
Gibberellin Increases the Bud Yield and Theanine Accumulation in Camellia sinensis (L.) Kuntze

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Dynamic Accumulation of Gibberellin and Theanine During Tea Bud Germination and Elongation
2.2. Gibberellin Treatment Promotes the Lateral Bud Germination and Elongation
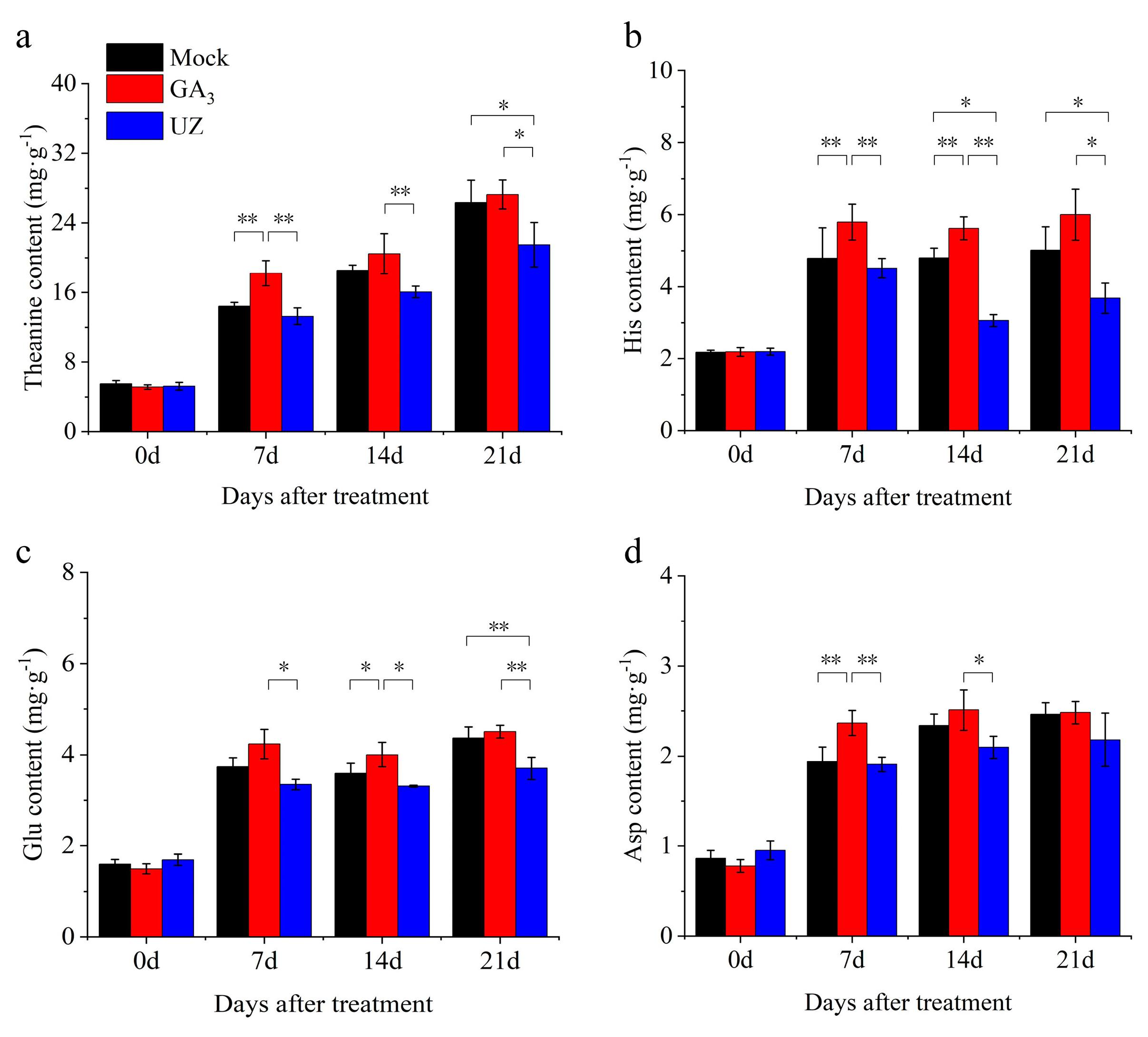
2.3. Gibberellin Treatment Increased the Theanine Content in Tea Buds and Leaves
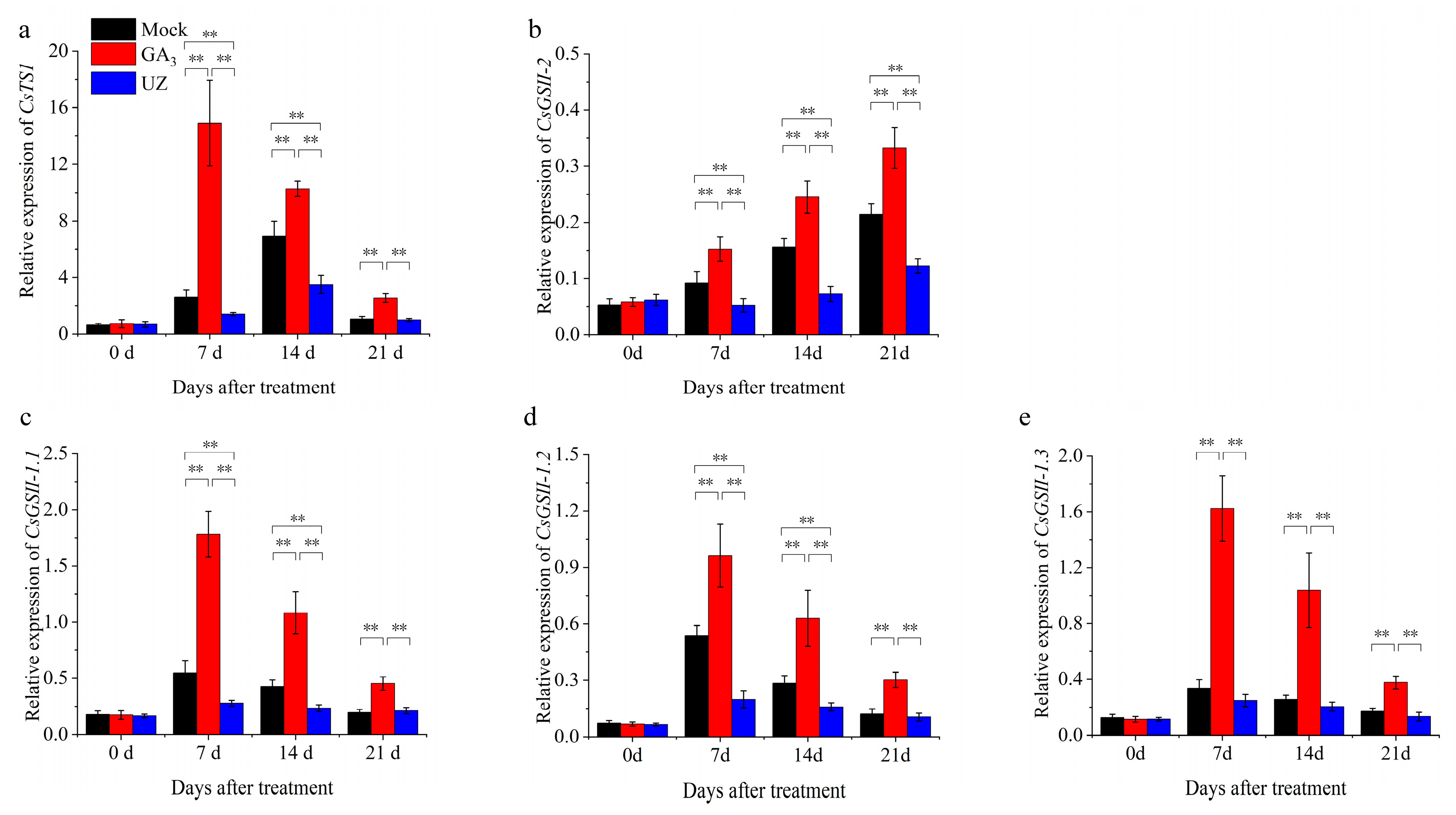
2.4. Effects of GA3 on Gene Expression in Biosynthesis of Theanine in Tea Plants
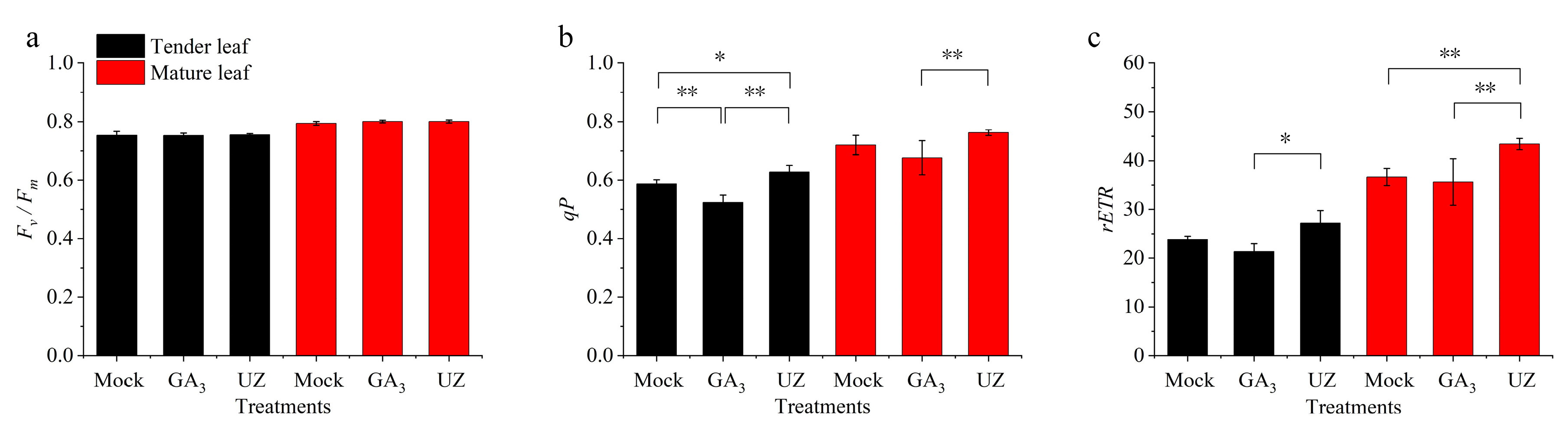
2.5. Gibberellin Affect the Chlorophyll Fluorescence Parameters in Tea Leaves
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Materials and Treatments
4.3. Sampling
4.4. Free Amino Acids Determination
4.5. RNA Extraction and Quantitative Real-Time PCR
4.6. Gibberellin Extraction and Quantification
4.7. Chlorophyll Fluorescence Determination
4.8. Chlorophyll Determination
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Anneleen, S.; Katia, V.; Ellen, L.; Joeri, V.L.; Yvette, M.; Ilse, S.; Ann, M. L-Theanine intake increases threshold for limbic seizures but decreases threshold for generalized seizures. Nutr. Neurosci. 2013, 16, 78–82. [Google Scholar]
- Li, W.; Xiang, F.; Zhong, M.; Zhou, L.; Liu, H.; Li, S.; Wang, X. Transcriptome and metabolite analysis identifies nitrogen utilization genes in tea plant (Camellia sinensis). Sci. Rep. 2017, 7, 1693. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, L.; Zhang, L.; Yan, P.; Ahammed, G.; Han, W. Methyl salicylate enhances flavonoid biosynthesis in tea leaves by stimulating the phenylpropanoid pathway. Molecules 2019, 24, 362. [Google Scholar] [CrossRef]
- Li, J.; Yang, Y.; Sun, K.; Chen, Y.; Chen, X.; Li, X. Exogenous melatonin enhances cold, salt and drought stress tolerance by improving antioxidant defense in tea plant (Camellia sinensis (L.) O. Kuntze). Molecules 2019, 24, 1826. [Google Scholar] [CrossRef]
- Kraujalyte, V.; Pelvan, E.; Alasalvar, C. Volatile compounds and sensory characteristics of various instant teas produced from black tea. Food Chem. 2016, 194, 864–872. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Huang, H.; Zhao, X.; Zhong, N.; Zheng, H.; Gong, Y. Distinct variation in taste quality of Congou black tea during a single spring season. Food Sci. Nutr. 2020, 8, 1848–1856. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, J. Japanese Tea Breeding History and the Future Perspective; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Wang, L.; Wei, K.; Cheng, H.; He, W.; Li, X.; Gong, W. Geographical tracing of Xihu Longjing tea using high performance liquid chromatography. Food Chem. 2014, 146, 98–103. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, M.; Ruan, J. Metabolomics analysis reveals the metabolic and functional roles of flavonoids in light-sensitive tea leaves. BMC Plant Biol. 2017, 17, 64. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Xia, E.; Lu, Y.; Tai, Y.; She, G.; et al. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, 4151–4158. [Google Scholar] [CrossRef]
- Eshita, S.; Robin, J.; Ashu, G. L-Theanine: An astounding sui generis integrant in tea. Food Chem. 2018, 242, 601–610. [Google Scholar]
- Wen, B.; Li, J.; Luo, Y.; Zhang, X.; Wang, K.; Liu, Z.; Huang, J. Identification and expression profiling of MYB transcription factors related to L-theanine biosynthesis in Camellia sinensis. Int. J. Biol. Macromol. 2020, 164, 4306–4317. [Google Scholar] [CrossRef]
- Oh, K.; Kato, T.; Xu, H.L. Transport of nitrogen assimilation in xylem vessels of green tea plants fed with NH4-N and NO3-N. Pedosphere 2008, 18, 222–226. [Google Scholar] [CrossRef]
- Narukawa, M.; Morita, K.; Hayashi, Y. L-theanine elicits an umami taste with inosine 5’-monophosphate. Biosci. Biotechnol. Biochem. 2008, 72, 3015–3017. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Fu, X.; Wang, X.; Liao, Y.; Yang, Z. Studies on the biochemical formation pathway of the amino acid l-theanine in tea (Camellia sinensis) and other plants. J. Agric. Food Chem. 2017, 65, 7210–7216. [Google Scholar] [CrossRef] [PubMed]
- Ashihara, H. Occurrence, biosynthesis and metabolism of theanine (γ-glutamyl-l-ethylamide) in plants: A comprehensive review. Nat. Prod. Commun. 2015, 10, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wu, Z.; Li, H.; Wang, Y.; Zhuang, J. L-theanine content and related gene expression: Novel insights into theanine biosynthesis and hydrolysis among different tea plant (Camellia sinensis L.) tissues and cultivars. Front. Plant Sci. 2017, 8, 498. [Google Scholar] [CrossRef]
- Shao, C.; Zhang, C.; Lv, Z.; Shen, C. Pre- and post-harvest exposure to stress influence quality-related metabolites in fresh tea leaves (Camellia sinensis). Sci. Hortic. 2021, 281, 109984. [Google Scholar] [CrossRef]
- Xia, E.; Zhang, H.; Sheng, J.; Li, K.; Zhang, Q.; Kim, C.; Zhang, Y.; Liu, Y.; Zhu, T.; Li, W. The tea tree genome provides insights into tea flavor and independent evolution of caffeine biosynthesis. Mol. Plant 2017, 10, 866–877. [Google Scholar] [CrossRef]
- Ruan, J.Y.; Gerendás, J.; Härdter, R.; Sattelmacher, B. Effect of root zone pH and form and concentration of nitrogen on accumulation of quality-related components in green tea. J. Sci. Food Agric. 2007, 87, 1505–1516. [Google Scholar] [CrossRef]
- Deng, W.; Fei, Y.; Wang, S.; Wan, X.; Zhang, Z.; Hu, X. Effect of shade treatment on theanine biosynthesis in Camellia sinensis seedlings. Plant Growth Regul. 2013, 71, 295–299. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, M.; Ruan, J. Integrated transcriptome and metabolic analyses reveals novel insights into free amino acid metabolism in huangjinya tea cultivar. Front. Plant Sci. 2017, 8, 291. [Google Scholar] [CrossRef]
- Asano, K.; Yamasaki, M.; Takuno, S.; Miura, K.; Katagiri, S.; Ito, T.; Doi, K.; Wu, J.; Ebana, K.; Matsumoto, T.; et al. Artificial selection for a green revolution gene during japonica rice domestication. Proc. Natl. Acad. Sci. USA 2011, 108, 11034–11039. [Google Scholar] [CrossRef]
- Tsuda, K.; Abraham-Juarez, M.J.; Maeno, A.; Dong, Z.; Aromdee, D.; Meeley, R.; Shiroishi, T.; Nonomura, K.I.; Hake, S. KNOTTED1 cofactors, BLH12 and BLH14, regulate internode patterning and vein anastomosis in maize. Plant Cell 2017, 29, 1105–1118. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Derose, R.; Suarez, A.; Ueno, T.; Chen, M.; Sun, T.P.; Wolfgang, M.J.; Mukherjee, C.; Meyers, D.J.; Inoue, T. Rapid and orthogonal logic gating with a gibberellin-induced dimerization system. Nat. Chem. Biol. 2012, 8, 465–470. [Google Scholar] [CrossRef]
- Vaistij, F.E.; Barros-Galvao, T.; Cole, A.F.; Gilday, A.D.; He, Z.; Li, Y.; Harvey, D.; Larson, T.R.; Graham, I.A. MOTHER-OF-FT-AND-TFL1 represses seed germination under far-red light by modulating phytohormone responses in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2018, 115, 8442–8447. [Google Scholar] [CrossRef]
- Yu, L.; Shi, Y.; Xiao, H.; Liu, F.; Liu, Z. Dynamic changes of endogenous GA3 and ABA contents in tea cultivars with different phenological characters and their impact on the regulation axillary buds sprouting. Acta Agro. Sin. 2008, 34, 277–283. [Google Scholar] [CrossRef]
- Cosgrove, D.J.; Sovonick-Dunford, S.A. Mechanism of gibberellin-dependent stem elongation in peas. Plant Physiol. 1989, 89, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Bulley, S.M.; Wilson, F.M.; Hedden, P.; Phillips, A.L.; Croker, S.J.; James, D.J. Modification of gibberellin biosynthesis in the grafted apple scion allows control of tree height independent of the rootstock. Plant Biotechnol. J. 2005, 3, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Li, J.; Liu, Y.; Zhang, Q.; Gao, Y.; Fang, K.; Cao, Q.; Qin, L.; Xing, Y. Roles of the GA-mediated SPL gene family and miR156 in the floral development of Chinese Chestnut (Castanea mollissima). Int. J. Mol. Sci. 2019, 20, 1577. [Google Scholar] [CrossRef] [PubMed]
- Rizza, A.; Jones, A.M. The makings of a gradient: Spatiotemporal distribution of gibberellins in plant development. Curr. Opin. Plant Biol. 2019, 47, 9–15. [Google Scholar] [CrossRef]
- Gòmez-Maldonado, J.; Cánovas, F.M.; Avila, C. Molecular analysis of the 5’-upstream region of a gibberellin-inducible cytosolic glutamine synthetase gene (GS1b) expressed in pine vascular tissue. Planta 2004, 218, 1036–1045. [Google Scholar] [CrossRef]
- El-Yazal, M.A.S.; El-Yazal, S.A.S.; Rady, M.M. Exogenous dormancy-breaking substances positively change endogenous phytohormones and amino acids during dormancy release in ‘Anna’ apple trees. Plant Growth Regul. 2014, 72, 211–220. [Google Scholar] [CrossRef]
- Zhang, Q.; Xiang, J.; Zhang, L.; Zhu, X.; Jochem, E.; van der Werf, W.; Duan, L. Optimizing soaking and germination conditions to improve gamma-aminobutyric acid content in japonica and indica germinated brown rice. J. Funct. Foods 2014, 10, 283–291. [Google Scholar] [CrossRef]
- Huang, Y.; Cai, S.; Ye, L.; Hu, H.; Li, C.; Zhang, G. The effects of GA and ABA treatments on metabolite profile of germinating barley. Food Chem. 2016, 192, 928–933. [Google Scholar] [CrossRef]
- Komatsu, S.; Zang, X.; Tanaka, N. Comparison of two proteomics techniques used to identify proteins regulated by gibberellin in rice. J. Proteome Res. 2006, 5, 270–276. [Google Scholar] [CrossRef]
- Nagar, P.K.; Kumar, A. Changes in endogenous gibberellin activity during winter dormancy in tea (Camellia sinensis (L.) O. Kuntze). Acta Physiol. Plant 2000, 22, 439–443. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Liu, H.; Lin, S.-J.; Han, M.-H.; Zhuang, J. Genomic analyses of the crosstalk between gibberellins and brassinosteroids metabolisms in tea plant (Camellia sinensis (L.) O. Kuntze). Sci. Hortic. 2020, 268, 109368. [Google Scholar] [CrossRef]
- Anzala, F.; Paven, M.-C.M.r.-L.; Fournier, S.; Rondeau, D.; Limami, A.M. Physiological and molecular aspects of aspartate-derived amino acid metabolism during germination and post-germination growth in two maize genotypes differing in germination efficiency. J. Exp. Bot. 2006, 57, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Garciarrubio, A.; Covarrubias, L.A.A. Abscisic acid inhibits germination of mature Arabidopsis seeds by limiting the availability of energy and nutrients. Planta 1997, 203, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, P.; Gu, M.; Lin, X.; Hou, B.; Zheng, Y.; Sun, Y.; Jin, S.; Ye, N. R2R3-MYB transcription factor family in tea plant (Camellia sinensis): Genome-wide characterization, phylogeny, chromosome location, structure and expression patterns. Genomics 2021, 113, 1565–1578. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Dong, C.; Yang, T.; Ma, J.; Zhang, Z. Seasonal theanine accumulation and related gene expression in the roots and leaf buds of tea plants (Camellia sinensis L.). Front. Plant Sci. 2019, 10, 1397. [Google Scholar] [CrossRef] [PubMed]
- Judd, M.J.; Meyer, D.H.; Meekings, J.S.; Richardson, A.C.; Walton, E.F. An FTIR study of the induction and release of kiwifruit buds from dormancy. J. Sci. Food Agric. 2010, 90, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Mi, X.; Guo, R.; Xia, X.; Liu, L.; An, Y.; Yan, X.; Wang, S.; Guo, L.; Wei, C. The biosynthesis of main taste compounds is coordinately regulated by mirnas and phytohormones in tea plant (Camellia sinensis). J. Agric. Food Chem. 2020, 68, 6221–6236. [Google Scholar] [CrossRef] [PubMed]
- Lam, H.M.; Coschigano, K.T.; Oliveira, I.C.; Melo-Oliveira, R.; Coruzzi, G.M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Z.; Li, P.; Zhang, Q. Determination for major chemical contaminants in tea (Camellia sinensis) matrices: A review. Food Res. Int. 2013, 53, 649–658. [Google Scholar] [CrossRef]
- Serrani, J.C.; Sanjuán, R.; Ruiz-Rivero, O.; Fos, M.; García-Martínez, J.L. Gibberellin regulation of fruit set and growth in tomato. Plant Physiol. 2007, 145, 246–257. [Google Scholar] [CrossRef]
- Deng, T.; Wu, D.; Duan, C.; Yan, X.; Du, Y.; Zou, J.; Guan, Y. Spatial profiling of gibberellins in a single leaf based on microscale matrix solid-phase dispersion and precolumn derivatization coupled with ultraperformance liquid chromatography-tandem mass spectrometry. Anal. Chem. 2017, 89, 9537–9543. [Google Scholar] [CrossRef]
- Li, D.; Guo, Z.; Liu, C.; Li, J.; Xu, W. Quantification of near-attomole gibberellins in floral organs dissected from a single Arabidopsis thaliana flower. Plant J. 2017, 91, 547–557. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chlorophyll a + b Content | Quenching Coefficient | Theanine Content | |
| Chlorophyll a + b content | 1 | ||
| Quenching coefficient | 0.999 * | 1 | |
| Theanine content | −0.980 * | −0.990 * | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Xiang, F.; Su, Y.; Luo, Z.; Luo, W.; Zhou, L.; Liu, H.; Xiao, L. Gibberellin Increases the Bud Yield and Theanine Accumulation in Camellia sinensis (L.) Kuntze. Molecules 2021, 26, 3290. https://doi.org/10.3390/molecules26113290
Li W, Xiang F, Su Y, Luo Z, Luo W, Zhou L, Liu H, Xiao L. Gibberellin Increases the Bud Yield and Theanine Accumulation in Camellia sinensis (L.) Kuntze. Molecules. 2021; 26(11):3290. https://doi.org/10.3390/molecules26113290
Chicago/Turabian StyleLi, Wei, Fen Xiang, Yi Su, Zhoufei Luo, Weigui Luo, Lingyun Zhou, Hongyan Liu, and Langtao Xiao. 2021. "Gibberellin Increases the Bud Yield and Theanine Accumulation in Camellia sinensis (L.) Kuntze" Molecules 26, no. 11: 3290. https://doi.org/10.3390/molecules26113290
APA StyleLi, W., Xiang, F., Su, Y., Luo, Z., Luo, W., Zhou, L., Liu, H., & Xiao, L. (2021). Gibberellin Increases the Bud Yield and Theanine Accumulation in Camellia sinensis (L.) Kuntze. Molecules, 26(11), 3290. https://doi.org/10.3390/molecules26113290