Chitosan-Based Agronanochemicals as a Sustainable Alternative in Crop Protection


Abstract
1. Introduction
2. Chitosan-Based Agronanochemicals
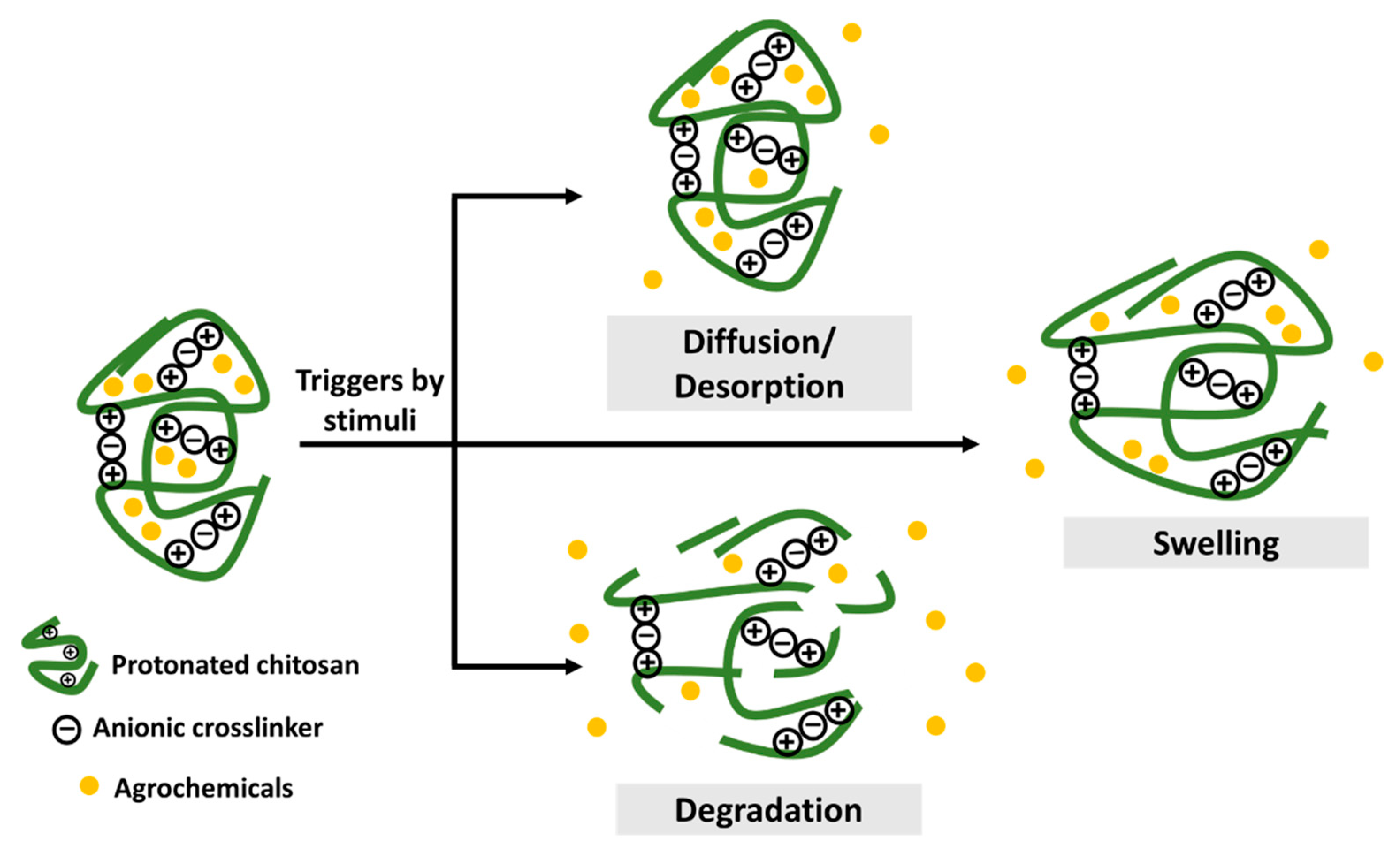
2.1. Controlled Release Formulations
2.2. Plant Growth Promoter
2.3. Biocides Against Plant Pathogens and Pests
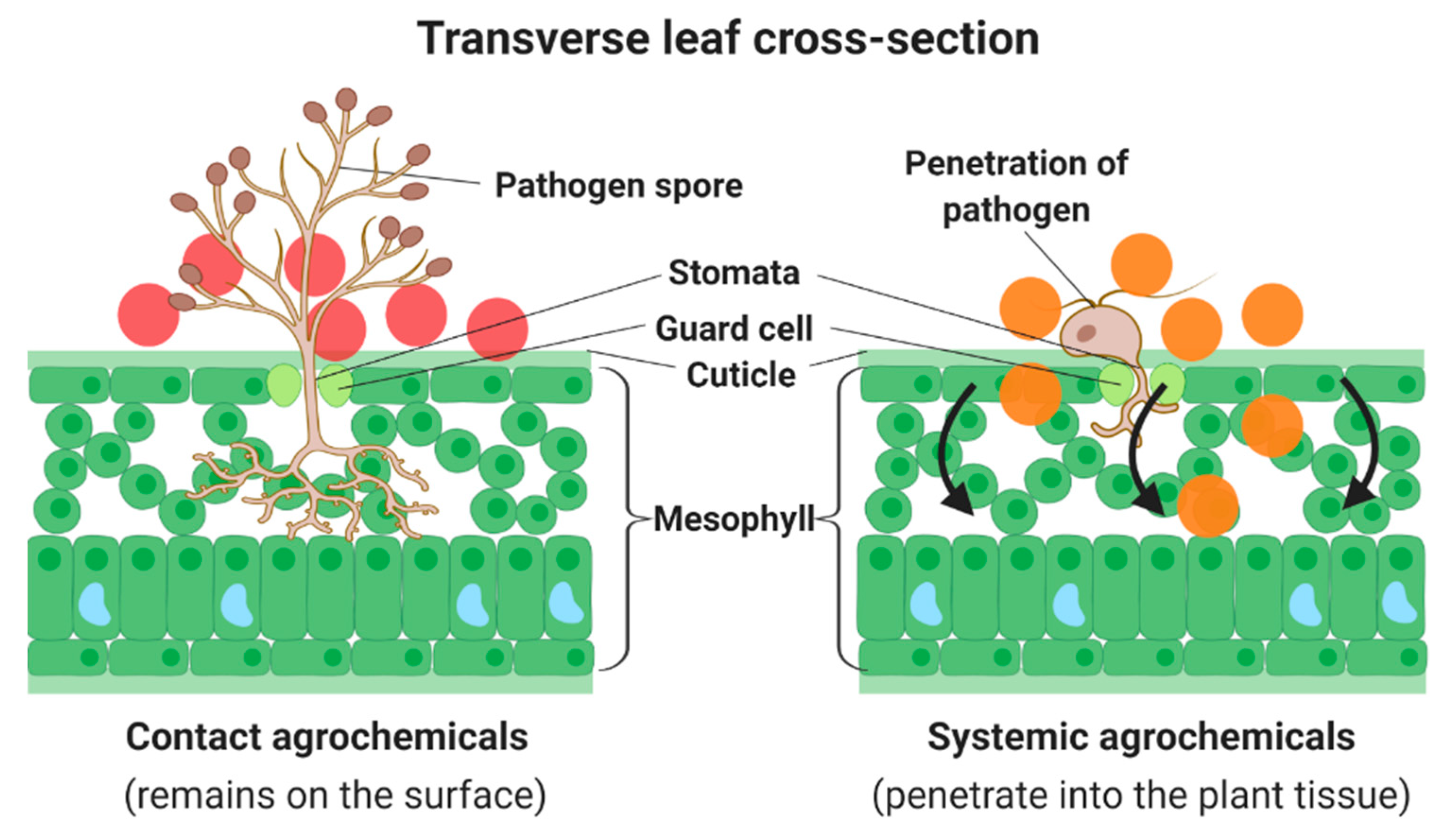
3. The Mechanism of Actions of Chitosan Against the Pathogens
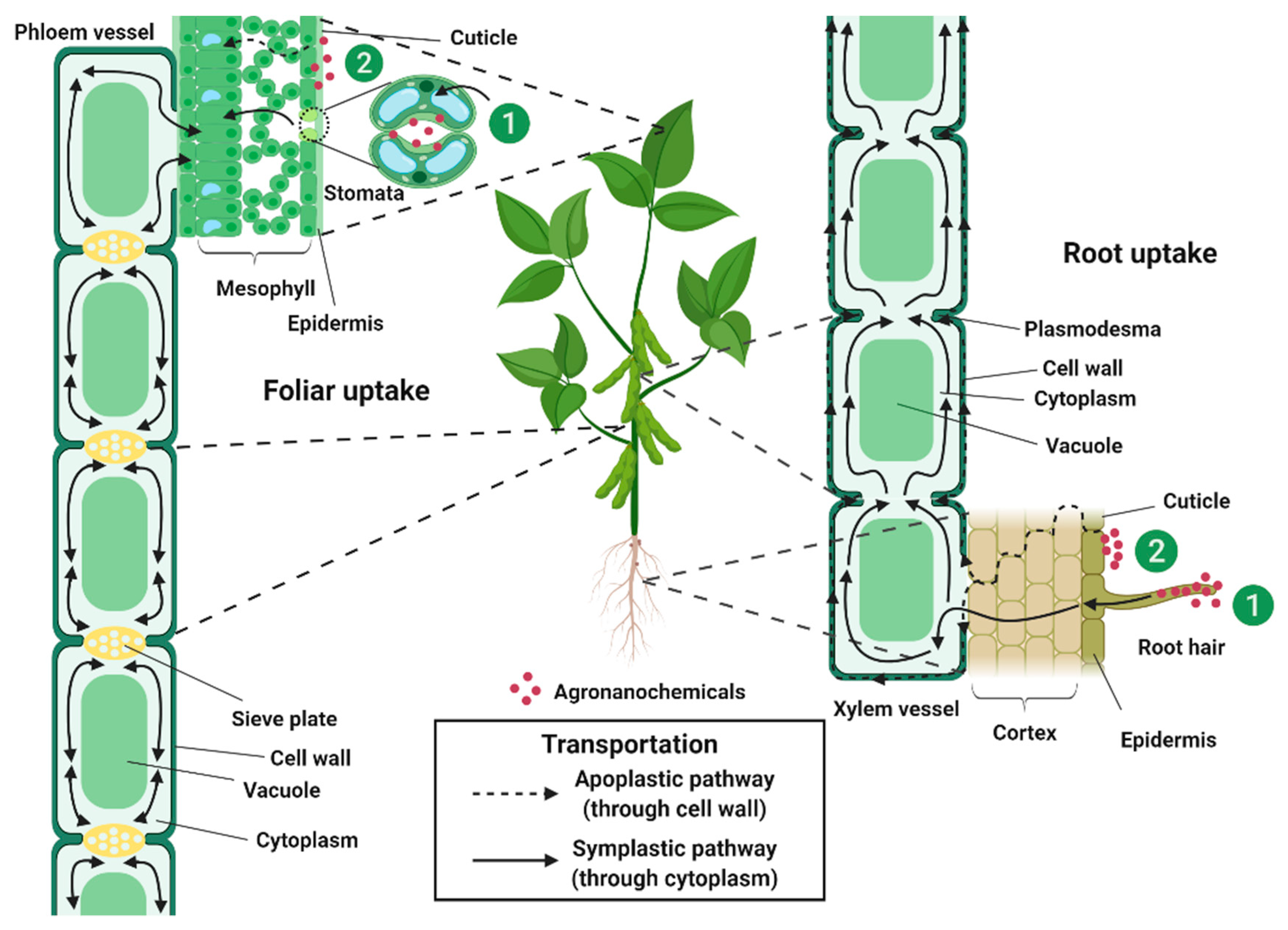
4. Uptake, Translocation and Transportation of Agronanochemicals in Plant
5. Phytoprotection, Cytoprotection and Genoprotection of Chitosan
6. Agronanochemicals Exert Negative Impacts on Human Health and Environment Wellbeing
7. Future Perspectives
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oerke, E.-C.; Dehne, H.-W. Safeguarding production—losses in major crops and the role of crop protection. Crop Prot. 2004, 23, 275–285. [Google Scholar] [CrossRef]
- Roy, R.N.; Finck, A.; Blair, G.; Tandon, H. Plant nutrition for food security. A guide for integrated nutrient management. FAO Fertil. Plant Nutr. Bull. 2006, 16, 368. [Google Scholar]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D. Worldwide pesticide usage and its impacts on ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef]
- Zhang, W. Global pesticide use: Profile, trend, cost/benefit and more. Proc. Int. Acad. Ecol. Environ. Sci. 2018, 8, 1. [Google Scholar]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Popp, J.; Pető, K.; Nagy, J. Pesticide productivity and food security. A review. Agron. Sustain. Dev. 2013, 33, 243–255. [Google Scholar] [CrossRef]
- Camara, M.C.; Campos, E.V.R.; Monteiro, R.A.; Santo Pereira, A.d.E.; de Freitas Proença, P.L.; Fraceto, L.F. Development of stimuli-responsive nano-based pesticides: Emerging opportunities for agriculture. J. Nanobiotechnol. 2019, 17, 100. [Google Scholar] [CrossRef]
- Ghormade, V.; Deshpande, M.V.; Paknikar, K.M. Perspectives for nano-biotechnology enabled protection and nutrition of plants. Biotechno. Adv. 2011, 29, 792–803. [Google Scholar] [CrossRef]
- De, A.; Bose, R.; Kumar, A.; Mozumdar, S. Targeted Delivery of Pesticides Using Biodegradable Polymeric Nanoparticles; Springer: New Delhi, India, 2014. [Google Scholar]
- Rodrigues, S.M.; Demokritou, P.; Dokoozlian, N.; Hendren, C.O.; Karn, B.; Mauter, M.S.; Sadik, O.A.; Safarpour, M.; Unrine, J.M.; Viers, J. Nanotechnology for sustainable food production: Promising opportunities and scientific challenges. Environ. Sci-Nano 2017, 4, 767–781. [Google Scholar] [CrossRef]
- Sangeetha, J.; Thangadurai, D.; Hospet, R.; Harish, E.R.; Purushotham, P.; Mujeeb, M.A.; Shrinivas, J.; David, M.; Mundaragi, A.C.; Thimmappa, S.C. Nanoagrotechnology for soil quality, crop performance and environmental management. In Nanotechnology; Springer: Singapore, 2017; pp. 73–97. [Google Scholar]
- Duhan, J.S.; Kumar, R.; Kumar, N.; Kaur, P.; Nehra, K.; Duhan, S. Nanotechnology: The new perspective in precision agriculture. Biotechnol. Rep. 2017, 15, 11–23. [Google Scholar] [CrossRef]
- Bernkop-Schnürch, A.; Dünnhaupt, S. Chitosan-based drug delivery systems. Eur. J. Pharm. Biopharm. 2012, 81, 463–469. [Google Scholar] [CrossRef]
- Aranaz, I.; Harris, R.; Heras, A. Chitosan amphiphilic derivatives. Chemistry and applications. Curr. Org. Chem. 2010, 14, 308–330. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Kaur, S.; Brar, S.K.; Verma, M. Green synthesis approach: Extraction of chitosan from fungus mycelia. Crit. Rev. Biotechnol. 2013, 33, 379–403. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2014. [Google Scholar]
- Yan, N.; Chen, X. Sustainability: Don’t waste seafood waste. Nature 2015, 524, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbio. 2010, 144, 51–63. [Google Scholar] [CrossRef]
- Campos, E.V.R.; de Oliveira, J.L.; Fraceto, L.F.; Singh, B. Polysaccharides as safer release systems for agrochemicals. Agron. Sustain. Dev. 2015, 35, 47–66. [Google Scholar] [CrossRef]
- Hernández-Téllez, C.N.; Plascencia-Jatomea, M.; Cortez-Rocha, M.O. Chitosan-based bionanocomposites: Development and perspectives in food and agricultural applications. In Chitosan in the Preservation of Agricultural Commodities; Elsevier: Cambridge, MA, USA, 2016; pp. 315–338. [Google Scholar]
- Kashyap, P.L.; Xiang, X.; Heiden, P. Chitosan nanoparticle based delivery systems for sustainable agriculture. Int. J. Biol. Macromol. 2015, 77, 36–51. [Google Scholar] [CrossRef]
- Agarwal, M.; Nagar, D.; Srivastava, N.; Agarwal, M. Chitosan nanoparticles based drug delivery: An update. Int, J. Adv. Multidiscip. Res. 2015, 2, 1–13. [Google Scholar]
- Dudhani, A.R.; Kosaraju, S.L. Bioadhesive chitosan nanoparticles: Preparation and characterization. Carbohydr. Polym. 2010, 81, 243–251. [Google Scholar] [CrossRef]
- Agnihotri, S.A.; Mallikarjuna, N.N.; Aminabhavi, T.M. Recent advances on chitosan-based micro-and nanoparticles in drug delivery. J. Control. Release 2004, 100, 5–28. [Google Scholar] [CrossRef]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, R.C.; Kumaraswamy, R.; Kumari, S.; Sharma, S.; Pal, A.; Raliya, R.; Biswas, P.; Saharan, V. Cu-chitosan nanoparticle boost defense responses and plant growth in maize (Zea mays L.). Sci. Rep. 2017, 7, 9754. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, P.; Dapkekar, A.; Oak, M.D.; Paknikar, K.M.; Rajwade, J.M. Zinc complexed chitosan/TPP nanoparticles: A promising micronutrient nanocarrier suited for foliar application. Carbohydr. Polym. 2017, 165, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Maluin, F.N.; Hussein, M.Z.; Yusof, N.A.; Fakurazi, S.; Idris, A.S.; Hilmi, Z.; Hailini, N.; Jeffery Daim, L.D. Preparation of chitosan–hexaconazole nanoparticles as fungicide nanodelivery system for combating Ganoderma disease in oil palm. Molecules 2019, 24, 2498. [Google Scholar] [CrossRef]
- Maluin, F.N.; Hussein, M.Z.; Yusof, N.A.; Fakurazi, S.; Idris, A.S.; Hilmi, N.H.Z.; Jeffery Daim, L.D. A Potent antifungal agent for basal stem rot disease treatment in oil palms based on chitosan-dazomet nanoparticles. Int. J. Mol. Sci. 2019, 20, 2247. [Google Scholar] [CrossRef]
- Maluin, F.N.; Hussein, M.Z.; Yusof, N.A.; Fakurazi, S.; Abu Seman, I.; Zainol Hilmi, N.H.; Jeffery Daim, L.D. Enhanced fungicidal efficacy on Ganoderma boninense by simultaneous co-delivery of hexaconazole and dazomet from their chitosan nanoparticles. RSC Adv. 2019, 9, 27083–27095. [Google Scholar] [CrossRef]
- Sun, C.; Shu, K.; Wang, W.; Ye, Z.; Liu, T.; Gao, Y.; Zheng, H.; He, G.; Yin, Y. Encapsulation and controlled release of hydrophilic pesticide in shell cross-linked nanocapsules containing aqueous core. Int. J. Pharm. 2014, 463, 108–114. [Google Scholar] [CrossRef]
- Kumar, S.; Chauhan, N.; Gopal, M.; Kumar, R.; Dilbaghi, N. Development and evaluation of alginate–chitosan nanocapsules for controlled release of acetamiprid. Int. J. Biol. Macromol. 2015, 81, 631–637. [Google Scholar] [CrossRef]
- Neri-Badang, M.C.; Chakraborty, S. Carbohydrate polymers as controlled release devices for pesticides. J. Carbohydr. Chem. 2019, 38, 67–85. [Google Scholar] [CrossRef]
- Minh, H.D.; Anh, D.N. Study on chitosan nanoparticles on biophysical characteristics and growth of Robusta coffee in green house. Biocatal. Agric. Biotechnol. 2013, 2, 289–294. [Google Scholar]
- Zayed, M.; Elkafafi, S.; Zedan, A.M.; Dawoud, S.F. Effect of nano chitosan on growth, physiological and biochemical parameters of Phaseolus vulgaris under salt stress. J. Plant Production 2017, 8, 577–585. [Google Scholar] [CrossRef]
- Abdel-Aziz, H.M.; Hasaneen, M.N.; Omer, A.M. Nano chitosan-NPK fertilizer enhances the growth and productivity of wheat plants grown in sandy soil. Spani. J. Agric. Res. 2016, 14, 0902. [Google Scholar] [CrossRef]
- Saharan, V.; Kumaraswamy, R.; Choudhary, R.C.; Kumari, S.; Pal, A.; Raliya, R.; Biswas, P. Cu-chitosan nanoparticle mediated sustainable approach to enhance seedling growth in maize by mobilizing reserved food. J. Agr. Food Chem. 2016, 64, 6148–6155. [Google Scholar] [CrossRef] [PubMed]
- Santo Pereira, A.E.; Silva, P.M.; Oliveira, J.L.; Oliveira, H.C.; Fraceto, L.F. Chitosan nanoparticles as carrier systems for the plant growth hormone gibberellic acid. Colloids Surf. B Biointerfaces 2017, 150, 141–152. [Google Scholar] [CrossRef]
- Pereira, A.; Sandoval-Herrera, I.; Zavala-Betancourt, S.; Oliveira, H.; Ledezma-Pérez, A.; Romero, J.; Fraceto, L. γ-Polyglutamic acid/chitosan nanoparticles for the plant growth regulator gibberellic acid: Characterization and evaluation of biological activity. Carbohydr. Polym. 2017, 157, 1862–1873. [Google Scholar] [CrossRef]
- Muthukrishnan, S.; Murugan, I.; Selvaraj, M. Chitosan nanoparticles loaded with thiamine stimulate growth and enhances protection against wilt disease in Chickpea. Carbohydr. Polym. 2019, 212, 169–177. [Google Scholar] [CrossRef]
- Sahab, A.; Waly, A.; Sabbour, M.; Nawar, L.S. Synthesis, antifungal and insecticidal potential of Chitosan (CS)-g-poly (acrylic acid)(PAA) nanoparticles against some seed borne fungi and insects of soybean. Int. J. Chem. Tech. Res 2015, 8, 589–598. [Google Scholar]
- Xing, K.; Shen, X.; Zhu, X.; Ju, X.; Miao, X.; Tian, J.; Feng, Z.; Peng, X.; Jiang, J.; Qin, S. Synthesis and in vitro antifungal efficacy of oleoyl-chitosan nanoparticles against plant pathogenic fungi. Int. J. Biol. Macromol. 2016, 82, 830–836. [Google Scholar] [CrossRef]
- Sathiyabama, M.; Parthasarathy, R. Biological preparation of chitosan nanoparticles and its in vitro antifungal efficacy against some phytopathogenic fungi. Carbohydr. Polym. 2016, 151, 321–325. [Google Scholar] [CrossRef]
- Barrera-Necha, L.L.; Correa-Pacheco, Z.N.; Bautista-Baños, S.; Hernández-López, M.; Jiménez, J.E.M.; Mejía, A.F.M. Synthesis and characterization of chitosan nanoparticles loaded botanical extracts with antifungal activity on Colletotrichum gloeosporioides and Alternaria species. Adv. Microbiol. 2018, 8, 286. [Google Scholar]
- Kheiri, A.; Jorf, S.M.; Malihipour, A.; Saremi, H.; Nikkhah, M. Synthesis and characterization of chitosan nanoparticles and their effect on Fusarium head blight and oxidative activity in wheat. Int. J. Biol. Macromol. 2017, 102, 526–538. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Guo, J.; Wu, D.; Tan, M.; Xiong, X.; Yin, Y.; He, G. Photo-responsive shell cross-linked micelles based on carboxymethyl chitosan and their application in controlled release of pesticide. Carbohydr. Polym. 2015, 132, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.-H.; Peng, L.-F. Synthesis and characterization of carboxymethyl chitosan carrying ricinoleic functions as an emulsifier for azadirachtin. Carbohydr. Polym. 2012, 88, 576–582. [Google Scholar] [CrossRef]
- Asgari-Targhi, G.; Iranbakhsh, A.; Ardebili, Z.O. Potential benefits and phytotoxicity of bulk and nano-chitosan on the growth, morphogenesis, physiology, and micropropagation of Capsicum annuum. Plant Physiol. Biochem. 2018, 127, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Khati, P.; Chaudhary, P.; Gangola, S.; Bhatt, P.; Sharma, A. Nanochitosan supports growth of Zea mays and also maintains soil health following growth. 3 Biotech 2017, 7, 81. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, H.; Hasaneen, M.N.; Omar, A. Effect of foliar application of nano chitosan NPK fertilizer on the chemical composition of wheat grains. Egypt. J. Bot. 2018, 58, 87–95. [Google Scholar] [CrossRef]
- Hasaneen, M.; Abdel-aziz, H.M.M.; Omer, A.M. Effect of foliar application of engineered nanomaterials: Carbon nanotubes NPK and chitosan nanoparticles NPK fertilizer on the growth of French bean plant. Biochem. Biotechnol. Res. 2016, 4, 68–76. [Google Scholar]
- Khalifa, N.S.; Hasaneen, M.N. The effect of chitosan–PMAA–NPK nanofertilizer on Pisum sativum plants. 3 Biotech 2018, 8, 193. [Google Scholar] [CrossRef]
- Kaur, P.; Duhan, J.S.; Thakur, R. Comparative pot studies of chitosan and chitosan-metal nanocomposites as nano-agrochemicals against Fusarium wilt of chickpea (Cicer arietinum L.). Biocatal. Agric. Biotechnol. 2018, 14, 466–471. [Google Scholar] [CrossRef]
- Kheiri, A.; Jorf, S.M.; Malihipour, A.; Saremi, H.; Nikkhah, M. Application of chitosan and chitosan nanoparticles for the control of Fusarium head blight of wheat (Fusarium graminearum) in vitro and greenhouse. Int. J. Biol. Macromol. 2016, 93, 1261–1272. [Google Scholar] [CrossRef]
- Brunel, F.; El Gueddari, N.E.; Moerschbacher, B.M. Complexation of copper (II) with chitosan nanogels: Toward control of microbial growth. Carbohydr. Polym. 2013, 92, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, M.K.; Joshi, A.; Sharma, S.; Saharan, V. Effect of laboratory synthesized Cu-Chitosan nanocomposites on control of PFSR disease of Maize caused by Fusarium verticillioids. Int. J. Curr. Microbiol. Appl. Sci 2017, 6, 1656–1664. [Google Scholar] [CrossRef]
- Manikandan, A.; Sathiyabama, M. Preparation of chitosan nanoparticles and its effect on detached rice leaves infected with Pyricularia grisea. Int. J. Biol. Macromol. 2016, 84, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Sathiyabama, M.; Manikandan, A. Chitosan nanoparticle induced defense responses in fingermillet plants against blast disease caused by Pyricularia grisea (Cke.) Sacc. Carbohydr. Polym. 2016, 154, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Sathiyabama, M.; Manikandan, A. Application of copper-chitosan nanoparticles stimulate growth and induce resistance in finger millet (Eleusine coracana Gaertn.) plants against blast disease. J. Agri. Food Chem. 2018, 66, 1784–1790. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.; Thi, T.V.; Nguyen, T.-T.; Le, T.D.; Vo, D.M.H.; Nguyen, D.H.; Nguyen, C.K.; Nguyen, D.C.; Nguyen, T.T.; Bach, L.G. Investigation of chitosan nanoparticles loaded with protocatechuic acid (PCA) for the resistance of Pyricularia oryzae fungus against rice blast. Polymers 2019, 11, 177. [Google Scholar]
- Xing, K.; Liu, Y.; Shen, X.; Zhu, X.; Li, X.; Miao, X.; Feng, Z.; Peng, X.; Qin, S. Effect of O-chitosan nanoparticles on the development and membrane permeability of Verticillium dahliae. Carbohydr. Polym. 2017, 165, 334–343. [Google Scholar] [CrossRef]
- Chauhan, N.; Dilbaghi, N.; Gopal, M.; Kumar, R.; Kim, K.-H.; Kumar, S. Development of chitosan nanocapsules for the controlled release of hexaconazole. Int. J. Biol. Macromol. 2017, 97, 616–624. [Google Scholar] [CrossRef]
- Xu, L.; Cao, L.-D.; Li, F.-M.; Wang, X.-J.; Huang, Q.-L. Utilization of chitosan-lactide copolymer nanoparticles as controlled release pesticide carrier for pyraclostrobin against Colletotrichum gossypii Southw. J. Disper. Sci. Technol. 2014, 35, 544–550. [Google Scholar] [CrossRef]
- Pham, D.C.; Nguyen, T.H.; Ngoc, U.T.P.; Le, N.T.T.; Tran, T.V.; Nguyen, D.H. Preparation, characterization and antifungal properties of chitosan-silver nanoparticles synergize fungicide against Pyricularia oryzae. J. Nanosci. Nanotechnol. 2018, 18, 5299–5305. [Google Scholar] [CrossRef]
- Liang, W.; Yu, A.; Wang, G.; Zheng, F.; Hu, P.; Jia, J.; Xu, H. A novel water-based chitosan-La pesticide nanocarrier enhancing defense responses in rice (Oryza sativa L) growth. Carbohydr. Polym. 2018, 199, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Ding, G.; Niu, J.; Zhang, W.; Tang, G.; Liang, Y.; Fan, C.; Dong, H.; Yang, J.; Li, J. Preparation and characterization of tebuconazole metal-organic framework-based microcapsules with dual-microbicidal activity. Chem. Eng. 2019, 359, 225–232. [Google Scholar] [CrossRef]
- Maruyama, C.R.; Guilger, M.; Pascoli, M.; Bileshy-José, N.; Abhilash, P.; Fraceto, L.F.; De Lima, R. Nanoparticles based on chitosan as carriers for the combined herbicides imazapic and imazapyr. Sci. Rep. 2016, 6, 19768. [Google Scholar] [CrossRef]
- Namasivayam, K.R.S.; Aruna, A.; Gokila. Evaluation of silver nanoparticles-chitosan encapsulated synthetic herbicide paraquate (AgNp-CS-PQ) preparation for the controlled release and improved herbicidal activity against Eichhornia crassipes. Res. J. Biotechnol. 2014, 9, 19–27. [Google Scholar]
- Liang, W.; Yu, A.; Wang, G.; Zheng, F.; Jia, J.; Xu, H. Chitosan-based nanoparticles of avermectin to control pine wood nematodes. Int. J. Biol. Macromol. 2018, 112, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: A review. Agron. Sustain. Dev. 2015, 35, 569–588. [Google Scholar] [CrossRef]
- Cruz-Romero, M.; Murphy, T.; Morris, M.; Cummins, E.; Kerry, J. Antimicrobial activity of chitosan, organic acids and nano-sized solubilisates for potential use in smart antimicrobially-active packaging for potential food applications. Food Control 2013, 34, 393–397. [Google Scholar] [CrossRef]
- Helander, I.; Nurmiaho-Lassila, E.-L.; Ahvenainen, R.; Rhoades, J.; Roller, S. Chitosan disrupts the barrier properties of the outer membrane of Gram-negative bacteria. Int. J. Food Microbiol. 2001, 71, 235–244. [Google Scholar] [CrossRef]
- Lopez-Moya, F.; Suarez-Fernandez, M.; Lopez-Llorca, L.V. Molecular mechanisms of chitosan interactions with fungi and plants. Int. J. Mol. Sci. 2019, 20, 332. [Google Scholar] [CrossRef]
- Hyun, M.W.; Yun, Y.H.; Kim, J.Y.; Kim, S.H. Fungal and plant phenylalanine ammonia-lyase. Mycobiology 2011, 39, 257–265. [Google Scholar] [CrossRef]
- Romanazzi, G.; Nigro, F.; Ippolito, A.; Divenere, D.; Salerno, M. Effects of pre-and postharvest chitosan treatments to control storage grey mold of table grapes. J. Food Sci. 2002, 67, 1862–1867. [Google Scholar] [CrossRef]
- Li, B.; Liu, B.; Shan, C.; Ibrahim, M.; Lou, Y.; Wang, Y.; Xie, G.; Li, H.Y.; Sun, G. Antibacterial activity of two chitosan solutions and their effect on rice bacterial leaf blight and leaf streak. Pest Manag. Sci. 2013, 69, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Li, S.j.; Zhu, T.H. Biochemical response and induced resistance against anthracnose (Colletotrichum camelliae) of camellia (Camellia pitardii) by chitosan oligosaccharide application. Forest Pathol. 2013, 43, 67–76. [Google Scholar] [CrossRef]
- Guan, Y.-J.; Hu, J.; Wang, X.-J.; Shao, C.-X. Seed priming with chitosan improves maize germination and seedling growth in relation to physiological changes under low temperature stress. J. Zhejiang Univ. Sci. B 2009, 10, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Foley, R.C.; Kidd, B.N.; Hane, J.K.; Anderson, J.P.; Singh, K.B. Reactive oxygen species play a role in the infection of the necrotrophic fungi, Rhizoctonia solani in wheat. PLoS ONE 2016, 11, e0152548. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Yang, L.; Kennedy, J.F.; Tian, S. Effects of chitosan and oligochitosan on growth of two fungal pathogens and physiological properties in pear fruit. Carbohydr. Polym. 2010, 81, 70–75. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, L.; Yan, H.; Kennedy, J.F.; Meng, X. Chitosan and oligochitosan enhance the resistance of peach fruit to brown rot. Carbohydr. Polym. 2013, 94, 272–277. [Google Scholar] [CrossRef]
- Yin, H.; Zhao, X.; Du, Y. Oligochitosan: A plant diseases vaccine—A review. Carbohydr. Polym. 2010, 82, 1–8. [Google Scholar] [CrossRef]
- Burketova, L.; Trda, L.; Ott, P.G.; Valentova, O. Bio-based resistance inducers for sustainable plant protection against pathogens. Biotechnol. Adv. 2015, 33, 994–1004. [Google Scholar] [CrossRef]
- Hadwiger, L.A.; Beckman, J.M. Chitosan as a component of pea-Fusarium solani interactions. Plant Physiol. 1980, 66, 205–211. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Couderchet, M.; Vernet, G.; Aziz, A. Chitosan stimulates defense reactions in grapevine leaves and inhibits development of Botrytis cinerea. Eur. J. Plant Pathol. 2006, 114, 405–413. [Google Scholar] [CrossRef]
- Awadalla, O.A.; Mahamoud, G. New chitosan derivatives induced resistance to Fusarium wilt disease through phytoalexin (Gossypol) production. Sains Malays. 2005, 34, 141–146. [Google Scholar]
- Yin, H.; Fretté, X.C.; Christensen, L.P.; Grevsen, K. Chitosan oligosaccharides promote the content of polyphenols in Greek oregano (Origanum vulgare ssp. hirtum). J. Agri. Food Chem. 2011, 60, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Nedukha, O. Callose: Localization, functions, and synthesis in plant cells. Cytol. Genet. 2015, 49, 49–57. [Google Scholar] [CrossRef]
- Iriti, M.; Faoro, F. Chitosan as a MAMP, searching for a PRR. Plant Signal. Behav. 2009, 4, 66–68. [Google Scholar] [CrossRef]
- Ellinger, D.; Voigt, C.A. Callose biosynthesis in Arabidopsis with a focus on pathogen response: What we have learned within the last decade. Ann. Bot. 2014, 114, 1349–1358. [Google Scholar] [CrossRef]
- Jabeen, N.; Ahmad, R. The activity of antioxidant enzymes in response to salt stress in safflower (Carthamus tinctorius L.) and sunflower (Helianthus annuus L.) seedlings raised from seed treated with chitosan. J. Sci. Food Agr. 2013, 93, 1699–1705. [Google Scholar] [CrossRef]
- Sharathchandra, R.; Raj, S.N.; Shetty, N.P.; Amruthesh, K.; Shetty, H.S. A Chitosan formulation Elexa™ induces downy mildew disease resistance and growth promotion in pearl millet. Crop Prot. 2004, 23, 881–888. [Google Scholar] [CrossRef]
- Abbasi, N.A.; Iqbal, Z.; Maqbool, M.; Hafiz, I.A. Postharvest quality of mango (Mangifera indica L.) fruit as affected by chitosan coating. Pak. J. Bot 2009, 41, 343–357. [Google Scholar]
- Langsdorf, G. Defence of cultivated plants. In Handbook of Agriculture; Yuste, M.-P., Gostincar, J., Eds.; Marcel Dekker, Inc., Urban & Fischer: München, Germany.
- Petit, A.-N.; Fontaine, F.; Vatsa, P.; Clément, C.; Vaillant-Gaveau, N. Fungicide impacts on photosynthesis in crop plants. Photosynth. Res. 2012, 111, 315–326. [Google Scholar] [CrossRef]
- Nair, R.; Varghese, S.H.; Nair, B.G.; Maekawa, T.; Yoshida, Y.; Kumar, D.S. Nanoparticulate material delivery to plants. Plant Sci. 2010, 179, 154–163. [Google Scholar] [CrossRef]
- Adisa, I.O.; Pullagurala, V.L.R.; Peralta-Videa, J.R.; Dimkpa, C.O.; Elmer, W.H.; Gardea-Torresdey, J.; White, J. Recent advances in nano-enabled fertilizers and pesticides: A critical review of mechanisms of action. Environ. Sci- Nano 2019, 6, 2002–2030. [Google Scholar] [CrossRef]
- Lv, J.; Christie, P.; Zhang, S. Uptake, translocation, and transformation of metal-based nanoparticles in plants: Recent advances and methodological challenges. Environ. Sci.-Nano 2019, 6, 41–59. [Google Scholar] [CrossRef]
- Eichert, T.; Kurtz, A.; Steiner, U.; Goldbach, H.E. Size exclusion limits and lateral heterogeneity of the stomatal foliar uptake pathway for aqueous solutes and water-suspended nanoparticles. Physiol. Plant. 2008, 134, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Eichert, T.; Goldbach, H.E. Equivalent pore radii of hydrophilic foliar uptake routes in stomatous and astomatous leaf surfaces–further evidence for a stomatal pathway. Physiol. Plant. 2008, 132, 491–502. [Google Scholar] [CrossRef]
- Popp, C.; Burghardt, M.; Friedmann, A.; Riederer, M. Characterization of hydrophilic and lipophilic pathways of Hedera helix L. cuticular membranes: Permeation of water and uncharged organic compounds. J. Exp. Bot. 2005, 56, 2797–2806. [Google Scholar]
- Miralles, P.; Church, T.L.; Harris, A.T. Toxicity, uptake, and translocation of engineered nanomaterials in vascular plants Environ. Sci. Technol. 2012, 46, 9224–9239. [Google Scholar] [CrossRef]
- Parthasarathi, T. Phytotoxicity of nanoparticles in agricultural crops. In Proceedings of the International Conference on Green Technology and Environmental Conservation (GTEC-2011), Chennai, India, 15–17 December 2011; pp. 51–60. [Google Scholar]
- Peterson, C.A.; Edgington, L. Transport of the systemic fungicide, benomyl, in bean plants. Phytopathology 1970, 60, 475–478. [Google Scholar] [CrossRef]
- Rees, R.; Rutledge, J.; Newnam, M. Compositions and Methods for Enhancing Plant Quality. WO2013067103A3, 27 July 2013. [Google Scholar]
- Abdel-Aziz, H.M.M.; Hasaneen, M.N.A.-G.; Omer, A.M. Foliar application of nano chitosan NPK fertilizer improves the yield of wheat plants grown on two different soils. Egypt. J. Exp. Biol. (Bot.) 2018, 14, 63–72. [Google Scholar]
- Chuang, Y.-H.; Liu, C.-H.; Sallach, J.B.; Hammerschmidt, R.; Zhang, W.; Boyd, S.A.; Li, H. Mechanistic study on uptake and transport of pharmaceuticals in lettuce from water. Environ. Int. 2019, 131, 104976. [Google Scholar] [CrossRef]
- Oliveira, M.E.F.; Silva, É.C.G.; Câmara, C.A.; Souza, I.A.d.; Amorim, R.V.S. Evaluation of acute toxicity of β-lapachone associated with chitosan as a cytoprotective agent. J. Bras. Patol. Med. Lab. 2018, 54, 279–287. [Google Scholar] [CrossRef]
- Damalas, C.; Koutroubas, S. Farmers’ exposure to pesticides: Toxicity types and ways of prevention. Toxics 2016, 4, 1. [Google Scholar] [CrossRef]
- Vuković, S.; Inđić, D.; Gvozdenac, S. Phytotoxic effects of fungicides, insecticides and nonpesticidal components on pepper depending on water quality. Pestic. Fitomed. 2014, 29, 145–153. [Google Scholar] [CrossRef]
- Hong, D.; Park, M.; Yang, S.H.; Lee, J.; Kim, Y.-G.; Choi, I.S. Artificial spores: Cytoprotective nanoencapsulation of living cells. Trends Biotechnol. 2013, 31, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Hong, D.; Lee, J.; Choi, I.S. Cell-in-shell hybrids: Chemical nanoencapsulation of individual cells. Acc. Chem. Res. 2016, 49, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Behboudi, F.; Sarvestani, Z.T.; Kassaee, M.Z.; Sanavi, S.A.M.M.; Sorooshzadeh, A. Phytotoxicity of chitosan and SiO2 nanoparticles to seed germination of wheat (Triticum aestivum L.) and barley (Hordeum vulgare L.) plants. Not. Sci. Biol. 2017, 9, 242–249. [Google Scholar] [CrossRef]
- Sunkireddy, P.; Kanwar, R.K.; Ram, J.; Kanwar, J.R. Ultra-small algal chitosan ocular nanoparticles with iron-binding milk protein prevents the toxic effects of carbendazim pesticide. Nanomedicine 2016, 11, 495–511. [Google Scholar] [CrossRef]
- Xu, C.; Cao, L.; Zhao, P.; Zhou, Z.; Cao, C.; Zhu, F.; Li, F.; Huang, Q. Synthesis and characterization of stimuli-responsive poly (2-dimethylamino-ethylmethacrylate)-grafted chitosan microcapsule for controlled pyraclostrobin release. Int. J. Mol. Sci. 2018, 19, 854. [Google Scholar] [CrossRef]
- Real, D.; Hoffmann, S.; Leonardi, D.; Salomon, C.; Goycoolea, F.M. Chitosan-based nanodelivery systems applied to the development of novel triclabendazole formulations. PLoS ONE 2018, 13, e0207625. [Google Scholar] [CrossRef]
- Manuja, A.; Kumar, B.; Kumar, R.; Chopra, M.; Dilbaghi, N.; Kumar, S.; Yadav, S.C. Encapsulated quinapyramine sulfate-loaded chitosan/mannitol nanoparticles: Biocompatibility and targeting efficiency in rabbit model of trypanosomosis. Antimicrob. Agents Chemother. 2018, AAC. 00466-18. [Google Scholar] [CrossRef]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide exposure, safety issues, and risk assessment indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef] [PubMed]
- Jeyaratnam, J. Acute Pesticide Poisoning: A Major Global Health Problem; World Health Statistics Organization: Geneva, Switzerland, 1990; Volume 43, pp. 139–144. [Google Scholar] [PubMed]
- Eddleston, M.; Karalliedde, L.; Buckley, N.; Fernando, R.; Hutchinson, G.; Isbister, G.; Konradsen, F.; Murray, D.; Piola, J.C.; Senanayake, N. Pesticide poisoning in the developing world—a minimum pesticides list. Lancet 2002, 360, 1163–1167. [Google Scholar] [CrossRef]
- Chen, H.; Yao, J.; Wang, F.; Choi, M.M.; Bramanti, E.; Zaray, G. Study on the toxic effects of diphenol compounds on soil microbial activity by a combination of methods. J. Hazard. Mater. 2009, 167, 846–851. [Google Scholar] [PubMed]
- Ramakrishnan, B.; Megharaj, M.; Venkateswarlu, K.; Sethunathan, N.; Naidu, R. Mixtures of environmental pollutants: Effects on microorganisms and their activities in soils. In Reviews of Environmental Contamination and Toxicology Volume 211; Springer: New York, NY, USA, 2011; pp. 63–120. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Nanoformulations, Molecular Weight (MW), Deacetylation Degree and Final pH of the Product | Plant and Application Type | Average Size * and Zeta Potential | Findings | Ref. |
|---|---|---|---|---|
| Nano-chitosan, 600 kDa, 85%, pH 6.0 | Robusta coffee (Coffea canephora), foliar spray | 420, 750 and 970 nm c | Increase chlorophyll content (30–50%), enhance nutrient uptake (10–27% N, 17–30% P, 30–45% K) and photosynthesis rate (30%). | [34] |
| Nano-chitosan, 110 kDa, 85%–90%, pH 4.0 | Chilli (Capsicum annuum), seed treatment | 163 nm a, +60.4 mV | Enhance in total root and leaf fresh mass up to 77% and 28%, respectively upon application of 1 mg/L of nano-chitosan. The increase of leaf catalase (33%) and peroxidase activities (23%) also been observed. | [48] |
| Nano-chitosan, 100–399 kDa, | Bean (Phaseolus vulgaris), seed treatment | 46 nm a | Promote seed germination (123% after 72 h) and radical length (231% after 72 h) under salinity stress. | [35] |
| Nano-chitosan, pH 7.0–9.0 | Maize (Zea mays), seed treatment | 80–100 nm d | Promote seed germination (37%), plant height (1.5-fold increase) and leaf area (2-fold increase). | [49] |
| Nano-chitosan, pH 4.8 | Chickpea (Cicer arietinum), seed treatment | 10–30 nm b, −37 mV | Enhance germination (100%), seedling vigor index (57%) and vegetative biomass of seedlings (3-fold). | [43] |
| Chitosan-polymethacrylic acid-NPK nanoparticles | Wheat (Triticumaestivum), foliar spray | 26 and 31 nm b | Enhance harvest index (24%), crop yield (59%), and mobilization index (42%). | [36] |
| 20 nm b | Enhance polysaccharides (10%) and total saccharides (11%). | [50] | ||
| French bean (Phaseolus vulgaris), foliar spray | 20 nm b | Enhance plant growth, nutrient uptake, and biomass accumulation. The nanoformulations was found on the leaf phloem via HRTEM image | [51] | |
| Pea (Pisum sativum), seed treatment | 20 nm b | Induce mitotic cell division (1.5 fold) and enhance of total soluble protein (i.e., legumin β, vicilin 1, 2 and 3, and convicilin) | [52] | |
| Chitosan-Cu nanoparticles, low MW, 80% | Maize (Surya local), seed treatment | 150 nm b, +22.6 mV | Increase α-amylase and protease activity as well as promote seedling growth. | [37] |
| Chitosan-Cu nanoparticles, 50–190 kDa, 80% | Maize (Zea mays), foliar spray | 361 nm a,+22.1 mV | pH-responsive sustained release of Cu was obtained. Promote seedling growth (significant increase in plant height, stem diameter, and root length). | [26] |
| Chitosan-Zn nanoparticles, 60 kDa, 85% | Wheat (Triticum durum), foliar spray | 325 nm a, +42.3 mV | Stomatal localization of nanoparticles was observed. Increase grain zinc content by up to 42%. | [27] |
| Chitosan-γ-polyglutamic acid-gibberellic acid nanoparticles, 290 kDa, 75%–85%, pH 4.5 | French bean (Phaseolus vulgaris), seed treatment | 134 nm a, −29.0 mV | 61% of the encapsulation efficiency of hormone in the nanoformulation. Offer sustained-release with 58% after 48 h. Exhibited high biological activity with 50–70% enhance of seed germination, leaf area, and root development compared to counterpart. | [39] |
| Chitosan-gibberellic acid nanoparticles, 27 kDa, 75%–85%, pH 4.5 | French bean (Phaseolus vulgaris), seed treatment | 450 nm a, +27.0 mV | 90% of the encapsulation efficiency of hormone in the nanoformulation. Offer stability up to 60 days with pH and temperature-controlled release mechanism. Upon treatment, the seedlings showed an increase of leaf area, chlorophyll and carotenoids amount. | [38] |
| Chitosan-thiamine nanoparticles, 27 kDa, 85% | Chickpea (Cicer arietinum), seed treatment | 596 nm a, +37.7 mV | 99% of the encapsulation efficiency of hormone in the nanoformulation. Enhance seeds germination and induce more defense enzymes (peroxidase, glucanase, chitinase, polyphenol oxidase, protease, and chitosanase activity) and increase 10-fold auxins level compared to the untreated seeds. | [40] |
| Plant Pathogen | Nanoformulations, Average Size *, Zeta Potential and its Application | In Vitro/In Vivo | Findings | Ref. |
|---|---|---|---|---|
| Alternaria solani, Fusarium oxysporum, and Pyricularia grisea, | Nano-CS, 10-30 nm b, –37 mV (fungicides) | In vitro | High inhibition on mycelial growth with the percentage of inhibition rate recorded at 92%, 87%, and 72% for P. grisea, F. oxysporum and A. solani, respectively. | [43] |
| Aphis gossypii | CS-polyacrylic acid nanoparticles, 50 nm a (insecticides) | In vivo, reared on castor leaves | The mean number of eggs/females reduce significantly under the laboratory conditions and field conditions with 76% and 61%, respectively. | [41] |
| Callosobruchus chinensis | In vivo, reared on castor leaves | The mean number of eggs/females reduce significantly under the laboratory conditions and store conditions with 74% and 70%, respectively. | [41] | |
| Callosobruchus maculatus: | In vivo, reared on soybean | The mean number of eggs/females reduce significantly under the laboratory condition and store condition with 84% and 74%, respectively. | [41] | |
| Colletotrichum Gloeosporioides and Alternaria spp. | Nano-CS, 406 nm a, –4.9 to –7.9 mV (fungicides) | In vitro | Higher inhibition on mycelial (up to 82%) and sporulation of fungus, compared to the counterpart. Enhance seeds germination. | [44] |
| Curvularia lunata | CS-Cu nanoparticles, 361 nm a, +22.1 mV (fungicides) | In vitro and In vivo (Maize, Zea mays) | Induce more defense response: 1.5–2 fold of peroxidase, a significant amount of superoxide dismutase, 2–3 fold of phenylalanine ammonia-lyase, and a significant amount of polyphenol oxidase. | [26] |
| Fusarium oxysporum | CS-CuO, 350 nm b, –26.8 mV; CS-ZnO, 441 nm b, –24.5 mV; and CS-Ag, 348 nm b, –49.1 mV (fungicides) | In vitro and In vivo (chickpea, Cicer arietinum) | In vitro results showed that the antifungal activity follows: CS-ZnO > CS-CuO > CS-Ag, while in vivo results showed that the wilt disease reduction follows: CS-CuO (47%) > CS-ZnO (40%) > CS-Ag (33%). | [53] |
| Fusarium graminearum | Nano-CS, 181 nm a, +45.6 mV (fungicides) | In vitro and in vivo (wheat) | 85% inhibition of mycelial growth in plate treated with 5000 mg/mL of CS nanoparticles (in vitro) and 53% reduction in disease severity on wheat (in vivo). Deformation and dehydration of fungus mycelial growth also can be seen. | [54] |
| Nano-CS, [1] 181 nm a, +45.6 mV; [2] 310 nma, +33.2 mV; [3] 340 nm a, +21.7 mV (fungicides) | In vitro and in vivo (wheat) | Inhibition rate (%) at 1000 mg/mL follows: (1) Nano-CS (71.1%) > (3) Nano-CS (17.7%) > (2) Nano-CS (14.1%) | [45] | |
| CS-Cu nanoparticles, 220 nm a, +40.0 mV (fungicides) | In vitro | Minimum inhibitory concentration after one week incubation follows: Cu (250 µg/mL) > CS-Cu nanoparticles (17.5 mg/mL) > chitosan (10 mg/mL). | [55] | |
| Fusarium verticillioids | CS-Cu nanoparticles, 296 nm a, +19.6 mV (fungicides) | In vivo (Maize, Zea mays) | At 4 and 8 h after treatment, the disease has been reduced by 48% and 50%, respectively. | [56] |
| Pyricularia grisea | Nano-CS, 83 nm a, –28.0 mV (fungicides) | In vitro and In vivo (rice, Oryza sativa) | No inhibitory activity was observed in the in vitro. However, in vivo results revealed its ability in suppressing the disease with zero percent disease incidence at 10 days after infection, where 100% disease incidence was observed in control. | [57] |
| In vitro and In vivo (finger millet, Eleusine coracana) | In the in vitro evaluation, 65% of radial growth inhibition was obtained. Meanwhile, delayed disease symptom (25 days) and low disease infection (23%) was observed in the in vivo evaluation, while for control, the symptoms started appear in 15 days and 100% disease infection was recorded. Enhance in peroxidase activity level (reached maximum on day 50) also been observed. | [58] | ||
| CS-Cu nanoparticles, 88 nm a, –29.0 mV (fungicides) | In vitro and In vivo (finger millet, Eleusine coracana) | Induce resistance against the pathogen attack: a 2-fold increase in chitinase and chitosanase and produce more protease inhibitors, peroxidase, β-1,3 glucanase, and polyphenol oxidase compared to the untreated plant. | [59] | |
| Pyricularia oryzae | Nano-CS, 28 nm b, +49.0 to +53.0 mV and CS-protocatechuic acid, 33 nm b, +11.0 mV (fungicides) | In vitro | The diameter of inhibition zone follows: CS-protocatechuic acid nanoparticles > protocatechuic acid > chitosan nanoparticles. Up to a 3-fold increase of the inhibition zone compared to the counterpart. | [60] |
| Verticillium dahliae | Nano-oleoyl-CS, 297 nm c (fungicides) | In vitro | The nanoparticles internalized the fungal cell, hence leads to the deformation of spore and hyphae, thickened cell walls, cease of organelles and cytoplasmic vacuolation. | [61] |
| Agrochemicals Type and Its Active Ingredient | Nanocarrier Formulations, Loading Content % (LC), Loading Efficiency % (LE), Encapsulation Efficiency % (EE), and its Average Size * | Plant Pathogen | In Vitro/In Vivo | Findings | Ref. |
|---|---|---|---|---|---|
| Fungicide, Dazomet | CS nanoparticles, [1] 276 nm b, 28% (LC), 78% (EE); [2] 32 nm b, 48% (LC), 98% (EE); [3] 31 nm b, 35% (LC), 85% (EE); [4] 7 nm b, 33% (LC), 83% (EE) | Ganoderma boninense | In vitro | Controlled release with saturation release of 97.9% and half release time (t1/2) of 11 h at pH 5.5. Increase fungicidal activity up to 30-fold compared to their counterparts. | [29] |
| Fungicides, Hexaconazole and Dazomet | CS nanoparticles, [1] 157 nm b, 17% (LC), 67% (EE); [2] 58 nm b, 17% (LC), 67% (EE); [3] 31 nm b, 17% (LC), 67% (EE); [4] 5 nm b, 13% (LC), 64% (EE) | Ganoderma boninense | In vitro | Controlled release with half release time (t1/2) up to 66 and 19 h for hexaconazole and dazomet, respectively, at pH 5.5. Increase fungicidal activity up to 40-fold compared to their counterparts. | [30] |
| Fungicide, Hexaconazole | CS nanoparticles, 100 nm b, 73% (EE) | Rhizoctonia solani | In vitro | Controlled release with prolongs the release time of hexaconazole up to 14 days at pH 8.3 while the conventional pesticides only last up to 5 days. Significant higher antifungal activity compared to the conventional counterpart. | [62] |
| Fungicide, Hexaconazole | CS nanoparticles, [1] 272 nm b, 11% (LC), 56% (EE); [2] 169 nm b, 17% (LC), 67% (EE); [3] 32 nm b, 15% (LC), 65% (EE); [4] 18 nm b, 15% (LC), 65% (EE) | Ganoderma boninense | In vitro | Controlled release with saturation release of 99.9% and half release time (t1/2) of 42 h at pH 5.5. Increase fungicidal activity up to 3-fold compared to their counterparts. | [28] |
| Fungicide, Pyraclostrobin | CS-lactide nanoparticles, [1] 128 nm a, 18% (LC), 45% (EE); [2] 90 nm a, 11% (LC), 85% (EE); [3] 77 nm a, 2% (LC), 91% (EE); | Colletotrichum gossypii | In vitro | Better stability of AI under light stress with 81% compared to the counterpart with 41%. Controlled release (75%) of AI up to 10 h at pH 8.3. High fungicidal activity with up to 85% inhibition rate at day 7 of incubation. | [63] |
| Fungicide, Pyraclostrobin | Quarternized CS-silica nanoparticles, 110 nm b, 27%–42% (LC) | Phomopsis asparagi | In vitro | Controlled release (72%) with prolongs release time up to 13 h. Inhibition percentage of fungi up to 95% | [46] |
| Fungicides, Tricyclazole and Hexaconazole | CS-Ag nanoparticles, 17 nm b | Pyricularia oryzae | In vitro | Significantly increased the inhibition zone by 2-fold compared to the counterpart | [64] |
| Fungicide, Avermectin | CS-lanthanum-nanoparticles, 333 nm a, 46% (LE), 65% (EE) | Magnaporthe grisea | In vitro and In vivo | Rapid release on the first 36 h followed by sustained release until day-10. No inhibitory of fungus was observed in the in vitro study. However, significant disease reduction was observed in the in vivo study (Rice, Oryza sativa). | [65] |
| Fungicide, Tebuconazole | CS-porphyrinic-pectin nanoparticles, 100 nm c, 30% (LE) | Xanthomonas campestris, Pseudomonas syringae, and Alternaria alternate | In vitro | Metal-organic frameworks (MOFs) capsule comprise of chitosan, porous porhpyrinic, and pectin demonstrated a stimuli-responsive sustained release of AI with prolonged-release time up to 174 h at pH 7. The nanocapsule exhibited high antimicrobials activities and no phytotoxic effect on Chinese cabbage. | [66] |
| Herbicides, Imazapic, and Imazapyr | CS-alginate nanoparticles, 378 nm a, 62% (EE) of imazapic, 71% (EE) of imazapyr;CS-tripolyphosphate nanoparticles, 479 nm a, 59% (EE) of imazapic, 70% (EE) of imazapyr | Bidens pilosa | In vivo | After 300 min under gentle agitation, 30% (imazapic) and 20% (imazapyr) were released in CS-alginate nanoparticles, while 59% (imazapic) and 9% (imazapyr) were released in CS-tripolyphosphate nanoparticles. Meanwhile, free imazapic and imazapyr were released up to 55% and 97%, respectively, hence, showing the slow-release formulation of the nanoparticulate system. The encapsulation of herbicides also reduced the toxicity of herbicides against the nontarget organism while maintaining its herbicidal activity on the tested weeds. | [67] |
| Herbicide, Paraquat | CS-Ag nanoparticles, 100 nm c, 90% (EE) | Eichhornia crassipes | In vivo | Improved herbicidal activity on the tested weed with a 90% release of paraquat was observed for up to 24 h. Improved the microbial population, bacteria, and yeast compared to its free herbicide. | [68] |
| Nematicide, Avermectin | CS-γ-polyglutamic acid nanoparticles, 61 and 56 nm b, 31% (LC), 35% (EE) | Caenorhabditis elegans | In vitro | The controlled release rate governed by pH. The mortality rate of nematodes was significantly increased by 29%, compared to its counterpart. | [69] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maluin, F.N.; Hussein, M.Z. Chitosan-Based Agronanochemicals as a Sustainable Alternative in Crop Protection. Molecules 2020, 25, 1611. https://doi.org/10.3390/molecules25071611
Maluin FN, Hussein MZ. Chitosan-Based Agronanochemicals as a Sustainable Alternative in Crop Protection. Molecules. 2020; 25(7):1611. https://doi.org/10.3390/molecules25071611
Chicago/Turabian StyleMaluin, Farhatun Najat, and Mohd Zobir Hussein. 2020. "Chitosan-Based Agronanochemicals as a Sustainable Alternative in Crop Protection" Molecules 25, no. 7: 1611. https://doi.org/10.3390/molecules25071611
APA StyleMaluin, F. N., & Hussein, M. Z. (2020). Chitosan-Based Agronanochemicals as a Sustainable Alternative in Crop Protection. Molecules, 25(7), 1611. https://doi.org/10.3390/molecules25071611