
Antiviral Effects of Green Tea EGCG and Its Potential Application against COVID-19


Abstract
:1. EGCG against DNA Virus
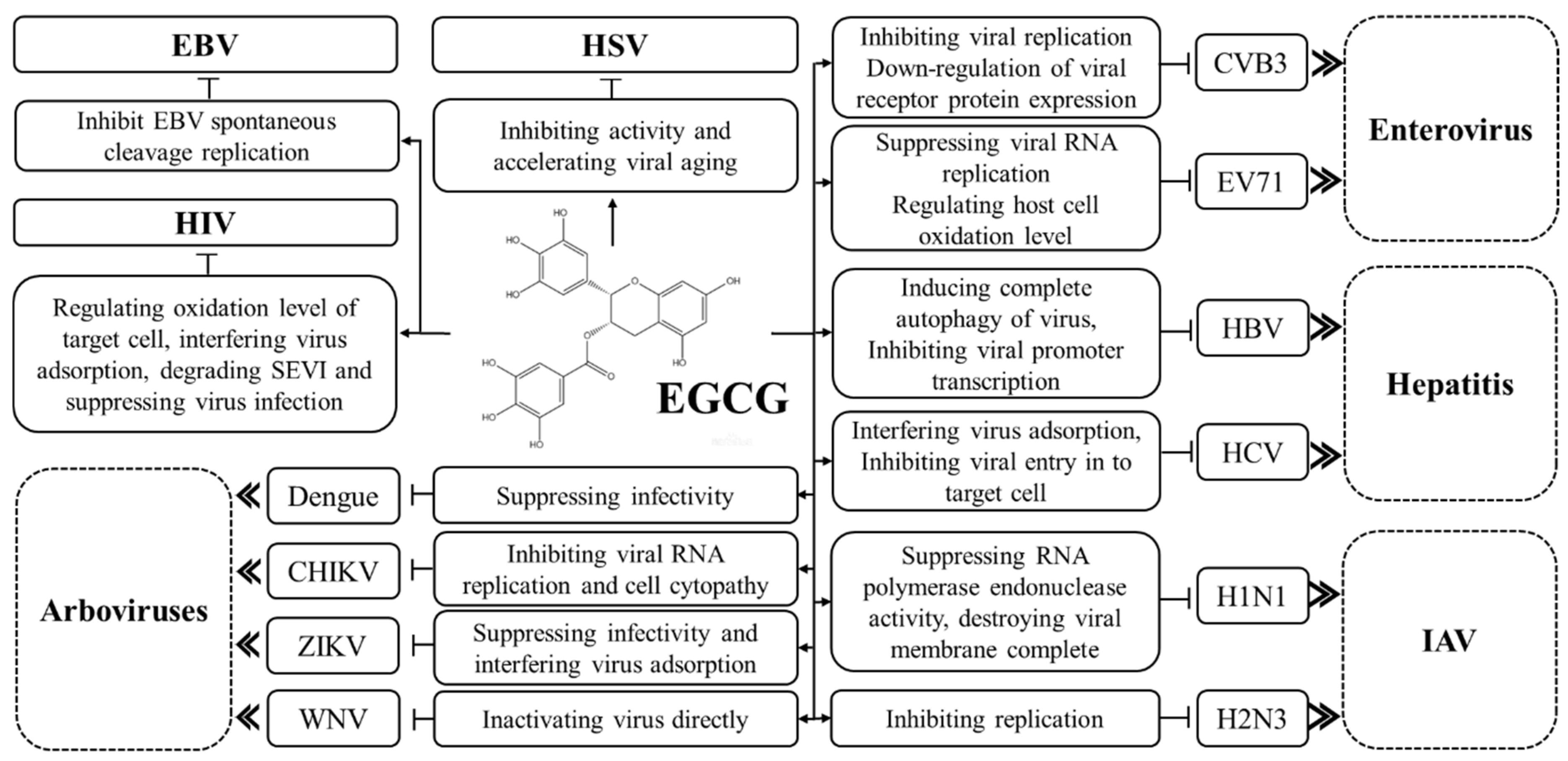
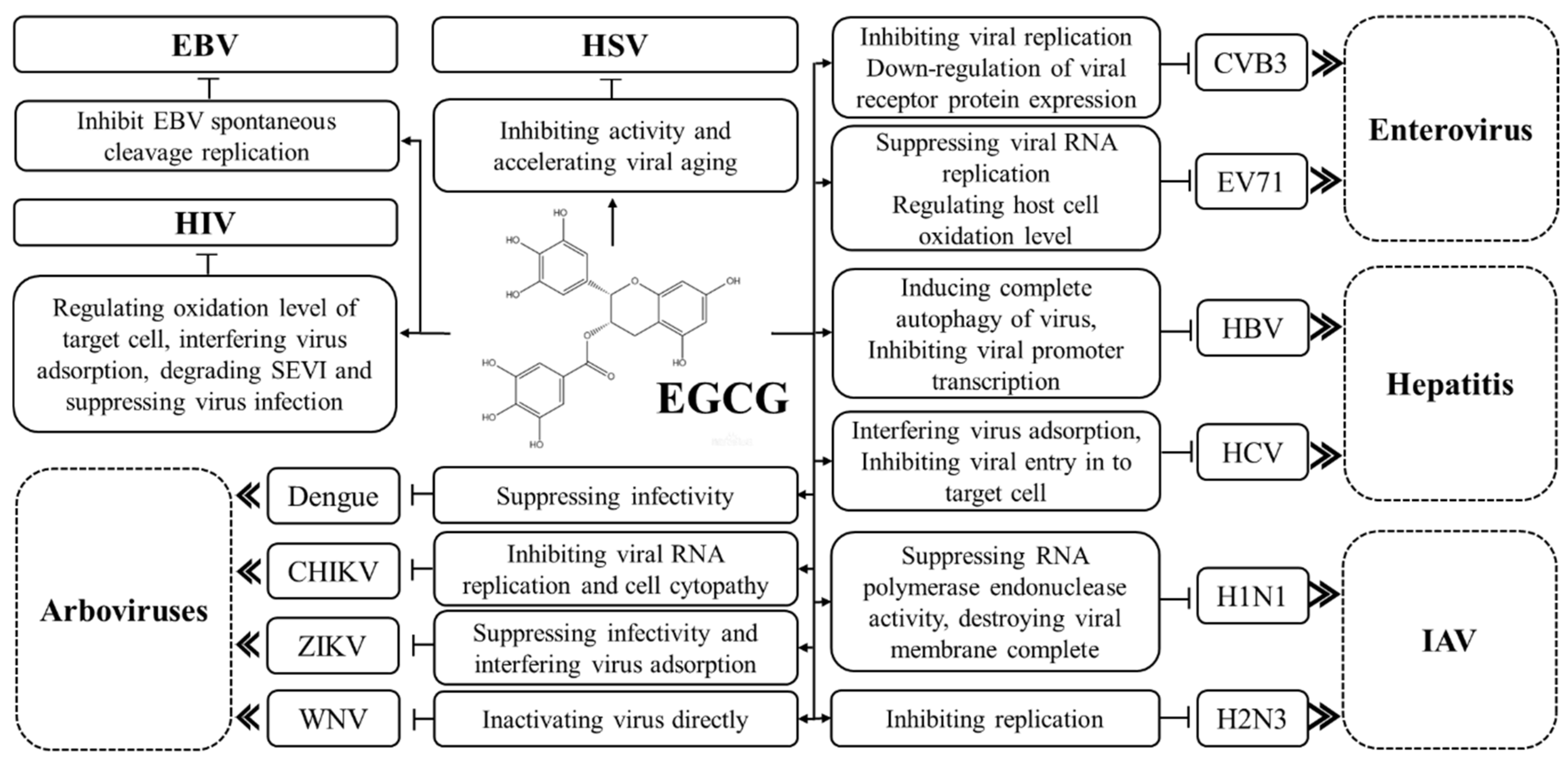
1.1. Effects of EGCG on Hepatitis B Virus
1.2. Effects of EGCG on Herpes Simplex Virus
1.3. Effects of EGCG on Epstein–Barr Virus
2. EGCG against RNA Virus
2.1. Effect of EGCG on Human Immunodeficiency Virus
2.2. Effect of EGCG on Hepatitis C Virus
2.3. Effect of EGCG on Influenza A Virus
3. EGCG against Coronaviruses
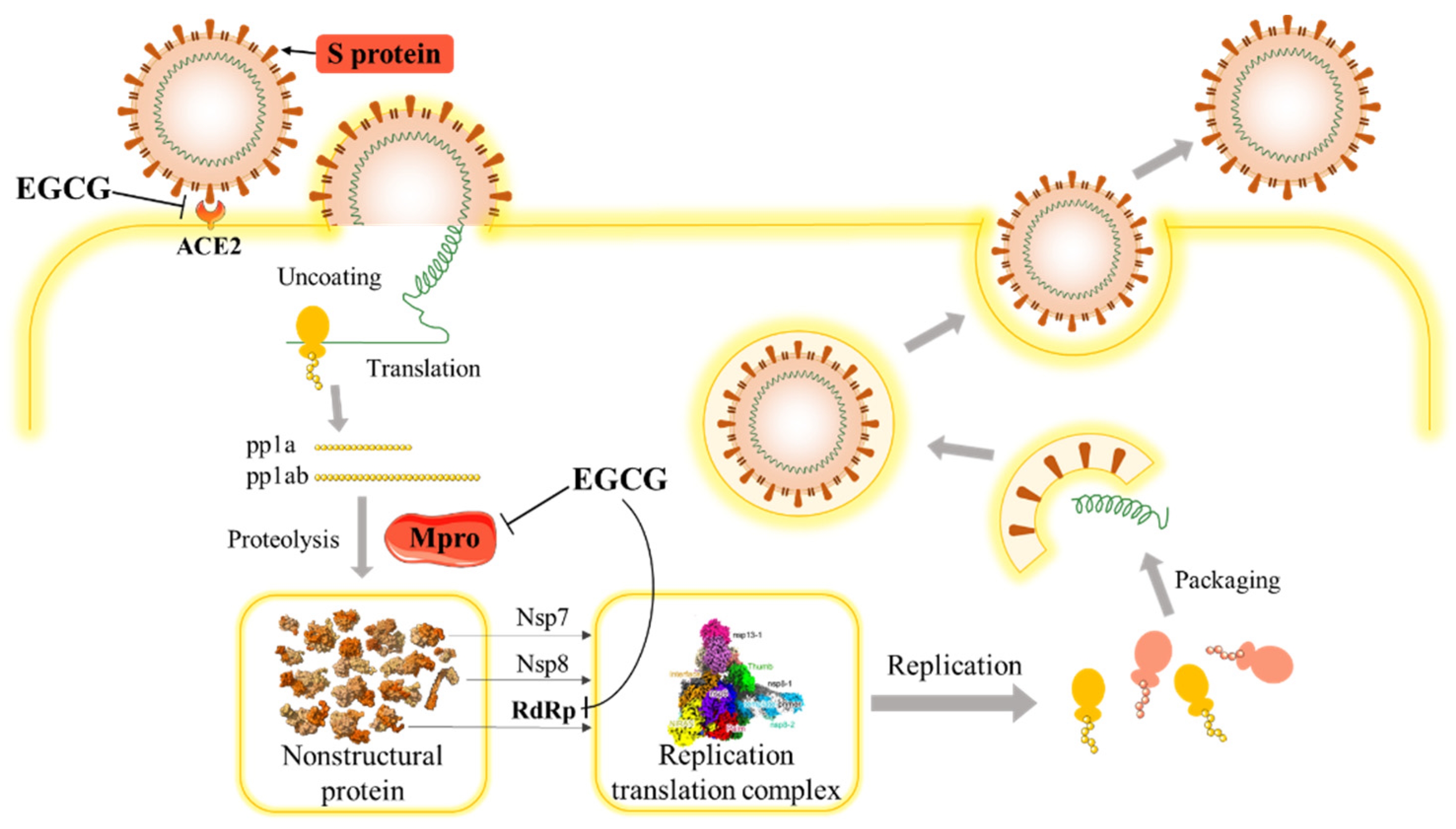
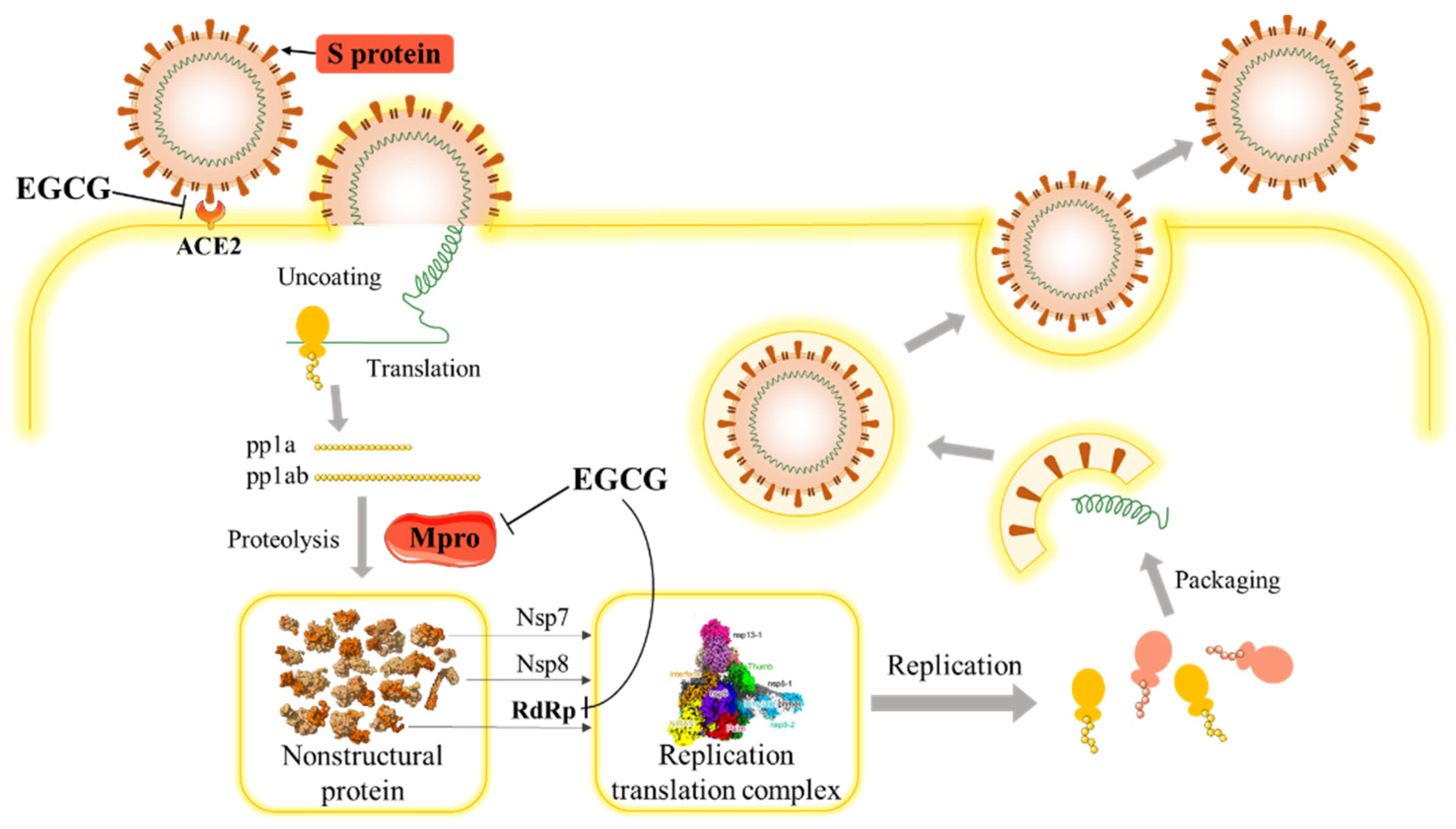
3.1. SARS-CoV-2 and COVID-19
3.2. The Potential Effect of EGCG on SARS-COV-2
4. Effects of EGCG on Enteroviruses
5. Effects of EGCG on Arboviruses
6. Antiviral Effects of EGCG epi-isomer GCG
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, L.; Ho, C.T.; Zhou, J.; Santos, J.S.; Armstrong, L.; Granato, D. Chemistry and Biological Activities of Processed Camellia sinensis Teas: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1474–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Z.M.; Zhong, Y.Z.; Duan, Y.H.; Chen, Q.H.; Li, F.N. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.Y.; Li, Q.S.; Lin, X.M.; Qiao, R.Y.; Yang, R.; Li, X.M.; Dong, Z.B.; Xiang, L.P.; Zheng, X.Q.; Lu, J.L.; et al. Antidiabetic effects of tea. Molecules 2017, 22, 849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polito, C.A.; Cai, Z.Y.; Shi, Y.L.; Li, X.M.; Yang, R.; Shi, M.; Li, Q.S.; Ma, S.C.; Xiang, L.P.; Wang, K.R.; et al. Association of tea consumption with risk of Alzheimer’s disease and anti-beta-amyloid effects of tea. Nutrients 2018, 10, 655. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Q.; Lu, J.L.; Liang, Y.R.; Li, Q.S. Suppressive effects of EGCG on cervical cancer. Molecules 2018, 23, 2334. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.X.; Shi, Y.L.; Zhang, L.J.; Wang, K.R.; Xiang, L.P.; Cai, Z.Y.; Lu, J.L.; Ye, J.H.; Liang, Y.R.; Zheng, X.Q. Inhibitory effects of (–)-Epigallocatechin-3-gallate on esophageal cancer. Molecules 2019, 24, 954. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Y.Y.; Xiang, J.; Wang, Z.S.; Jin, J.; Wang, Y.Q.; Li, Q.S.; Li, D.; Fang, Z.T.; Lu, J.L.; Ye, J.H.; et al. Theacrine from Camellia Kucha and its health beneficial effects. Front. Nutr. 2020, 7. [Google Scholar] [CrossRef]
- Li, Q.S.; Wang, Y.Q.; Liang, Y.R.; Lu, J.L. The anti-allergic potential of tea: A review of its components, mechanisms and risks. Food Funct. 2021, 12, 57–69. [Google Scholar] [CrossRef]
- Wang, X.; Dong, W.; Zhang, X.; Zhu, Z.; Chen, Y.; Liu, X.; Guo, C. Antiviral mechanism of tea polyphenols against porcine reproductive and respiratory syndrome virus. Pathogens 2021, 10, 202. [Google Scholar] [CrossRef]
- Dong, J.J.; Ye, J.H.; Lu, J.L.; Zheng, X.Q.; Liang, Y.R. Isolation of antioxidant catechins from green tea and its decaffeination. Food Bioprod. Process. 2011, 89, 62–66. [Google Scholar] [CrossRef]
- Khan, M.; Adil, S.F.; Alkhathlan, H.Z.; Tahir, M.N.; Saif, S.; Khan, M.; Khan, S.T. COVID-19: A global challenge with old history, epidemiology and progress so far. Molecules 2021, 26, 39. [Google Scholar] [CrossRef]
- Zhou, W.K.; Wang, A.L.; Xia, F.; Xiao, Y.N.; Tang, S.Y. Effects of media reporting on mitigating spread of COVID-19 in the early phase of the outbreak. Math. Biosci. Eng. 2020, 17, 2693–2707. [Google Scholar] [CrossRef]
- World Health Organization. Coronavirus Disease (COVID-19) Pandemic. Last Update: 11 June 2021. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 11 June 2021).
- Gopinath, G. The Great Lockdown: Worst Economic Downturn Since the Great Depression. IMFBlog: Insights and Analysis on Economics and Finance. 14 April 2021. Available online: https://blogs.imf.org/2020/04/14/the-great-lockdown-worst-economic-downturn-since-the-great-depression/ (accessed on 11 June 2021).
- Patel, B.; Sharma, S.; Nair, N.; Majeed, J.; Goyal, R.K.; Dhobi, M. Therapeutic opportunities of edible antiviral plants for COVID-19. Mol. Cell Biochem. 2021, 476, 2345–2364. [Google Scholar] [CrossRef] [PubMed]
- Roschek, B., Jr.; Fink, R.C.; McMichael, M.D.; Li, D.; Alberte, R.S. Elderberry flavonoids bind to and prevent H1N1 infection in vitro. Phytochemistry 2009, 70, 1255–1261. [Google Scholar] [CrossRef]
- Fanunza, E.; Iampietro, M.; Distinto, S.; Corona, A.; Quartu, M.; Maccioni, E.; Horvat, B.; Tramontano, E. Quercetin blocks ebola virus infection by counteracting the vp24 interferon-inhibitory function. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Jo, S.; Kim, H.; Kim, S.; Shin, D.H.; Kim, M.S. Characteristics of flavonoids as potent MERS-CoV 3C-like protease inhibitors. Chem. Biol. Drug Des. 2019, 94, 2023–2030. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Gu, W.; Li, C.; Li, X.; Xing, G.; Li, Y.; Song, Y.; Zheng, W. Epigallocatechin gallate inhibits hepatitis B virus via farnesoid X receptor alpha. J. Nat. Med. 2016, 70, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Hu, J.; Shu, W.; Gao, B.; Xiong, S. Epigallocatechin-3-gallate opposes HBV-induced incomplete autophagy by enhancing lysosomal acidification, which is unfavorable for HBV replication. Cell Death Dis. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, J.Y.; Zhao, K.J.; Wang, J.B.; Ma, Z.J.; Xiao, X.H. Green tea polyphenol, Epigallocatechin-3-gallate, possesses the antiviral activity necessary to fight against the hepatitis B virus replication in vitro. J. Zhejiang Univ. Sci. B 2014, 15, 533–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Sallberg, M.; Hughes, J.; Jones, J.; Guidotti, L.G.; Chisari, F.V.; Billaud, J.N.; Milich, D.R. Immune tolerance split between hepatitis B virus precore and core proteins. J. Virol. 2005, 79, 3016–3027. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Li, L.X.; Liao, Q.J.; Liu, C.L.; Chen, X.L. Epigallocatechin gallate inhibits HBV DNA synthesis in a viral replication-inducible cell line. World J. Gastroenterol. 2011, 17, 1507–1514. [Google Scholar] [CrossRef]
- Isaacs, C.E.; Wen, G.Y.; Xu, W.; Jia, J.H.; Rohan, L.; Corbo, C.; Di Maggio, V.; Jenkins, E.C., Jr.; Hillier, S. Epigallocatechin gallate inactivates clinical isolates of herpes simplex virus. Antimicrob. Agents Chemother. 2008, 52, 962–970. [Google Scholar] [CrossRef] [Green Version]
- Haberichter, J.; Roberts, S.; Abbasi, I.; Dedthanou, P.; Pradhan, P.; Nguyen, M.L. The telomerase inhibitor MST-312 interferes with multiple steps in the herpes simplex virus life cycle. J. Virol. 2015, 89, 9804–9816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, P.; Nguyen, M.L. Herpes simplex virus virucidal activity of MST-312 and Epigallocatechin gallate. Virus Res. 2018, 249, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Gosslau, A.; Jao, D.L.E.; Huang, M.T.; Ho, C.T.; Evans, D.; Rawson, N.E.; Chen, K.Y. Effects of the black tea polyphenol theaflavin-2 on apoptotic and inflammatory pathways in vitro and in vivo. Mol. Nutr. Food Res. 2011, 55, 198–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutok, J.L.; Wang, F. Spectrum of Epstein-Barr virus-associated diseases. Annu. Rev. Pathol. Mech. Dis. 2006, 1, 375–404. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.M.D. Quantitative analysis of Epstein-Barr virus DNA in plasma and serum-applications to tumor detection and monitoring. Ann. N. Y. Acad. Sci. 2001, 945, 68–72. [Google Scholar] [CrossRef]
- Chang, L.K.; Wei, T.T.; Chiu, Y.F.; Tung, C.P.; Chuang, J.Y.; Hung, S.K.; Li, C.; Liu, S.T. Inhibition of Epstein-Barr virus lytic cycle by (–)-Epigallocatechin gallate. Biochem. Biophys. Res. Commun. 2003, 301, 1062–1068. [Google Scholar] [CrossRef]
- Liu, S.; Li, H.; Chen, L.; Yang, L.; Li, L.; Tao, Y.; Li, W.; Li, Z.; Liu, H.; Tang, M.; et al. (–)-Epigallocatechin-3-gallate inhibition of Epstein-Barr virus spontaneous lytic infection involves ERK1/2 and PI3-K/Akt signaling in EBV-positive cells. Carcinogenesis 2013, 34, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.S.; Wu, T.C.; Sang, W.W.; Ruan, Z. EGCG inhibits Tat-induced LTR transactivation: Role of Nrf2, AKT, AMPK signaling pathway. Life Sci. 2012, 90, 747–754. [Google Scholar] [CrossRef]
- Hamza, A.; Zhan, C.G. How can (–)-Epigallocatechin gallate from green tea prevent HIV-1 infection? Mechanistic insights from computational modeling and the implication for rational design of anti-HIV-1 entry inhibitors. J. Phys. Chem. B 2006, 110, 2910–2917. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.P.; McCormick, T.G.; Nance, C.L.; Shearer, W.T. Epigallocatechin gallate, the main polyphenol in green tea, binds to the T-cell receptor, CD4: Potential for HIV-1 therapy. J. Allergy Clin. Immunol. 2006, 118, 1369–1374. [Google Scholar] [CrossRef] [PubMed]
- Castellano, L.M.; Hammond, R.M.; Holmes, V.M.; Weissman, D.; Shorter, J. Epigallocatechin-3-gallate rapidly remodels PAP85-120, SEM1(45-107), and SEM2(49-107) seminal amyloid fibrils. Biol. Open 2015, 4, 1206–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.M.; Qiu, J.Y.; Tan, S.Y.; Liu, S.W.; Li, L. Semen-derived enhancer of viral infection a key factor in sexual transmission of HIV. Chin. J. Virol. 2012, 28, 84–88. [Google Scholar]
- Hauber, I.; Hohenberg, H.; Holstermann, B.; Hunstein, W.; Hauber, J. The main green tea polyphenol Epigallocatechin-3-gallate counteracts semen-mediated enhancement of HIV infection. Proc. Natl. Acad. Sci. USA 2009, 106, 9033–9038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Hattori, T.; Kodama, E.N. Epigallocatechin gallate inhibits the HIV reverse transcription step. Antivir. Chem. Chemother. 2011, 21, 239–243. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.W.; Lu, H.; Zhao, Q.; He, Y.X.; Niu, J.K.; Debnath, A.K.; Wu, S.G.; Jiang, S.B. Theaflavin derivatives in black tea and catechin derivatives in green tea inhibit HIV-1 entry by targeting gp41. BBA-Gen. Subjects 2005, 1723, 270–281. [Google Scholar] [CrossRef]
- Lavanchy, D. Evolving epidemiology of hepatitis C virus. Clin. Microbiol. Infect. 2011, 17, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Salloum, S.; Kluge, S.F.; Kim, A.Y.; Roggendorf, M.; Timm, J. The resistance mutation R155K in the NS3/4A protease of hepatitis C virus also leads the virus to escape from HLA-A*68-restricted CD8 T cells. Antivir. Res. 2010, 87, 272–275. [Google Scholar] [CrossRef]
- Calland, N.; Albecka, A.; Belouzard, S.; Wychowski, C.; Duverlie, G.; Descamps, V.; Hober, D.; Dubuisson, J.; Rouille, Y.; Seron, K. (–)-Epigallocatechin-3-gallate is a new inhibitor of hepatitis C virus entry. Hepatology 2012, 55, 720–729. [Google Scholar] [CrossRef]
- Ciesek, S.; von Hahn, T.; Colpitts, C.C.; Schang, L.M.; Friesland, M.; Steinmann, J.; Manns, M.P.; Ott, M.; Wedemeyer, H.; Meuleman, P.; et al. The green tea polyphenol, epigallocatechin-3-gallate, inhibits hepatitis C virus entry. Hepatology 2011, 54, 1947–1955. [Google Scholar] [CrossRef]
- Okamoto, T.; Nishimura, Y.; Ichimura, T.; Suzuki, K.; Miyamura, T.; Suzuki, T.; Moriishi, K.; Matsuura, Y. Hepatitis C virus RNA replication is regulated by FKBP8 and Hsp90. EMBO J. 2006, 25, 5015–5025. [Google Scholar] [CrossRef] [Green Version]
- Roh, C.; Jo, S.K. (–)-Epigallocatechin gallate inhibits hepatitis C virus (HCV) viral protein NS5B. Talanta 2011, 85, 2639–2642. [Google Scholar] [CrossRef]
- Shiha, G.; Soliman, R.; Elbasiony, M.; Darwish, N.H.E.; Mousa, S.A. Addition of Epigallocatechin gallate 400 mg to Sofosbuvir 400 mg +Daclatisvir 60 mg with or without Ribavirin in treatment of patients with chronic Hepatitis C improves the safety profile: A pilot study. Sci. Rep. 2019, 9, 13593–13601. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.F.; Shao, S.H.; Xu, P.; Yang, X.Q.; Qian, L.S. Catechin-enriched green tea extract as a safe and effective agent for antimicrobial and anti-inflammatory treatment. Afr. J. Pharm. Pharmacol. 2011, 5, 1452–1461. [Google Scholar] [CrossRef] [Green Version]
- Kuzuhara, T.; Iwai, Y.; Takahashi, H.; Hatakeyama, D.; Echigo, N. Green tea catechins inhibit the endonuclease activity of influenza A virus RNA polymerase. PLoS. Curr. 2009, 1, RRN1052. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.X.; Wei, F.; Li, N.; Li, J.L.; Chen, L.J.; Liu, Y.Y.; Luo, F.; Xiong, H.R.; Hou, W.; Yang, Z.Q. Amelioration of influenza virus-induced reactive oxygen species formation by Epigallocatechin gallate derived from green tea. Acta Pharmacol. Sin. 2012, 33, 1533–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Kim, S.Y.; Lee, H.W.; Shin, J.S.; Kim, P.; Jung, Y.S.; Jeong, H.S.; Hyun, J.K.; Lee, C.K. Inhibition of influenza virus internalization by (–)-Epigallocatechin-3-gallate. Antivir. Res. 2013, 100, 460–472. [Google Scholar] [CrossRef]
- Song, J.M.; Lee, K.H.; Seong, B.L. Antiviral effect of catechins in green tea on influenza virus. Antivir. Res. 2005, 68, 66–74. [Google Scholar] [CrossRef]
- Su, S.; Wong, G.; Shi, W.F.; Liu, J.; Lai, A.C.K.; Zhou, J.Y.; Liu, W.J.; Bi, Y.H.; Gao, G.F. Epidemiology, genetic recombination, and pathogenesis of coronaviruses. Trends Microbiol. 2016, 24, 409–502. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.R.; Navas-Martin, S. Coronavirus pathogenesis and the emerging pathogen severe acute respiratory syndrome coronavirus. Microbiol. Mol. Biol. Rev. 2005, 69, 635–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–290. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.L.; Wang, Y.M.; Li, X.W.; Ren, L.L.; Zhao, J.P.; Hu, Y.; Zhang, L.; Fan, G.H.; Xu, J.Y.; Gu, X.Y.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Conti, P.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Frydas, I.; Kritas, S.K. Induction of pro-inflammatory cytokines (IL-1 and IL-6) and lung inflammation by Coronavirus-19 (COVI-19 or SARS-CoV-2): Anti-inflammatory strategies. J. Biol. Regul. Homeost. Agents 2020, 34, 327–331. [Google Scholar] [PubMed]
- Zhang, W.; Zhao, Y.; Zhang, F.C.; Wang, Q.; Li, T.S.; Liu, Z.Y.; Wang, J.L.; Qin, Y.; Zhang, X.; Yan, X.W.; et al. The use of anti-inflammatory drugs in the treatment of people with severe coronavirus disease 2019 (COVID-19): The perspectives of clinical immunologists from China. Clin. Immunol. 2020, 214. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical characteristics of 138 hospitalized patients with 2019 Novel Coronavirus-infected pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Prabakaran, P.; Mao, X.D.; Dimitrov, D.S. A model of the ACE2 structure and function as a SARS-CoV receptor. Biochem. Biophys. Res. Commun. 2004, 314, 235–241. [Google Scholar] [CrossRef]
- Maiti, S.; Banerjee, A. Epigallocatechin gallate and theaflavin gallate interaction in SARS-CoV-2 spike protein central channel with reference to the hydroxychloroquine interaction: Bioinformatics and molecular docking study. Drug Dev. Res. 2020. [Google Scholar] [CrossRef]
- Subbaiyan, A.; Ravichandran, K.; Singh, S.V.; Sankar, M.; Thomas, P.; Dhama, K.; Malik, Y.S.; Singh, R.K.; Chaudhuri, P. In silico molecular docking analysis targeting SARS-CoV-2 spike protein and selected herbal constituents. J. Pure Appl. Microbiol. 2020, 14, 989–998. [Google Scholar] [CrossRef]
- Ghosh, R.; Chakraborty, A.; Biswas, A.; Chowdhuri, S. Evaluation of green tea polyphenols as novel coronavirus (SARS CoV-2) main protease (Mpro) inhibitors—An in silico docking and molecular dynamics simulation study. J. Biomol. Struct. Dyn. 2020, 1–13. [Google Scholar] [CrossRef]
- Upadhyay, S.; Tripathi, P.K.; Singh, M.; Raghavendhar, S.; Bhardwaj, M.; Patel, A.K. Evaluation of medicinal herbs as a potential therapeutic option against SARS-CoV-2 targeting its main protease. Phytother. Res. 2020, 34, 3411–3419. [Google Scholar] [CrossRef]
- Bhardwaj, V.K.; Singh, R.; Sharma, J.; Rajendran, V.; Purohit, R.; Kumar, S. Identification of bioactive molecules from tea plant as SARS-CoV-2 main protease inhibitors. J. Biomol. Struct. Dyn. 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Park, Y.I.; Cha, Y.E.; Park, R.; Namkoong, S.; Lee, J.I.; Park, J. Tea polyphenols EGCG and theaflavin inhibit the activity of SARS-CoV-2 3CL-protease In vitro. Evid. Based Complementary Altern. Med. CAM 2020, 2020, 5630838. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Li, W.; Ashok, M.; Zhu, C.; Sama, A. A major ingredient of green tea rescues mice from lethal sepsis partly by inhibiting HMGB1. PLoS ONE 2007, 2, e1153. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Sk, M.F.; Sonawane, A.; Kar, P.; Sadhukhan, S. Plant-derived natural polyphenols as potential antiviral drugs against SARS-CoV-2 via RNA-dependent RNA polymerase (RdRp) inhibition: An in silico analysis. J. Biomol. Struct. Dyn. 2020. [Google Scholar] [CrossRef] [PubMed]
- Lung, J.; Lin, Y.S.; Yang, Y.H.; Chou, Y.L.; Shu, L.H.; Cheng, Y.C.; Liu, H.T.; Wu, C.Y. The potential chemical structure of anti-SARS-CoV-2 RNA-dependent RNA polymerase. J. Med. Virol. 2020, 92, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.T.; Li, W.; Wang, H.C. More tea for septic patients? Green tea may reduce endotoxin-induced release of high mobility group box 1 and other pro-inflammatory cytokines. Med. Hypotheses 2006, 66, 660–663. [Google Scholar] [CrossRef] [Green Version]
- Wyganowska-Swiatkowska, M.; Nohawica, M.; Grocholewicz, K.; Nowak, G. Influence of herbal medicines on HMGB1 Release, SARS-CoV-2 viral attachment, acute respiratory failure, and sepsis. a literature review. Int. J. Mol. Sci. 2020, 21, 4639. [Google Scholar] [CrossRef]
- He, X.R.; Gao, B.; Zhou, L.; Xiong, S.D. Green tea polyphenol Epigallocatechin-3-gallate-alleviated Coxsackievirus B3-induced myocarditis through inhibiting viral replication but not through inhibiting inflammatory responses. J. Cardiovasc. Pharmacol. 2017, 69, 41–47. [Google Scholar] [CrossRef]
- Ho, H.Y.; Cheng, M.L.; Weng, S.F.; Leu, Y.L.; Chiu, D.T.Y. Antiviral effect of Epigallocatechin gallate on Enterovirus 71. J. Agric. Food Chem. 2009, 57, 6140–6147. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, B.M.; Batista, M.N.; Braga, A.C.S.; Nogueira, M.L.; Rahal, P. The green tea molecule EGCG inhibits Zika virus entry. Virology 2016, 496, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.; Silva, K.; von Rhein, C.; Kuemmerer, B.M.; Schnierle, B.S. The green tea catechin, epigallocatechin gallate inhibits Chikungunya virus infection. Antivir. Res. 2015, 113, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.W.; Hsieh, P.S.; Lin, C.C.; Hu, M.K.; Huang, S.M.; Wang, Y.M.; Liang, C.Y.; Gong, Z.Y.; Ho, Y.J. Synergistic effects of combination treatment using EGCG and Suramin against the Chikungunya virus. Biochem. Biophys. Res. Commun. 2017, 491, 595–602. [Google Scholar] [CrossRef]
- Vazquez-Calvo, A.; de Oya, N.J.; Martin-Acebes, M.A.; Garcia-Moruno, E.; Saiz, J.C. Antiviral properties of the natural polyphenols Delphinidin and Epigallocatechin gallate against the flaviviruses West Nile virus, Zika virus, and Dengue virus. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Fassina, G.; Buffa, A.; Benelli, R.; Varnier, O.E.; Noonan, D.M.; Albini, A. Polyphenolic antioxidant (–)-epigallocatechin-3-gallate from green tea as a candidate anti-HIV agent. Aids 2002, 16, 939–941. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Sharma, N.; Aarthy, M.; Singh, S.K.; Giri, R. Mechanistic insights into Zika virus NS3 helicase inhibition by Epigallocatechin-3-gallate. ACS Omega 2020, 5, 11217–11226. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Honda, M.; Ikigai, H.; Hara, Y.; Shimamura, T. Inhibitory effects of (–)-Epigallocatechin gallate on the life cycle of human immunodeficiency virus type 1 (HIV-1). Antivir. Res. 2002, 53, 19–34. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Compound | Total Binding Energy (kcal/mol) | Van der Waal’s Force (kcal/mol) | H-Bond Energy (kcal/mol) | AverCon Pair (kcal/mol) |
|---|---|---|---|---|
| EGCG | −130.566 | −91.7244 | −38.8417 | 25.7273 |
| Curcumin | −115.198 | −87.5695 | −27.6288 | 26.963 |
| Apigenin | −108.614 | −82.1108 | −26.503 | 33.9 |
| Chrysophanol | −107.385 | −90.5916 | −16.7935 | 36.7895 |
| Emodin | −105.462 | −87.2314 | −18.2303 | 32.85 |
| Zingerone | −102.184 | −77.9523 | −24.2321 | 26.9524 |
| Gingerol | −98.0333 | −84.1818 | −13.8515 | 27.6667 |
| Ursolic acid | −89.9499 | −72.2658 | −17.6841 | 23.5714 |
| Ajoene | −74.2819 | −68.2819 | −6 | 32.4615 |
| Aloe emodin | −69.2503 | −69.2503 | 0 | 29.2308 |
| Allicin | −62.4326 | −62.4326 | 0 | 40.3333 |
| Diallyltrisulfide | −53.2872 | −53.2872 | 0 | 39.2222 |
| Virus | Cell Line | Effect | Reference |
|---|---|---|---|
| CHIKV | U2OS | Inhibiting viral RNA replication | [75] |
| CHIKV | HEK 293T | Blocking viral entry into target cells | [74] |
| CVB3 | Vero | Inhibiting viral RNA replication, improving the survival rate of infected cells | [72] |
| EBV | B95.8 | Down regulating RNA synthesis, inhibiting EBV lytic protein expression | [31] |
| HBV | HepG2.2.15 | Inhibiting HBs Ag and HBe Ag secretion | [21] |
| HBV | HepG2.2.15 | Opposing HBV-induced incomplete autophagy, reducing HBV replication | [20] |
| HBV | HepG2.N10 | Interfering core promoter transcription | [19] |
| HBV | HepG2.117 | Inhibiting RNA, DNA and cccDNA synthesis | [23] |
| HCV | Huh7.5 | Interfering virus adsorption, preventing cell-to-cell transmission | [43] |
| HCV | Huh7.19 | Inhibiting viral entry in to target cell | [42] |
| HIV | MAGI | Reducing Nf-kB expression and suppressing HIV-1 gene transcription | [32] |
| HIV | peripheral blood lymphocytes | Inhibiting viral replication | [77] |
| HIV | HeLa-CD4-LTR-beta-gal | Suppressing viral infection ability, inhibiting viral reverse transcription | [38] |
| HIV | THP-1 | Inhibiting viral transcription | [78] |
| HSV | Vero, CV-1 | In-activating viral | [24] |
| ZIKA | Vero E6 | Inhibiting viral entry in to target cell | [73] |
| H1N1 | HEK 293T | Destroying viral membrane integrity, preventing viral adsorption to cell surface | [50] |
| H1N1, | MDCK | Inhibiting erythrocyte agglutination and viral replication | [51] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.-Q.; Li, Q.-S.; Zheng, X.-Q.; Lu, J.-L.; Liang, Y.-R. Antiviral Effects of Green Tea EGCG and Its Potential Application against COVID-19. Molecules 2021, 26, 3962. https://doi.org/10.3390/molecules26133962
Wang Y-Q, Li Q-S, Zheng X-Q, Lu J-L, Liang Y-R. Antiviral Effects of Green Tea EGCG and Its Potential Application against COVID-19. Molecules. 2021; 26(13):3962. https://doi.org/10.3390/molecules26133962
Chicago/Turabian StyleWang, Ying-Qi, Qing-Sheng Li, Xin-Qiang Zheng, Jian-Liang Lu, and Yue-Rong Liang. 2021. "Antiviral Effects of Green Tea EGCG and Its Potential Application against COVID-19" Molecules 26, no. 13: 3962. https://doi.org/10.3390/molecules26133962
APA StyleWang, Y.-Q., Li, Q.-S., Zheng, X.-Q., Lu, J.-L., & Liang, Y.-R. (2021). Antiviral Effects of Green Tea EGCG and Its Potential Application against COVID-19. Molecules, 26(13), 3962. https://doi.org/10.3390/molecules26133962