
Identification of 3-Methoxycarpachromene and Masticadienonic Acid as New Target Inhibitors against Trypanothione Reductase from Leishmania Infantum Using Molecular Docking and ADMET Prediction


Abstract

1. Introduction
2. Results and Discussion
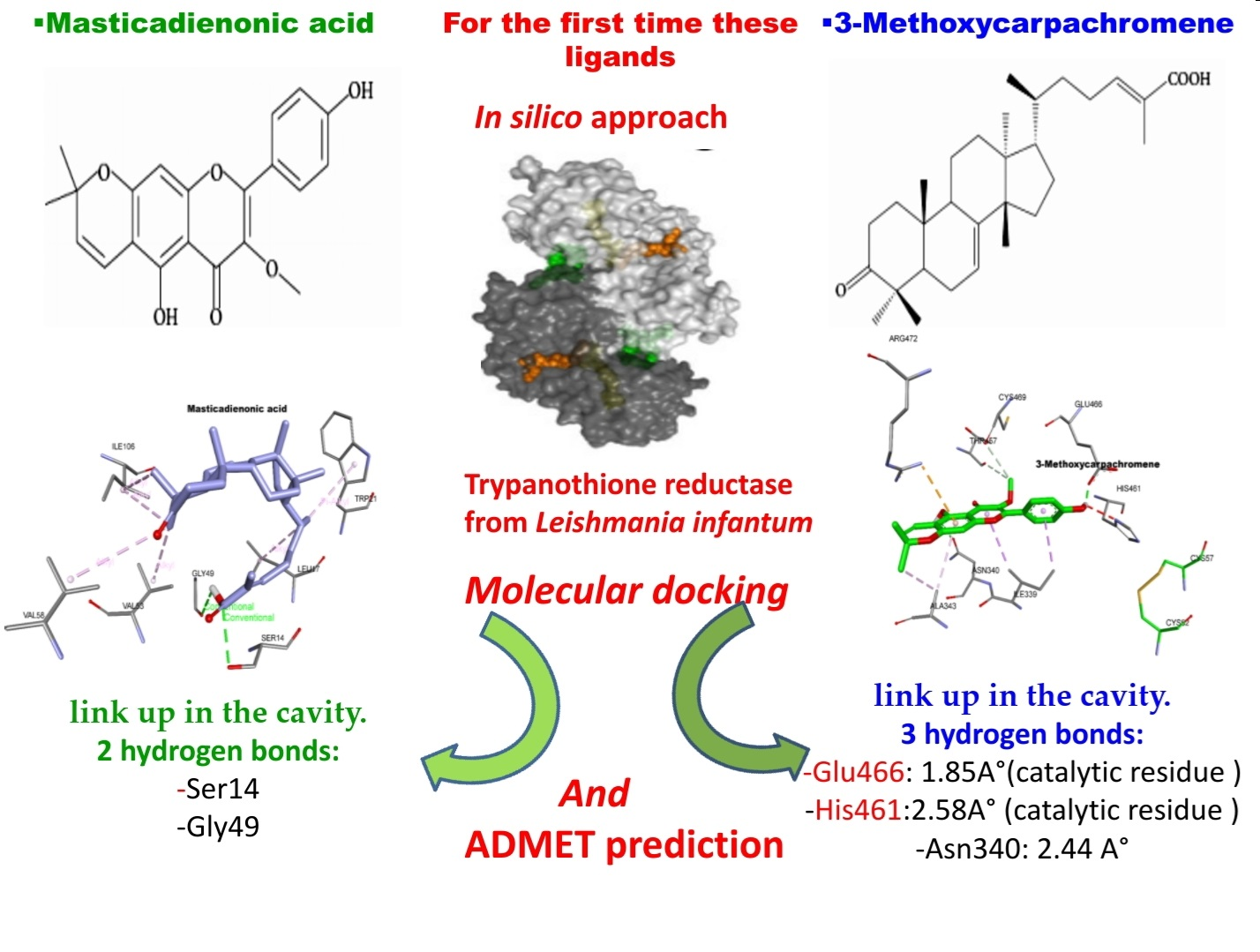
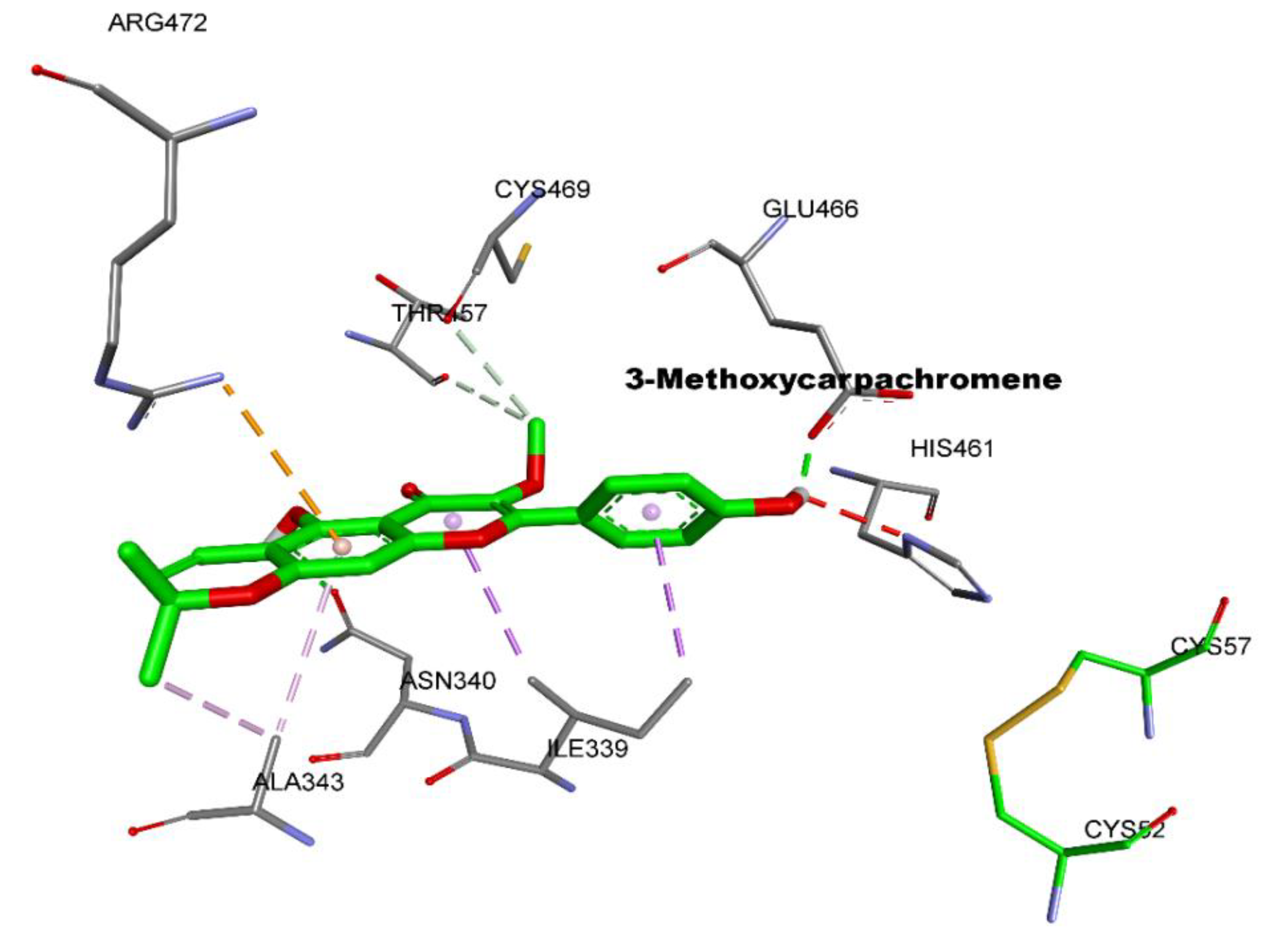
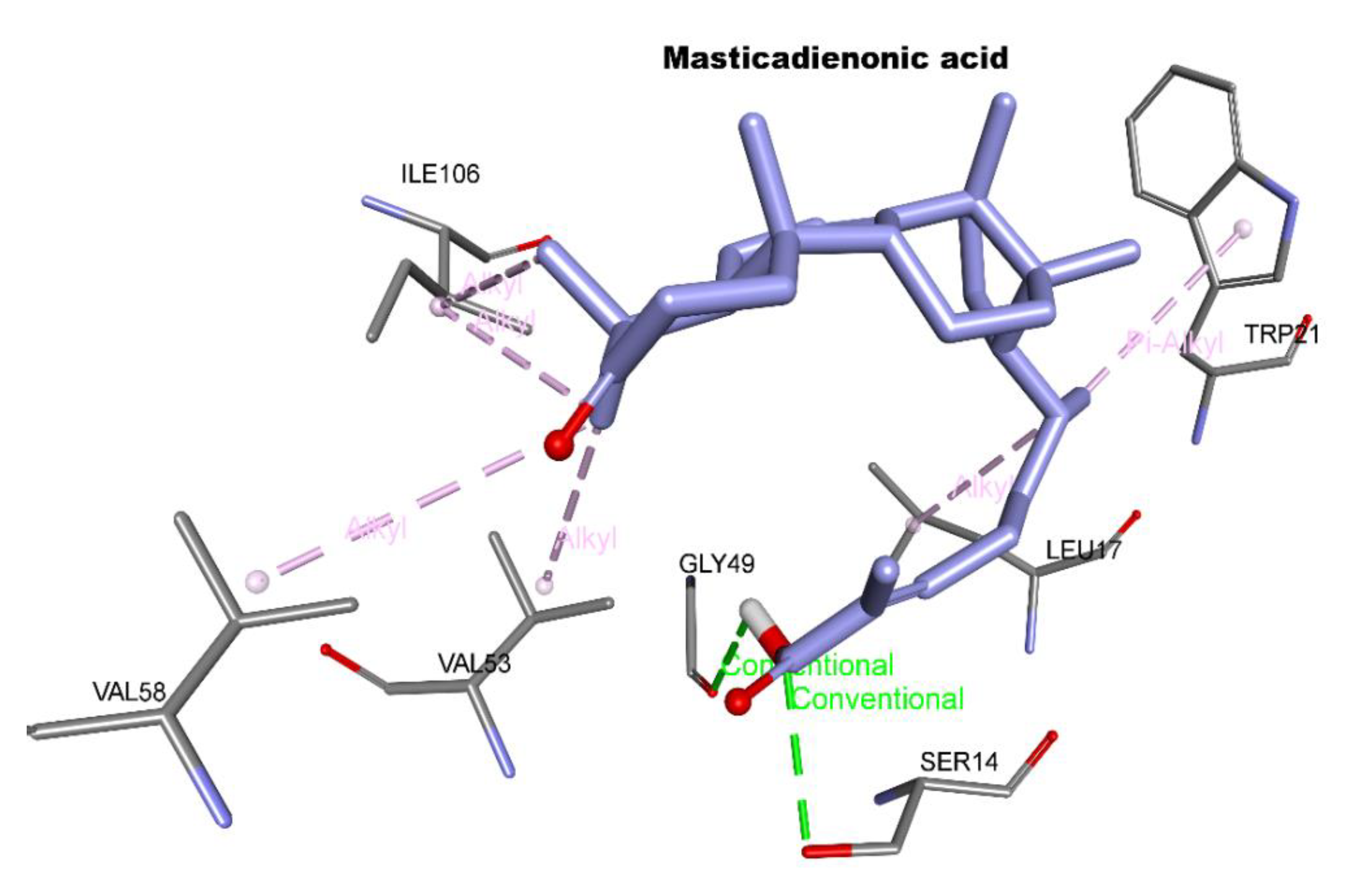
2.1. Molecular Docking
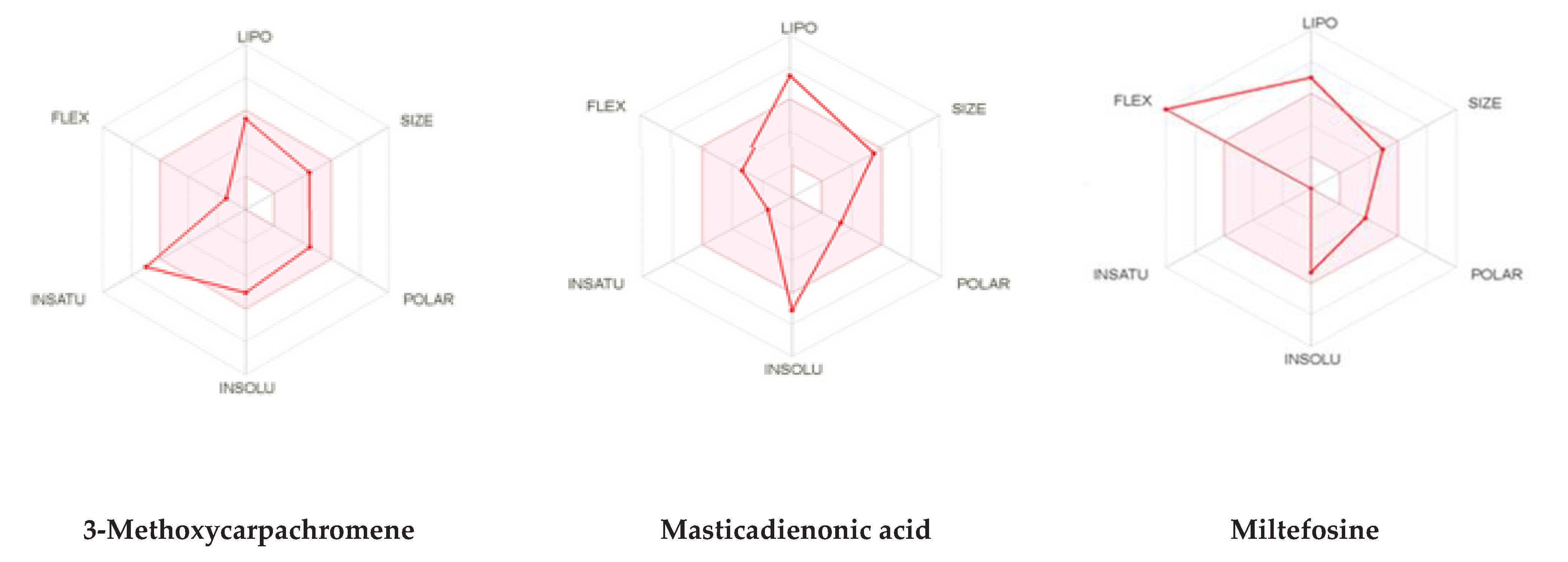
2.2. ADMET Study
3. Materials and Methods
3.1. Molecular Docking
3.2. ADMET Study
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Akbari, M.; Oryan, A.; Hatam, G. Application of nanotechnology in treatment of leishmaniasis: A review. Acta Trop. 2017, 172, 86–90. [Google Scholar] [CrossRef]
- Soosaraei, M.; Khasseh, A.A.; Fakhar, M.; Hezarjaribi, H.Z. A decade bibliometric analysis of global research on leishmaniasis in Web of Science database. Ann. Med. Surg. 2018, 26, 30–37. [Google Scholar] [CrossRef]
- Ikeogu, N.M.; Akaluka, G.N.; Edechi, C.A.; Salako, E.S.; Onyilagha, C.; Barazandeh, A.F.; Uzonna, J.E. Leishmania immunity: Advancing immunotherapy and vaccine development. Microorganisms 2020, 8, 1201. [Google Scholar] [CrossRef] [PubMed]
- Eddaikra, N.; Ait-Oudhia, K.; Kherrachi, I.; Oury, B.; Moulti-Mati, F.; Benikhlef, R.; Harrat, Z.; Sereno, D. Antimony susceptibility of Leishmania isolates collected over a 30-year period in Algeria. PLoS Negl. Trop. Dis. 2018, 12, e0006310. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, N.; Qureshi, N.A.; Iqbal, A.; Ashraf, A.; Fatima, H. Evaluation of Safety, Antileishmanial, and Chemistry of Ethanolic Leaves Extracts of Seven Medicinal Plants: An In-vitro Study. Open Chem. J. 2020, 7, 26–36. [Google Scholar] [CrossRef]
- Camacho, M.; Phillipson, J.; Croft, S.; Solis, P.; Marshall, S.; Ghazanfar, S. Screening of plant extracts for antiprotozoal and cytotoxic activities. J. Ethnopharmacol. 2003, 89, 185–191. [Google Scholar] [CrossRef]
- Berbert, T.R.N.; de Mello, T.F.P.; Wolf Nassif, P.; Mota, C.A.; Silveira, A.V.; Duarte, G.C.; Demarchi, I.G.; Aristides, S.M.A.; Lonardoni, M.V.C.; Vieira Teixeira, J.J. Pentavalent antimonials combined with other therapeutic alternatives for the treatment of cutaneous and mucocutaneous leishmaniasis: A systematic review. Dermatol. Res. Pract. 2018, 2018, 1–21. [Google Scholar] [CrossRef]
- Guery, R.; Henry, B.; Martin-Blondel, G.; Rouzaud, C.; Cordoliani, F.; Harms, G.; Gangneux, J.-P.; Foulet, F.; Bourrat, E.; Baccard, M. Liposomal amphotericin B in travelers with cutaneous and muco-cutaneous leishmaniasis: Not a panacea. PLoS Negl. Trop. Dis. 2017, 11, e0006094. [Google Scholar] [CrossRef] [PubMed]
- Wiwanitkit, V. Interest in paromomycin for the treatment of visceral leishmaniasis (kala-azar). Ther. Clin. Risk Manag. 2012, 8, 323–328. [Google Scholar] [CrossRef]
- Raj, S.; Sasidharan, S.; Balaji, S.N.; Dubey, V.K.; Saudagar, P. Review on natural products as an alternative to contemporary anti-leishmanial therapeutics. J. Proteins Proteom. 2020, 11, 135–158. [Google Scholar] [CrossRef]
- Lira, R.; Sundar, S.; Makharia, A.; Kenney, R.; Gam, A.; Saraiva, E.; Sacks, D. Evidence that the High Incidence of Treatment Failures in Indian Kala-Azar Is Due to the Emergence of Antimony-Resistant Strains ofLeishmaniadonovani. J. Infect. Dis. 1999, 180, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Perry, M.R.; Wyllie, S.; Raab, A.; Feldmann, J.; Fairlamb, A.H. Chronic exposure to arsenic in drinking water can lead to re-sistance to antimonial drugs in a mouse model of visceral leishmaniasis. Proc. Natl. Acad. Sci. USA 2013, 110, 19932–19937. [Google Scholar] [CrossRef]
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M.; WHO Leishmaniasis Control Team. Leishmaniasis Worldwide and Global Estimates of Its Incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef]
- Mukherjee, B.; Mukherjee, K.; Nanda, P.; Mukhopadhayay, R.; Ravichandiran, V.; Bhattacharyya, S.N.; Roy, S. Probing the molecular mechanism of aggressive infection by antimony resistant Leishmania donovani. Cytokine 2020, 155245. [Google Scholar] [CrossRef]
- Ogungbe, I.V.; Erwin, W.R.; Setzer, W.N. Antileishmanial phytochemical phenolics: Molecular docking to potential protein targets. J. Mol. Graph. Model. 2014, 48, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Fairlamb, A.H.; Blackburn, P.; Ulrich, P.; Chait, B.T.; Cerami, A. Trypanothione: A novel bis(glutathionyl)spermidine cofactor for glutathione reductase in trypanosomatids. Science 1985, 227, 1485–1487. [Google Scholar] [CrossRef]
- Schmidt, A.; Krauth-Siegel, R.L. Enzymes of the Trypanothione Metabolism as Targets for Antitrypanosomal Drug Development. Curr. Top. Med. Chem. 2002, 2, 1239–1259. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.F.; Castro-Pinto, D.; Echevarria, A.; dos Reis, C.M.; DelCistia, C.N.; Sant’Anna, C.M.R.; Teixeira, F.; Castro, H.; Canto-Cavalheiro, M.; Leon, L.L.; et al. Investigation of trypanothione reductase inhibitory activity by 1,3,4-thiadiazolium-2-aminide derivatives and molecular docking studies. Bioorg. Med. Chem. 2012, 20, 1760–1766. [Google Scholar] [CrossRef] [PubMed]
- Colotti, G.; Baiocco, P.; Fiorillo, A.; Boffi, A.; Poser, E.; Chiaro, F.D.; Ilari, A. Structural insights into the enzymes of the tryp-anothione pathway: Targets for antileishmaniasis drugs. Future Med. Chem. 2013, 5, 1861–1875. [Google Scholar] [CrossRef]
- Moreno-Izquierdo, M.A.; Alonso, A.; Alcolea, P.J.; Abramov, A.; De Lacoba, M.G.; Abendroth, J.; Zhang, S.; Edwards, T.; Lorimer, D.; Myler, P.J.; et al. Tyrosine aminotransferase from Leishmania infantum: A new drug target candidate. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 347–354. [Google Scholar] [CrossRef]
- Khademvatan, S.; Eskandari, K.; Hazrati-Tappeh, K.; Rahim, F.; Foroutan, M.; Yousefi, E.; Asadi, N. In silico and in vitro comparative activity of green tea components against Leishmania infantum. J. Glob. Antimicrob. Resist. 2019, 18, 187–194. [Google Scholar] [CrossRef]
- Prada, C.F.; Álvarez-Velilla, R.; Balana-Fouce, R.; Prieto, C.; Alvarez, E.C.; Escudero-Martínez, J.M.; Requena, J.M.; Ordóñez, C.; Desideri, A.; Pérez-Pertejo, Y.; et al. Gimatecan and other camptothecin derivatives poison Leishmania DNA-topoisomerase IB leading to a strong leishmanicidal effect. Biochem. Pharmacol. 2013, 85, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Gundampati, R.K.; Chandrasekaran, S.; Jagannadham, M.V. Molecular docking study on the interaction between trypano-thione reductase and mangiferin for antileishmanial activity. Bangladesh J. Pharmacol. 2013, 8, 40–43. [Google Scholar]
- Chelghoum, M.; Guenane, H.; Harrat, M.; Yousfi, M. Total Tocopherols, Carotenoids, and Fatty Acids Content Variation of Pistacia atlantica from Different Organs’ Crude Oils and Their Antioxidant Activity during Development Stages. Chem. Biodivers. 2020, 2000117. [Google Scholar] [CrossRef]
- Labdelli, A.; Zemour, K.; Simon, V.; Cerny, M.; Adda, A.; Merah, O. Pistacia AtlanticaDesf., a Source of Healthy Vegetable Oil. Appl. Sci. 2019, 9, 2552. [Google Scholar] [CrossRef]
- Ben Ahmed, Z.; Yousfi, M.; Viaene, J.; Dejaegher, B.; Demeyer, K.; Heyden, Y.V. Four Pistacia atlantica subspecies (atlantica, cabulica, kurdica and mutica): A review of their botany, ethnobotany, phytochemistry and pharmacology. J. Ethnopharmacol. 2021, 265, 113329. [Google Scholar] [CrossRef] [PubMed]
- Mahjoub, F.; Rezayat, K.A.; Yousefi, M.; Mohebbi, M.; Salari, R. Pistacia atlanticaDesf. A review of its traditional uses, phy-tochemicals and pharmacology. J. Med. Life 2018, 11, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Bozorgi, M.; Memariani, Z.; Mobli, M.; Surmaghi, M.H.S.; Shams-Ardekani, M.R.; Rahimi, R. FivePistaciaspecies (P. vera, P.atlantica, P. terebinthus, P. khinjuk, and P. lentiscus): A Review of Their Traditional Uses, Phytochemistry, and Pharmacology. Sci. World J. 2013, 2013, 1–33. [Google Scholar] [CrossRef]
- Nachvak, S.M.; Hosseini, S.; Nili-Ahmadabadi, A.; Dastan, D.; Rezaei, M. Chemical composition and antioxidant activity of Pistacia atlantica subsp. Kurdica from Awraman. J. Rep. Pharm. Sci. 2018, 7, 222–230. [Google Scholar]
- Ogungbe, I.V.; Setzer, W.N. In-silico Leishmania Target Selectivity of Antiparasitic Terpenoids. Molecules 2013, 18, 7761–7847. [Google Scholar] [CrossRef] [PubMed]
- Bernal, F.; Coy-Barrera, E. In-Silico Analyses of Sesquiterpene-Related Compounds on Selected Leishmania Enzyme-Based Targets. Molecules 2014, 19, 5550–5569. [Google Scholar] [CrossRef] [PubMed]
- Morais, T.R.; Da Costa-Silva, T.A.; Tempone, A.G.; Borborema, S.E.T.; Scotti, M.T.; De Sousa, R.M.F.; Araujo, A.C.C.; De Oliveira, A.; De Morais, S.A.L.; Sartorelli, P.; et al. Antiparasitic Activity of Natural and Semi-Synthetic Tirucallane Triterpenoids from Schinus terebinthifolius (Anacardiaceae): Structure/Activity Relationships. Molecules 2014, 19, 5761–5776. [Google Scholar] [CrossRef]
- Sharifi, M.S.; Hazell, S.L. Isolation, Analysis and Antimicrobial Activity of the Acidic Fractions of Mastic, Kurdica, Mutica and Cabolica Gums from Genus Pistacia. Glob. J. Health Sci. 2011, 4, 217–228. [Google Scholar] [CrossRef]
- Adams, M.; Plitzko, I.; Kaiser, M.; Brun, R.; Hamburger, M. HPLC-profiling for antiplasmodial com-pounds—3-Methoxycarpachromene from Pistacia atlantica. Phytochem. Lett. 2009, 2, 159–162. [Google Scholar] [CrossRef]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef]
- Venkatesan, S.K.; Shukla, A.K.; Dubey, V.K. Molecular docking studies of selected tricyclic and quinone derivatives on tryp-anothione reductase of Leishmania infantum. J. Comput. Chem. 2010, 31, 2463–2475. [Google Scholar]
- Battista, T.; Colotti, G.; Ilari, A.; Fiorillo, A. Targeting Trypanothione Reductase, a Key Enzyme in the Redox Trypanosomatid Metabolism, to Develop New Drugs against Leishmaniasis and Trypanosomiases. Molecules 2020, 25, 1924. [Google Scholar] [CrossRef]
- Bailey, S.; Smith, K.; Fairlamb, A.H.; Hunter, W.N. Substrate interactions between trypanothione reductase and N1-glutathionylspermidine disulphide at 0.28-nm resolution. JBIC J. Biol. Inorg. Chem. 1993, 213, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Patterson, S.; Alphey, M.S.; Jones, D.C.; Shanks, E.J.; Street, I.P.; Frearson, J.A.; Wyatt, P.G.; Gilbert, I.H.; Fairlamb, A.H. Di-hydroquinazolines as a novel class of Trypanosoma brucei trypanothione reductase inhibitors: Discovery, synthesis, and characterization of their binding mode by protein crystallography. J. Med. Chem. 2011, 54, 6514–6530. [Google Scholar] [CrossRef]
- Saravanamuthu, A.; Vickers, T.J.; Bond, C.S.; Peterson, M.R.; Hunter, W.N.; Fairlamb, A.H. Two interacting binding sites for quinacrine derivatives in the active site of trypanothione reductase: A template for drug design. J. Biol. Chem. 2004, 279, 29493–29500. [Google Scholar] [CrossRef] [PubMed]
- Turcano, L.; Torrente, E.; Missineo, A.; Andreini, M.; Gramiccia, M.; DiMuccio, T.; Genovese, I.; Fiorillo, A.; Harper, S.; Bresciani, A.; et al. Identification and binding mode of a novel Leishmania Trypanothione reductase inhibitor from high throughput screening. PLoS Negl. Trop. Dis. 2018, 12, e0006969. [Google Scholar] [CrossRef] [PubMed]
- Baiocco, P.; Poce, G.; Alfonso, S.; Cocozza, M.; Porretta, G.C.; Colotti, G.; Biava, M.; Moraca, F.; Botta, M.; Yardley, V.; et al. Inhibition ofLeishmaniainfantumTrypanothione Reductase by Azole-Based Compounds: A Comparative Analysis with Its Physiological Substrate by X-ray Crystallography. ChemMedChem 2013, 8, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Krieger, S.; Schwarz, W.; Ariyanayagam, M.R.; Fairlamb, A.H.; Krauth-Siegel, R.L.; Clayton, C. Trypanosomes lacking trypanothione reductase are avirulent and show increased sensitivity to oxidative stress. Mol. Microbiol. 2002, 35, 542–552. [Google Scholar] [CrossRef]
- Dumas, C.; Ouellette, M.; Tovar, J.; Cunningham, M.L.; Fairlamb, A.H.; Tamar, S.; Olivier, M.; Papadopoulou, B. Disruption of the trypanothione reductase gene of Leishmania decreases its ability to survive oxidative stress in macrophages. EMBO J. 1997, 16, 2590–2598. [Google Scholar] [CrossRef] [PubMed]
- Tovar, J.; Wilkinson, S.; Mottram, J.C.; Fairlamb, A.H. Evidence that trypanothione reductase is an essential enzyme in Leishmania by targeted replacement of the tryA gene locus. Mol. Microbiol. 1998, 29, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Tovar, J.; Cunningham, M.L.; Smith, A.C.; Croft, S.L.; Fairlamb, A.H. Down-regulation of Leishmania donovani trypanothione reductase by heterologous expression of a trans-dominant mutant homologue: Effect on parasite intracellular survival. Proc. Natl. Acad. Sci. USA 1998, 95, 5311–5316. [Google Scholar] [CrossRef]
- Müller, S.; Liebau, E.; Walter, R.D.; Krauth-Siegel, R.L. Thiol-based redox metabolism of protozoan parasites. Trends Parasitol. 2003, 19, 320–328. [Google Scholar] [CrossRef]
- Bond, C.S.; Zhang, Y.; Berriman, M.; Cunningham, M.L.; Fairlamb, A.H.; Hunter, W.N. Crystal structure of Trypanosoma cruzi trypanothione reductase in complex with trypanothione, and the structure-based discovery of new natural product inhibitors. Structure 1999, 7, 81–89. [Google Scholar] [CrossRef]
- Meanwell, N.A. Improving Drug Design: An Update on Recent Applications of Efficiency Metrics, Strategies for Replacing Problematic Elements, and Compounds in Nontraditional Drug Space. Chem. Res. Toxicol. 2016, 29, 564–616. [Google Scholar] [CrossRef] [PubMed]
- Saccoliti, F.; Angiulli, G.; Pupo, G.; Pescatori, L.; Madia, V.N.; Messore, A.; Colotti, G.; Fiorillo, A.; Scipione, L.; Gramiccia, M.; et al. Inhibition of Leishmania infantum trypanothione reductase by diaryl sulfide derivatives. J. Enzym. Inhib. Med. Chem. 2017, 32, 304–310. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem substance and compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina:Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar]
- Martins, C.; Carreiras, M.C.; Leon, R.; Ríos, C.D.L.; Bartolini, M.; Andrisano, V.; Iriepa, I.; Moraleda, I.; Gálvez, E.; Garcia, M.; et al. Synthesis and biological assessment of diversely substituted furo[2,3-b]quinolin-4-amine and pyrrolo[2,3-b]quinolin-4-amine derivatives, as novel tacrine analogues. Eur. J. Med. Chem. 2011, 46, 6119–6130. [Google Scholar] [CrossRef] [PubMed]
- Nickel, J.; Gohlke, B.; Erehman, J.; Banerjee, P.; Rong, W.W.; Goede, A.; Dunkel, M.; Preissner, R. SuperPred: Update on drug classification and target prediction. Nucleic Acids Res. 2014, 42, W26–W31. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Enzyme | References | Number of Published Papers (Google Scholar) |
|---|---|---|
| Tyrosine aminotransferase | [20] | 298 |
| Trypanothione synthetase | [21] | 1340 |
| DNA topoisomerases | [22] | 1400 |
| Trypanothione reductase | [23] | 1790 |
| Ligand Name | 2D Structure | References |
|---|---|---|
| Masticadienonic acid |  | [33] |
| 3-Methoxycarpa-chromene |  | [34] |
| Ligand | Free Binding Energy (kcal mol−1) | Closest Residues | Hydrophobic Interactions | Hydrogen Bonds | Length (Å) |
|---|---|---|---|---|---|
| 3-Methoxycarpachromene | −8.4 | Glu466, His461, Asn340, Arg472, Cys469,Thr457, Ile339, Ala343 | Π-Alkyl, Π-sigma, Π-cation, alkyl-alkyl | Glu466 His461 Asn340 | 1.85 2.58 2.44 |
| Masticadienonic acid | −6.2 | Trp21, Leu17, Gly49, Val53, Val58, Ile106, Ser14 | Π-Alkyl, alkyl-alkyl | Ser14 Gly49 | 2.96 2.9 |
| Models | 3-Methoxycarpachromene | Masticadienonic Acid | Miltefosine |
|---|---|---|---|
| A. Absorption | |||
| Blood–Brain Barrier | No | No | No |
| Human Intestinal Absorption | high | low | low |
| Skin Permeation | −5.60 cm/s | −3.68 cm/s | −3.97 cm/s |
| B. Metabolism | |||
| P-gp Substrate | Non Substrate | Non Substrate | yes |
| CYP450 1A2 Inhibitor | Non Inhibitor | Non Inhibitor | Non Inhibitor |
| CYP450 2C19 Inhibitor | Inhibitor | Non Inhibitor | Non Inhibitor |
| CYP450 2C9 Inhibitor | Non Inhibitor | Inhibitor | Non Inhibitor |
| CYP450 2D6 Inhibitor | Non Inhibitor | Non Inhibitor | Non Inhibitor |
| CYP450 2C19 Inhibitor | Non Inhibitor | Non Inhibitor | Non Inhibitor |
| CYP450 3A4 Inhibitor | Inhibitor | Non Inhibitor | Non Inhibitor |
| Lipinski Rule | Accepted | Accepted | Accepted |
| C.Toxicity | |||
| HERG_inhibition | Non | Non | Non |
| Ames test | Non mutagen | Non mutagen | Non mutagen |
| Carcinogenicity (Mouse) | Yes | Yes | Non |
| Carcinogenicity (Rat) | Non | Non | Non |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maamri, S.; Benarous, K.; Yousfi, M. Identification of 3-Methoxycarpachromene and Masticadienonic Acid as New Target Inhibitors against Trypanothione Reductase from Leishmania Infantum Using Molecular Docking and ADMET Prediction. Molecules 2021, 26, 3335. https://doi.org/10.3390/molecules26113335
Maamri S, Benarous K, Yousfi M. Identification of 3-Methoxycarpachromene and Masticadienonic Acid as New Target Inhibitors against Trypanothione Reductase from Leishmania Infantum Using Molecular Docking and ADMET Prediction. Molecules. 2021; 26(11):3335. https://doi.org/10.3390/molecules26113335
Chicago/Turabian StyleMaamri, Sarra, Khedidja Benarous, and Mohamed Yousfi. 2021. "Identification of 3-Methoxycarpachromene and Masticadienonic Acid as New Target Inhibitors against Trypanothione Reductase from Leishmania Infantum Using Molecular Docking and ADMET Prediction" Molecules 26, no. 11: 3335. https://doi.org/10.3390/molecules26113335
APA StyleMaamri, S., Benarous, K., & Yousfi, M. (2021). Identification of 3-Methoxycarpachromene and Masticadienonic Acid as New Target Inhibitors against Trypanothione Reductase from Leishmania Infantum Using Molecular Docking and ADMET Prediction. Molecules, 26(11), 3335. https://doi.org/10.3390/molecules26113335