Composition and Biosynthesis of Scent Compounds from Sterile Flowers of an Ornamental Plant Clematis florida cv. ‘Kaiser’


 , and
, and
Abstract
1. Introduction
2. Results
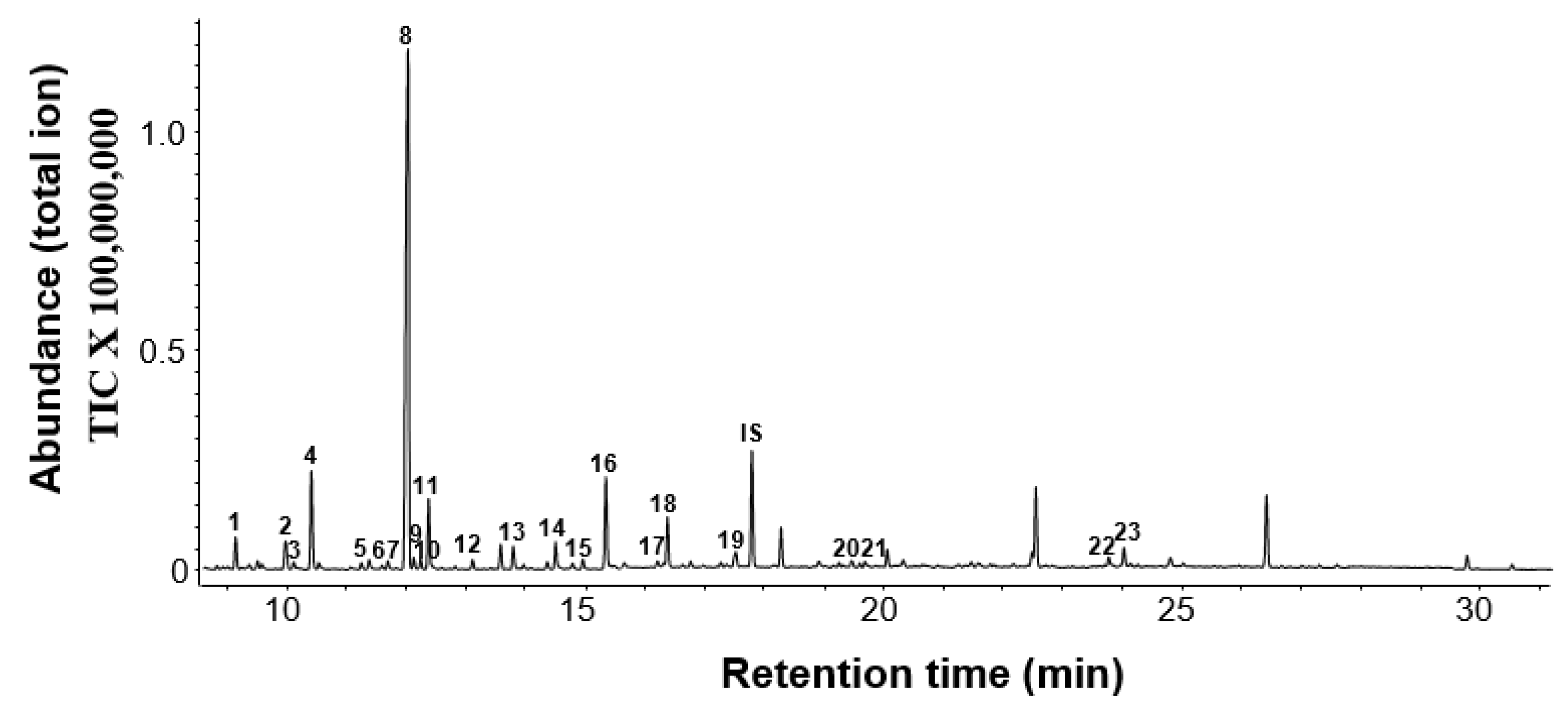
2.1. Chemical Composition of Floral Volatiles of ‘Kaiser’
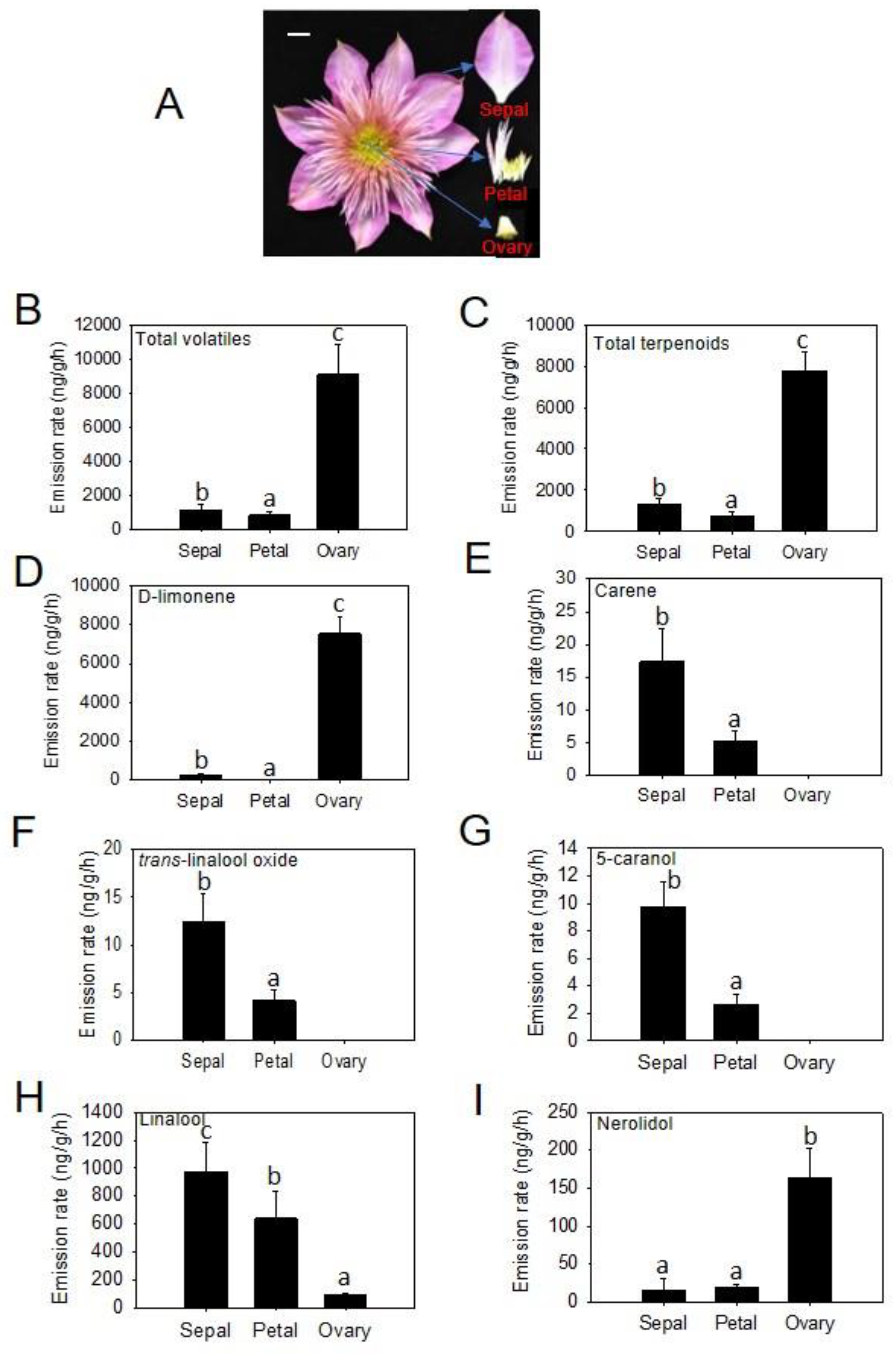
2.2. Spatial Specificity of Volatile Emission in Different Floral Organs from ‘Kaiser’
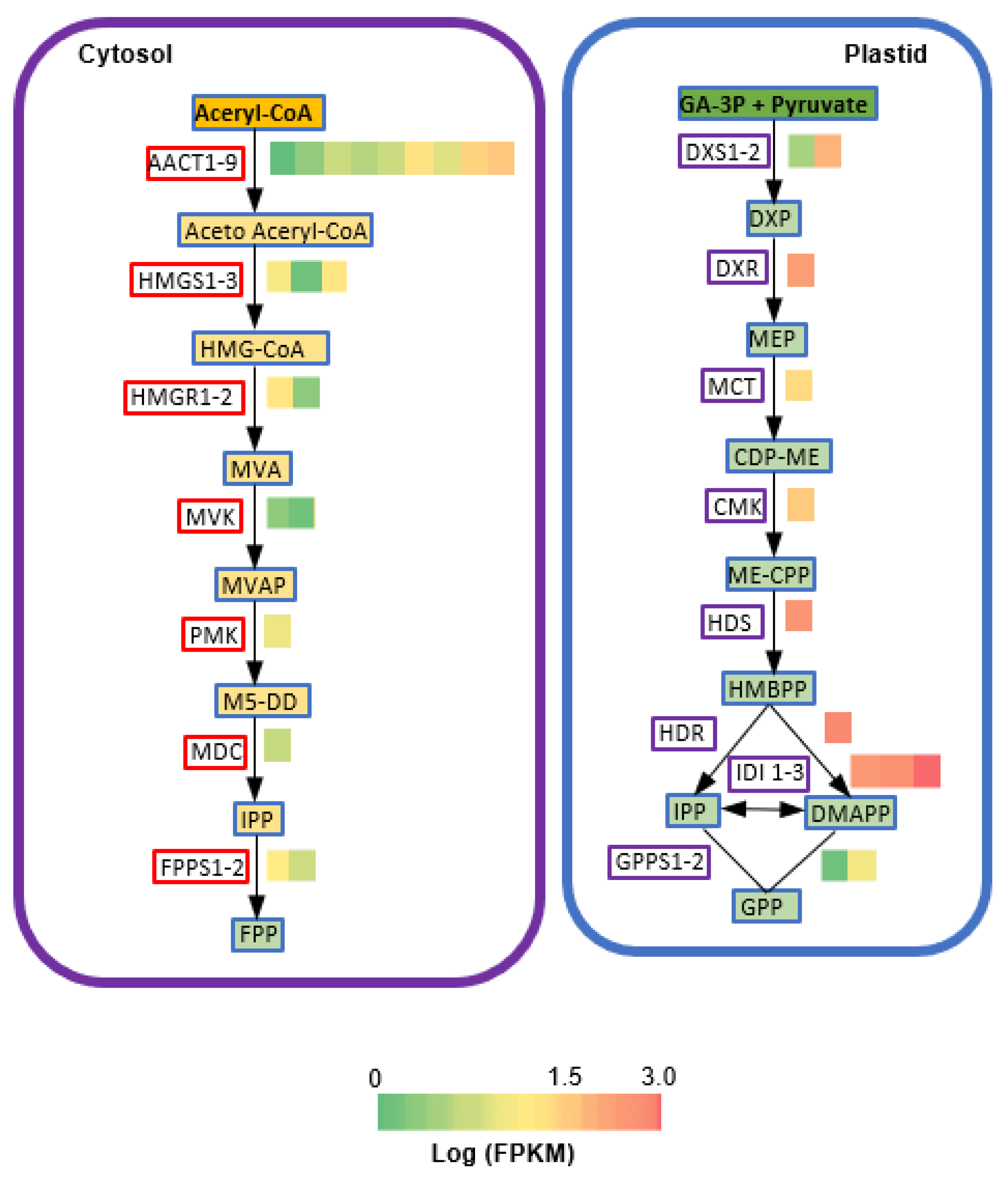
2.3. Transcriptome Sequencing and Annotation of Assembled Unigenes
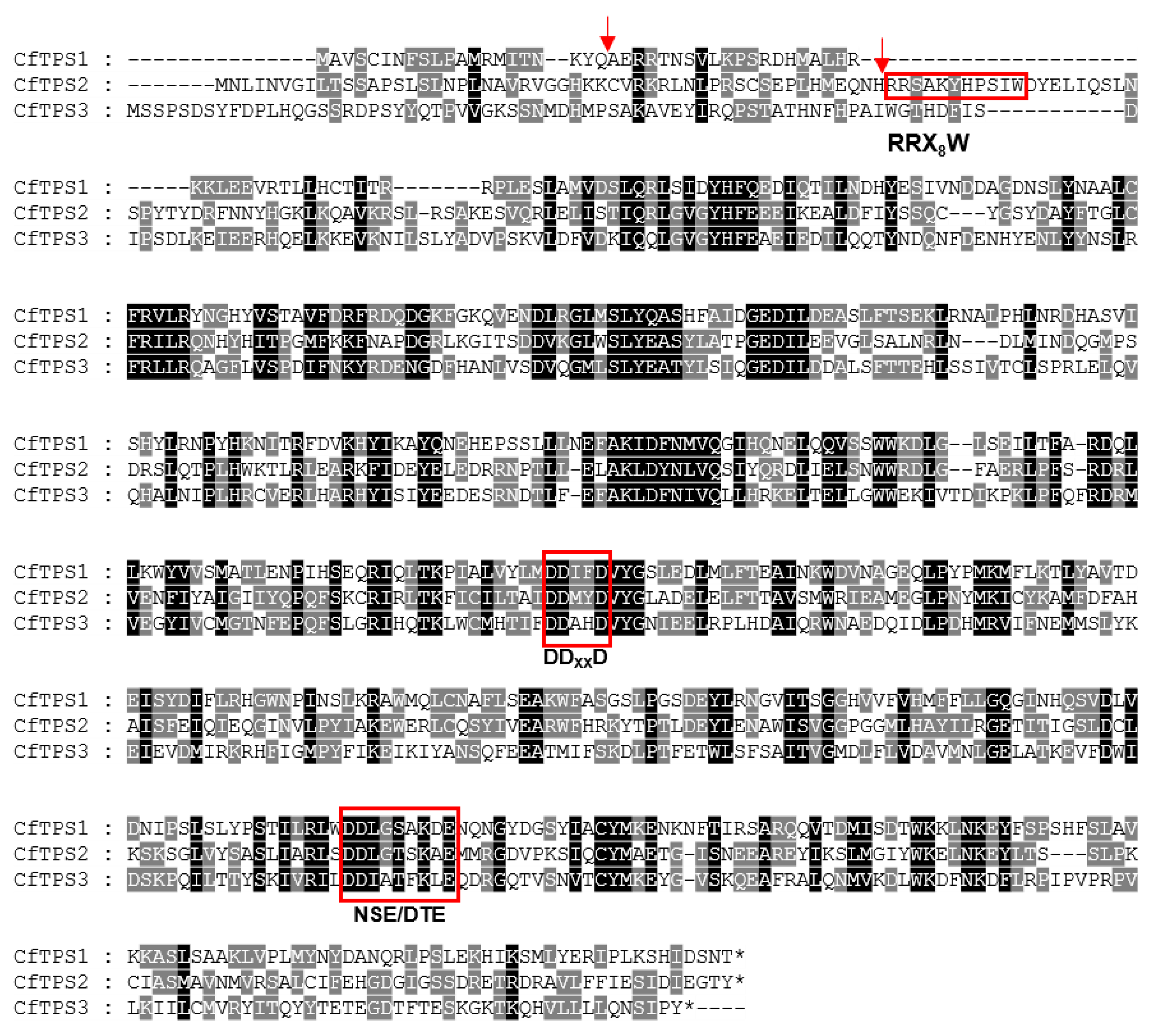
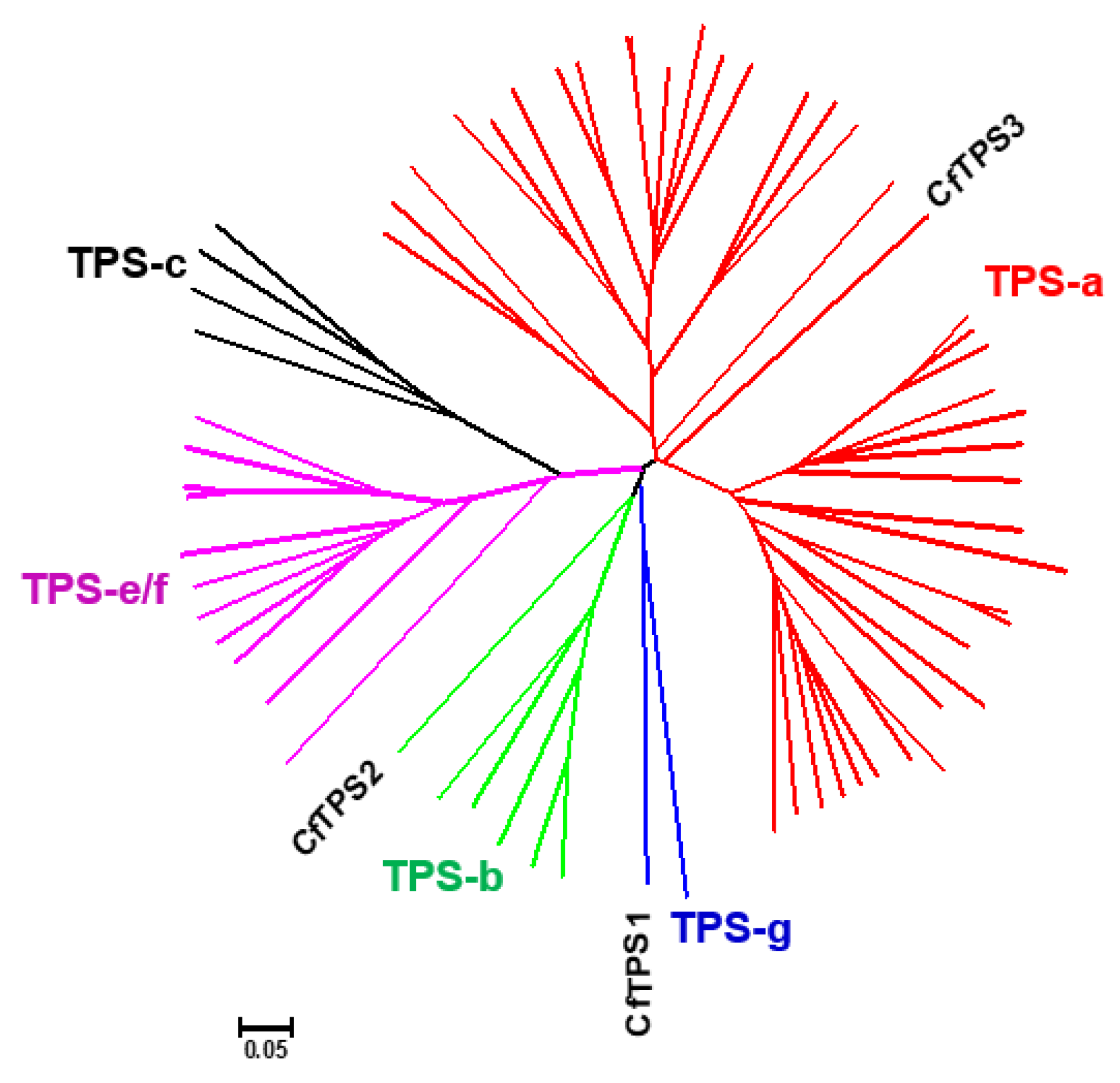
2.4. Identification of CfTPS Genes
2.5. Catalytic Activities of CfTPSs
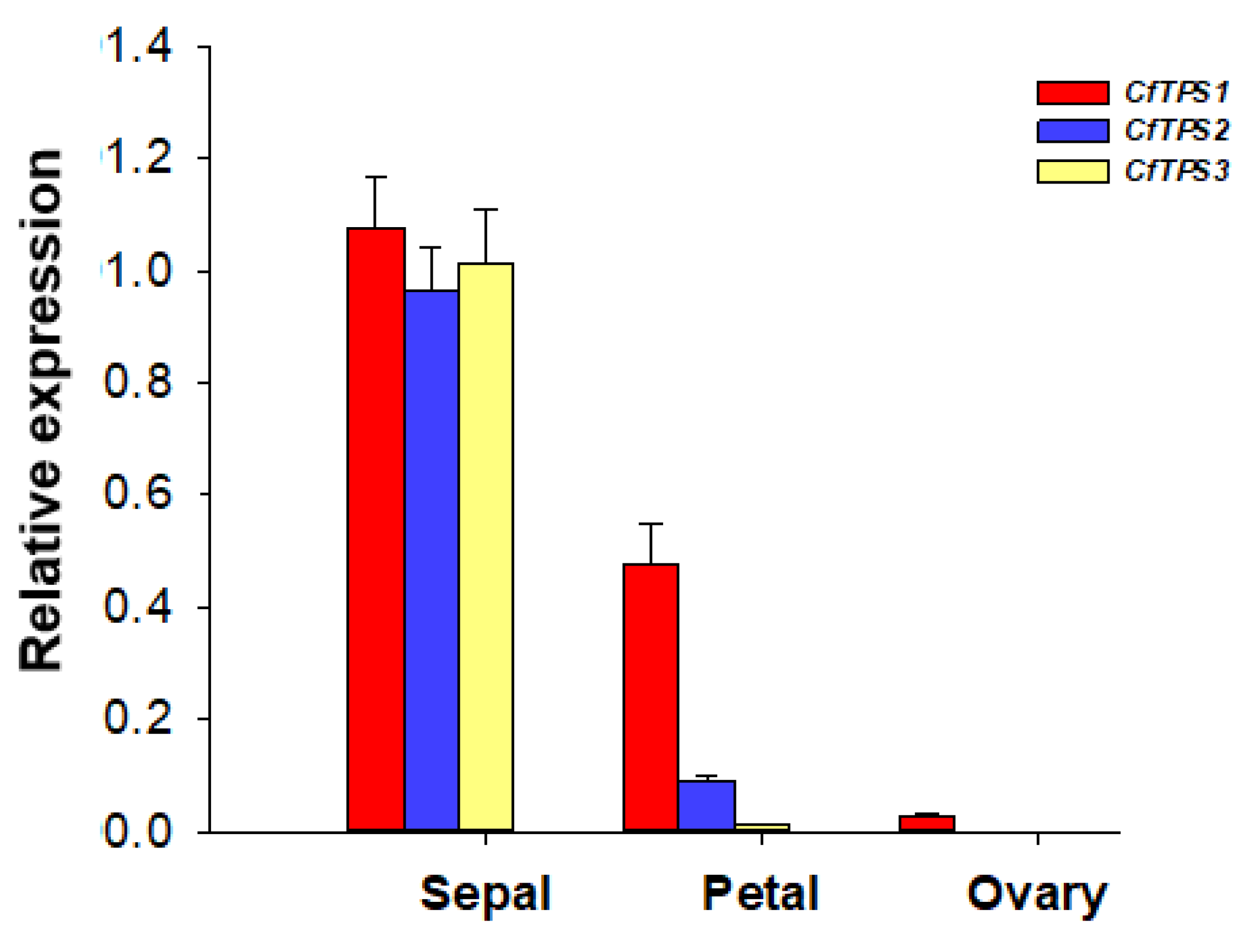
2.6. Expression of CfTPS Genes in Different Floral Organs
3. Discussion
4. Materials and Methods
4.1. Plants and Growth
4.2. Floral Volatile Collection and Identification
4.3. RNA Preparation, Transcriptome Sequencing, and Analysis
4.4. Identification of CfTPS and Sequence Analysis
4.5. Protein Expression and Terpene Synthase Enzyme Assays
4.6. Quantitative Real-time PCR of CfTPS Genes in Different Floral Organs
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wang, W.C.; Bruce, B. Clematis Linnaeus. Flora China 2001, 6, 333–386. [Google Scholar]
- Huang, Z.H.; Zhang, Y.L.; Shen, X.J. Pharmacognostic Identification of Clematis florida var. plena. Subtrop Plant Sci. 2013, 42, 104–108. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Zhao, C.X.; Liang, Y.Z.; Yang, H.; Fang, H.Z.; Yi, L.Z.; Zhong, D.Z. Comparative analysis of volatile components from Clematis species growing in china. Anal. Chim. Acta 2007, 595, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Rakia, S.; Ferdaws, G.; Rim, K.; Abdelfattah, E.F.; Tahya, B.; Christophe, C.; Emna, A.; Raoudha, M.J. Tunisian Clematis flammula essential oil enhances wound healing: GC-MS analysis, biochemical and histological assessment. J. Oleo Sci. 2018, 67, 1483–1499. [Google Scholar] [CrossRef]
- Chawla, R.; Kumar, S.; Sharma, A. The genus Clematis (Ranunculaceae): Chemical and pharmacological perspectives. J Ethnopharmacol. 2012, 143, 116–150. [Google Scholar] [CrossRef]
- Zhao, M.; Da-Wa, Z.M.; Guo, D.L.; Fang, D.M.; Chen, X.Z.; Xu, H.X.; Gu, Y.C.; Xia, B.; Chen, L.; Ding, L.S.; et al. Cytotoxic triterpenoid saponins from Clematis tangutica. Phytochemistry 2016, 130, 228–237. [Google Scholar] [CrossRef]
- Xiong, J.; Bui, V.B.; Liu, X.H.; Hong, Z.L.; Yang, G.X.; Hu, J.F. Lignans from the stems of Clematis armandii (“Chuan-Mu-Tong”) and their anti-neuroinflammatory activities. J. Ethnopharmacol. 2014, 153, 737–743. [Google Scholar] [CrossRef]
- Qiu, L.; Yuan, H.M.; Liang, J.M.; Cheng, X.L.; Wang, P.; Du, Y.F.; Qiang, F. Clemochinenosides C and D, two new macrocyclic glucosides from Clematis chinensis. J. Asian Nat. Prod. Res. 2017, 23, 1–7. [Google Scholar] [CrossRef]
- Li, W.T.; Yang, B.Y.; Zhu, W.; Gong, M.H.; Xu, X.D.; Lu, X.H.; Sun, L.L.; Tian, J.K.; Zhang, L. A new indole alkaloidal glucoside from the aerial parts of Clematis terniflora DC. Nat. Prod. Res. 2013, 27, 2333–2337. [Google Scholar] [CrossRef]
- Sun, K.H.; Ma, X.H.; Zeng, X.M.; Lin, Z.Y.; Cai, Y.M.; Zhang, H.T.; Lin, X.Y.; Feng, S.B.; Zhong, T.H.; Zhang, Y.H. A new indole-type alkaloid from the roots of Clematis florida var. plena. Nat. Prod. Res. 2018, 33, 2925–2931. [Google Scholar] [CrossRef]
- Caruso, C.M.; Parachnowitsch, A.L. Do Plants Eavesdrop on Floral Scent Signals? Trends Plant Sci. 2016, 21, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function. Plant Cell Environ. 2014, 37, 1936–1949. [Google Scholar] [CrossRef] [PubMed]
- Dexter, R.; Qualley, A.; Kish, C.M.; Ma, C.J.; Koeduka, T.; Nagegowda, D.A.; Dudareva, N.; Pichersky, E.; Clark, D. Characterization of a petunia acetyltransferase involved in the biosynthesis of the floral volatile isoeugenol. Plant J. 2007, 49, 265–275. [Google Scholar] [CrossRef] [PubMed]
- D’AuriaJ, C. Acyltransferases in plants: A good time to be BAHD. Curr. Opin. Plant Biol. 2006, 9, 331–340. [Google Scholar] [CrossRef]
- Pott, M.B.; Hippauf, F.; Saschenbrecker, S.; Chen, F.; Ross, J.; Kiefer, I.; Slusarenko, A.; Noel, J.P.; Pichersky, E.; Effmert, U.; et al. Biochemical and structural characterization of benzenoid carboxyl methyltransferases involved in floral scent production in Stephanotis floribunda and Nicotiana suaveolens. Plant Physiol. 2004, 135, 1946–1955. [Google Scholar] [CrossRef][Green Version]
- Koeduka, T.; Kajiyama, M.; Suzuki, H.; Furuta, T.; Tsuge, T.; Matsui, K. Benzenoid biosynthesis in the flowers Oferiobotrya japonica: Molecular cloning and functional characterization of p-methoxybenzoic acid carboxyl methyltransferase. Planta 2016, 244, 725–736. [Google Scholar] [CrossRef]
- Gao, F.Z.; Liu, B.F.; Li, M.; Gao, X.Y.; Fang, Q.; Liu, C.; Ding, H.; Wang, L.; Gao, X. Identification and characterization of terpene synthase genes accounting for volatile terpene emissions in flowers of Freesia x hybrida. J. Exp. Bot. 2018, 69, 4249–4265. [Google Scholar] [CrossRef]
- Dhandapani, S.; Jin, J.; Sridhar, V.; Rajani, S.; Chua, N.H.; Jang, I.C. Integrated metabolome and transcriptome analysis of Magnolia champaca identifies biosynthetic pathways for floral volatile organic compounds. BMC Genomics 2017. [CrossRef]
- Jin, J.J.; Kim, M.J.; Dhandapani, S.; Tjhang, J.G.; Yin, J.L.; Wong, L.; Sarojam, R.; Chua, N.H.; Jang, I.C. The floral transcriptome of ylang ylang (Cananga odorata var. fruticosa) uncovers biosynthetic pathways for volatile organic compounds and a multifunctional and novel sesquiterpene synthase. J. Exp. Bot. 2015, 66, 3959–3975. [Google Scholar] [CrossRef]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; D’Auria, J.C.; Farooq, A.; Pichersky, E.; Gershenzon, J. Biosynthesis and emission of terpenoid volatiles from arabidopsis flowers. Plant Cell. 2003, 15, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.L.; Liu, C.; Zheng, R.R.; Cai, X.; Luo, J.; Zou, J.J.; Wang, C.Y. Emission and accumulation of monoterpene and the key terpene synthase (TPS) associated with monoterpene biosynthesis in Osmanthus fragrans Lour. Front. Plant Sci. 2016, 6, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pazouki, L.; Memari, H.R.; Kannaste, A.; Bichele, R.; Niinemets, U. Germacrene A synthase in yarrow (Achillea millefolium) is an enzyme with mixed substrate specificity: Gene cloning, functional characterization and expression analysis. Front. Plant Sci. 2015, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Azuma, H.; Thien, L.B.; Kawano, S. Floral scents, leaf volatiles and thermogenic flowers in Magnoliaceae. Plant Spec. Biol. 1999, 14, 121–127. [Google Scholar] [CrossRef]
- Azuma, H.; Toyota, M.; Asakawa, Y.; Yamaoka, R.; Jose, G.G.F.; Dieringer, G. Chemical divergence in floral scents of magnolia and allied genera (magnoliaceae). Plant Spec. Biol. 2010, 12, 69–83. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, T.; Fan, Q.; Qi, X.; Zhang, F.; Fang, W.; Jiang, J.F.; Chen, F.D.; Chen, S.M. Identification of floral scent in chrysanthemum cultivars and wild relatives by gas chromatography-mass spectrometry. Molecules 2015, 20, 5346–5359. [Google Scholar] [CrossRef]
- Parachnowitsch, A.L.; Burdon, R.C.F.; Raguso, R.A.; Kessler, A. Natural selection on floral volatile production in Penstemon digitalis: Highlighting the role of linalool. Plant Signal. Behav. 2013. [Google Scholar] [CrossRef]
- Raguso, R.A.; Pichersky, E. New Perspectives in Pollination Biology: Floral Fragrances. A day in the life of a linalool molecule: Chemical communication in a plant-pollinator system. Part 1: Linalool biosynthesis in flowering plants. Plant Spec. Biol. 2010, 14, 95–120. [Google Scholar] [CrossRef]
- Raguso, R.A. More lessons from linalool: Insights gained from a ubiquitous floral volatile. Curr. Opin. Plant Biol. 2016, 32, 31–36. [Google Scholar] [CrossRef]
- Yang, T.; Stoopen, G.; Thoen, M.; Wiegers, G.; Jongsma, M.A. Chrysanthemum expressing a linalool synthase gene ‘smells good’, but ‘tastes bad’ to western flower thrips. Plant Biotechnol. J. 2013, 11, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.F.; Zhao, N.; Wang, F.; Chen, F. Emission and regulation of volatile chemicals from globe amaranth flowers. J. Amer. Soc. Hort. Sci. 2011, 136, 16–22. [Google Scholar] [CrossRef]
- Zhuang, X.F.; Klingeman, W.E.; Hu, J.; Chen, F. Emission of volatile chemicals from flowering dogwood (Cornus florida L.) flowers. J. Agric. Food Chem. 2008, 56, 9570–9574. [Google Scholar] [CrossRef]
- Huang, Y.; Liao, M.; Yang, Q.Q.; Xiao, J.J.; Hu, Z.Y.; Zhou, L.J.; Cao, H.Q. Transcriptome profiling reveals differential gene expression of detoxification enzymes in Sitophilus zeamais responding to terpinen-4-ol fumigation. Pestic Biochem. Phys. 2018, 149, 44–53. [Google Scholar] [CrossRef]
- Li, G.L.; Kollner, T.G.; Yin, Y.B.; Jiang, Y.F.; Chen, H.; Xu, Y.; Gershenzon, J.; Pichersky, E.; Chen, F. Nonseed plant Selaginella moellendorfii has both seed plant and microbial types of terpene synthases. Proc. Natl. Acad. Sci. USA 2012, 109, 14711–14715. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compounds | Retention Index | Emission Rate (ng/g/h) c | |
|---|---|---|---|---|
| Calc a | Lit b | |||
| Terpenoids | ||||
| 1 | Limonene | 1033 | 1031 | 28.46 ± 12.62 |
| 2 | trans-Linalool oxide | 1053 | 1081 | 3.41 ± 0.64 |
| 3 | 3-Carene | 1079 | 1017 | 2.96 ± 0.30 |
| 4 | 5-Caranol | 1095 | 1125 | 4.12 ± 0.53 |
| 5 | Linalool | 1107 | 1101 | 423.22 ± 33.71 |
| 6 | 6-Ethenyldihydro-2,2,6-trimethyl-2H-Pyran-3(4H)-one | 1115 | 1108 | 4.87 ± 0.64 |
| 7 | Nerolidol | 1572 | 1564 | 10.11 ± 0.74 |
| Benzenoids | ||||
| 8 | Benzyl alcohol | 1038 | 1036 | 2.62 ± 0.22 |
| 9 | 2-Phenylacetaldehyde | 1049 | 1047 | 52.43 ± 7.49 |
| 10 | Phenylethyl alcohol | 1119 | 1116 | 37.45 ± 5.49 |
| 11 | 1,4-Dimethoxybenzene | 1170 | 1158 | 12.73 ± 1.87 |
| 12 | 2-(2-Butoxyethoxy)ethanol | 1195 | 1192 | 15.36 ± 2.47 |
| 13 | 2-Phenoxyethanol | 1226 | 1225 | 48.69 ± 11.24 |
| 14 | 2-Phenylethylacetate | 1265 | 1258 | 26.97 ± 4.12 |
| 15 | 1,3,5-Trimethoxybenzene | 1417 | 1392 | 3.75 ± 0.37 |
| Fatty acid derivatives | ||||
| 16 | 2-Hydroxyethyl methacrylate | 1019 | 985 | 1.76 ± 0.26 |
| 17 | Nonanal | 1110 | 1104 | 2.70 ± 0.33 |
| 18 | Decanal | 1212 | 1206 | 5.24 ± 0.75 |
| 19 | Tetradecane | 1406 | 1400 | 2.39 ± 0.41 |
| 20 | 2,6,10,15-Tetramethylheptadecane | 1506 | 1660 | 4.87 ± 0.71 |
| Nitrogen-containing compounds | ||||
| 21 | Benzyl cyanide | 1146 | 1144 | 7.49 ± 1.12 |
| 22 | Phenylacetaldoxime | 1253 | 1402 | 1.32 ± 0.25 |
| 23 | 2-Phenylnitroethane | 1311 | 1283 | 10.32 ± 1.86 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Qian, R.; Zhang, W.; Wei, G.; Ma, X.; Zheng, J.; Köllner, T.G.; Chen, F. Composition and Biosynthesis of Scent Compounds from Sterile Flowers of an Ornamental Plant Clematis florida cv. ‘Kaiser’. Molecules 2020, 25, 1711. https://doi.org/10.3390/molecules25071711
Jiang Y, Qian R, Zhang W, Wei G, Ma X, Zheng J, Köllner TG, Chen F. Composition and Biosynthesis of Scent Compounds from Sterile Flowers of an Ornamental Plant Clematis florida cv. ‘Kaiser’. Molecules. 2020; 25(7):1711. https://doi.org/10.3390/molecules25071711
Chicago/Turabian StyleJiang, Yifan, Renjuan Qian, Wanbo Zhang, Guo Wei, Xiaohua Ma, Jian Zheng, Tobias G. Köllner, and Feng Chen. 2020. "Composition and Biosynthesis of Scent Compounds from Sterile Flowers of an Ornamental Plant Clematis florida cv. ‘Kaiser’" Molecules 25, no. 7: 1711. https://doi.org/10.3390/molecules25071711
APA StyleJiang, Y., Qian, R., Zhang, W., Wei, G., Ma, X., Zheng, J., Köllner, T. G., & Chen, F. (2020). Composition and Biosynthesis of Scent Compounds from Sterile Flowers of an Ornamental Plant Clematis florida cv. ‘Kaiser’. Molecules, 25(7), 1711. https://doi.org/10.3390/molecules25071711