
Lupeol Accumulation Correlates with Auxin in the Epidermis of Castor

 ,
,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
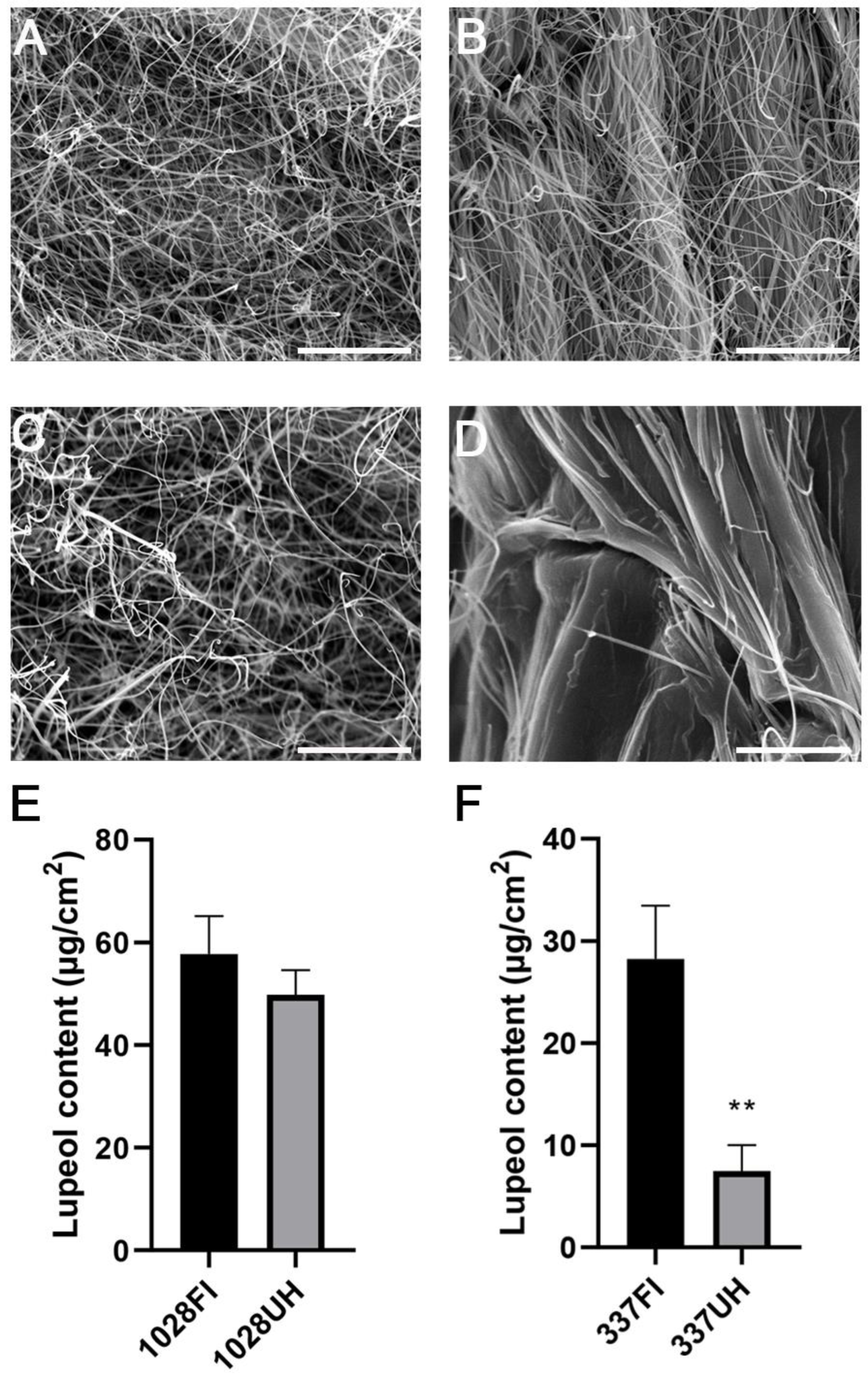
2.1. Castor Accession Line 337 Has Less Lupeol Content on Its Hypocotyl Compared to the Stem
2.2. RNA-Sequencing, Transcript Assembly, and Annotation
2.3. Comparative Transcriptome Enrichment Analysis between Upper Hypocotyl and First Internode of the Stem of Accession Line 337
2.4. Genes Closely Related with Lupeol Biosynthesis in Castor Epidermis
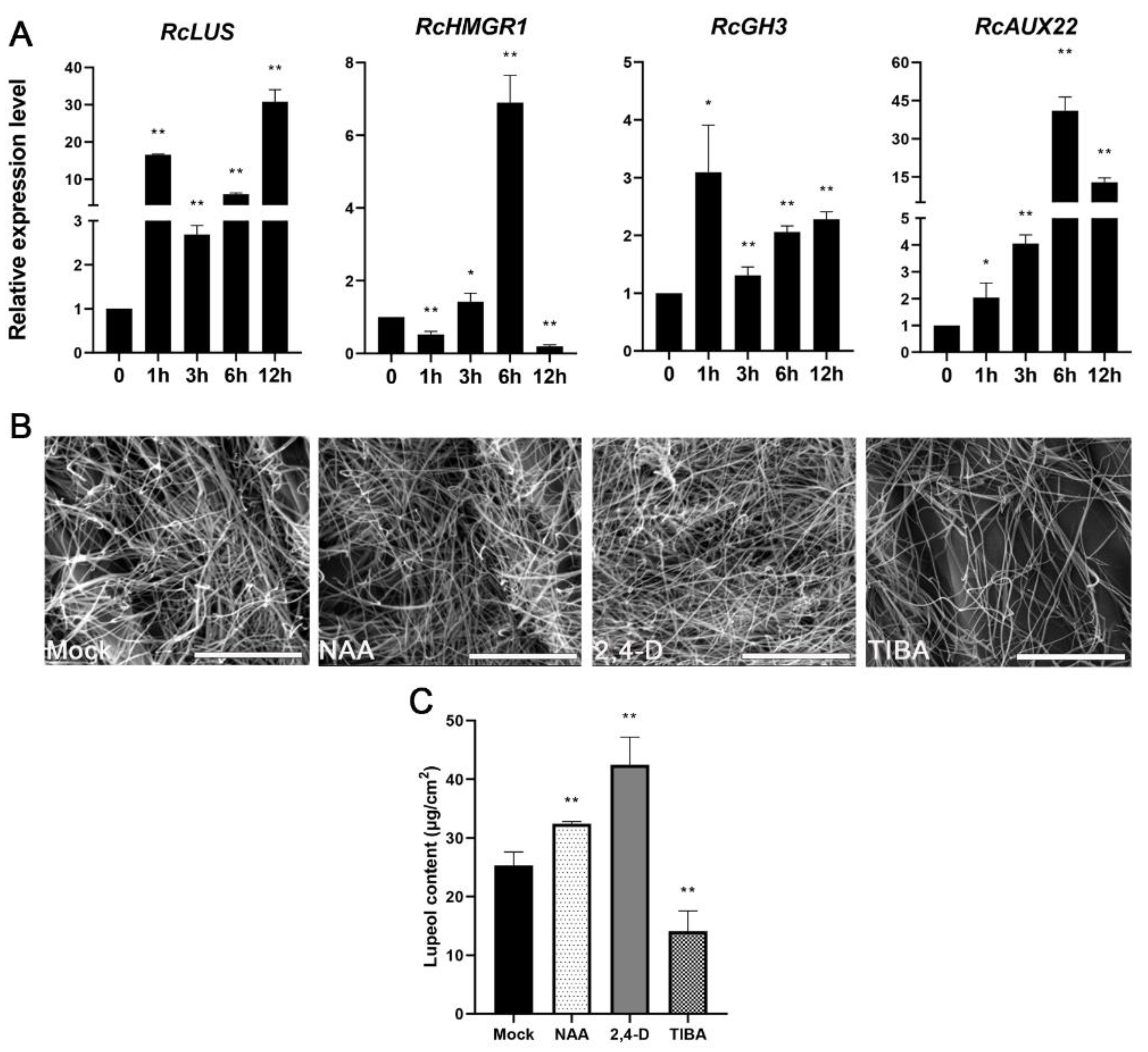
2.5. Auxin Signaling or Early Response Pathway Is Highly Enriched in the Upregulated DEGs
2.6. Auxin Treatment Enhanced Lupeol Accumulation in the Epidermis of Castor Leaves
3. Materials and Methods
3.1. Plant Materials
3.2. Cuticular Wax Analysis
3.3. Scanning Electron Microscopy (SEM)
3.4. RNA Extraction, cDNA Library Preparation, and Transcriptome Sequencing
3.5. Transcriptome Assembly, Functional Annotation, and DEG Analysis
3.6. Quantitative Reverse Transcription-PCR (qRT-PCR)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saleem, M. Lupeol, a novel anti-inflammatory and anti-cancer dietary triterpene. Cancer Lett. 2009, 285, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.M.; Xu, H.G.; Wang, L.; Li, Y.J.; Sun, P.H.; Wu, X.M.; Wang, G.J.; Chen, W.M.; Ye, W.C. Betulinic acid and its derivatives as potential antitumor agents. Med. Res. Rev. 2015, 35, 1127–1155. [Google Scholar] [CrossRef]
- Liu, K.; Zhang, X.; Xie, L.; Deng, M.; Chen, H.; Song, J.; Long, J.; Li, X.; Luo, J. Lupeol and its derivatives as anticancer and anti-inflammatory agents: Molecular mechanisms and therapeutic efficacy. Pharmacol. Res. 2021, 164, 105373. [Google Scholar] [CrossRef] [PubMed]
- Thimmappa, R.; Geisler, K.; Louveau, T.; O’Maille, P.; Osbourn, A. Triterpene biosynthesis in plants. Annu. Rev. Plant. Biol. 2014, 65, 225–257. [Google Scholar] [CrossRef]
- Vranova, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant. Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef]
- Liu, X.; Li, R.; Lu, W.; Zhou, Z.; Jiang, X.; Zhao, H.; Yang, B.; Lü, S. Transcriptome analysis identifies key genes involved in the regulation of epidermal lupeol biosynthesis in Ricinus communis. Ind. Crop. Prod. 2021, 160, 113100. [Google Scholar] [CrossRef]
- Guhling, O.; Hobl, B.; Yeats, T.; Jetter, R. Cloning and characterization of a lupeol synthase involved in the synthesis of epicuticular wax crystals on stem and hypocotyl surfaces of Ricinus communis. Arch. Biochem. Biophys. 2006, 448, 60–72. [Google Scholar] [CrossRef]
- Fan, W.; Lu, J.; Pan, C.; Tan, M.; Lin, Q.; Liu, W.; Li, D.; Wang, L.; Hu, L.; Wang, L.; et al. Sequencing of Chinese castor lines reveals genetic signatures of selection and yield-associated loci. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Chan, A.P.; Crabtree, J.; Zhao, Q.; Lorenzi, H.; Orvis, J.; Puiu, D.; Melake-Berhan, A.; Jones, K.M.; Redman, J.; Chen, G.; et al. Draft genome sequence of the oilseed species Ricinus communis. Nat. Biotechnol. 2010, 28, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Edqvist, J.; Blomqvist, K.; Nieuwland, J.; Salminen, T.A. Plant lipid transfer proteins: Are we finally closing in on the roles of these enigmatic proteins? J. Lipid Res. 2018, 59, 1374–1382. [Google Scholar] [CrossRef]
- Lee, E.J.; Kim, K.Y.; Zhang, J.; Yamaoka, Y.; Gao, P.; Kim, H.; Hwang, J.U.; Suh, M.C.; Kang, B.; Lee, Y. Arabidopsis seedling establishment under waterlogging requires ABCG5-mediated formation of a dense cuticle layer. New Phytol. 2021, 229, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Buda, G.J.; Barnes, W.J.; Fich, E.A.; Park, S.; Yeats, T.H.; Zhao, L.; Domozych, D.S.; Rose, J.K. An ATP binding cassette transporter is required for cuticular wax deposition and desiccation tolerance in the moss Physcomitrella patens. Plant. Cell 2013, 25, 4000–4013. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Molina, I.; Ranathunge, K.; Castillo, I.Q.; Rothstein, S.J.; Reed, J.W. ABCG transporters are required for suberin and pollen wall extracellular barriers in Arabidopsis. Plant. Cell 2014, 26, 3569–3588. [Google Scholar] [CrossRef] [PubMed]
- Salminen, T.A.; Eklund, D.M.; Joly, V.; Blomqvist, K.; Matton, D.P.; Edqvist, J. Deciphering the Evolution and Development of the Cuticle by Studying Lipid Transfer Proteins in Mosses and Liverworts. Plants 2018, 7, 6. [Google Scholar] [CrossRef]
- Hagen, G.; Guilfoyle, T. Auxin-responsive gene expression: Genes, promoters and regulatory factors. Plant Mol. Biol. 2002, 49, 373–385. [Google Scholar] [CrossRef]
- Woodward, A.W.; Bartel, B. A receptor for auxin. Plant. Cell 2005, 17, 2425–2429. [Google Scholar] [CrossRef]
- Staswick, P.E.; Serban, B.; Rowe, M.; Tiryaki, I.; Maldonado, M.T.; Maldonado, M.C.; Suza, W. Characterization of an Arabidopsis enzyme family that conjugates amino acids to indole-3-acetic acid. Plant. Cell 2005, 17, 616–627. [Google Scholar] [CrossRef]
- Stortenbeker, N.; Bemer, M. The SAUR gene family: The plant’s toolbox for adaptation of growth and development. J. Exp. Bot. 2019, 70, 17–27. [Google Scholar] [CrossRef]
- Chae, K.; Isaacs, C.G.; Reeves, P.H.; Maloney, G.S.; Muday, G.K.; Nagpal, P.; Reed, J.W. Arabidopsis SMALL AUXIN UP RNA63 promotes hypocotyl and stamen filament elongation. Plant. J. 2012, 71, 684–697. [Google Scholar] [CrossRef]
- Roig-Villanova, I.; Bou-Torrent, J.; Galstyan, A.; Carretero-Paulet, L.; Portolés, S.; Rodríguez-Concepción, M.; Martínez-García, J.F. Interaction of shade avoidance and auxin responses: A role for two novel atypical bHLH proteins. EMBO J. 2007, 26, 4756–4767. [Google Scholar] [CrossRef]
- Sauerwein, M.; Yamazaki, T.; Shimomura, K. Hernandulcin in hairy root cultures of Lippia dulcis. Plant. Cell Rep. 1991, 9, 579–581. [Google Scholar] [CrossRef]
- Shimomura, M.S.K.I.K. Indole alkaloids in hairy roots of Amsonia elliptica. Phytochemistry 1991, 30, 1153–1155. [Google Scholar]
- Washida, D.; Shimomura, K.; Takido, M.; Kitanaka, S. Auxins affected ginsenoside production and growth of hairy roots in Panax hybrid. Biol. Pharm. Bull. 2004, 27, 657–660. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.H.; Adhikari, P.B.; Park, S.B.; Han, J.Y.; Choi, Y.E. Production of the dammarene sapogenin (protopanaxadiol) in transgenic tobacco plants and cultured cells by heterologous expression of PgDDS and CYP716A47. Plant. Cell Rep. 2015, 34, 1551–1560. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Cassol, D.; Cruz, F.P.; Espindola, K.; Mangeon, A.; Müller, C.; Ehlers Loureiro, M.; Corrêa, R.L.; Sachetto-Martins, G. Identification of reference genes for quantitative RT-PCR analysis of microRNAs and mRNAs in castor bean (Ricinus communis L.) under drought stress. Plant. Physiol. Biochem. 2016, 106, 101–107. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene-ID | log2FC * | p-Value | Gene Description |
|---|---|---|---|
| LOC8280179 | 2.297 | 0.0002 | auxin-responsive protein IAA7 |
| LOC8271341 | 1.957 | 0.0005 | auxin-induced protein AUX22 |
| LOC8285710 | 2.956 | 0.0002 | indole-3-acetic acid-amido synthetase GH3.17 |
| LOC107262116 | 3.171 | 0.0001 | auxin-induced protein 15A-like |
| LOC107260799 | 4.896 | 0.0001 | auxin-induced protein 15A-like |
| LOC107262113 | 6.189 | 0.0002 | auxin-induced protein 15A-like |
| LOC107262111 | 3.754 | 0.0002 | auxin-induced protein 15A-like |
| LOC107262117 | 5.028 | 0.0008 | auxin-induced protein 15A-like |
| LOC107261577 | 4.484 | 0.0001 | auxin-responsive protein SAUR21-like |
| LOC107261578 | 6.831 | 0.0000 | auxin-responsive protein SAUR21-like |
| LOC107261581 | 7.976 | 0.0000 | auxin-responsive protein SAUR21-like |
| LOC107261586 | 8.247 | 0.0000 | auxin-responsive protein SAUR21-like |
| LOC107261587 | 7.936 | 0.0000 | auxin-responsive protein SAUR21-like |
| LOC8283211 | 3.697 | 0.0003 | auxin-responsive protein SAUR62-like |
| LOC8283214 | 7.806 | 0.0000 | auxin-responsive protein SAUR63-like |
| LOC8283212 | 6.678 | 0.0000 | auxin-responsive protein SAUR63-like |
| LOC8266878 | 8.459 | 0.0000 | auxin-responsive protein SAUR68-like |
| LOC8266881 | 8.433 | 0.0000 | auxin-responsive protein SAUR68-like |
| LOC8266894 | 8.010 | 0.0000 | auxin-responsive protein SAUR68-like |
| LOC8266888 | 2.562 | 0.0003 | auxin-responsive protein SAUR68-like |
| LOC8266882 | 6.531 | 0.0000 | auxin-responsive protein SAUR68-like |
| LOC8266895 | 7.277 | 0.0000 | auxin-responsive protein SAUR68-like |
| LOC8266875 | 6.060 | 0.0000 | auxin-responsive protein SAUR68-like |
| LOC8266880 | 4.946 | 0.0004 | auxin-responsive protein SAUR68-like |
| LOC8266887 | 6.710 | 0.0000 | auxin-responsive protein SAUR68-like |
| LOC8283213 | 5.393 | 0.0003 | auxin-responsive protein SAUR68-like |
| LOC8266876 | 6.118 | 0.0003 | auxin-responsive protein SAUR68-like |
| LOC8266884 | 5.706 | 0.0008 | auxin-responsive protein SAUR68-like |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Pan, C.; Lu, J.; Zaman, W.; Zhao, H.; Zhang, J.; Lü, S. Lupeol Accumulation Correlates with Auxin in the Epidermis of Castor. Molecules 2021, 26, 2978. https://doi.org/10.3390/molecules26102978
Li D, Pan C, Lu J, Zaman W, Zhao H, Zhang J, Lü S. Lupeol Accumulation Correlates with Auxin in the Epidermis of Castor. Molecules. 2021; 26(10):2978. https://doi.org/10.3390/molecules26102978
Chicago/Turabian StyleLi, Donghai, Cheng Pan, Jianjun Lu, Wajid Zaman, Huayan Zhao, Jixing Zhang, and Shiyou Lü. 2021. "Lupeol Accumulation Correlates with Auxin in the Epidermis of Castor" Molecules 26, no. 10: 2978. https://doi.org/10.3390/molecules26102978
APA StyleLi, D., Pan, C., Lu, J., Zaman, W., Zhao, H., Zhang, J., & Lü, S. (2021). Lupeol Accumulation Correlates with Auxin in the Epidermis of Castor. Molecules, 26(10), 2978. https://doi.org/10.3390/molecules26102978