
Ginger, a Possible Candidate for the Treatment of Dementias?


Abstract
1. Introduction
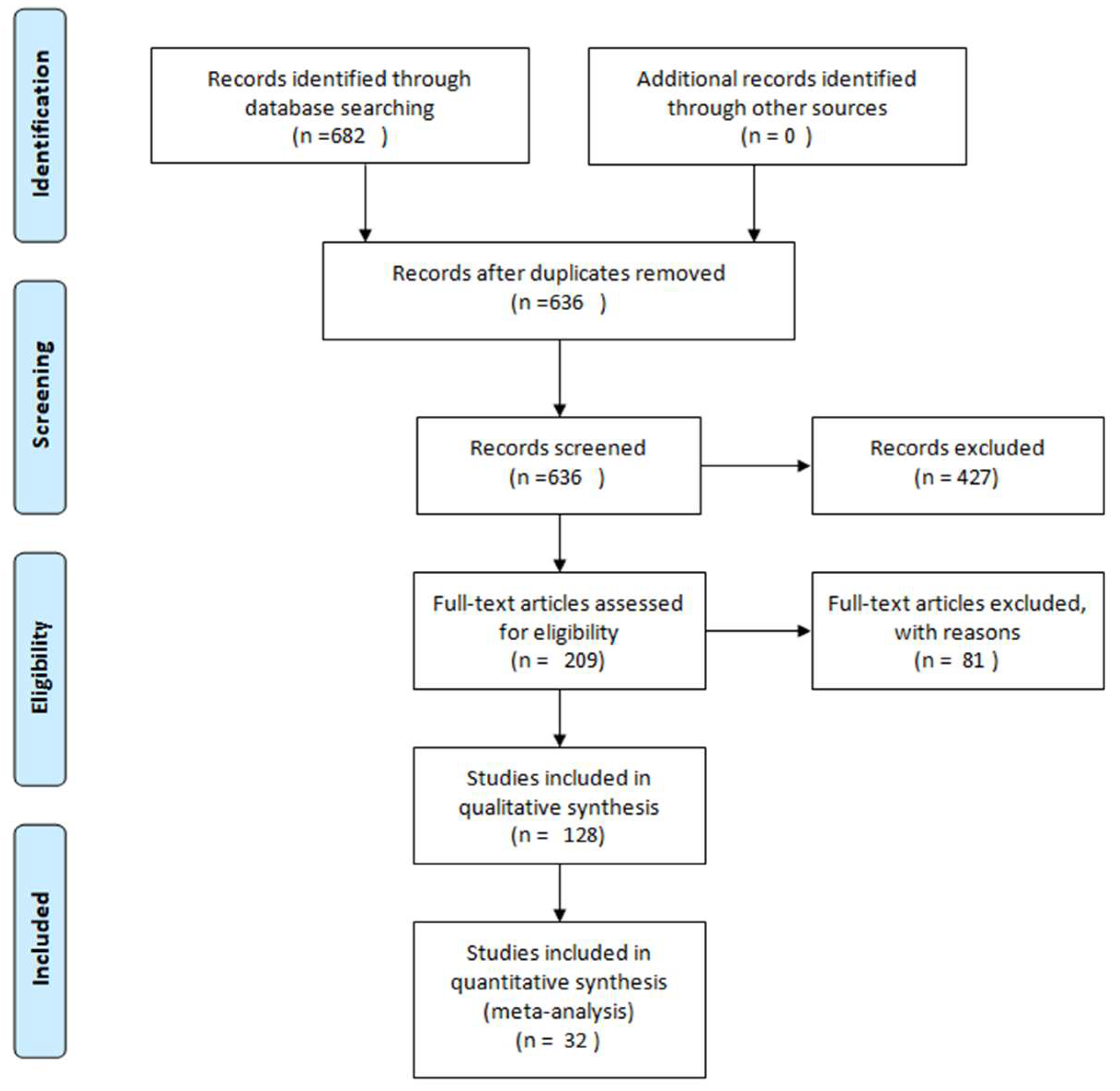
2. Methodology
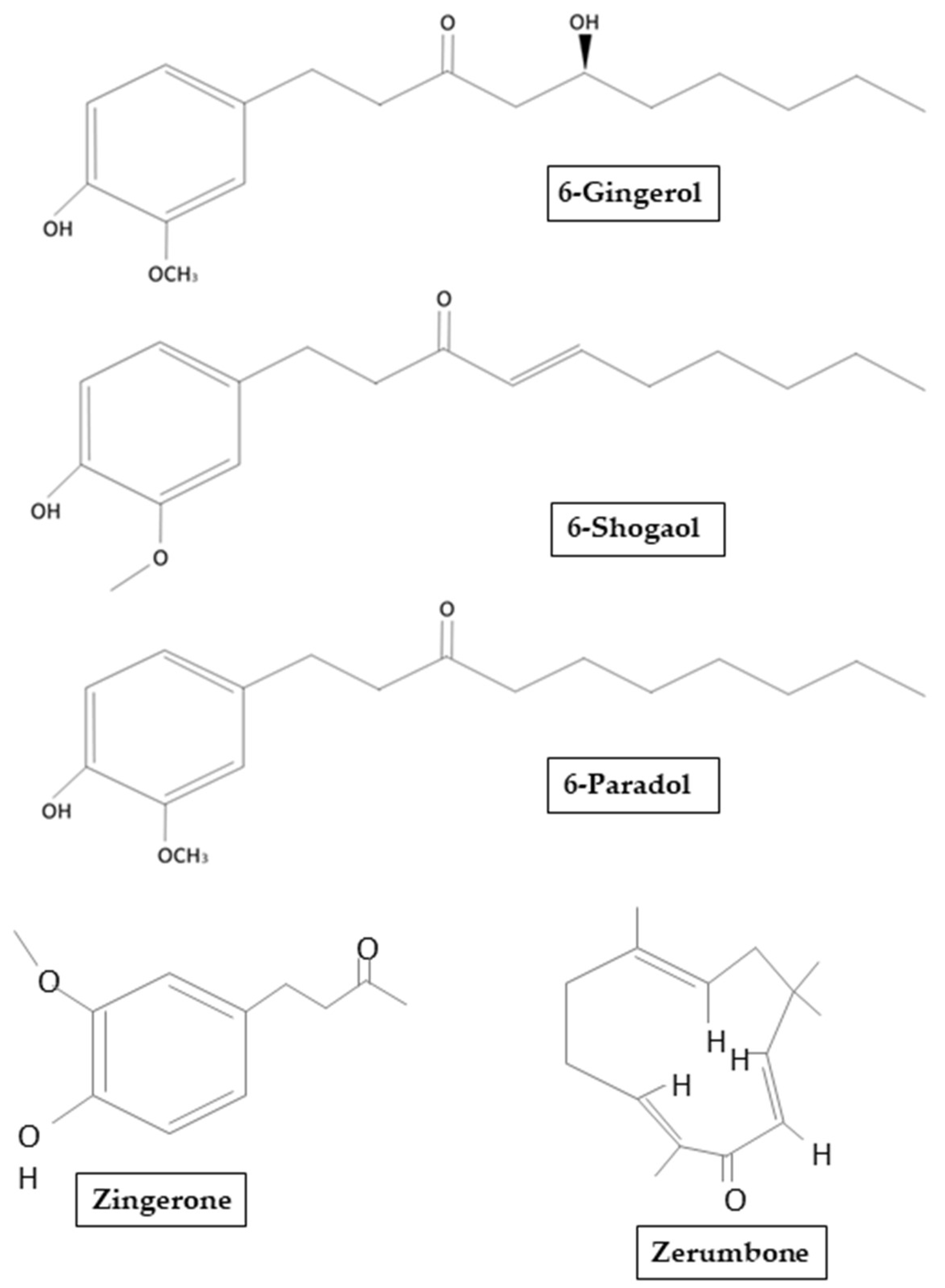
3. Ginger: Chemical Structure, Properties, and Therapeutic Potential
4. Alzheimer’s Disease
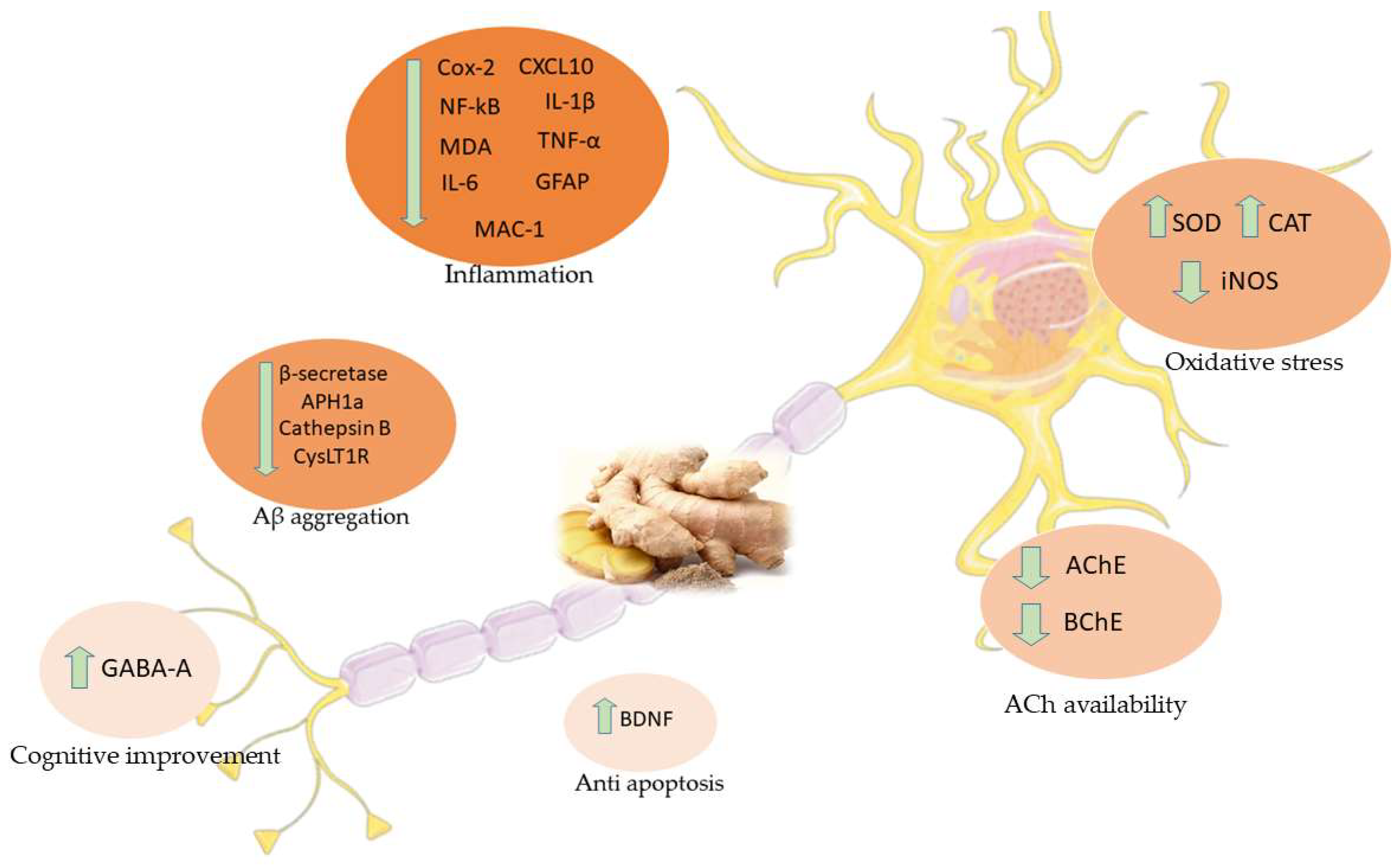
5. Effects of Ginger in AD Studies
5.1. Computational Studies
5.2. In Vivo and In Vitro Studies
6. Vascular Dementia
7. Effects of Ginger in VD Studies
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DSM-5 | Diagnostic and Statistical Manual of Mental Disorders |
| AD | Alzheimer’s Disease |
| VD | Vascular Dementia |
| NFTs | Neurofibrillary tangles |
| Aβ | Beta-amyloid |
| BBB | Blood-brain barrier |
| COX-1 | Cyclooxygenase 1 |
| APOE | Apolipoprotein E |
| PSEN 1 | Presenilin 1 |
| PSEN 2 | Presenilin 2 |
| APP | Amyloid precursor protein |
| GSK-3 | Glycogen synthase kinase-3 |
| ROS | Reactive oxygen species reactive |
| MDA | Malondialdehyde |
| NSAIDs | Non-steroidal anti-inflammatory drugs |
| COX-2 | Cyclooxygenase-2 |
| 5-LOX | 5-lipoxygenase |
| LTs | Leukotrienes |
| SAD | Late-onset sporadic Alzheimer Disease |
| FAD | Early-onset familial Alzheimer Disease |
| APH1a | Gamma-secretase subunit APH-1A |
| AChE | Acetylcholinesterase |
| ACh | Acetylcholine |
| Zo.Cr | Dried ginger |
| BChE | Butyrylcholinesterase |
| SOD | Superoxide dismutase |
| CAT | Catalase |
| ERK | eEtracellular signal-regulated kinase |
| BDNF | Brain-derived neurotrophic factor |
| GRE | Ginger root extract |
| NF-κB | Nuclear factor kappa-B |
| IL | Interleukin |
| GFAP | Glial fibrillary acid protein |
| Mac-1 | Macrophage-1 antigen |
| CysLT1R | Cysteinyl leukotriene 1 receptor |
| CysLTs | Cysteinyl leukotrienes |
| AA | Arachidonic acid |
| SORL1 | Sortilin-related receptor 1 |
| siRNA | Small interfering RNA |
| BACE1 | Beta-site amyloid precursor protein cleaving enzyme 1 |
| GABA | Gamma-Aminobutyric acid |
| MAPK | Mitogen-activated protein kinase |
| PGE2 | Prostaglandin E2 |
| iNOS | Inducible nitric oxide synthetase |
| JNK | C-jun N-terminal kinase |
| ER | Estrogen receptor |
| Bcl-2 | B-cell lymphoma 2 |
| BAX | BCL2-associated X: apoptosis regulator |
| NRF2 | Nuclear factor erythroid 2-related factor 2 |
| GCL | C-glutamylcysteine ligase |
| HO-1 | Heme Oxygenase-1 |
| NO | Nitric oxide |
| GE | Ginger extracts |
| LPS | Lipopolysaccharide |
| TNF-α | Tumour necrosis factor-α |
| MCP-1 | Monocyte chemoattractant protein-1 |
| CCL2 | C-C motif chemokine ligand 2 |
| MIP-1α | Macrophage inflammatory protein-1α |
| CCL3 | C-C motif chemokine ligand 3 |
| ATP | Adenosine 5’-triphosphate |
| NLRP3 | Nucleotide-binding domain and leucine-rich repeat-containing family, pyrin do-main-containing 3 |
| ELNs | Exosome-like nanoparticles |
| G-ELNs | Ginger rhizomes |
| ASC | Apoptotic speck protein containing a caspase recruitment domain |
| TACE | TNF-α converting enzyme |
| NMDA | N-methyl-D-aspartate |
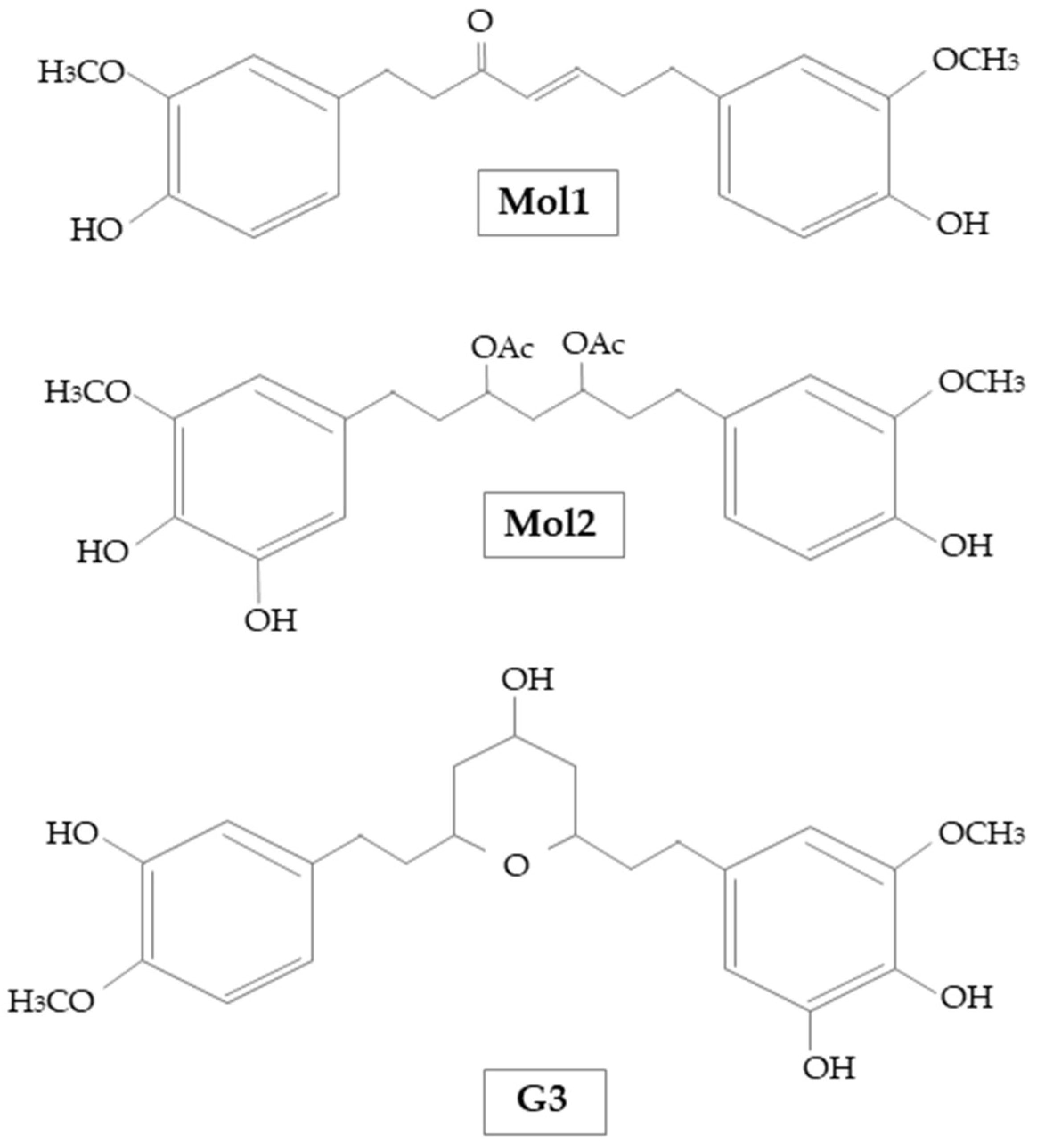
| Mol1 | [(E)-1,7-bis (4-hydroxy-3-methoxyphenyl) hept-4-en-3-on] |
| Mol2 | [1-(3,4-dihydroxy-5-methoxyphenyl)-7-(4-hydroxy-3-ethoxyphenyl) heptane-3,5-diyl diacetate] |
| G3 | 5-[(2S,4R,6R)-4-hydroxy-6-[2-(4-methoxyphenyl)ethyl]oxan-2-yl]-3-methoxybenzene-1,2-diol |
| PPARγ | Peroxisome proliferation activator receptor γ |
| MCI | Mild Cognitive Impairment |
| VaMCI | Vascular Mild Cognitive Impairment |
| VCI | Vascular Cognitive Impairment |
| MetS | Metabolic syndrome |
| PMG | Mulberry fruit and ginger |
| Rt. MCAO | Right middle cerebral artery |
| GPx | Glutathione peroxidase |
References
- Libro, R.; Giacoppo, S.; Soundara Rajan, T.; Bramanti, P.; Mazzon, E. Natural Phytochemicals in the Treatment and Prevention of Dementia: An Overview. Molecules 2016, 21, 518. [Google Scholar] [CrossRef]
- Sacuiu, S.F. Dementias. Handb. Clin. Neurol. 2016, 138, 123–151. [Google Scholar] [CrossRef] [PubMed]
- Gale, S.A.; Acar, D.; Daffner, K.R. Dementia. Am. J. Med. 2018, 131, 1161–1169. [Google Scholar] [CrossRef]
- Aarsland, D. Epidemiology and Pathophysiology of Dementia-Related Psychosis. J. Clin. Psychiatry 2020, 81. [Google Scholar] [CrossRef] [PubMed]
- Soria Lopez, J.A.; Gonzalez, H.M.; Leger, G.C. Alzheimer’s disease. Handb. Clin. Neurol. 2019, 167, 231–255. [Google Scholar] [CrossRef]
- Iadecola, C. The pathobiology of vascular dementia. Neuron 2013, 80, 844–866. [Google Scholar] [CrossRef]
- Gorelick, P.B.; Scuteri, A.; Black, S.E.; Decarli, C.; Greenberg, S.M.; Iadecola, C.; Launer, L.J.; Laurent, S.; Lopez, O.L.; Nyenhuis, D.; et al. Vascular contributions to cognitive impairment and dementia: A statement for healthcare professionals from the american heart association/american stroke association. Stroke 2011, 42, 2672–2713. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed]
- Talebi, M.; Ilgun, S.; Ebrahimi, V.; Talebi, M.; Farkhondeh, T.; Ebrahimi, H.; Samarghandian, S. Zingiber officinale ameliorates Alzheimer’s disease and Cognitive Impairments: Lessons from preclinical studies. Biomed. Pharmacother. 2021, 133, 111088. [Google Scholar] [CrossRef]
- De Lima, R.M.T.; Dos Reis, A.C.; de Menezes, A.P.M.; Santos, J.V.O.; Filho, J.; Ferreira, J.R.O.; de Alencar, M.; da Mata, A.; Khan, I.N.; Islam, A.; et al. Protective and therapeutic potential of ginger (Zingiber officinale) extract and [6]-gingerol in cancer: A comprehensive review. Phytother. Res. 2018, 32, 1885–1907. [Google Scholar] [CrossRef] [PubMed]
- Nagendra chari, K.L.; Manasa, D.; Srinivas, P.; Sowbhagya, H.B. Enzyme-assisted extraction of bioactive compounds from ginger (Zingiber officinale Roscoe). Food Chem. 2013, 139, 509–514. [Google Scholar] [CrossRef]
- Choi, J.G.; Kim, S.Y.; Jeong, M.; Oh, M.S. Pharmacotherapeutic potential of ginger and its compounds in age-related neurological disorders. Pharmacol. Ther. 2018, 182, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Pagano, E.; Souto, E.B.; Durazzo, A.; Sharifi-Rad, J.; Lucarini, M.; Souto, S.B.; Salehi, B.; Zam, W.; Montanaro, V.; Lucariello, G.; et al. Ginger (Zingiber officinale Roscoe) as a nutraceutical: Focus on the metabolic, analgesic, and antiinflammatory effects. Phytother. Res. 2020, 35, 2403–2417. [Google Scholar] [CrossRef] [PubMed]
- Mohd Yusof, Y.A. Gingerol and Its Role in Chronic Diseases. Adv. Exp. Med. Biol. 2016, 929, 177–207. [Google Scholar] [CrossRef]
- Rai, S.; Mukherjee, K.; Mal, M.; Wahile, A.; Saha, B.P.; Mukherjee, P.K. Determination of 6-gingerol in ginger (Zingiber officinale) using high-performance thin-layer chromatography. J. Sep. Sci. 2006, 29, 2292–2295. [Google Scholar] [CrossRef]
- Jung, M.Y.; Lee, M.K.; Park, H.J.; Oh, E.B.; Shin, J.Y.; Park, J.S.; Jung, S.Y.; Oh, J.H.; Choi, D.S. Heat-induced conversion of gingerols to shogaols in ginger as affected by heat type (dry or moist heat), sample type (fresh or dried), temperature and time. Food Sci. Biotechnol. 2018, 27, 687–693. [Google Scholar] [CrossRef]
- Mao, Q.Q.; Xu, X.Y.; Cao, S.Y.; Gan, R.Y.; Corke, H.; Beta, T.; Li, H.B. Bioactive Compounds and Bioactivities of Ginger (Zingiber officinale Roscoe). Foods 2019, 8, 185. [Google Scholar] [CrossRef]
- Mohd Yusof, Y.A. Gingerol and Its Role in Chronic Diseases. In Drug Discovery from Mother Nature; Gupta, S.C., Prasad, S., Aggarwal, B.B., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 177–207. [Google Scholar] [CrossRef]
- Chrubasik, S.; Pittler, M.H.; Roufogalis, B.D. Zingiberis rhizoma: a comprehensive review on the ginger effect and efficacy profiles. Phytomed. Int. J. Phytother. Phytopharm. 2005, 12, 684–701. [Google Scholar] [CrossRef]
- Bordia, A.; Verma, S.K.; Srivastava, K.C. Effect of ginger (Zingiber officinale Rosc.) and fenugreek (Trigonella foenumgraecum L.) on blood lipids, blood sugar and platelet aggregation in patients with coronary artery disease. Prostaglandins Leukot. Essent. Fat. Acids 1997, 56, 379–384. [Google Scholar] [CrossRef]
- Nakazawa, T.; Ohsawa, K. Metabolism of [6]-gingerol in rats. Life Sci. 2002, 70, 2165–2175. [Google Scholar] [CrossRef]
- Iqbal, K.; Grundke-Iqbal, I. Alzheimer’s disease, a multifactorial disorder seeking multitherapies. Alzheimer’s Dement. 2010, 6, 420–424. [Google Scholar] [CrossRef]
- Chauhan, N.; Wang, K.C.; Wegiel, J.; Malik, M.N. Walnut extract inhibits the fibrillization of amyloid beta-protein, and also defibrillizes its preformed fibrils. Curr. Alzheimer Res. 2004, 1, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Muller, U.C.; Deller, T.; Korte, M. Not just amyloid: physiological functions of the amyloid precursor protein family. Nat. Rev. Neurosci. 2017, 18, 281–298. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.S.; Stachowiak, A.; Mamun, A.A.; Tzvetkov, N.T.; Takeda, S.; Atanasov, A.G.; Bergantin, L.B.; Abdel-Daim, M.M.; Stankiewicz, A.M. Autophagy and Alzheimer’s Disease: From Molecular Mechanisms to Therapeutic Implications. Front. Aging Neurosci. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Lindwall, G.; Cole, R.D. Phosphorylation affects the ability of tau protein to promote microtubule assembly. J. Biol Chem 1984, 259, 5301–5305. [Google Scholar] [CrossRef]
- Khan, S.S.; Bloom, G.S. Tau: The Center of a Signaling Nexus in Alzheimer’s Disease. Front. Neurosci. 2016, 10, 31. [Google Scholar] [CrossRef] [PubMed]
- Avila, J.; Lucas, J.J.; Perez, M.; Hernandez, F. Role of tau protein in both physiological and pathological conditions. Physiol. Rev. 2004, 84, 361–384. [Google Scholar] [CrossRef]
- Iqbal, K.; Alonso Adel, C.; Chen, S.; Chohan, M.O.; El-Akkad, E.; Gong, C.X.; Khatoon, S.; Li, B.; Liu, F.; Rahman, A.; et al. Tau pathology in Alzheimer disease and other tauopathies. Biochim. Et Biophys. Acta 2005, 1739, 198–210. [Google Scholar] [CrossRef]
- Avila, J. Tau phosphorylation and aggregation in Alzheimer’s disease pathology. FEBS Lett. 2006, 580, 2922–2927. [Google Scholar] [CrossRef]
- Terzo, S.; Amato, A.; Mulè, F. From obesity to Alzheimer’s disease through insulin resistance. J. Diabetes Its Complicat. 2021. [Google Scholar] [CrossRef]
- Anekonda, T.S.; Reddy, P.H. Can herbs provide a new generation of drugs for treating Alzheimer’s disease? Brain Res. Rev. 2005, 50, 361–376. [Google Scholar] [CrossRef]
- Grzanna, R.; Lindmark, L.; Frondoza, C.G. Ginger—an herbal medicinal product with broad anti-inflammatory actions. J. Med. Food 2005, 8, 125–132. [Google Scholar] [CrossRef]
- Afzal, M.; Al-Hadidi, D.; Menon, M.; Pesek, J.; Dhami, M.S. Ginger: An ethnomedical, chemical and pharmacological review. Drug Metab. Drug Interact. 2001, 18, 159–190. [Google Scholar] [CrossRef]
- Flynn, D.L.; Rafferty, M.F.; Boctor, A.M. Inhibition of human neutrophil 5-lipoxygenase activity by gingerdione, shogaol, capsaicin and related pungent compounds. Prostaglandins Leukot. Med. 1986, 24, 195–198. [Google Scholar] [CrossRef]
- Charlier, C.; Michaux, C. Dual inhibition of cyclooxygenase-2 (COX-2) and 5-lipoxygenase (5-LOX) as a new strategy to provide safer non-steroidal anti-inflammatory drugs. Eur. J. Med. Chem. 2003, 38, 645–659. [Google Scholar] [CrossRef]
- Martel-Pelletier, J.; Lajeunesse, D.; Reboul, P.; Pelletier, J.P. Therapeutic role of dual inhibitors of 5-LOX and COX, selective and non-selective non-steroidal anti-inflammatory drugs. Ann. Rheum. Dis. 2003, 62, 501–509. [Google Scholar] [CrossRef]
- Radmark, O.; Samuelsson, B. 5-Lipoxygenase: Mechanisms of regulation. J. Lipid Res. 2009, 50, S40–S45. [Google Scholar] [CrossRef]
- Jeon, S.G.; Song, E.J.; Lee, D.; Park, J.; Nam, Y.; Kim, J.I.; Moon, M. Traditional oriental medicines and Alzheimer’s disease. Aging Dis. 2019, 10, 307–328. [Google Scholar] [CrossRef] [PubMed]
- Dabaghian, F.; Azadi, A.; Setooni, M.; Zarshenas, M.M. An overview on multi-ingredient memory enhancers and anti-Alzheimer’s formulations from traditional persian pharmacy. Trends Pharm. Sci. 2017, 3, 215–220. [Google Scholar]
- Azam, F.; Amer, A.M.; Abulifa, A.R.; Elzwawi, M.M. Ginger components as new leads for the design and development of novel multi-targeted anti-Alzheimer’s drugs: a computational investigation. Drug Des. Dev. Ther. 2014, 8, 2045–2059. [Google Scholar] [CrossRef] [PubMed]
- Cuya, T.; Baptista, L.; Franca, T.C.C. A molecular dynamics study of components of the ginger (zingiber officinale) extract inside human acetylcholinesterase: implications for Alzheimer disease. J. Biomol. Struct. Dyn. 2018, 36, 3843–3855. [Google Scholar] [CrossRef] [PubMed]
- Cuya, T.; Franca, T.C.C. A molecular modeling study of components of the ginger (zingiber officinale) extract inside human butyrylcholinesterase: Implications for Alzheimer disease. J. Biomol. Struct. Dyn. 2020, 38, 2809–2815. [Google Scholar] [CrossRef]
- Hwang, J.; Youn, K.; Ji, Y.; Lee, S.; Lim, G.; Lee, J.; Ho, C.T.; Leem, S.H.; Jun, M. Biological and computational studies for dual cholinesterases inhibitory effect of zerumbone. Nutrients 2020, 12, 1215. [Google Scholar] [CrossRef]
- Aguiar-Furucho, M.A.; Pelaez, F.J.R. Alzheimer’s Disease as a Result of Stimulus Reduction in a GABA-A-Deficient Brain: A Neurocomputational Model. Neural Plast. 2020, 2020, 8895369. [Google Scholar] [CrossRef] [PubMed]
- Halawany, A.M.E.; Sayed, N.S.E.; Abdallah, H.M.; Dine, R.S.E. Protective effects of gingerol on streptozotocin-induced sporadic Alzheimer’s disease: Emphasis on inhibition of beta-amyloid, COX-2, alpha-, beta - secretases and APH1a. Sci. Rep. 2017, 7, 2902. [Google Scholar] [CrossRef] [PubMed]
- Ghayur, M.N.; Gilani, A.H.; Ahmed, T.; Khalid, A.; Nawaz, S.A.; Agbedahunsi, J.M.; Choudhary, M.I.; Houghton, P.J. Muscarinic, Ca(++) antagonist and specific butyrylcholinesterase inhibitory activity of dried ginger extract might explain its use in dementia. J. Pharm. Pharmacol. 2008, 60, 1375–1383. [Google Scholar] [CrossRef]
- Sutalangka, C.; Wattanathorn, J. Neuroprotective and cognitive-enhancing effects of the combined extract of cyperus rotundus and zingiber officinale. BMC Complementary Altern. Med. 2017, 17, 135. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Seo, Y.; Lee, C.; Park, G.H.; Jang, J.H. Neuroprotective effect and molecular mechanism of [6]-gingerol against scopolamine-induced amnesia in C57BL/6 mice. Evid.-Based Complementary Altern. Med. Ecam 2018, 2018, 8941564. [Google Scholar] [CrossRef]
- Zeng, G.F.; Zhang, Z.Y.; Lu, L.; Xiao, D.Q.; Zong, S.H.; He, J.M. Protective effects of ginger root extract on Alzheimer disease-induced behavioral dysfunction in rats. Rejuvenation Res. 2013, 16, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Choi, J.G.; Moon, M.; Kim, H.G.; Lee, W.; Bak, H.R.; Sung, H.; Park, C.H.; Kim, S.Y.; Oh, M.S. An optimized combination of ginger and peony root effectively inhibits amyloid-beta accumulation and amyloid-beta-mediated pathology in abetaPP/PS1 double-transgenic mice. J. Alzheimer’s Dis. 2016, 50, 189–200. [Google Scholar] [CrossRef]
- Moon, M.; Kim, H.G.; Choi, J.G.; Oh, H.; Lee, P.K.; Ha, S.K.; Kim, S.Y.; Park, Y.; Huh, Y.; Oh, M.S. 6-Shogaol, an active constituent of ginger, attenuates neuroinflammation and cognitive deficits in animal models of dementia. Biochem. Biophys. Res. Commun. 2014, 449, 8–13. [Google Scholar] [CrossRef]
- Na, J.Y.; Song, K.; Lee, J.W.; Kim, S.; Kwon, J. 6-Shogaol has anti-amyloidogenic activity and ameliorates Alzheimer’s disease via CysLT1R-mediated inhibition of cathepsin B. Biochem. Biophys. Res. Commun. 2016, 477, 96–102. [Google Scholar] [CrossRef]
- Na, J.Y.; Song, K.; Lee, J.W.; Kim, S.; Kwon, J. Sortilin-related receptor 1 interacts with amyloid precursor protein and is activated by 6-shogaol, leading to inhibition of the amyloidogenic pathway. Biochem. Biophys. Res. Commun. 2017, 484, 890–895. [Google Scholar] [CrossRef]
- Huh, E.; Lim, S.; Kim, H.G.; Ha, S.K.; Park, H.Y.; Huh, Y.; Oh, M.S. Ginger fermented with Schizosaccharomyces pombe alleviates memory impairment via protecting hippocampal neuronal cells in amyloid beta1-42 plaque injected mice. Food Funct. 2018, 9, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Jafarian, S.; Ling, K.H.; Hassan, Z.; Perimal-Lewis, L.; Sulaiman, M.R.; Perimal, E.K. Effect of zerumbone on scopolamine-induced memory impairment and anxiety-like behaviours in rats. Alzheimer’s Dement. 2019, 5, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wu, X.H.; Zhao, X.J.; Xu, L.; Pan, C.L.; Zhang, Z.Y. Zerumbone ameliorates behavioral impairments and neuropathology in transgenic APP/PS1 mice by suppressing MAPK signaling. J. Neuroinflammation 2020, 17, 61. [Google Scholar] [CrossRef]
- Kim, M.K.; Chung, S.W.; Kim, D.H.; Kim, J.M.; Lee, E.K.; Kim, J.Y.; Ha, Y.M.; Kim, Y.H.; No, J.K.; Chung, H.S.; et al. Modulation of age-related NF-kappaB activation by dietary zingerone via MAPK pathway. Exp. Gerontol. 2010, 45, 419–426. [Google Scholar] [CrossRef]
- Mizuno, D.; Konoha-Mizuno, K.; Mori, M.; Yamazaki, K.; Haneda, T.; Koyama, H.; Kawahara, M. An in vitro system comprising immortalized hypothalamic neuronal cells (GT1-7 cells) for evaluation of the neuroendocrine effects of essential oils. Evid.-Based Complementary Altern. Med. Ecam 2015, 2015, 343942. [Google Scholar] [CrossRef]
- Lee, C.; Park, G.H.; Kim, C.Y.; Jang, J.H. [6]-Gingerol attenuates β-amyloid-induced oxidative cell death via fortifying cellular antioxidant defense system. Food Chem. Toxicol. 2011, 49, 1261–1269. [Google Scholar] [CrossRef]
- Zeng, G.F.; Zong, S.H.; Zhang, Z.Y.; Fu, S.W.; Li, K.K.; Fang, Y.; Lu, L.; Xiao, D.Q. The role of 6-gingerol on inhibiting amyloid beta protein-induced apoptosis in PC12 cells. Rejuvenation Res. 2015, 18, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Kim, J.Y.; Han, Y.S. Alzheimer’s disease drug discovery from herbs: Neuroprotectivity from beta-amyloid (1-42) insult. J. Altern. Complementary Med. 2007, 13, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Mathew, M.; Subramanian, S. In vitro evaluation of anti-Alzheimer effects of dry ginger (zingiber officinale roscoe) extract. Indian J. Exp. Biol. 2014, 52, 606–612. [Google Scholar] [PubMed]
- Guo, J.P.; Yu, S.; McGeer, P.L. Simple in vitro assays to identify amyloid-beta aggregation blockers for Alzheimer’s disease therapy. J. Alzheimer’s Dis. 2010, 19, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Grzanna, R.; Phan, P.; Polotsky, A.; Lindmark, L.; Frondoza, C.G. Ginger extract inhibits beta-amyloid peptide-induced cytokine and chemokine expression in cultured THP-1 monocytes. J. Altern. Complementary Med. 2004, 10, 1009–1013. [Google Scholar] [CrossRef]
- Ho, S.C.; Chang, Y.H. Comparison of inhibitory capacities of 6-, 8- and 10-gingerols/shogaols on the canonical NLRP3 inflammasome-mediated IL-1β secretion. Molecules 2018, 23, 466. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, Y.; Yu, J. Exosome-like nanoparticles from ginger rhizomes inhibited NLRP3 inflammasome activation. Mol. Pharm. 2019, 16, 2690–2699. [Google Scholar] [CrossRef]
- Petersen, R.C.; Smith, G.E.; Waring, S.C.; Ivnik, R.J.; Tangalos, E.G.; Kokmen, E. Mild cognitive impairment: Clinical characterization and outcome. Arch. Neurol. 1999, 56, 303–308. [Google Scholar] [CrossRef]
- O’Brien, J.T.; Erkinjuntti, T.; Reisberg, B.; Roman, G.; Sawada, T.; Pantoni, L.; Bowler, J.V.; Ballard, C.; DeCarli, C.; Gorelick, P.B.; et al. Vascular cognitive impairment. Lancet. Neurol. 2003, 2, 89–98. [Google Scholar] [CrossRef]
- Zlokovic, B.V. Neurovascular pathways to neurodegeneration in Alzheimer’s disease and other disorders. Nat. Rev. Neurosci. 2011, 12, 723–738. [Google Scholar] [CrossRef] [PubMed]
- Bowler, J.V. Modern concept of vascular cognitive impairment. Br. Med Bull. 2007, 83, 291–305. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Daneman, R.; Prat, A. The blood-brain barrier. Cold Spring Harb. Perspect. Biol. 2015, 7, a020412. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B.V. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef]
- Marshall, R.S. Effects of altered cerebral hemodynamics on cognitive function. J. Alzheimer’s Dis. 2012, 32, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, Y.; Chen, J.P.; Li, D.Z.; Jiang, Q.; Wu, T.; Zhou, X.Z. Oxygen glucose deprivation/re-oxygenation-induced neuronal cell death is associated with Lnc-D63785 m6A methylation and miR-422a accumulation. Cell Death Dis. 2020, 11, 816. [Google Scholar] [CrossRef] [PubMed]
- Faraci, F.M. Reactive oxygen species: Influence on cerebral vascular tone. J. Appl. Physiol. 2006, 100, 739–743. [Google Scholar] [CrossRef] [PubMed]
- Sim, F.J.; Zhao, C.; Penderis, J.; Franklin, R.J. The age-related decrease in CNS remyelination efficiency is attributable to an impairment of both oligodendrocyte progenitor recruitment and differentiation. J. Neurosci. Off. J. Soc. Neurosci. 2002, 22, 2451–2459. [Google Scholar] [CrossRef]
- Freeman, L.R.; Keller, J.N. Oxidative stress and cerebral endothelial cells: Regulation of the blood-brain-barrier and antioxidant based interventions. Biochim. Et Biophys. Acta 2012, 1822, 822–829. [Google Scholar] [CrossRef]
- Ueno, M.; Chiba, Y.; Matsumoto, K.; Murakami, R.; Fujihara, R.; Kawauchi, M.; Miyanaka, H.; Nakagawa, T. Blood-brain barrier damage in vascular dementia. Neuropathol. Off. J. Jpn. Soc. Neuropathol. 2016, 36, 115–124. [Google Scholar] [CrossRef]
- Wattanathorn, J.; Palachai, N.; Thukham-Mee, W.; Muchimapura, S. Memory-enhancing effect of a phytosome containing the combined extract of mulberry fruit and ginger in an animal model of ischemic stroke with metabolic syndrome. Oxidative Med. Cell. Longev. 2020, 2020, 3096826. [Google Scholar] [CrossRef]
- Palachai, N.; Wattanathorn, J.; Muchimapura, S.; Thukham-Mee, W. Phytosome loading the combined extract of mulberry fruit and ginger protects against cerebral ischemia in metabolic syndrome rats. Oxidative Med. Cell. Longev. 2020, 2020, 5305437. [Google Scholar] [CrossRef]
- Wattanathorn, J.; Jittiwat, J.; Tongun, T.; Muchimapura, S.; Ingkaninan, K. Zingiber officinale mitigates brain damage and improves memory impairment in focal cerebral ischemic rat. Evid.-Based Complementary Altern. Med. 2011, 2011, 429505. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, L.; Zhang, N.; Zhang, Q.; Zhao, H.; Zhang, Q. Houshiheisan compound prescription protects neurovascular units after cerebral ischemia. Neural Regen. Res. 2014, 9, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Gaire, B.P.; Kwon, O.W.; Park, S.H.; Chun, K.H.; Kim, S.Y.; Shin, D.Y.; Choi, J.W. Neuroprotective effect of 6-paradol in focal cerebral ischemia involves the attenuation of neuroinflammatory responses in activated microglia. PloS ONE 2015, 10, e0120203. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Model | Compound | Route/Dose | Effects | Reference |
|---|---|---|---|---|
| Molecular docking | Ginger compounds | . | Evaluation of molecular targets based on the measure of the binding energy is AChE. | [41] |
| Molecular docking | Active compounds of ginger extracts | . | Inhibition of AChE activity by the Trp86 choline binding site. | [42] |
| Molecular docking | Ginger compounds | . | Inhibition of BChE activity. | [43] |
| In vitro target enzyme assays, silico docking and simulation of absorption, distribution, metabolism, excretion, and toxicity | Zerumbone | . | Reduction of AChE and BChE activity. | [44] |
| Computational model: koniocortex | Terpenoid-based nutrients | . | Expression of GABA-A receptor and improve cognitive function. | [45] |
| Male Swiss albino mice | Gingerol | Intraperitoneal 10 and 20 mg/kg | Reduction of the levels of Aβ42, β-secretase, APH1a and COX-2. Increased of α-secretase activity. No behavioural and memory deficits. Probable improvement of neuroinflammation and amyloidogenesis. | [46] |
| Sprague Dawley rats stomach | Zo.Cr | 0.03–5.00 mg/mL | Possible calcium antagonism and cholinesterase inhibition. | [47] |
| Male Wistar rats | Extract of Cyperus rotundus and Zingiber officinale (CP1) | Gavage 100, 200 and 300 mg/kg | Oxidative stress reduction in hippocampus. Improved cholinergic function. Improved memory deficit. | [48] |
| Male C57BL/6 mice | fresh ginger, dried ginger, 6-gingerol | Gavage fresh ginger 100 or 200 mg/kg dried ginger 100 or 200 mg/kg 6-gingerol 10 or 25 mg/kg | Upregulation of BDNF. Prevention of memory deficits. | [49] |
| Sprague-Dawley rats | Ginger root extract (GRE) | Intragastric GRE high-dose group 4 gr/kg, GRE medium-dose group 2 gr/kg, GRE low-dose group 1 gr/kg huperzine A group 100 mg/kg | Reduction of inflammatory markers (high-dose group). Improved behavioural tests (high-dose group). | [50] |
| Male APP/PSEN1 mice | Ginger and peony root (OCGP) | Gavage 50 or 100 mg/kg/day 14 weeks | Aβ aggregation inhibition in hippocampus. Behavioural and memory deficits prevention. Reduction of inflammatory marker expression. Reduction of astrocytes activation. | [51] |
| Male ICR mice | 6-shogaol | Gavage 10 mg/kg/day | Reduced astrocytes and microglia activation. Inflammation and neuronal death reduction. Improvement in learning and memory. | [52] |
| APP/PSEN1 mice and HT22 cells | 6-shogaol | Gavage 5 and 20 mg/kg and pre-treatment 10 μM | Improving of behavioural and memory deficits (in vivo). Increased cell availability (in vitro). Inhibition of cytotoxicity Aβ1–42-induced (in vitro). | [53] |
| HT22 cells and APP/PSEN1 mice | 6-shogaol | 1, 5, 10 and 20 mM Gavage 5 or 20 mg/kg every 2 days for 2 months | Aβ aggregation suppression. | [54] |
| Male ICR mice | Ginger + 6-paradol unfermented ginger fermented ginger | Gavage ginger + 6-paradol 100 mg/kg unfermented ginger 100 mg/kg fermented ginger 50,100,200 mg/kg per day | Neuroprotective effect (ginger + 6-paradol). Protection to neurodegeneration in hippocampus (fermented ginger). Improved memory deficits (fermented ginger). | [55] |
| Sprague-Dawley rats | Zerumbone | Intraperitoneal 1 and 10 mg/kg | Improved the memory deficits. Ameliorate anxiety-like behaviours. | [56] |
| Transgenic APP/PSEN1 mice and N9 microglial cells and primary microglial cells from C57BL/6J mice | Zerumbone | Gavage 25 mg/kg and 1, 3 o 10 μg/ml | Improved memory, learning, social interaction, and nesting (in vivo). Reduced Aβ aggregate formation and neuroinflammation in hippocampus and cortex (in vivo). Neuroinflammation suppression (in vitro). | [57] |
| YPEN-1 rat prostate endothelial cells and rat kidney cells from male Fischer 344 rats | Zingerone | YPEN-1 cells pre-treated 1, 5, 10, or 20 μM and rat kidney cells obtained from male rats fed with zingerone 2 or 8 mg/kg/day for 10 days | Reduction in inflammation and oxidative stress. | [58] |
| GT1–7 cells | Ginger essential oil | 25 ppm | Reduction of peroxide-induced neurotoxicity. Enhanced tamoxifen neurotoxicity. | [59] |
| SH-SY5Y cells | [6]-gingerol | Pre-treatment 3, 10 μM | Improved cell viability. Reduction Aβ25–35 citotoxicity. Oxidative stress reduction. | [60] |
| PC12 cells | 6-gingerol | Pre-treatment 40, 80, 120, 200, and 300 μM | Oxidative stress reduction. Decreased Tau hyperphosphorylation. Decreased Aβ formation. | [61] |
| PC12 cells and primary neuronal cells | Ginger extract | 20 g/mL | Improved cell viability. | [62] |
| Rat primary hippocampal cells | Dry ginger | 0.02, 0.1, and 0.2 mg in 2 μL of DMSO | Prevention of Aβ-aggregation. Neurotoxicity reduction. Improved cell survival. | [63] |
| Brain tissue from AD transgenic mice | Ginger extract | Dilutions (1:20, 1:40 and 1:100) | Aggregation of Aβ prevention. | [64] |
| Human monocytic THP-1 cells | Ginger extract composed by Zingiber officinale and Alpinia galangal | Pre-treatment 255 mg | Reduction of inflammation. | [65] |
| THP-1 cells | Shogaols/gingerols | Pre-treatment 20 μM | Reduction of inflammation. | [66] |
| Bone marrow-derived macrophages | Exosome-like nanoparticles of ginger rhizomes | / | Inhibition of inflammasome formation. | [67] |
| Model | Compound | Route, Dose, Timing | Results | Reference |
|---|---|---|---|---|
| Male Wistar rats Rt. MCAO | PMG | Gavage. 50, 100, 200 mg/kg 21 days | Improved cognitive deficits. Reduced AChE activity. Decreased inflammation markers. Increased antioxidant levels. | [80] |
| Male Wistar Rats Rt. MCAO | PMG | Gavage. 50, 100, 200 mg/kg 21 days | Improved cerebral ischemic damage. Improved cerebral edema. Decreased neurological deficits. Inflammation and oxidative stress reduction. | [81] |
| Male Wistar Rats Rt. MCAO | Ginger rhizome | Gavage. 200 mg/kg Before and after surgery | Improved cognitive function. Reduced oxidative stress. Increased hippocampal neuron density. Reduced cerebral infarction volume. | [82] |
| Rat Focal ischemia | Houshiheisan | Gavage. 2.59, 7.7 and 10.5 g/kg Before and after surgery. | Reduced damage to the ischemic penumbra. Vascular protection from Aβ deposition. Stabilization of neurovascular unit. | [83] |
| Mice MCAO/reperfusion and BV2 cells LPS stimulation 100 ng/mL | 6-paradol | Gavage. 1, 5, 10 mg/kg after reperfusion and Pre-treatment 10 ug/mL | Reduced microglia activation. Reduced brain damage. Improved motor and sensory function. Neuroinflammation reduction. Reduction of iNOS. Reduction pro-inflammatory cytokines | [84] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schepici, G.; Contestabile, V.; Valeri, A.; Mazzon, E. Ginger, a Possible Candidate for the Treatment of Dementias? Molecules 2021, 26, 5700. https://doi.org/10.3390/molecules26185700
Schepici G, Contestabile V, Valeri A, Mazzon E. Ginger, a Possible Candidate for the Treatment of Dementias? Molecules. 2021; 26(18):5700. https://doi.org/10.3390/molecules26185700
Chicago/Turabian StyleSchepici, Giovanni, Valentina Contestabile, Andrea Valeri, and Emanuela Mazzon. 2021. "Ginger, a Possible Candidate for the Treatment of Dementias?" Molecules 26, no. 18: 5700. https://doi.org/10.3390/molecules26185700
APA StyleSchepici, G., Contestabile, V., Valeri, A., & Mazzon, E. (2021). Ginger, a Possible Candidate for the Treatment of Dementias? Molecules, 26(18), 5700. https://doi.org/10.3390/molecules26185700