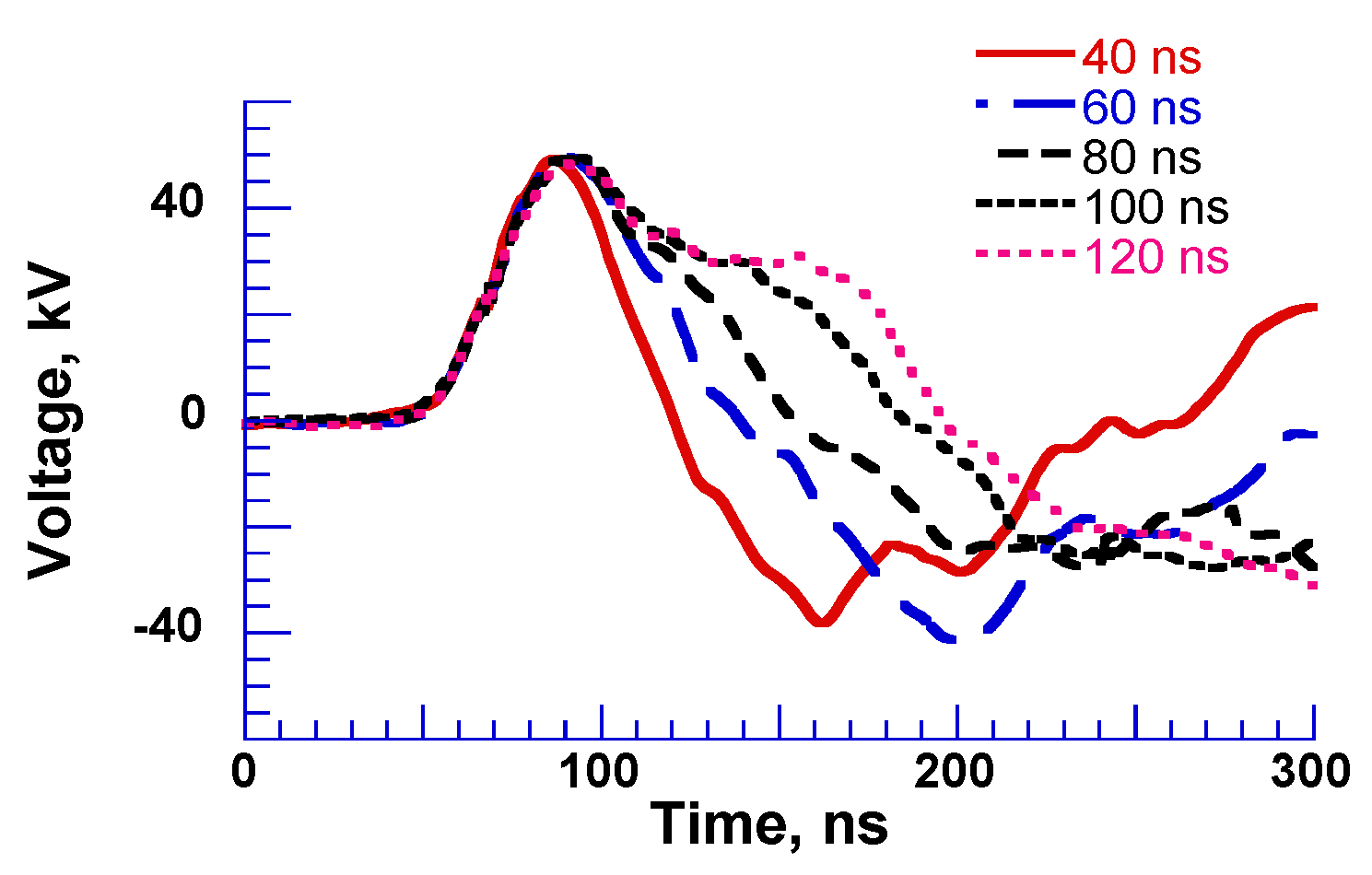
2.3. Pulse Compression and Forming
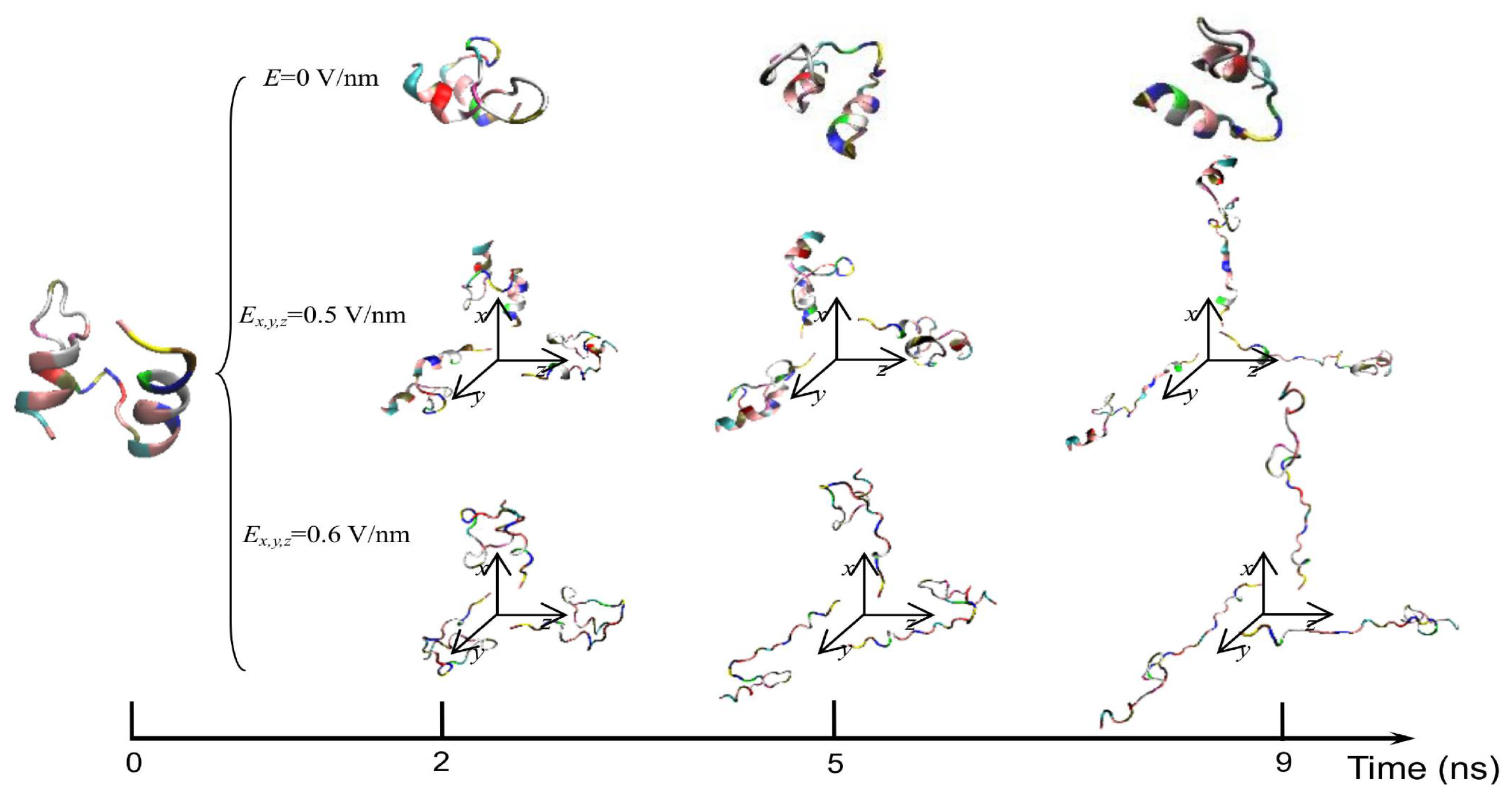
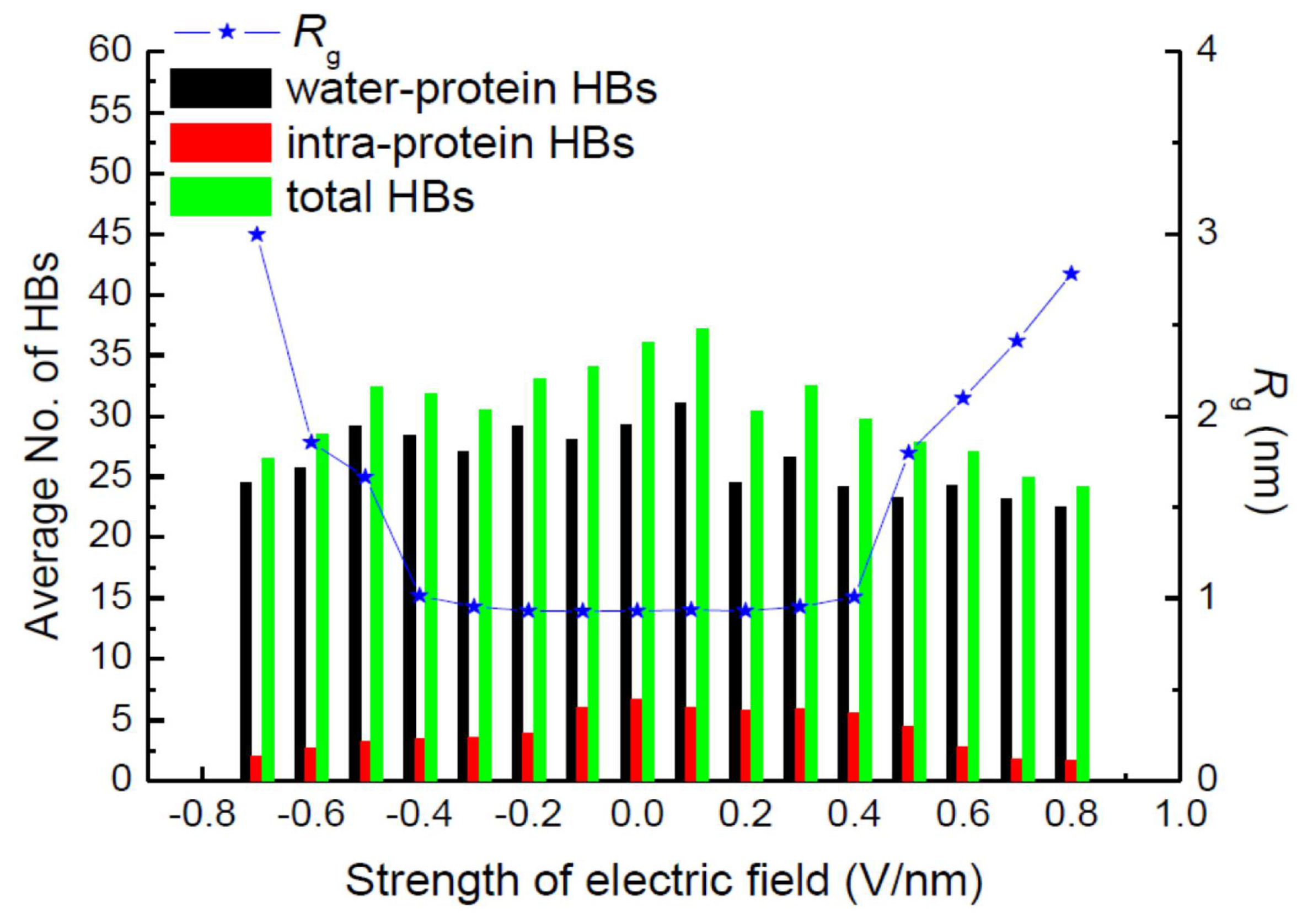
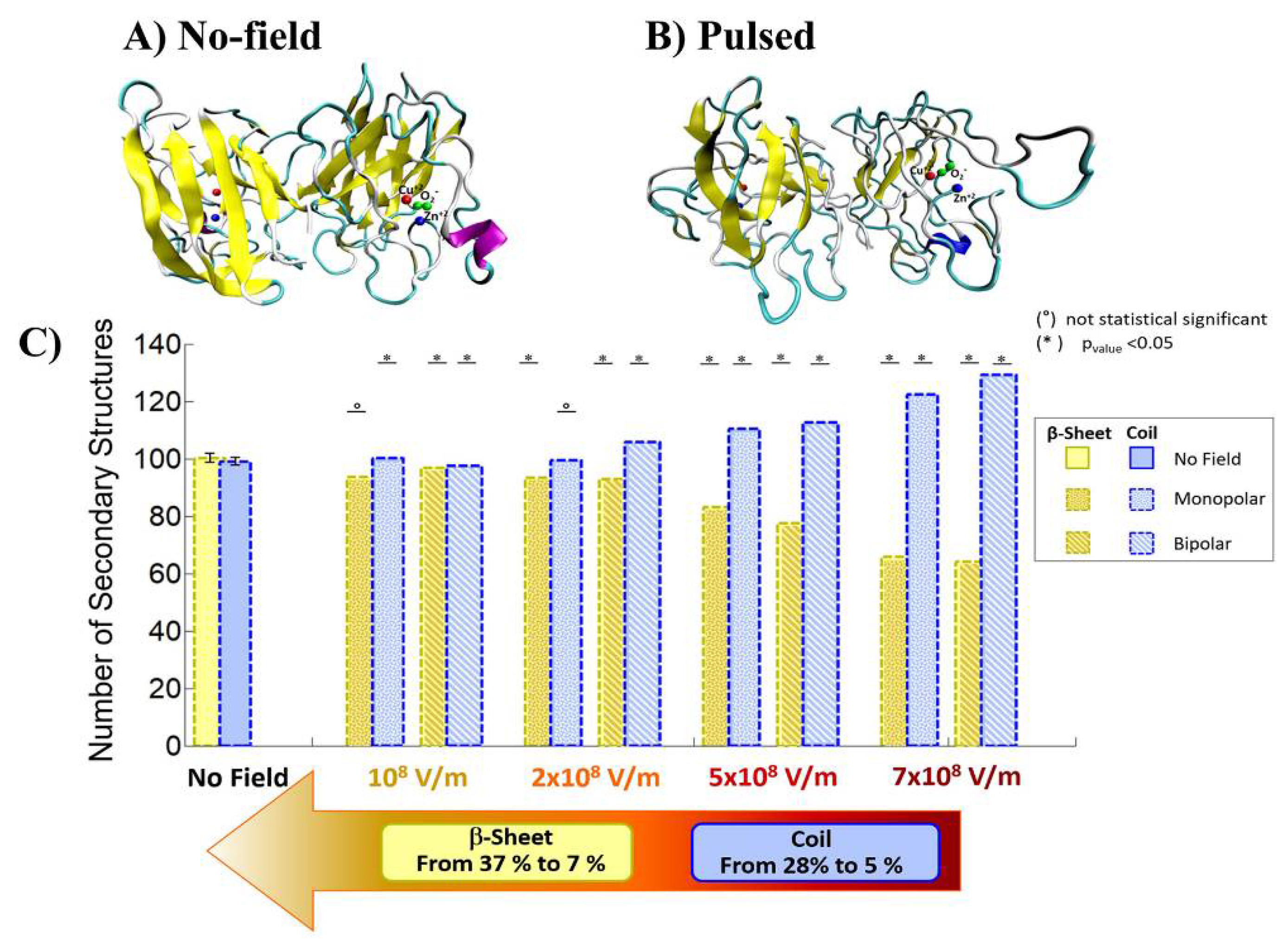
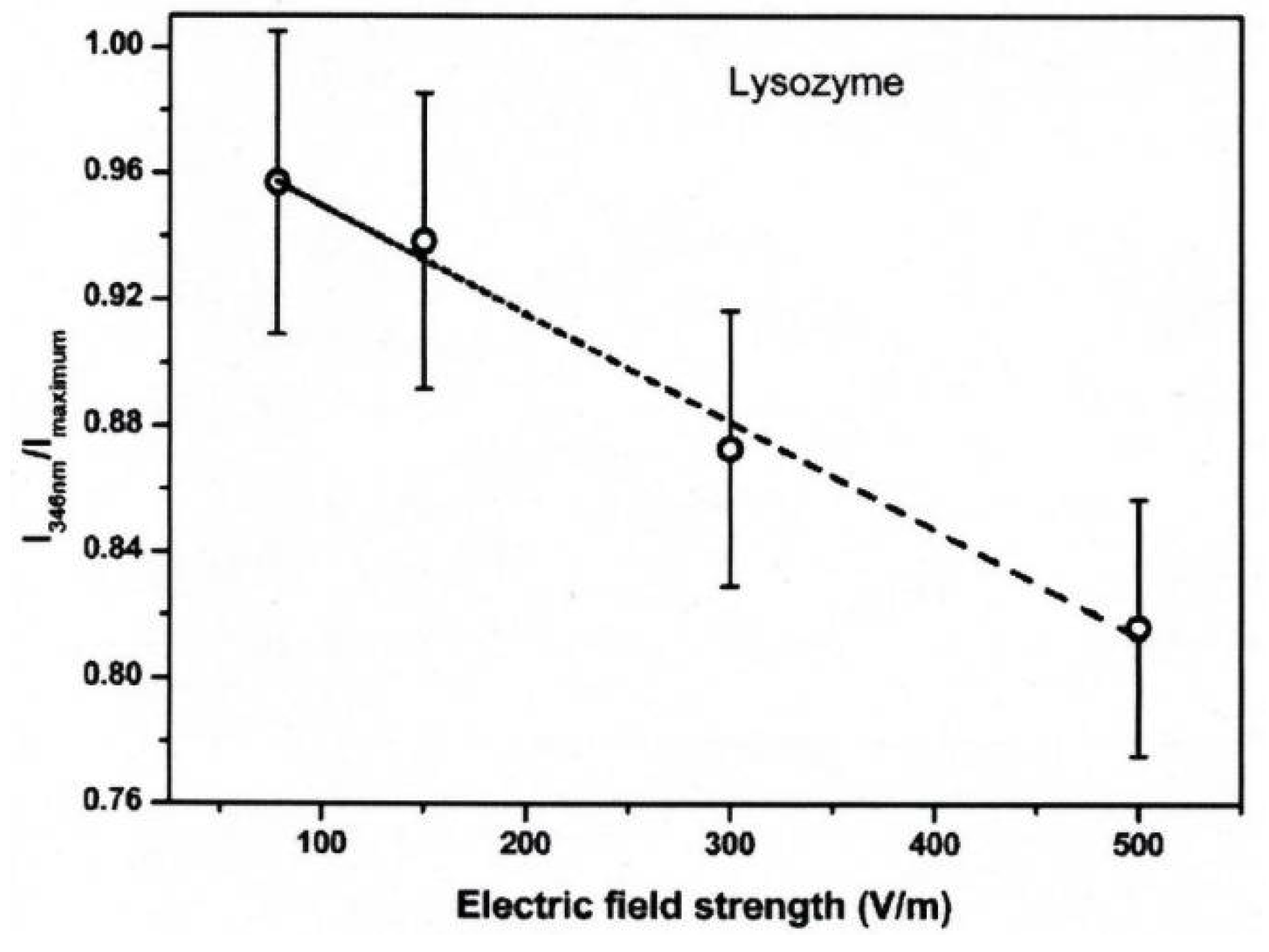
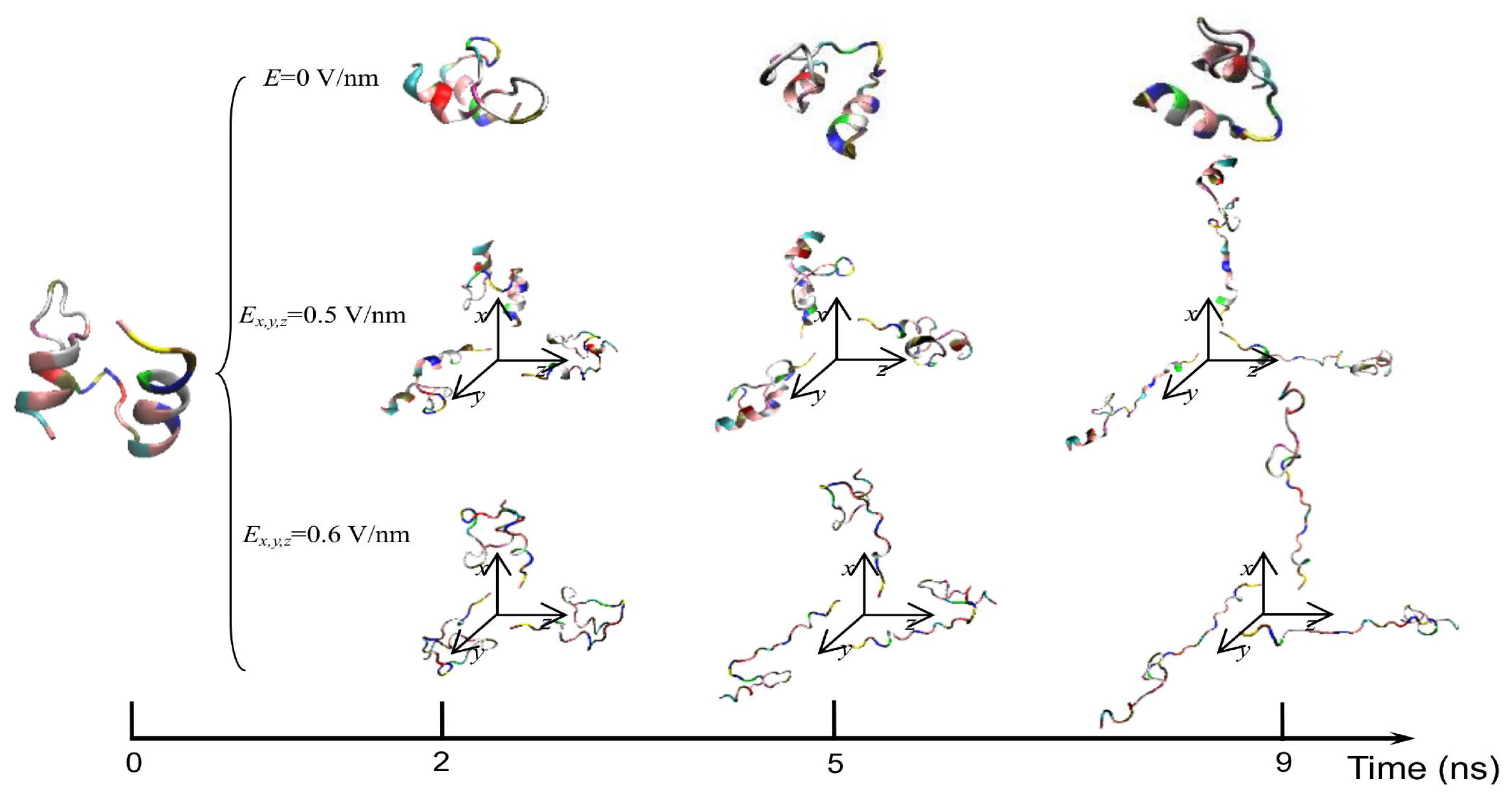
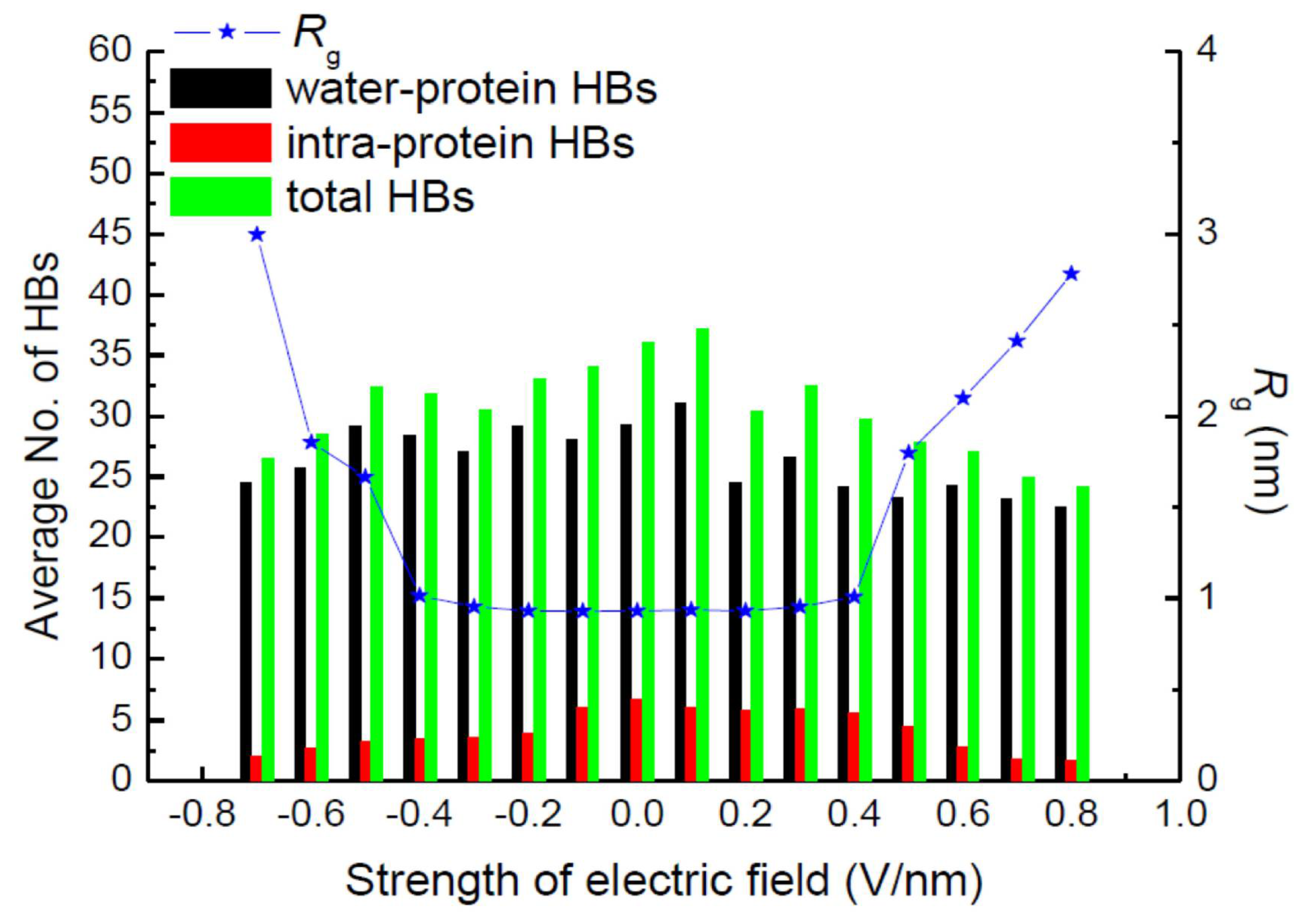
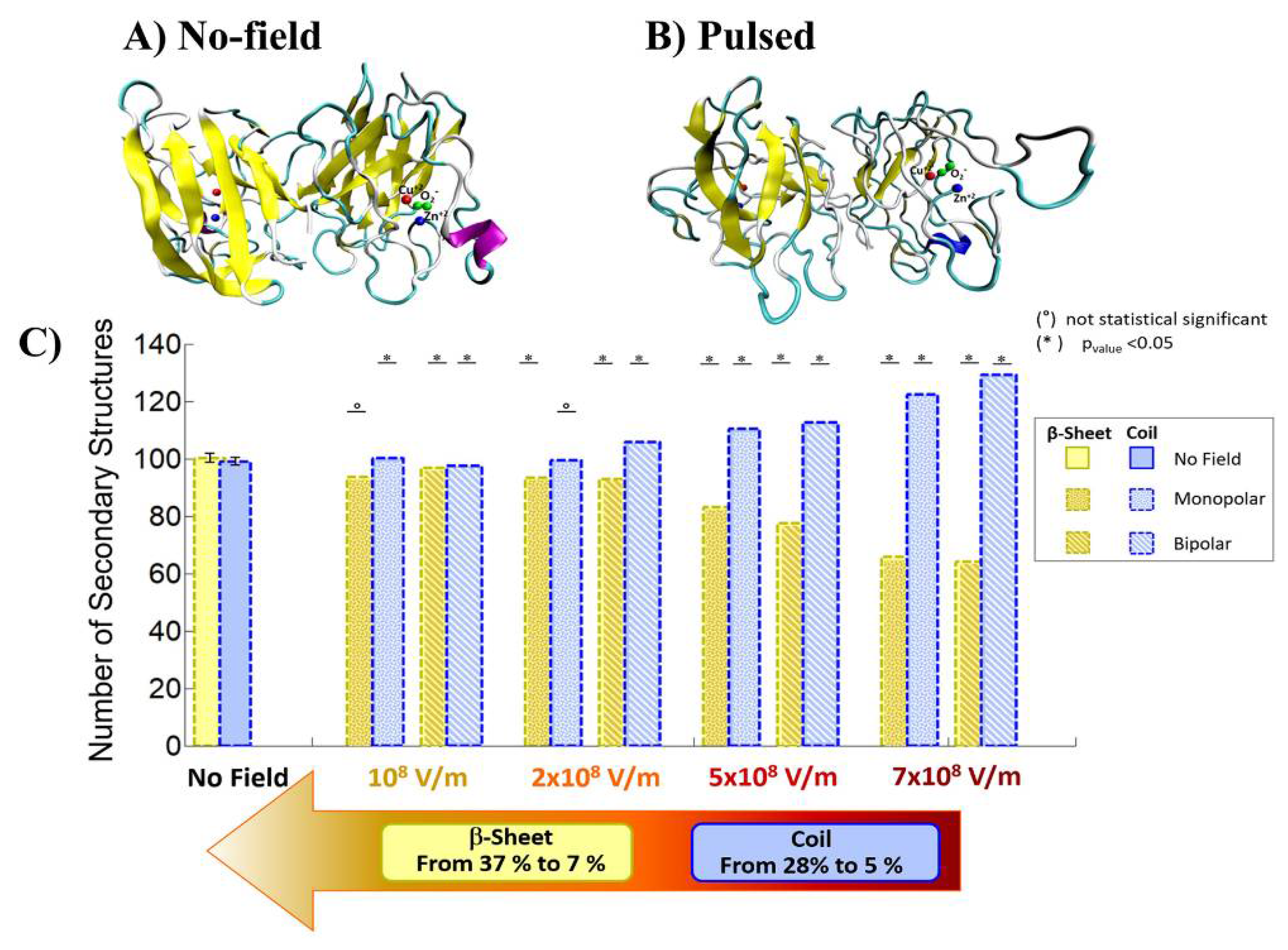
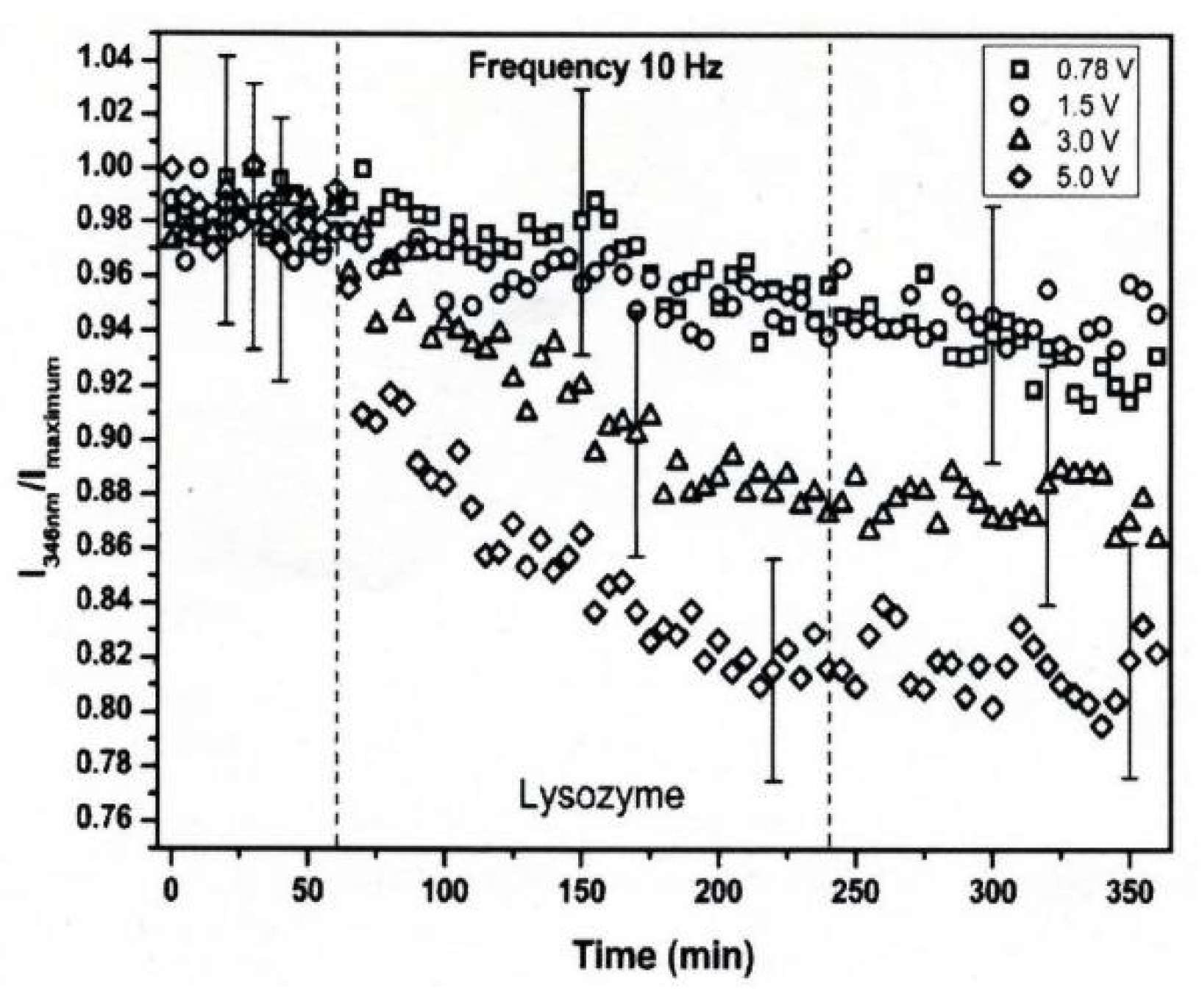
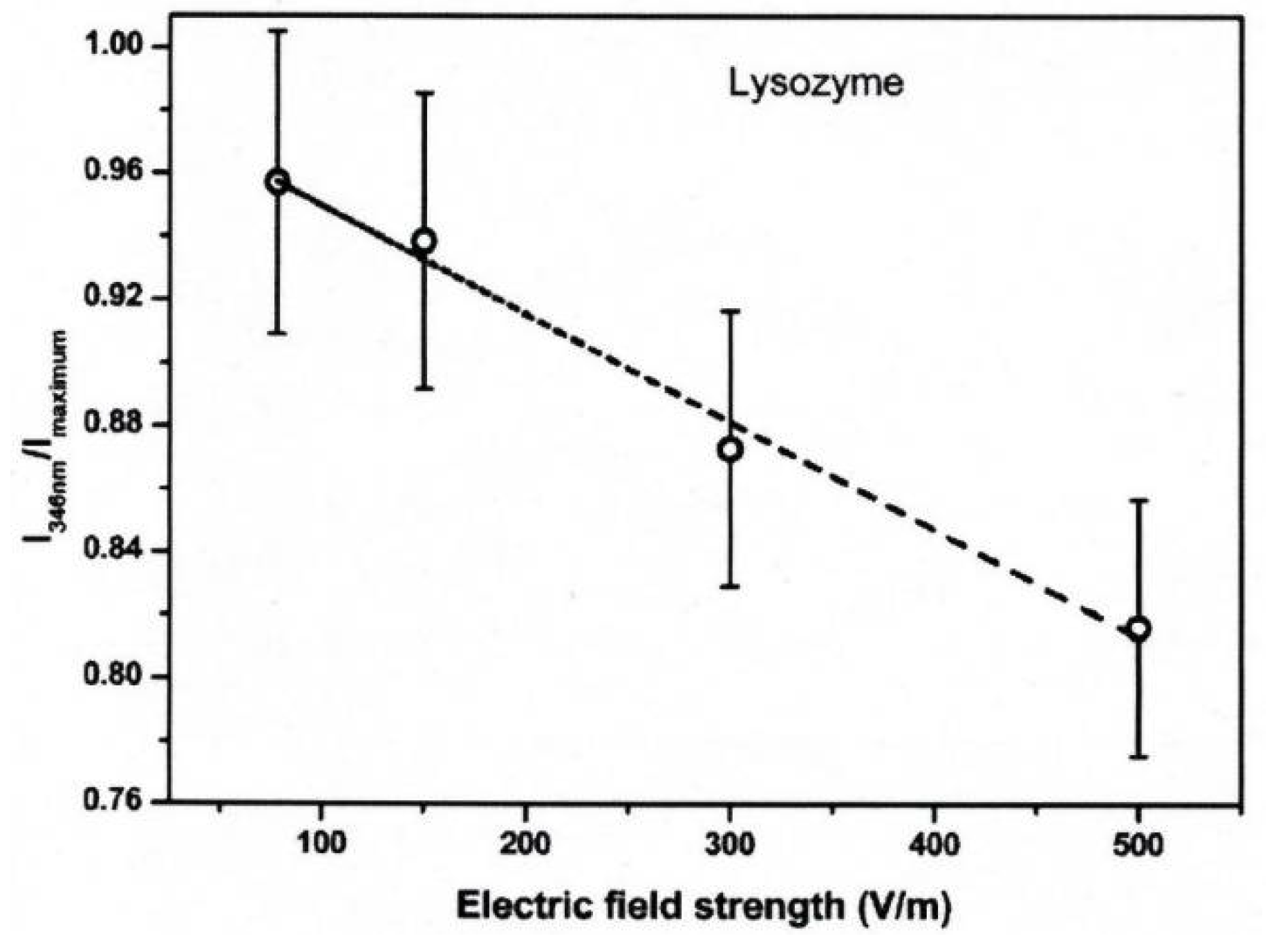
3. Protein Conformational Change by PEF Irradiation
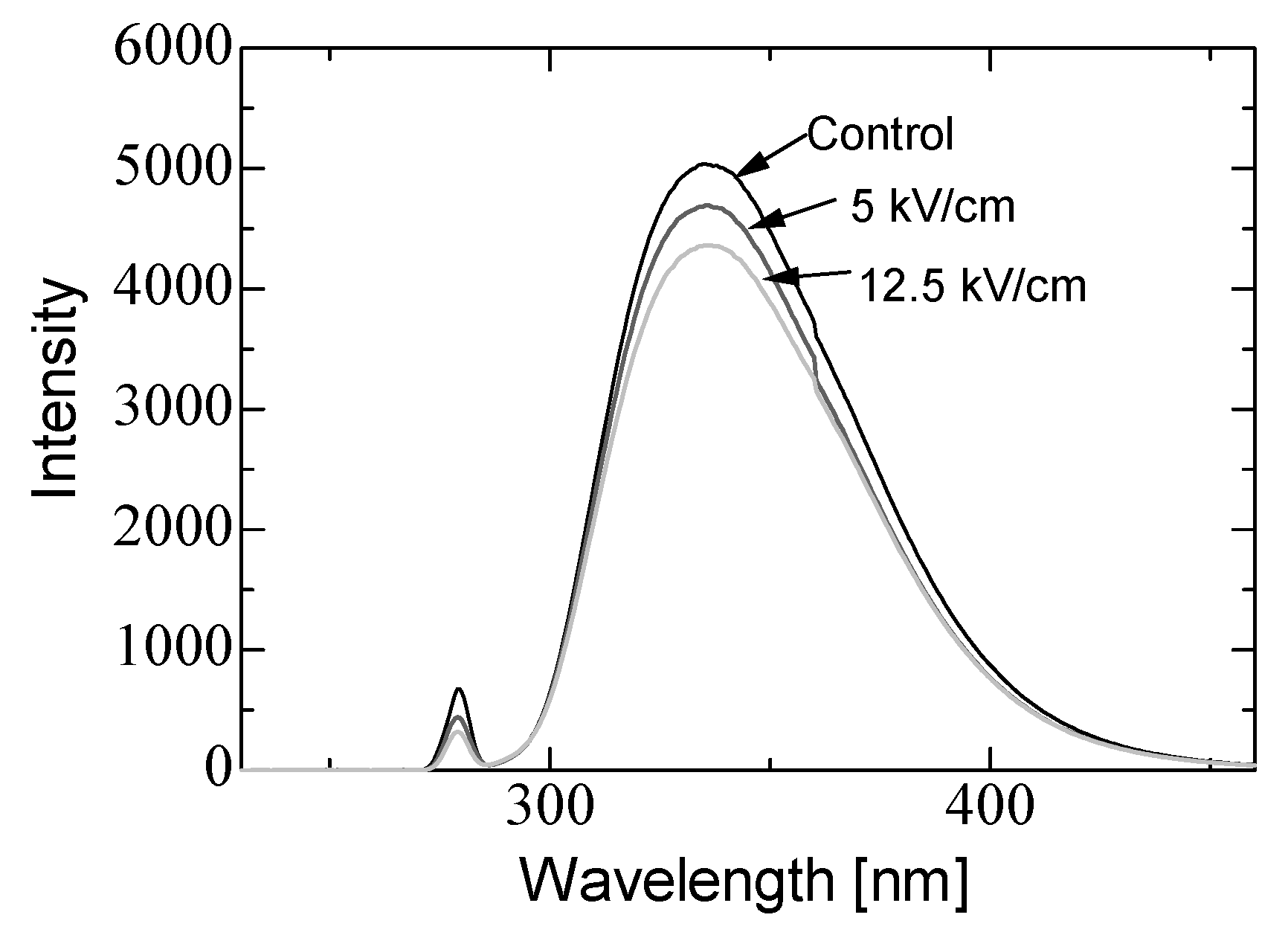
3.1. Conformational Change in Proteins
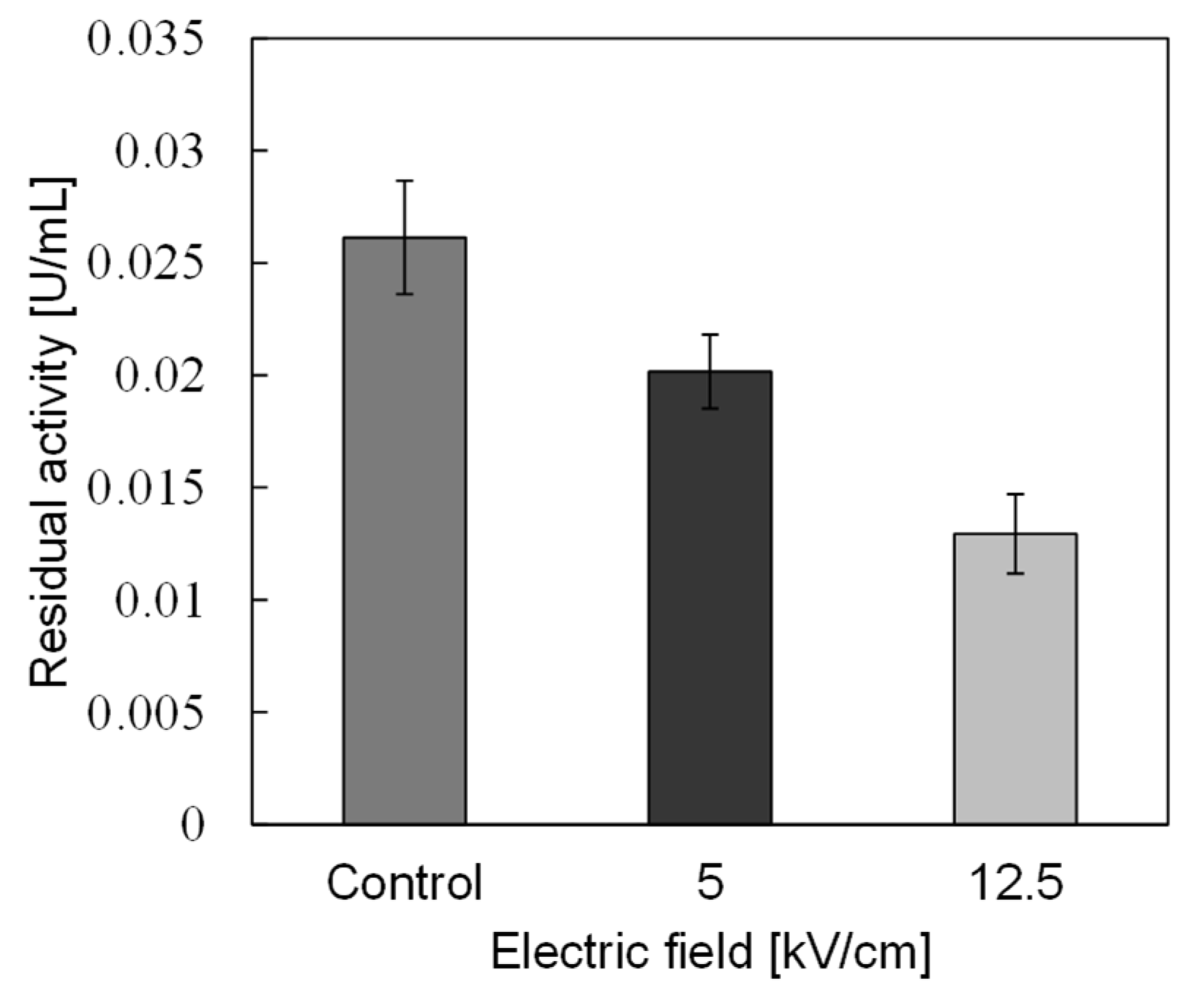
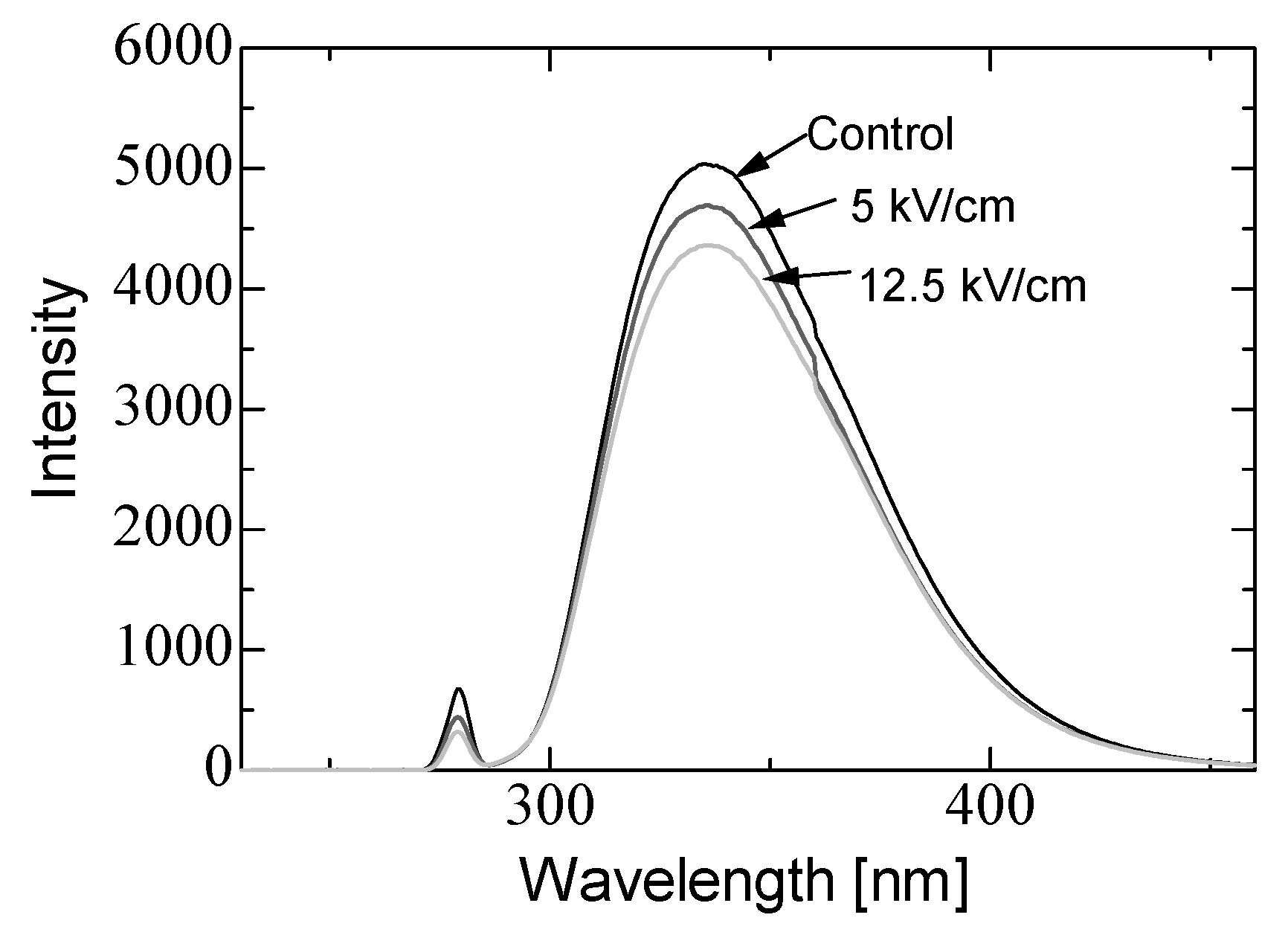
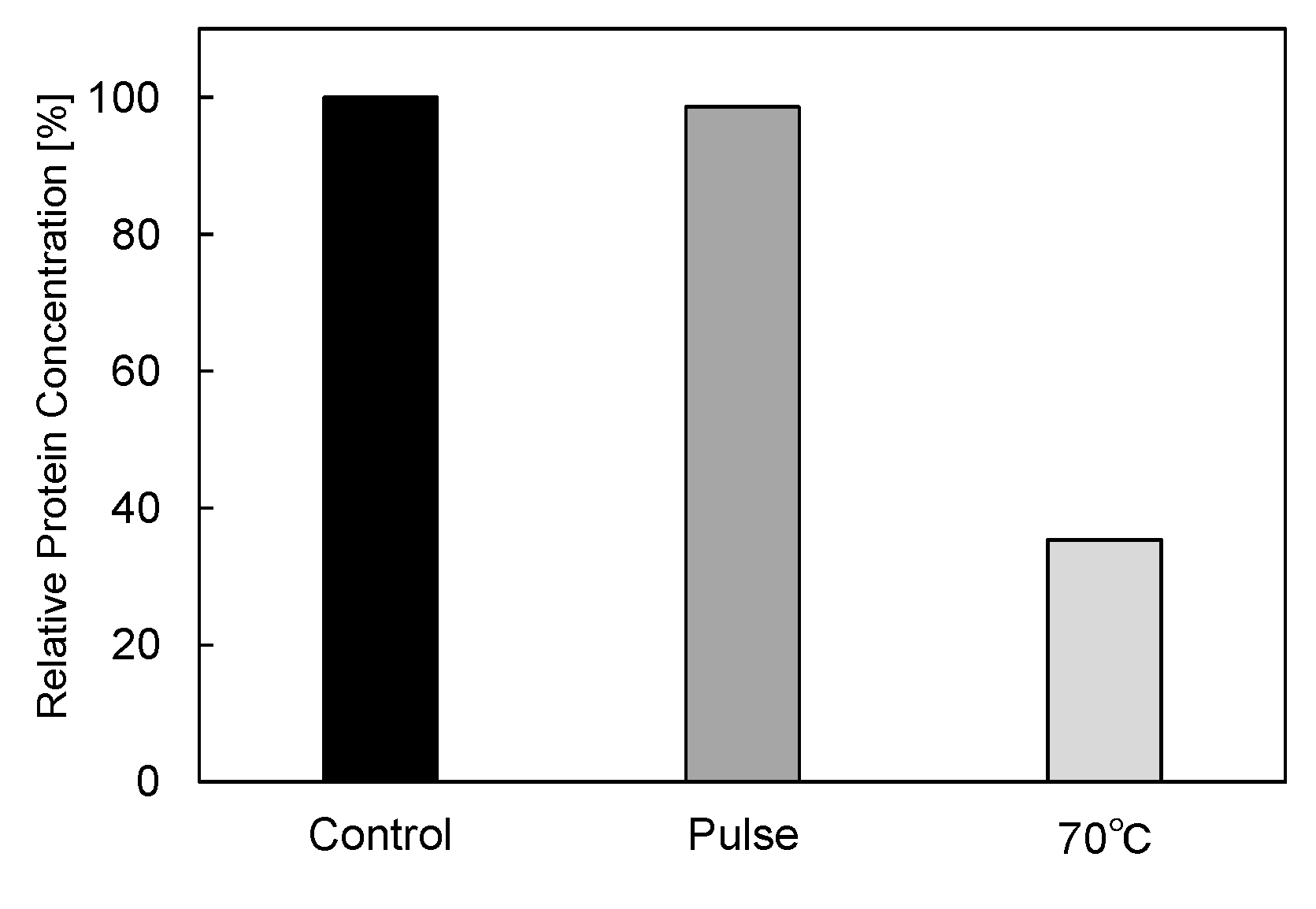
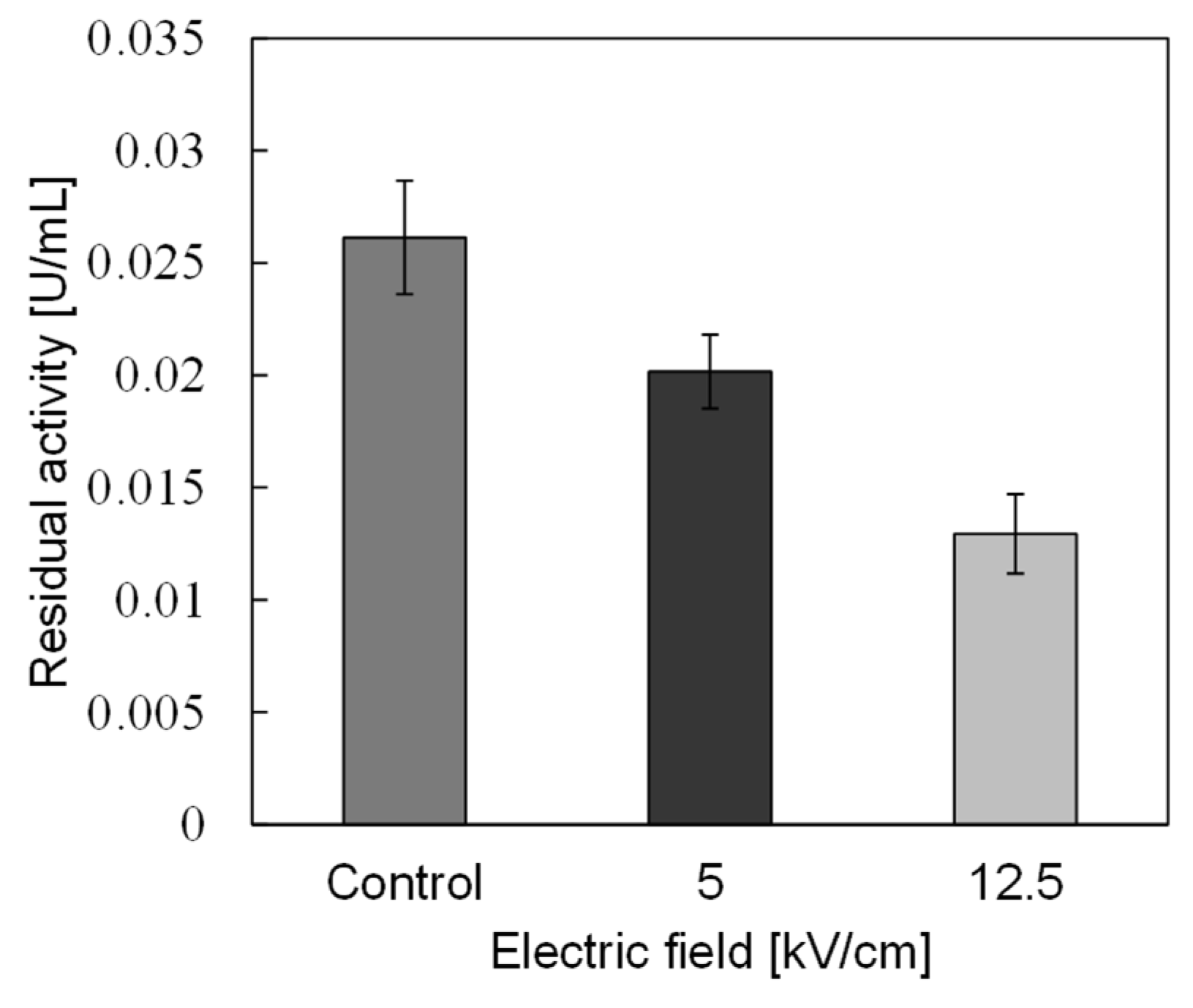
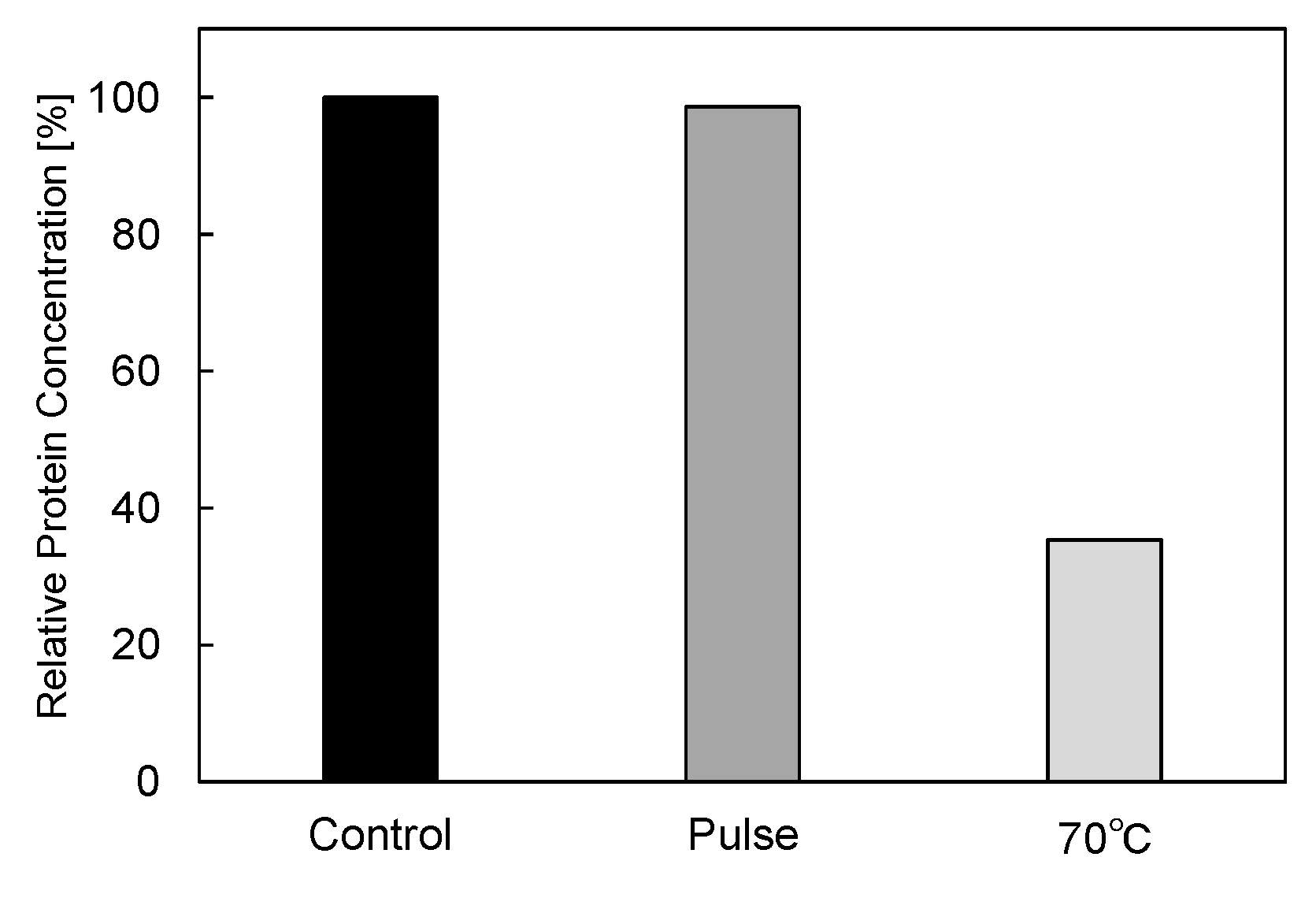
3.2. PEF Treatment for α-Amylase Inactivation via Conformational Changes
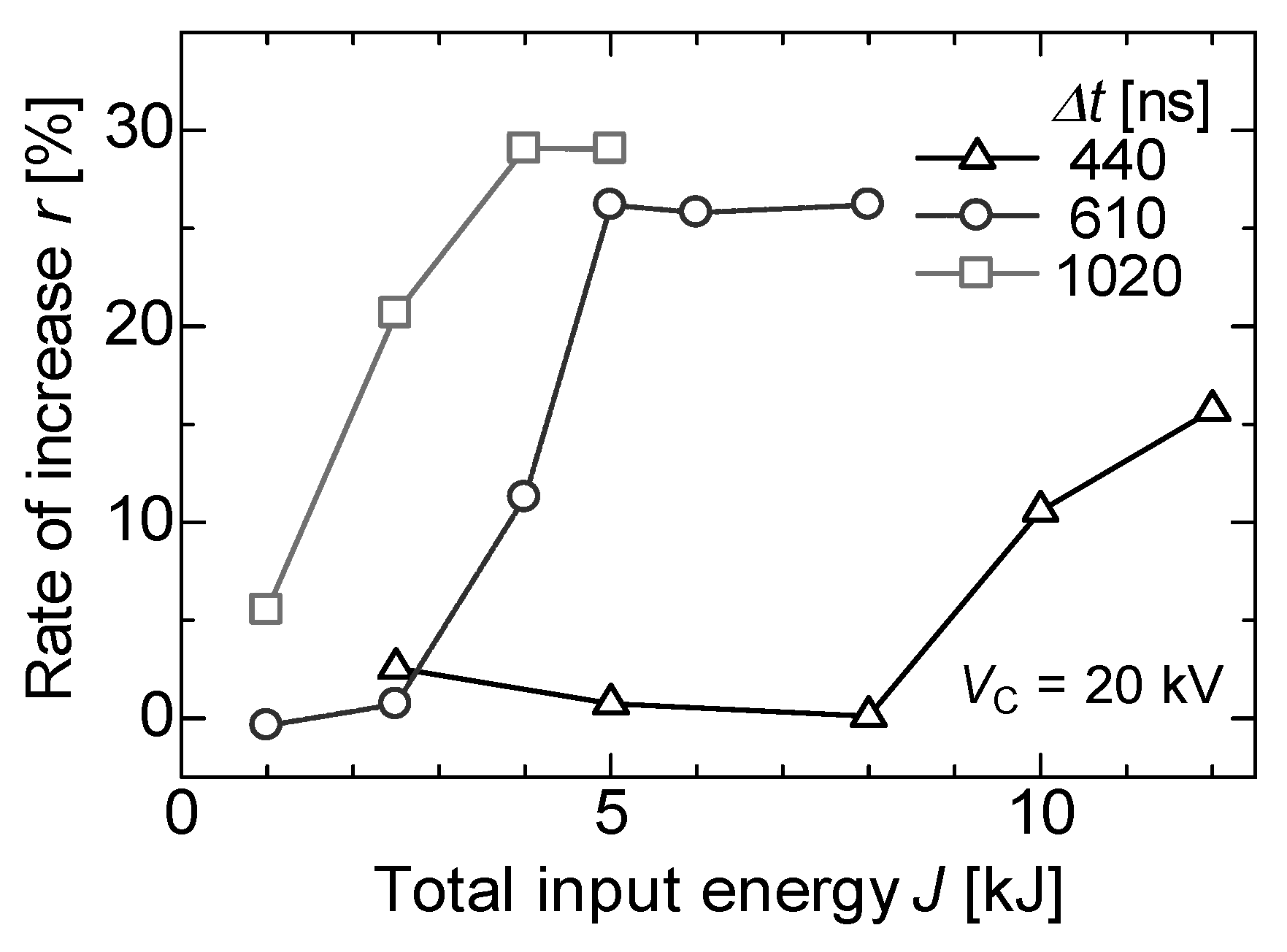
3.3. Enzyme Inactivation by a PEF under Various Conditions
4. PEF Poration Process of Cell Membranes and Its Applications
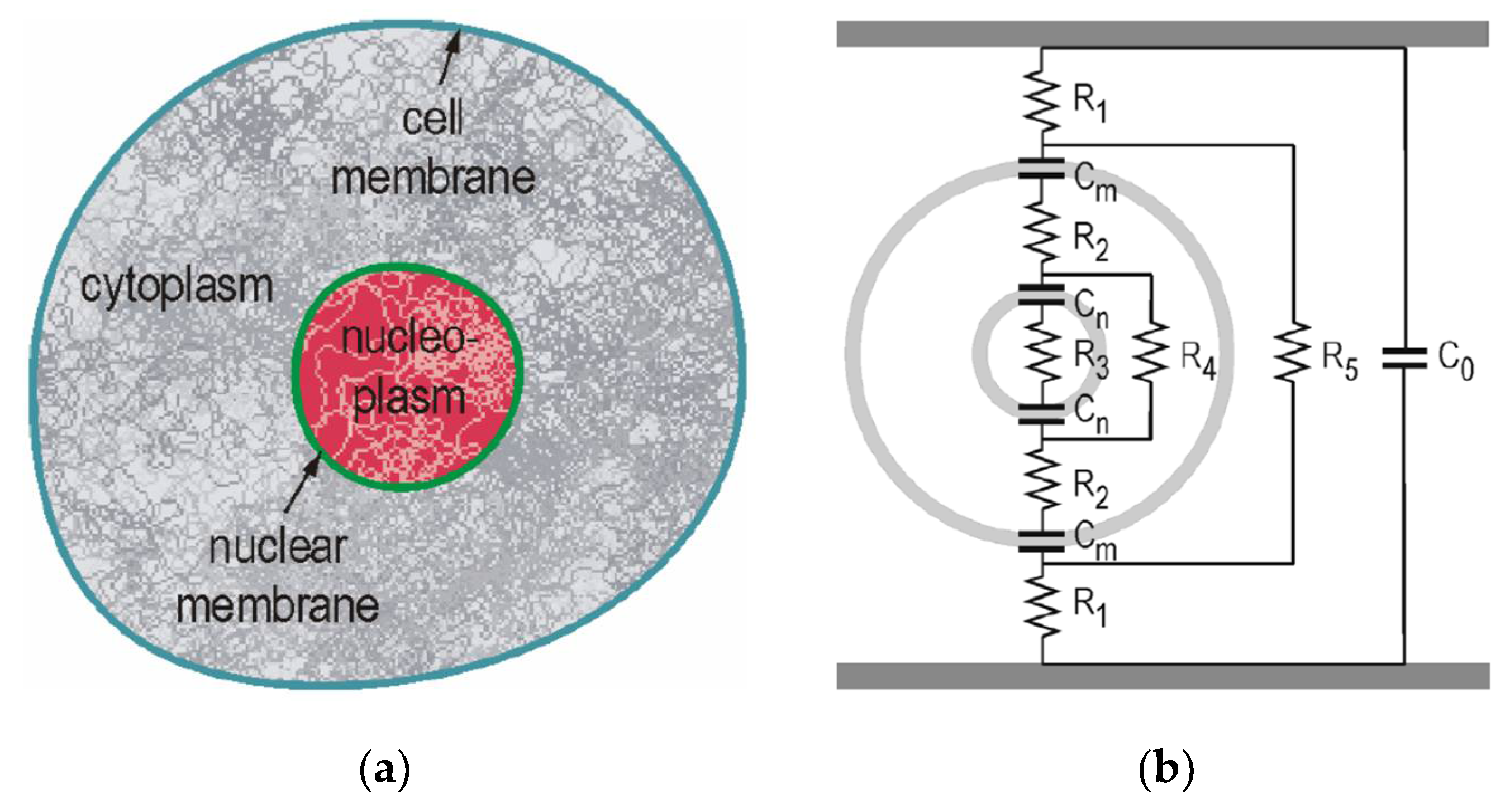
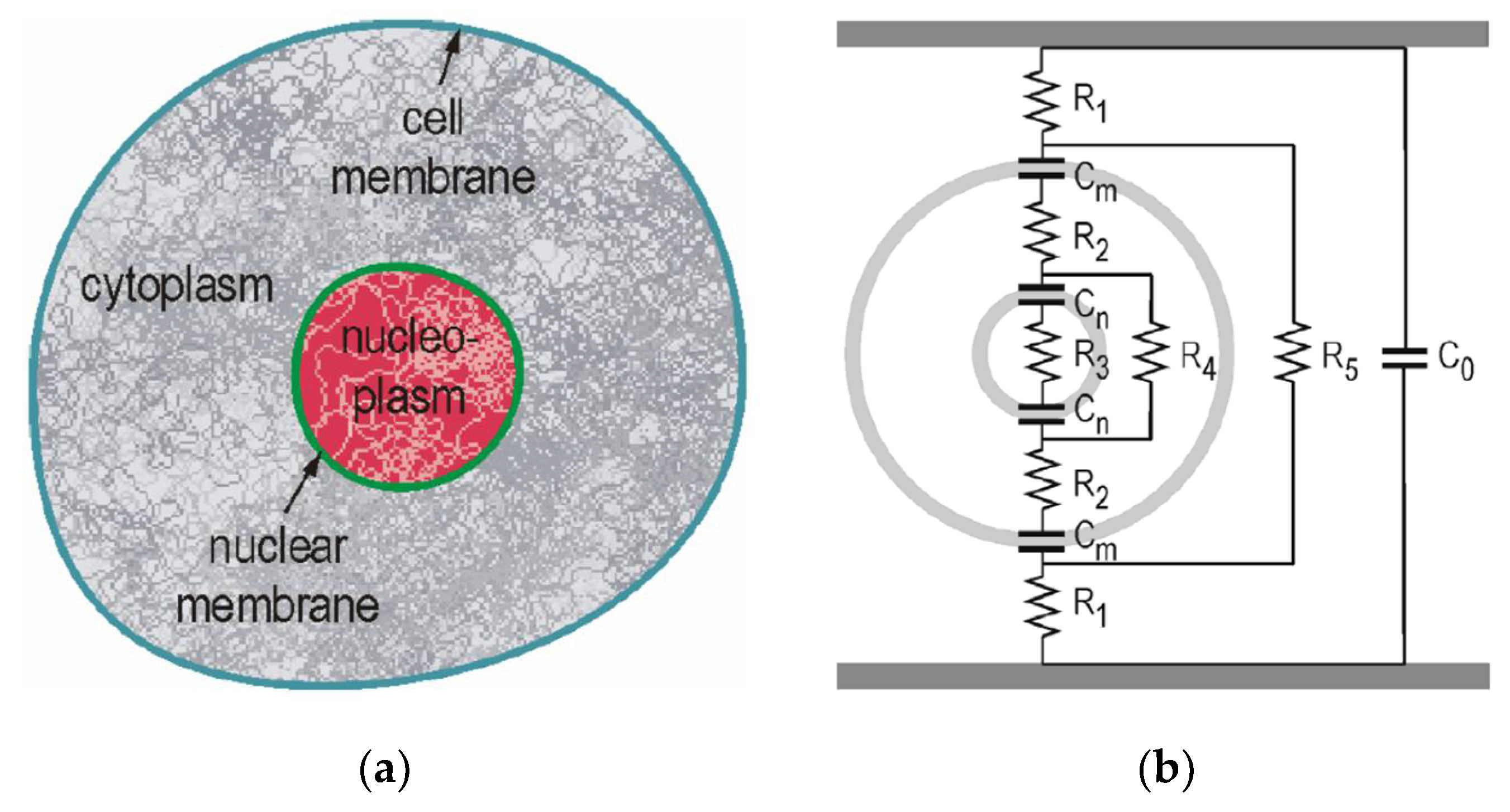
4.1. Voltage Buildup across the Cell Membrane
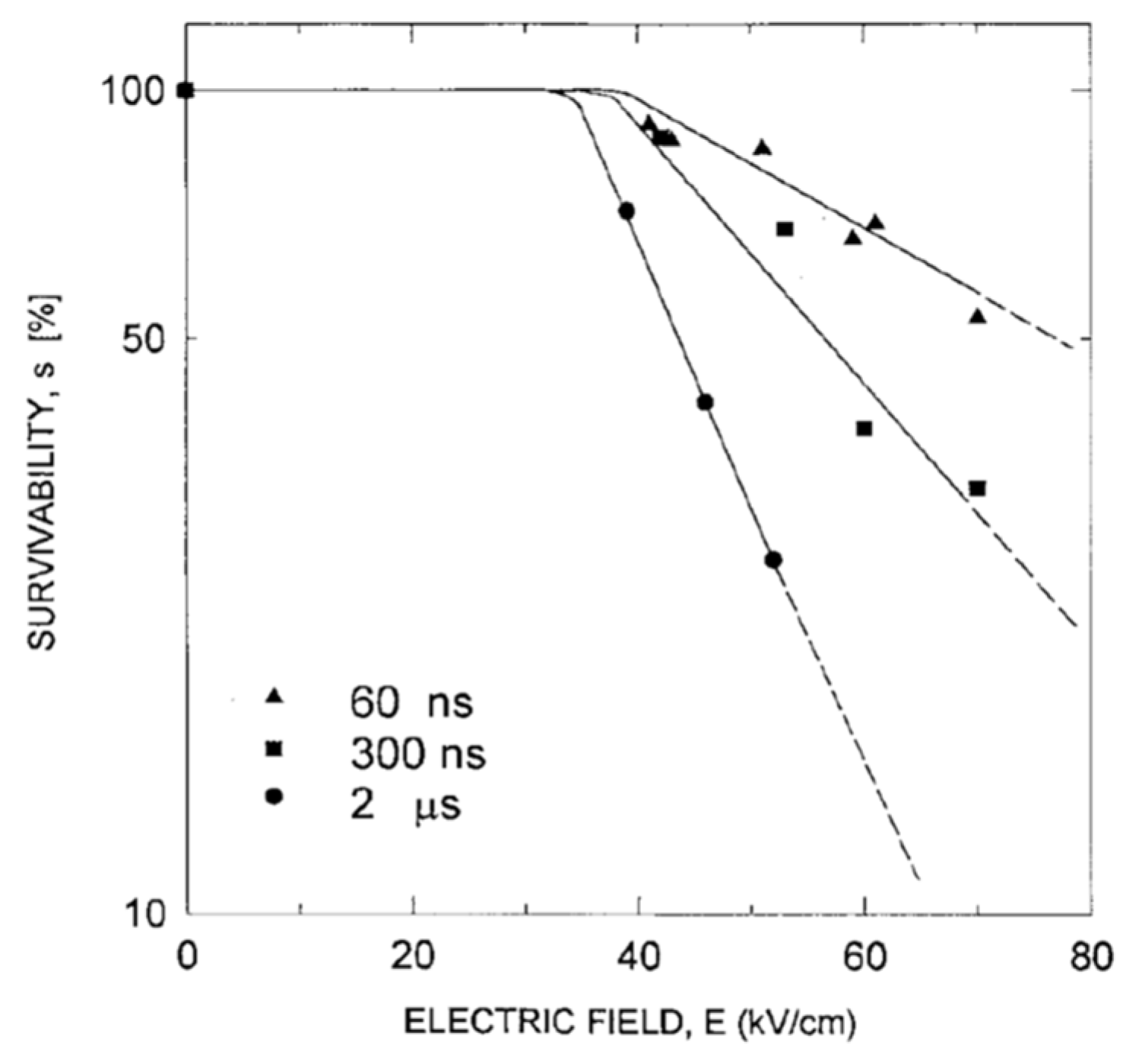
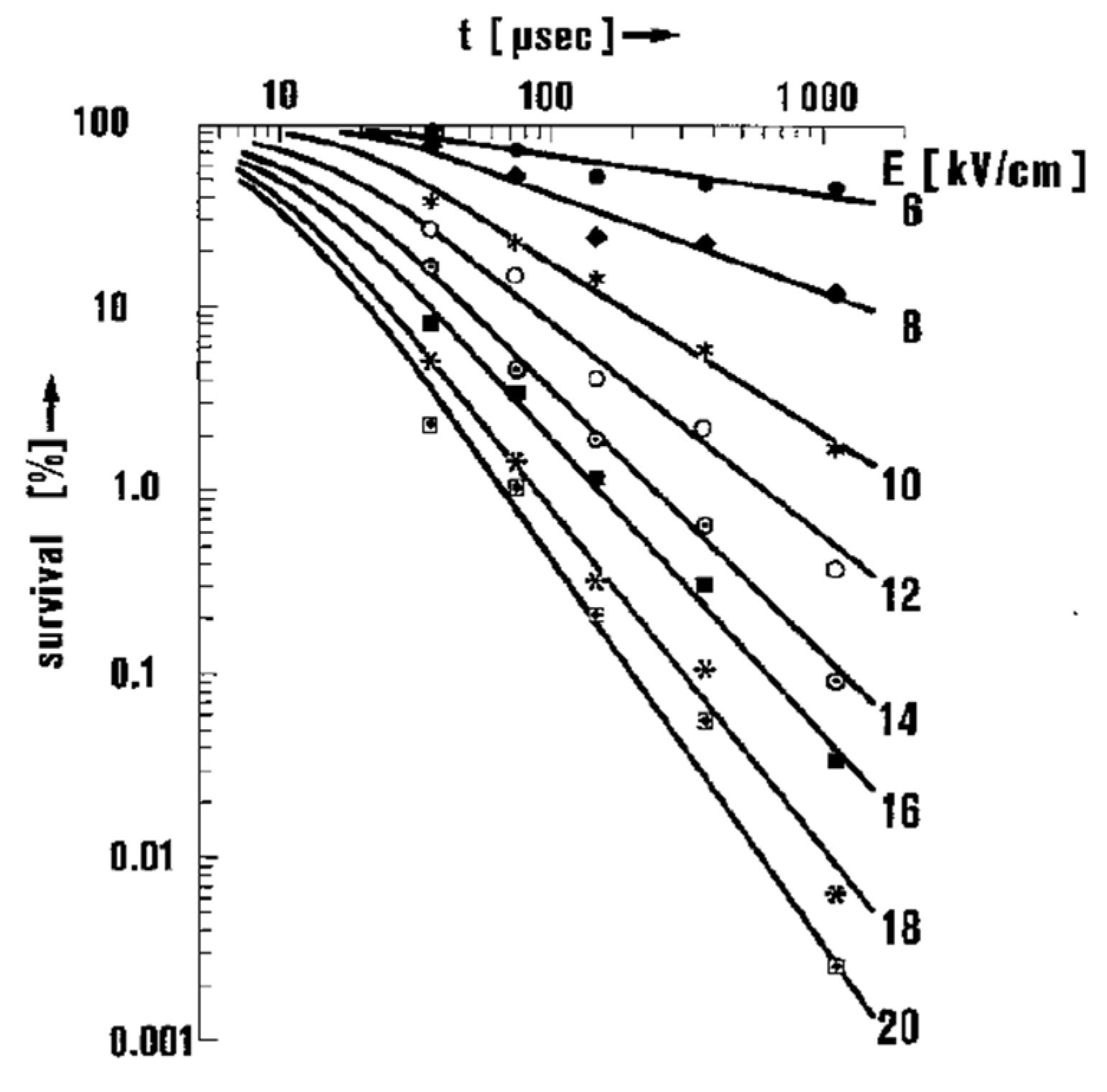
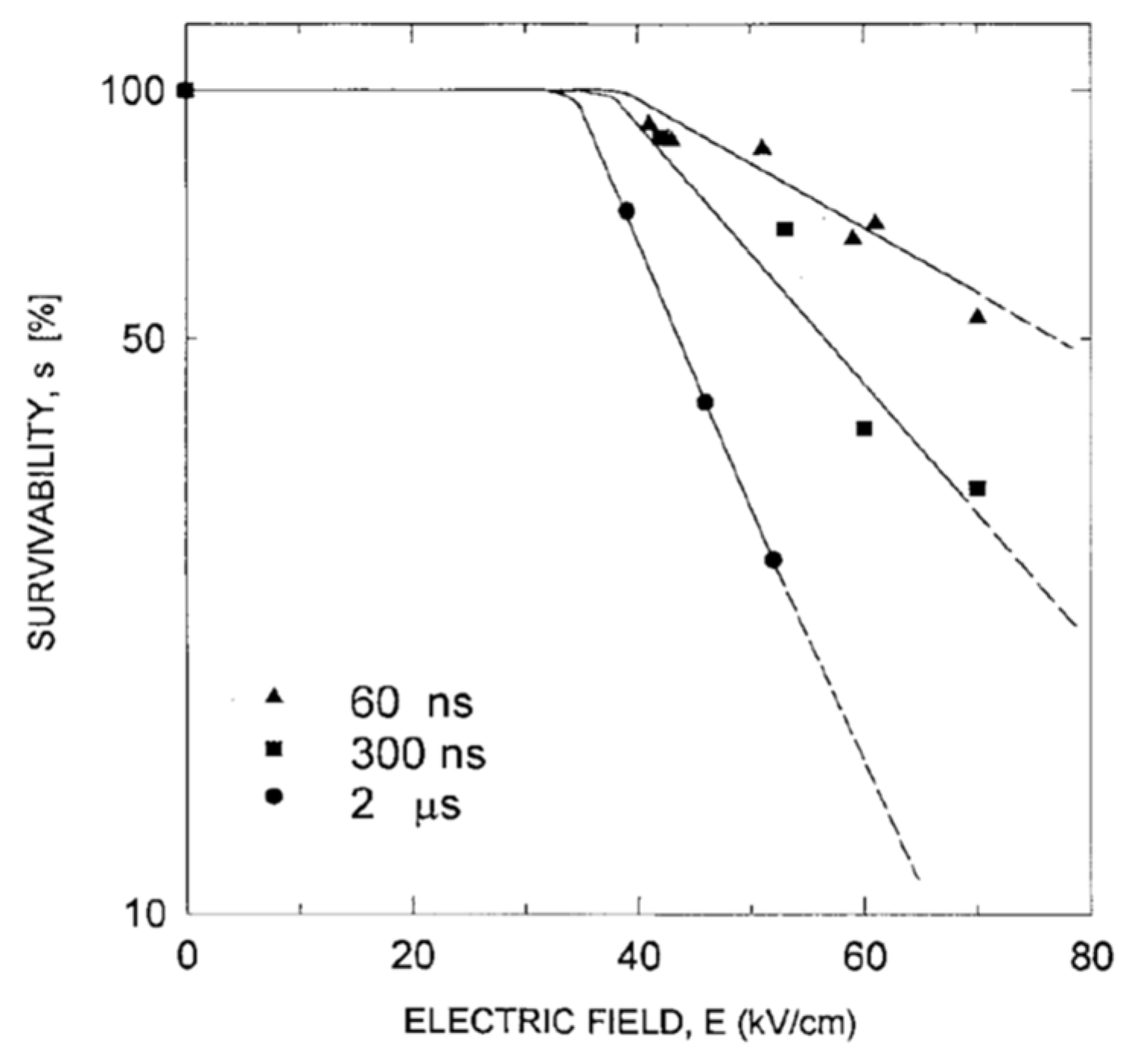
4.2. Critical PEF Strength for Microorganism Survivability
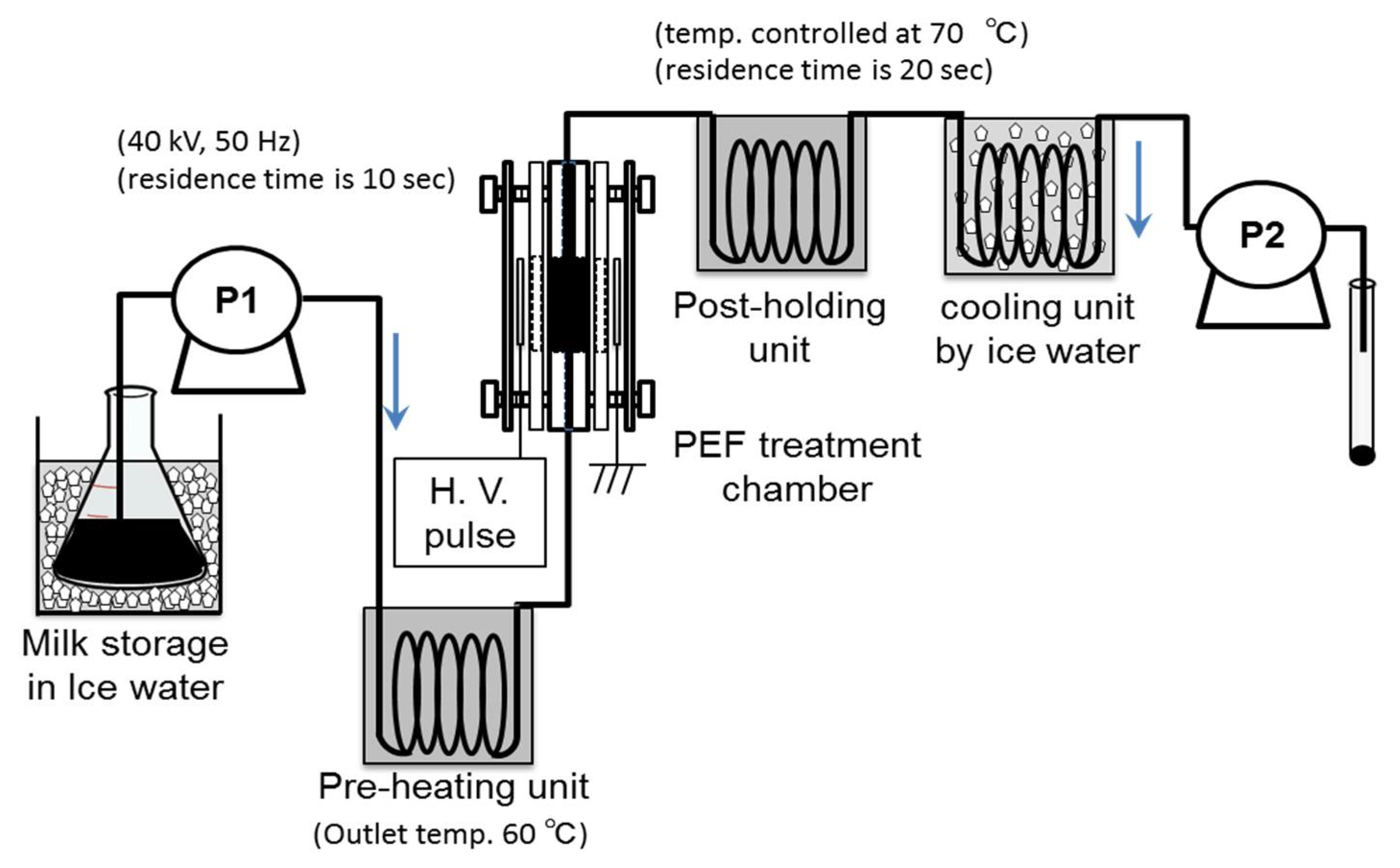
4.3. PEF Pasteurization
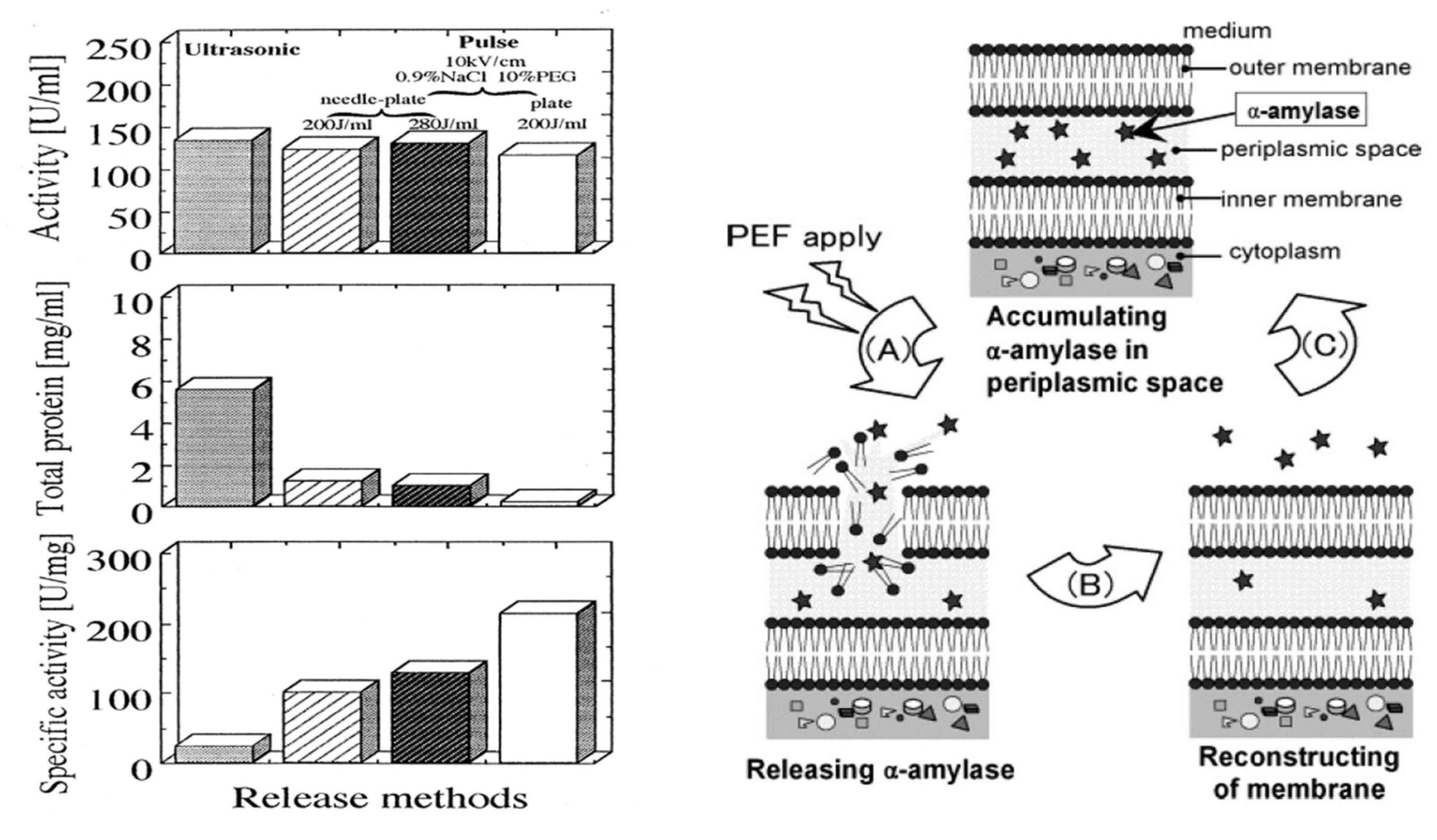
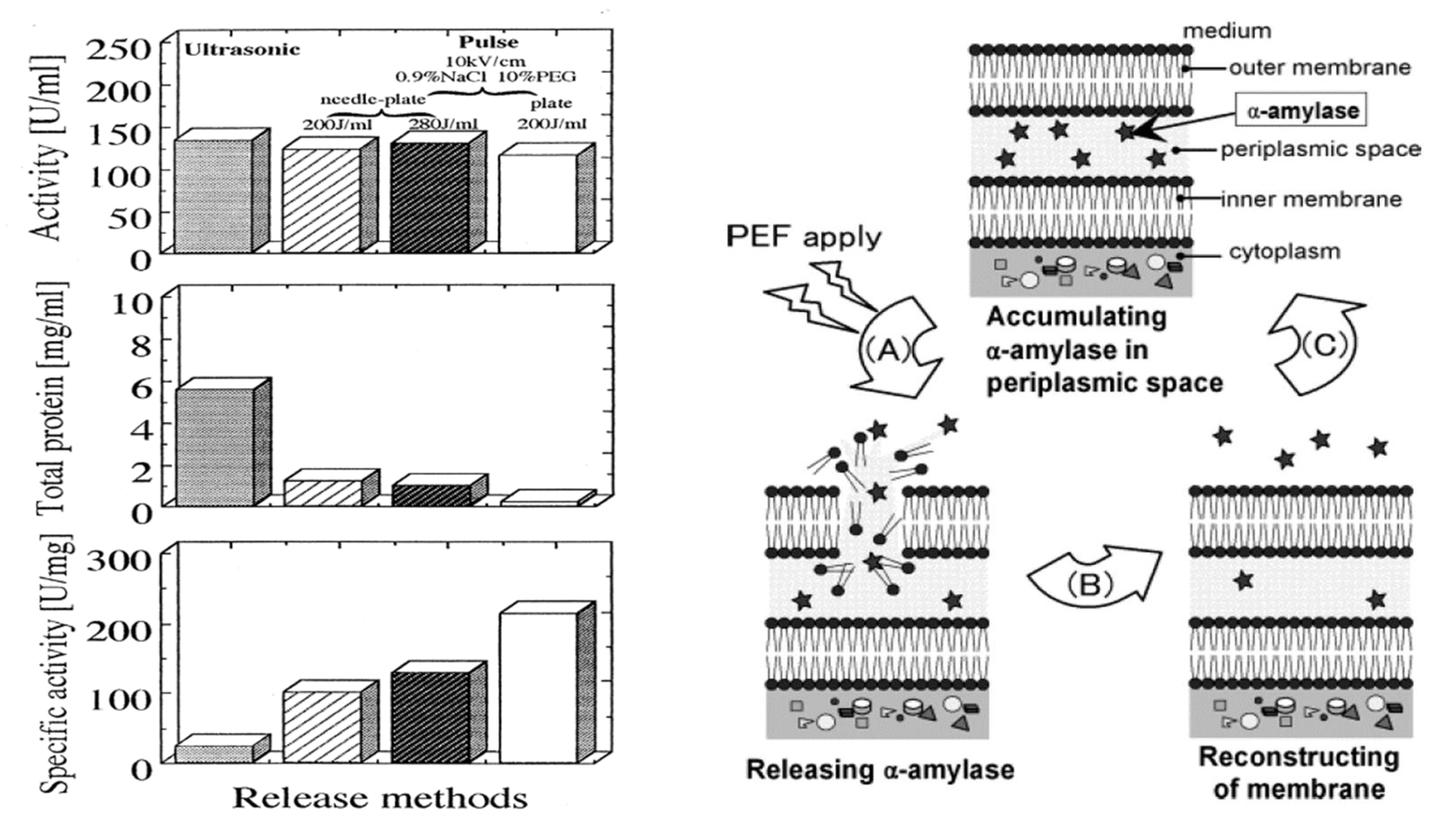
4.4. PEF Extraction of Intracellular Contents
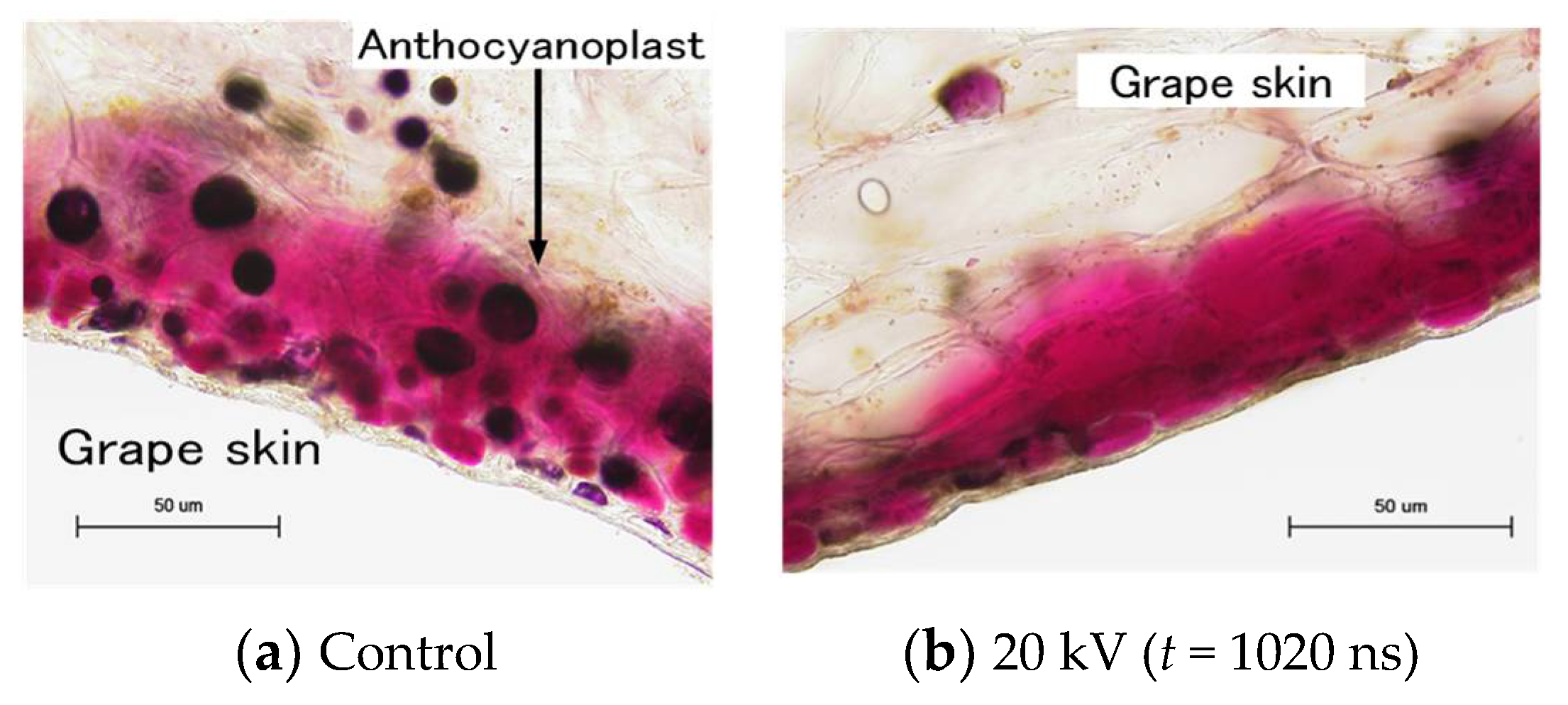
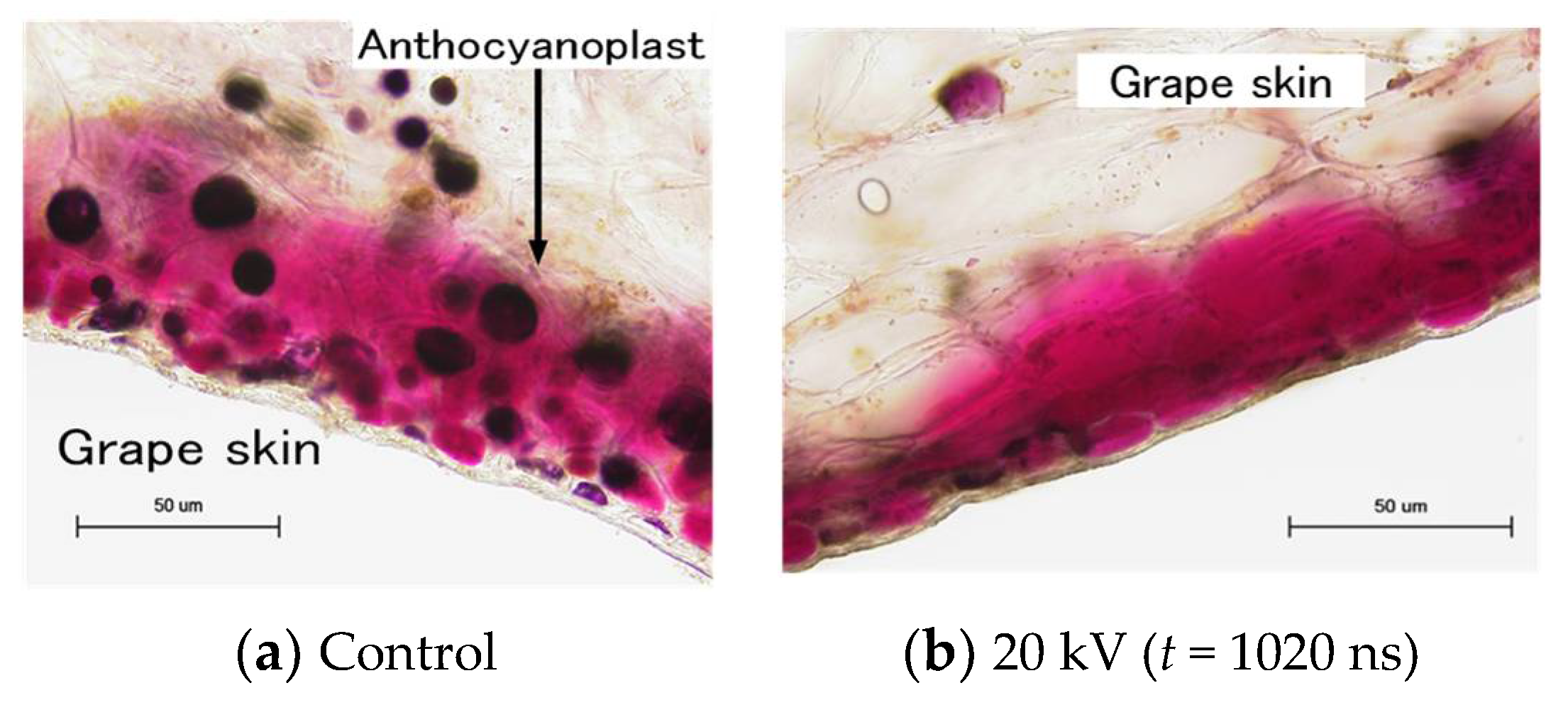
4.5. PEF Pre-Treatment for Food Drying Processes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Takaki, K.; Hayashi, N.; Wang, D.; Ohshima, T. High-voltage technologies for agriculture and food processing. J. Phys. D Appl. Phys. 2019, 52, 473001. [Google Scholar] [CrossRef]
- Yano, K.; Rems, L.; Kotnik, T.; Miklavčič, D.; Weaver, J.C.; Smith, K.C.; Son, R.S.; Gowrishankar, T.R.; Vernier, P.T.; Levine, Z.A.; et al. Biological Responses. In Bioelectrics, 1st ed.; Akiyama, H., Heller, R., Eds.; Springer: Tokyo, Japan, 2017; pp. 155–274. [Google Scholar]
- Graves, D.B. The emerging role of reactive oxygen and nitrogen species in redox biology and some implications for plasma applications to medicine and biology. J. Phys. D Appl. Phys. 2012, 45, 263001. [Google Scholar] [CrossRef]
- Akiyama, H.; Katsuki, S.; Redondo, L.; Akiyama, M.; Pemen, A.J.M.; Huiskamp, T.; Beckers, F.J.C.M.; Heesch, E.J.M.; Winands, G.J.J.; Voeten, S.J.; et al. Pulsed Power Technology. In Bioelectrics, 1st ed.; Akiyama, H., Heller, R., Eds.; Springer: Tokyo, Japan, 2017; pp. 41–107. [Google Scholar]
- Zimmermann, U. Electric field-mediated fusion and related electrical phenomena. Biochem. Biophys. Acta 1982, 64, 227–277. [Google Scholar] [CrossRef]
- Guionet, A.; Fujiwara, T.; Sato, H.; Takahashi, K.; Takaki, K.; Matsui, M.; Tanino, T.; Ohshima, T. Pulsed electric fields act on tryptophan to inactivate α-amylase. J. Electrost. 2021, 112, 103597. [Google Scholar] [CrossRef]
- Yang, N.; Huang, K.; Lyu, C.; Wang, J. Pulsed electric field technology in the manufacturing processes of wine, beer, and rice wine: A review. Food Control 2016, 61, 28–38. [Google Scholar] [CrossRef]
- Ohshima, T.; Tamura, T.; Sato, M. Influence of pulsed electric field on various enzyme activities. J. Electrost. 2007, 65, 156–161. [Google Scholar] [CrossRef]
- Ohshima, T.; Tanino, T.; Guionet, A.; Takahashi, K.; Takaki, K. Mechanism of pulsed electric field enzyme activity change and pulsed discharge permeabilization of agricultural products. Jpn. J. Appl. Phys. 2021, 60, 060501. [Google Scholar] [CrossRef]
- Weltmann, K.; Kindel, E.; Woedtke, T. Atmospheric pressure plasma sources: Prospective tools for plasma medicine. Pure Appl. Chem. 2010, 82, 1223–1237. [Google Scholar] [CrossRef]
- Bluhm, H. Pulsed Power System, 1st ed.; Springer: Berlin, Germany, 2006; pp. 1–6. [Google Scholar]
- Lehr, J.; Rpn, P. Foundations of Pulsed Power Technology, 1st ed.; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. xxv–xxxi. [Google Scholar]
- Kuffel, E.; Zaengl, W.S.; Kuffel, D. High Voltage Engineering: Fundamentals, 2nd ed.; Elsevier: Oxford, UK, 2000; pp. 281–366. [Google Scholar]
- Grahl, T.; Markl, H. Killing of microorganisms by pulsed electric fields. Appl. Microbiol. Biotechnol. 1996, 45, 148–157. [Google Scholar] [CrossRef]
- Miura, T.; Sato, T.; Arima, K.; Mukaigawa, S.; Takaki, K.; Fujiwara, T. Duty Factor Effect on Ozone Production Using Dielectric Barrier Discharge Reactor Driven by IGBT Pulse Modulator. J. Adv. Oxid. Technol. 2007, 10, 311–315. [Google Scholar] [CrossRef]
- Takaki, K.; Kanesawa, K.; Mukaigawa, S.; Fujiwara, T. Energy Efficiency of Corona Discharge Reactor Driven by Inductive Energy Storage System Pulsed Power Generator. IEEE Trans. Dielectr. Electr. Insul. 2007, 14, 834–845. [Google Scholar] [CrossRef]
- Takaki, K.; Takahashi, K.; Ueno, T.; Akiyama, M.; Sakugawa, T. Design and Practice of Pulsed Power Circuit. J. Plasma Fusion Res. 2011, 87, 202–215. [Google Scholar]
- Takahashi, K.; Yagi, I.; Takaki, K.; Satta, N. Development of Pulsed Discharge Inside Bubble in Water. IEEE Trans. Plasma Sci. 2011, 39, 2654–2655. [Google Scholar] [CrossRef]
- Rukin, S.N. High-Power Nanosecond Pulse Generators Based on Semiconductor Opening Switches. Instrum. Exp. Tech. 1999, 42, 439–467. [Google Scholar]
- Tendero, C.; Tixier, C.; Tristant, P.; Desmaison, J.; Leprince, P. Atmospheric pressure plasmas: A review. Spectrochim. Acta B 2006, 61, 2–30. [Google Scholar] [CrossRef]
- Takahashi, K.; Saito, R.; Onodera, T.; Takaki, K.; Kitai, H.; Sakamoto, K. Development of compact inductive energy storage pulsed-power generator driven by 13 kV SiC-MOSFET. Rev. Sci. Instrum. 2021, 92, 064706. [Google Scholar] [CrossRef]
- Takaki, K.; Kanesawa, K.; Yamazaki, N.; Mukaigawa, S.; Fujiwara, T.; Takahasi, K.; Yamasita, K.; Nagane, K. Improvement of edible mushroom yield by electric stimulations. J. Plasma Fusion Res. Ser. 2009, 8, 556–559. [Google Scholar]
- Takahashi, K.; Miyamoto, K.; Takaki, K.; Takahashi, K. Development of Compact High-Voltage Power Supply for Stimulation to Promote Fruiting Body Formation in Mushroom Cultivation. Materials 2018, 11, 2471. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Namihira, T.; Sakugawa, T.; Katsuki, S.; Akiyama, H.K. Loss Characteristics of a Magnetic Core for Pulsed Power Applications. IEEE Trans. Plasma Sci. 2007, 35, 1791–1796. [Google Scholar] [CrossRef]
- Huiskamp, T. Nanosecond pulsed streamer discharges Part I: Generation, source-plasma interaction and energy-efficiency optimization. Plasma Sources Sci. Technol. 2020, 29, 023002. [Google Scholar] [CrossRef]
- Namihira, T.; Tsukamoto, S.; Wang, D.; Katsuki, S.; Hackam, R.; Akiyama, H.; Uchida, Y.; Koike, M. Improvement of NOX removal efficiency using short width pulsed power. IEEE Trans. Plasma Sci. 2000, 28, 434–442. [Google Scholar] [CrossRef]
- Syed, Q.A.; Ishaq, A.; Rahman, U.U.; Aslam, S.; Shukat, R. Pulsed electric field technology in food preservation: A review. J. Nutr. Health Food Eng. 2017, 6, 168–172. [Google Scholar]
- Mohamed, M.E.A.; Eissa, A.H.A. Pulsed electric fields for food processing technology. In Structure and Function of Food Engineering; Eissa, A.A., Ed.; Intech Open: London, UK, 2012; Chapter 11; pp. 275–304. [Google Scholar]
- Bekard, I.; Dunstan, D.E. Electric field induced changes in protein conformation. Soft Matter 2014, 10, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; You, L.; Dou, W.; Sun, T.; Xu, P. Effects of an Electric Field on the Conformational Transition of the Protein: A Molecular Dynamics Simulation Study. Polymers 2019, 11, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Z.; Buehler, M.J. Molecular Dynamics Simulation of the α-Helix to β-Sheet Transition in Coiled Protein Filaments: Evidence for a Critical Filament Length Scale. Phys. Rev. Lett. 2010, 104, 198304. [Google Scholar] [CrossRef] [PubMed]
- Marracino, P.; Paffi, A.; Reale, R.; Liberti, M.; d’Inzeo, G.; Apollonio, F. Technology of High–Intensity Electric–Field Pulses: A Way to Control Protein Unfolding. J. Phys. Chem. Biophys. 2013, 3, 1000117. [Google Scholar]
- Valle, E.D.; Marracino, P.; Pakhomova, O.; Liberti, M.; Apollonio, F. Nanosecond pulsed electric signals can affect electrostatic environment of proteins below the threshold of conformational effects: The case study of SOD1 with a molecular simulation study. PLoS ONE 2019, 14, e0221685. [Google Scholar]
- Sheu, S.-Y.; Yang, D.-Y.; Selzle, H.L.; Schlag, E.W. Energetics of hydrogen bonds in peptides. Proc. Natl. Acad. Sci. USA 2003, 100, 12683–12687. [Google Scholar] [CrossRef] [Green Version]
- Ding, F.; Borreguero, J.M.; Buldyrey, S.V.; Stanley, H.E.; Dokholyan, N.V. Mechanism for the α-Helix to β-Hairpin Transition. Proteins 2003, 53, 220–228. [Google Scholar] [CrossRef]
- Van Loey, A.; Verachtert, B.; Hendrickx, M. Effects of high electric field pulses on enzymes. Trends Food Sci. Technol. 2002, 12, 94–102. [Google Scholar] [CrossRef]
- Yeom, H.W.; Zhang, Q.H. Enzymatic inactivation by pulsed electric fields: A review. In Pulsed Electric Fields in Food Processing; Barbosa-Cánovas, G.V., Zhang, Q.H., Eds.; Technomic Publishing: Lancaster, UK, 2001; pp. 57–63. [Google Scholar]
- Vega-Mercado, H.M.; Martin-Belloso, O.; Qin, B.L.; Chang, F.J.; Góngora-Nieto, M.M.; Barbara-Cánovas, G.V.; Swanson, B.G. Non-thermal food preservation: Pulsed electric fields. Trends Food Sci. Technol. 1997, 8, 151–157. [Google Scholar] [CrossRef]
- Castro, A.J.; Swanson, B.G.; Barbosa-Cánovas, G.V.; Zhang, Q.H. Pulsed electric fields modification of milk alkaline phosphatase activity. In Pulsed Electric Fields in Food Processing; Barbosa-Cánovas, G.V., Zhang, Q.H., Eds.; Technomic Publishing: Lancaster, UK, 2001; pp. 65–82. [Google Scholar]
- Dong, M.; Xu, Y.; Zhang, Y.; Han, M.; Wang, P.; Xu, X.; Zhou, G. Physicochemical and structural properties of myofibrillar proteins isolated from pale, soft, exudative (PSE)-like chicken breast meat: Effects of pulsed electric field (PEF). Innov. Food Sci. Emerg. Technol. 2020, 59, 102277. [Google Scholar] [CrossRef]
- Kubo, T.; Mizobata, T.; Kawata, Y. Refolding of yeast enolase in the presence of the chaperonin GroE. The nucleotide specificity of GroE and the role of GroES. J. Biol. Chem. 1993, 268, 19346–19351. [Google Scholar] [CrossRef]
- Yeom, H.W.; Streaker, C.B.; Zhang, Q.H.; Min, D.B. Effects of pulsed electric fields on the activities of microorganisms and pectin methyl esterase in orange juice. J. Food Sci. 2000, 65, 1359–1363. [Google Scholar] [CrossRef]
- Elez-Martínez, P.; Aguiló-Aguayo, I.; Martin-Belloso, O. Inactivation of orange juice peroxidase by high-intensity pulsed electric fields as influenced by process parameters. J. Sci. Food Agric. 2006, 86, 71–81. [Google Scholar] [CrossRef]
- Shamsi, K.; Versteeg, C.; Sherkat, F.; Wan, J. Alkaline phosphatase and microbial inactivation by pulsed electric field in bovine milk. Innov. Food Sci. Emerg. Technol. 2008, 9, 217–223. [Google Scholar] [CrossRef]
- Ho, S.Y.; Mittal, G.S.; Cross, J.D. Effects of high field electric pulses on the activity of selected enzymes. J. Food Eng. 1997, 31, 69–84. [Google Scholar] [CrossRef]
- Agcam, E.; Akyıldız, A.; Evrendilek, G.A. Effects of PEF and heat pasteurization on PME activity in orange juice with regard to a new inactivation kinetic model. Food Chem. 2014, 165, 70–76. [Google Scholar] [CrossRef]
- Sharma, P.; Oey, I.; Bremer, P.; Everett, D.W. Reduction of bacterial counts and inactivation of enzymes in bovine whole milk using pulsed electric fields. Int. Dairy J. 2014, 39, 146–156. [Google Scholar] [CrossRef]
- Schoenbach, K.H.; Peterkin, F.E.; Alden, R.W., III; Beebe, S.J. The Effect of Pulsed Electric Fields on Biological Cells: Experiments and Applications. IEEE Trans. Plasma Sci. 1997, 25, 284–292. [Google Scholar] [CrossRef]
- Schoenbach, K.H.; Hargrave, B.; Joshi, R.P.; Kolb, J.F.; Nuccitelli, R.; Osgood, C.; Pakhomov, A.; Stacey, M.; Swanson, R.J.; White, J.A.; et al. Bioelectric Effects of Intense Nanosecond Pulses. IEEE Trans. Dielectr. Electr. Ins. 2007, 14, 1088–1109. [Google Scholar] [CrossRef] [Green Version]
- Schoenbach, K.H.; Xiao, S.; Joshi, R.P.; Camp, J.T.; Heeren, T.; Kolb, J.F.; Beebe, S.J. The Effect of Intense Subnanosecond Electrical Pulses on Biological Cells. IEEE Trans. Plasma Sci. 2008, 36, 414–422. [Google Scholar] [CrossRef]
- Hülsheger, H.; Potel, J.; Niemann, E.G. Killing of bacteria with electric pulses of high electric field strength. Radiat. Environ. Biophys. 1981, 20, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Barbosa-Canovas, G.V.; Altunakar, B. Pulsed electric fields processing of foods: An overview. In Pulsed Electric Field Technology for Food Industry: Fundamentals and Applications; Raso, J., Heinz, V., Eds.; Springer: New York, NY, USA, 2006; pp. 3–26. [Google Scholar]
- Barba, F.J.; Parniakov, O.; Pereira, S.A.; Wiktor, A.; Grimi, N.; Boussetta, N.; Saraiva, J.A.; Raso, J.; Martin-Belloso, O.; Witrowa-Rajchert, D.; et al. Current applications and new opportunities for the use of pulsed electric fields in food science and industry. Food Res. Int. 2015, 77, 773–798. [Google Scholar] [CrossRef]
- Min, S.; Evrendilek, G.A.; Zhang, H.Q. Pulsed Electric Fields: Processing System, Microbial and Enzyme Inhibition, and Shelf Life Extension of Foods. IEEE Trans. Plasma Sci. 2007, 35, 59–73. [Google Scholar] [CrossRef]
- Castro, A.J.; Barbosa-Cánovas, G.V.; Swanson, B.G. Microbial inactivation of foods by pulsed electric field. J. Food Process. Preserv. 1993, 17, 47–73. [Google Scholar] [CrossRef]
- Zhao, W.; Yang, R.; Shen, X.; Zhang, S.; Chen, X. Lethal and sublethal injury and kinetics of Escherichia coli, Listeria monocytogenes and Staphylococcus aureus in milk by pulsed electric fields. Food Control 2013, 32, 6–12. [Google Scholar] [CrossRef]
- Sharma, P.; Bremer, P.; Oey, I.; Everett, D.W. Bacterial inactivation in whole milk using pulsed electric field processing. Int. Dairy J. 2014, 35, 49–56. [Google Scholar] [CrossRef]
- Cregenzán-Alberti, O.; Halpin, R.M.; Whyte, P.; Lyng, J.; Noci, F. Suitability of ccRSM as a tool to predict inactivation and its kinetics for Escherichia coli, Staphylococcus aureus and Pseudomonas fluorescens in homogenized milk treated by manothermosonication (MTS). Food Control 2014, 39, 41–48. [Google Scholar] [CrossRef]
- Ohshima, T.; Tanino, T.; Kameda, T.; Harashima, H. Engineering of operation condition in milk pasteurization with PEF treatment. Food Control 2016, 68, 297–302. [Google Scholar] [CrossRef]
- Ohshima, T.; Sato, K.; Terauchi, H.; Sato, M. Physical and chemical modifications of high-voltage pulse sterilization. J. Electrostat. 1997, 42, 159–166. [Google Scholar] [CrossRef]
- Sato, M.; Ishida, N.M.; Sugiarto, A.T.; Ohshima, T.; Taniguchi, H. High-efficiency sterilizer by high-voltage pulse using concentrated-field electrode system. IEEE Trans. Ind. Appl. 2001, 37, 1646–1650. [Google Scholar] [CrossRef]
- Kitajima, N.; Ueda, K.; Ohshima, T.; Sato, M. Development of textile electrode for microbial inactivation with pulsed electric field Textile. Res. J. 2007, 77, 528–534. [Google Scholar]
- Zimmermann, U.; Pilwat, G.; Riemann, F. Preparation of erythrocyte ghosts by dielectric breakdown of the cell membrane. Biochim. Biophys. Acta 1975, 375, 209–219. [Google Scholar] [CrossRef]
- Ohshima, T.; Sato, M.; Saito, M. Selective release of intracellular protein using pulsed electric field. J. Electrostat. 1995, 35, 103–112. [Google Scholar] [CrossRef]
- Ohshima, T.; Hama, Y.; Sato, M. Releasing profiles of gene products from recombinant Escherichia coli in a high-voltage pulsed electric field. Biochem. Eng. J. 2000, 5, 149–155. [Google Scholar] [CrossRef]
- Shiina, S.; Ohshima, T.; Sato, M. Extracellular release of recombinant α-amylase from Escherichia coli using pulsed electric field. Biotechnol. Prog. 2004, 20, 1528–1533. [Google Scholar] [CrossRef]
- Shiina, S.; Ohshima, T.; Sato, M. Extracellular production of α-amylase during fed-batch cultivation of recombinant Escherichia coli using pulsed electric field. J. Electrost. 2007, 65, 30–36. [Google Scholar] [CrossRef]
- Mahnič-Kalamiza, S.; Vorobiev, E.; Miklavčič, D. Electroporation in food processing and biorefinery. J. Membr. Biol. 2014, 247, 1279–1304. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, A.; Hatayama, H.; Takaki, K.; Koide, K.; Kawamura, Y. Influence of pulse width on polyphenol extraction from agricultural products by pulsed electric field. IEEJ Trans. Fundam. Mater. 2013, 133, 32–37. (In Japanese) [Google Scholar] [CrossRef]
- Vega-Mercado, H.; Gongora-Nieto, M.M.; Barbosa-Canovas, G.V. Advances in dehydration of foods. J. Food Eng. 2001, 49, 271–289. [Google Scholar] [CrossRef]
- Orikasa, T.; Koide, S.; Sugawara, H.; Yoshida, M.; Kato, K.; Matsushima, U.; Okada, M.; Watanabe, T.; Ando, Y.; Shiina, T.; et al. Applicability of vacuum-microwave drying for tomato fruit based on evaluations of energy cost, color, functional components, and sensory qualities. J. Food Process. Preserv. 2018, 42, 1–12. [Google Scholar] [CrossRef]
- Aghilinategh, N.; Rafiee, S.; Gholikhani, A.; Hosseinpur, S.; Omid, M.; Mohtasebi, S.S.; Maleki, N. A comparative study of dried apple using hot air, intermittent and continuous microwave: Evaluation of kinetic parameters and physicochemical quality attributes. Food Sci. Nutr. 2015, 3, 519–526. [Google Scholar] [CrossRef]
- Drouzas, A.E.; Tsami, E.; Saravacos, G.D. Microwave/vacuum drying of model fruit gels. J. Food Eng. 1999, 39, 117–122. [Google Scholar] [CrossRef]
- Ade-Omowaye, B.I.O.; Rastogi, N.K.; Angersbach, A.; Knorr, D. Combined effects of pulsed electric field pre-treatment and partial osmotic dehydration on air drying behaviour of red bell pepper. J. Food Eng. 2003, 60, 89–98. [Google Scholar] [CrossRef]
- Wiktor, A.; Dadan, M.; Nowacka, M.; Rybak, K.; Witrowa-Rajchert, D. The impact of combination of pulsed electric field and ultrasound treatment on air drying kinetics and quality of carrot tissue. LWT Food Sci. Technol. 2019, 110, 71–79. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Wang, L.H.; Zeng, X.A.; Han, Z.; Wang, M.S. Effect of pulsed electric fields (PEFs) on the pigments extracted from spinach (Spinacia oleracea L.). Innov. Food Sci. Emerg. Technol. 2017, 43, 26–34. [Google Scholar] [CrossRef]
- Huang, W.; Feng, Z.; Aila, R.; Hou, Y.; Carne, A.; Bekhit, A.E.D.A. Effect of pulsed electric fields (PEF) on physico-chemical properties, β-carotene and antioxidant activity of air-dried apricots. Food Chem. 2019, 291, 253–262. [Google Scholar] [CrossRef]
- Janositz, A.; Noack, A.K.; Knorr, D. Pulsed electric fields and their impact on the diffusion characteristics of potato slices. LWT Food Sci. Technol. 2011, 44, 1939–1945. [Google Scholar] [CrossRef]
- Ade-Omowaye, B.I.O.; Angersbach, A.; Taiwo, K.A.; Knorr, D. Use of pulsed electric field pre-treatment to improve dehydration characteristics of plant based foods. Trends Food Sci. Technol. 2001, 12, 285–295. [Google Scholar] [CrossRef]
- Lebovka, N.I.; Shynkaryk, N.V.; Vorobiev, E. Pulsed electric field enhanced drying of potato tissue. J. Food Eng. 2007, 78, 606–613. [Google Scholar] [CrossRef]
- Gachovska, T.K.; Simpson, M.V.; Ngadi, M.O.; Raghavan, G.S.V. Pulsed electric field treatment of carrots before drying and rehydration. J. Sci. Food Agric. 2009, 89, 2372–2376. [Google Scholar] [CrossRef]
- Lamanauskas, N.; Satkauskas, S.; Bobinaite, R.; Viskelis, P. Pulsed electric field (PEF) impact on Actinidia kolomikta drying efficiency. J. Food Process Eng. 2015, 38, 243–249. [Google Scholar] [CrossRef]
- Telfser, A.; Galindo, F.G. Effect of reversible permeabilization in combination with different drying methods on the structure and sensorial quality of dried basil (Ocimum basilicum L.) leaves. LWT Food Sci. Technol. 2019, 99, 148–155. [Google Scholar] [CrossRef]
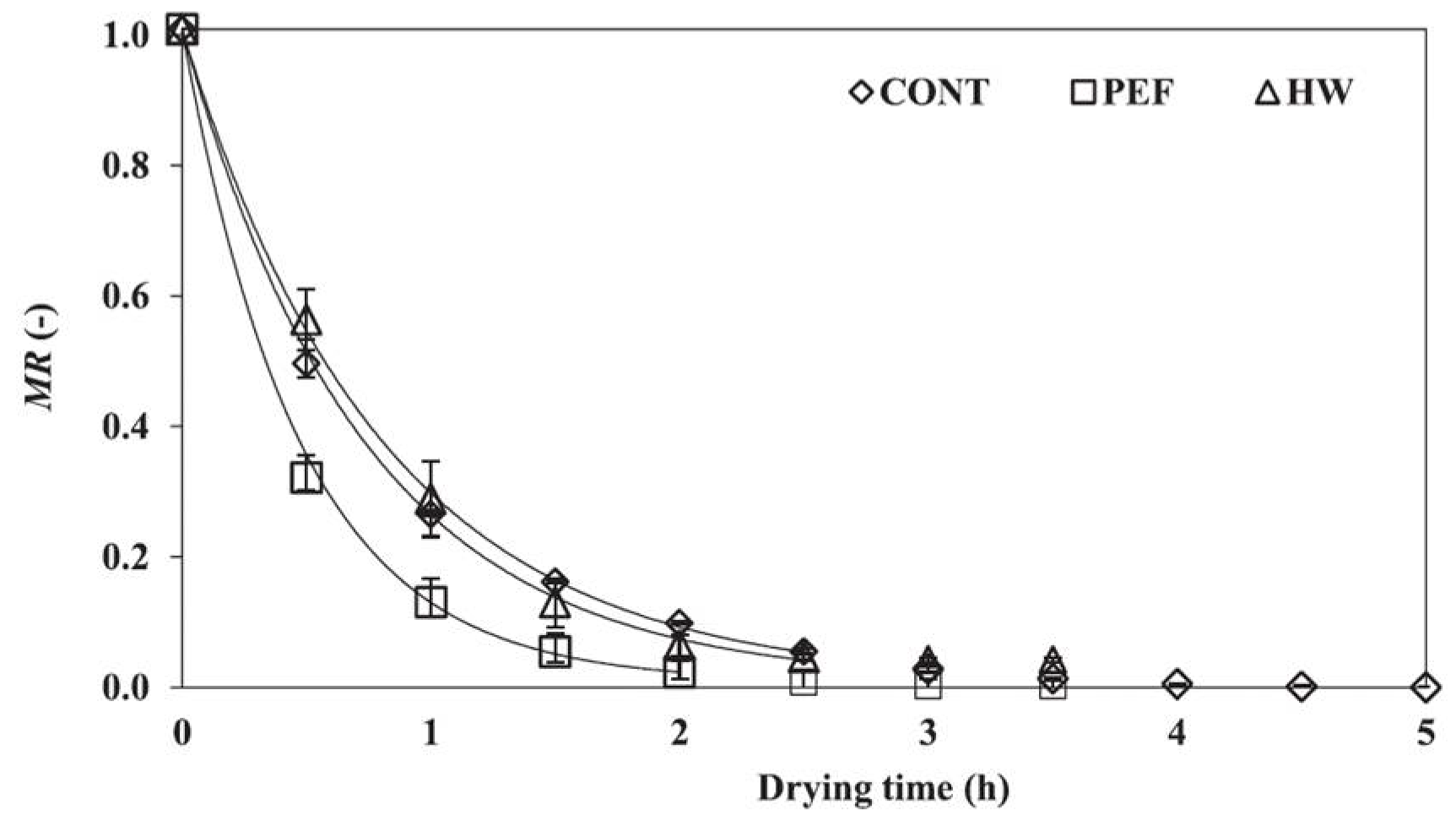
- Yamada, T.; Yamakage, K.; Takahashi, K.; Takaki, K.; Orikasa, T.; Kamagata, J.; Aoki, H. Influence of drying rate on hot air drying processing of fresh foods using pulsed electric field. Inst. Electr. Eng. Jpn. 2020, 15, 123–125. [Google Scholar] [CrossRef]
- Yamakage, K.; Yamada, T.; Takahashi, K.; Takaki, K.; Komuro, M.; Sasaki, K.; Aoki, H.; Kamagata, J.; Koide, S.; Orikasa, T. Impact of pre-treatment with pulsed electric field on drying rate and changes in spinach quality during hot air drying. Innov. Food Sci. Emerg. Technol. 2021, 68, 102615. [Google Scholar] [CrossRef]
- Yongguang, Y.; Yong, H.; Jingbo, L. A novel protecting method for visual green color in spinach puree treated by high intensity pulsed electric fields. J. Food Eng. 2007, 79, 1256–1260. [Google Scholar]
- Zhang, F.; Tian, M.; Du, M.; Fang, T. Enhancing the activity of pectinase using pulsed electric field (PEF) treatment. J. Food Eng. 2017, 205, 56–63. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drying Constant k | Ratio of Drying Constant | |
|---|---|---|
| CONT | 1.24 | - |
| PEF | 2.11 | 1.70 |
| HW | 1.37 | 1.10 |
| Ratio of Surface Area (−) | Residual Ratio of L-AsA (−) | |
|---|---|---|
| CONT | 0.352 ± 0.015 a | 0.74 ± 0.04 a |
| REF | 0.489 ± 0.002 b | 0.74 ± 0.04 a |
| HW | 0.349 ± 0.004 a | 0.52 ± 0.03 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takaki, K.; Takahashi, K.; Guionet, A.; Ohshima, T. Pulsed Power Applications for Protein Conformational Change and the Permeabilization of Agricultural Products. Molecules 2021, 26, 6288. https://doi.org/10.3390/molecules26206288
Takaki K, Takahashi K, Guionet A, Ohshima T. Pulsed Power Applications for Protein Conformational Change and the Permeabilization of Agricultural Products. Molecules. 2021; 26(20):6288. https://doi.org/10.3390/molecules26206288
Chicago/Turabian StyleTakaki, Koichi, Katsuyuki Takahashi, Alexis Guionet, and Takayuki Ohshima. 2021. "Pulsed Power Applications for Protein Conformational Change and the Permeabilization of Agricultural Products" Molecules 26, no. 20: 6288. https://doi.org/10.3390/molecules26206288
APA StyleTakaki, K., Takahashi, K., Guionet, A., & Ohshima, T. (2021). Pulsed Power Applications for Protein Conformational Change and the Permeabilization of Agricultural Products. Molecules, 26(20), 6288. https://doi.org/10.3390/molecules26206288