Anti-Quorum Sensing Activity of Stevia Extract, Stevioside, Rebaudioside A and Their Aglycon Steviol

 ,
, 
 ,
,  , and
, and
Abstract
1. Introduction
2. Results
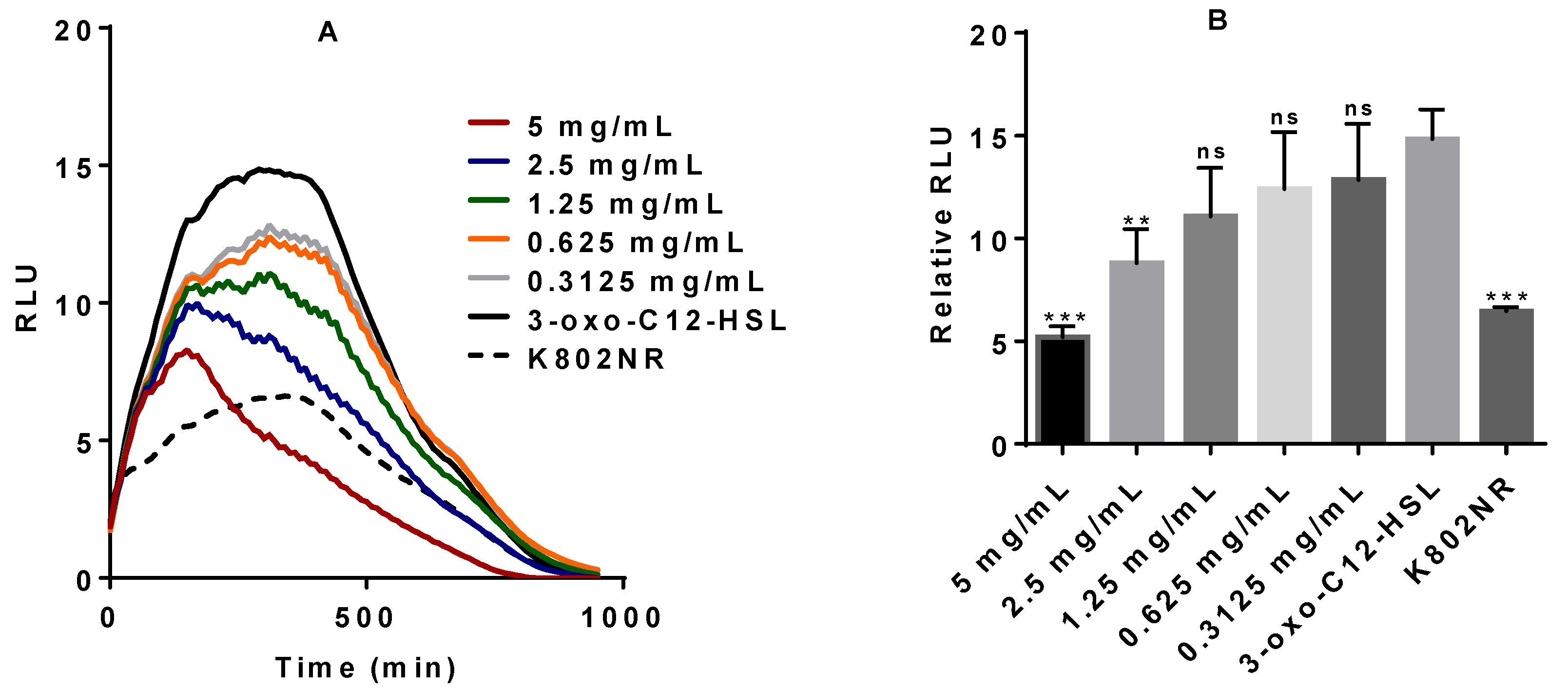
2.1. Anti-QS Activity of Commercial Stevia Herbal Supplement (CSHS)
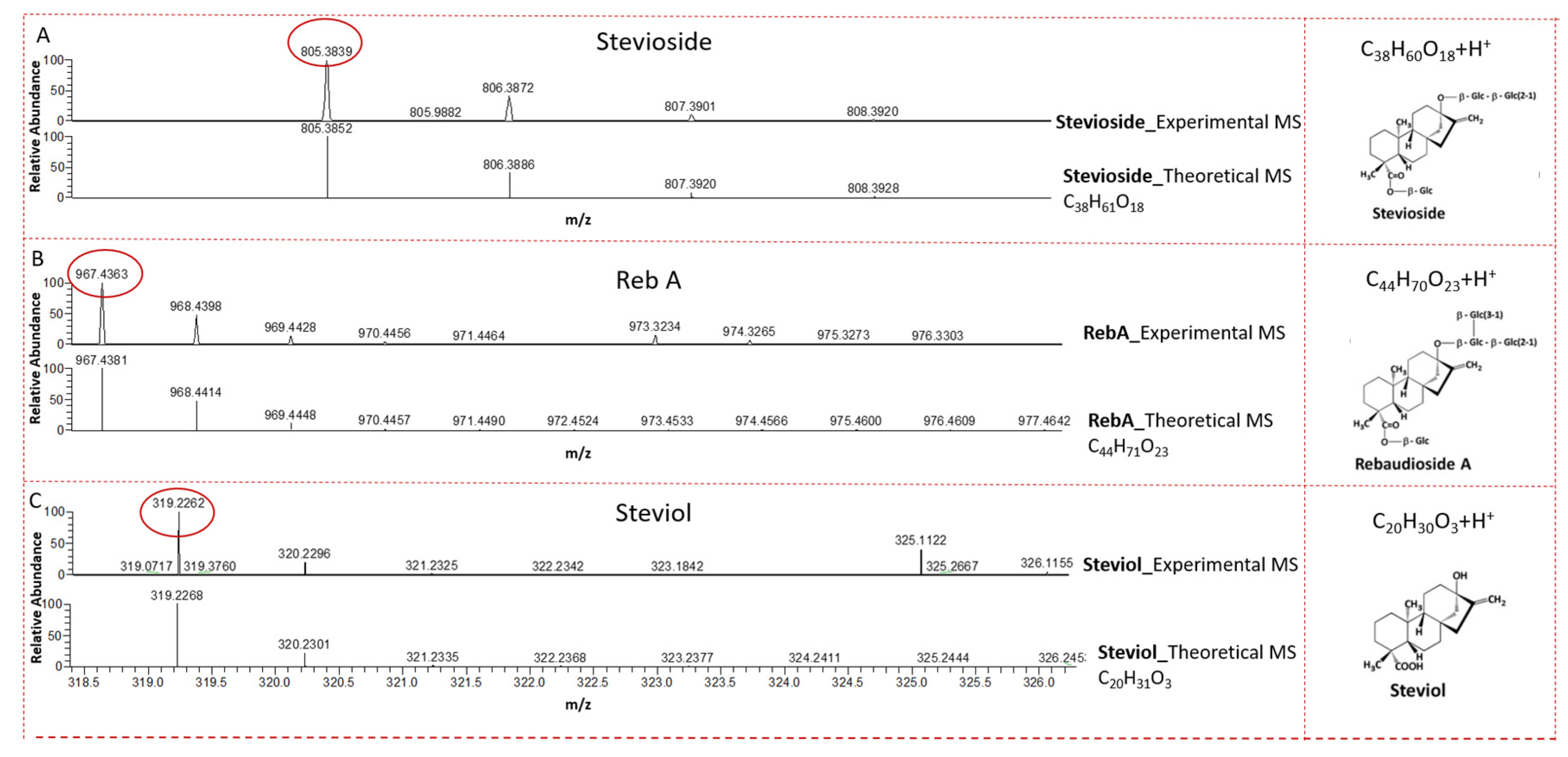
2.2. LC-MS Analysis of CSHS
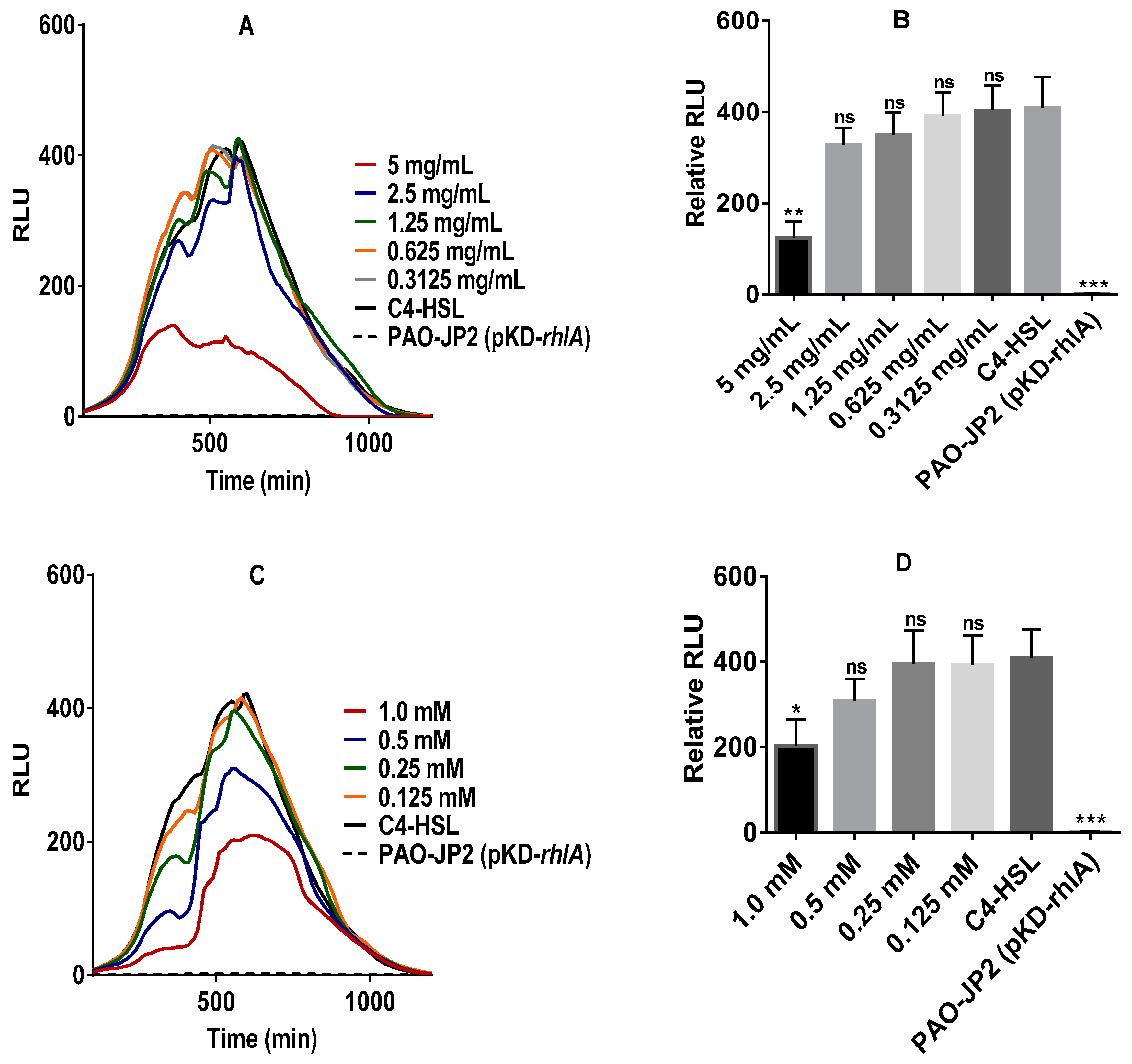
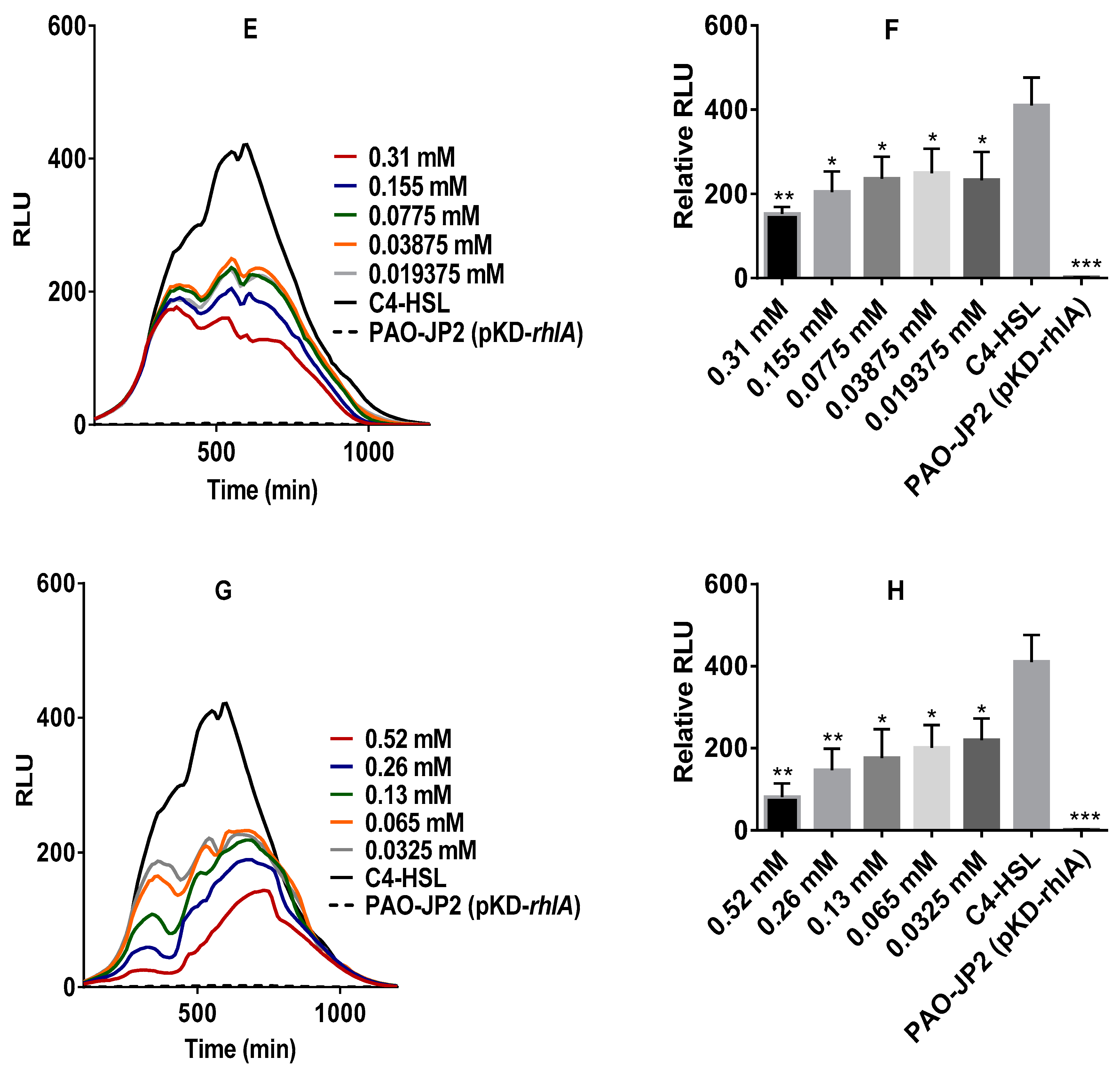
2.3. Anti-QS Activity of Pure Stevia-Derived Components
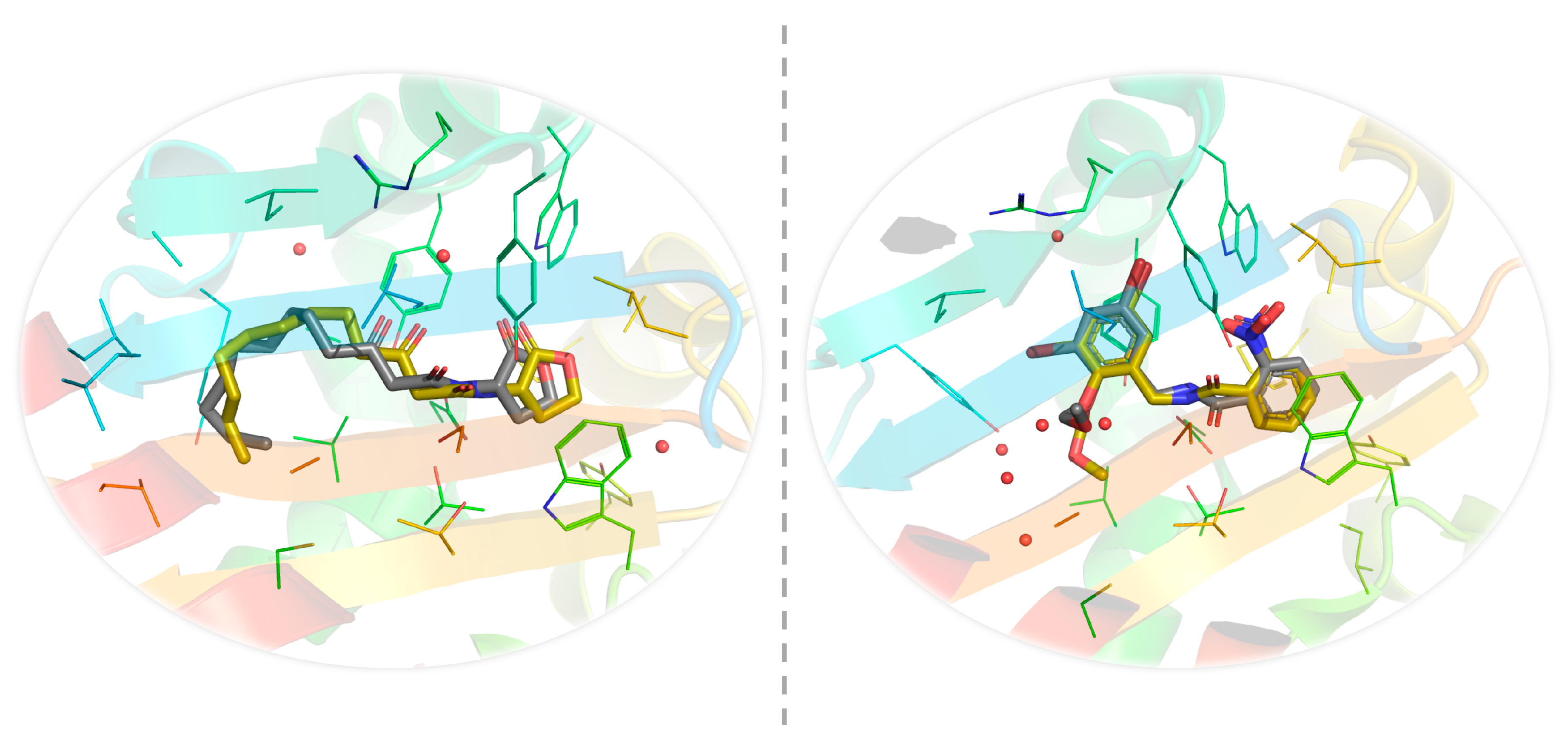
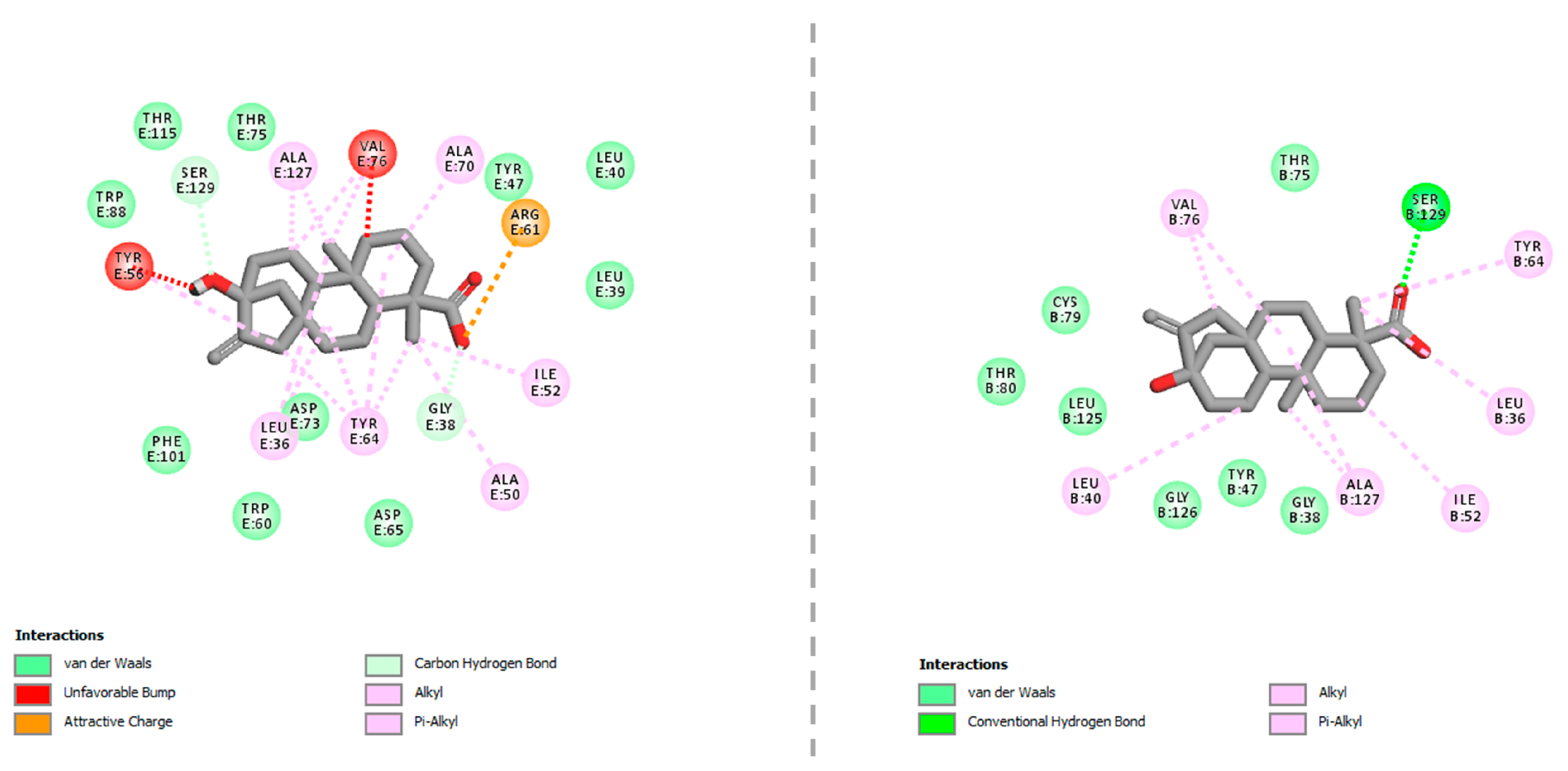
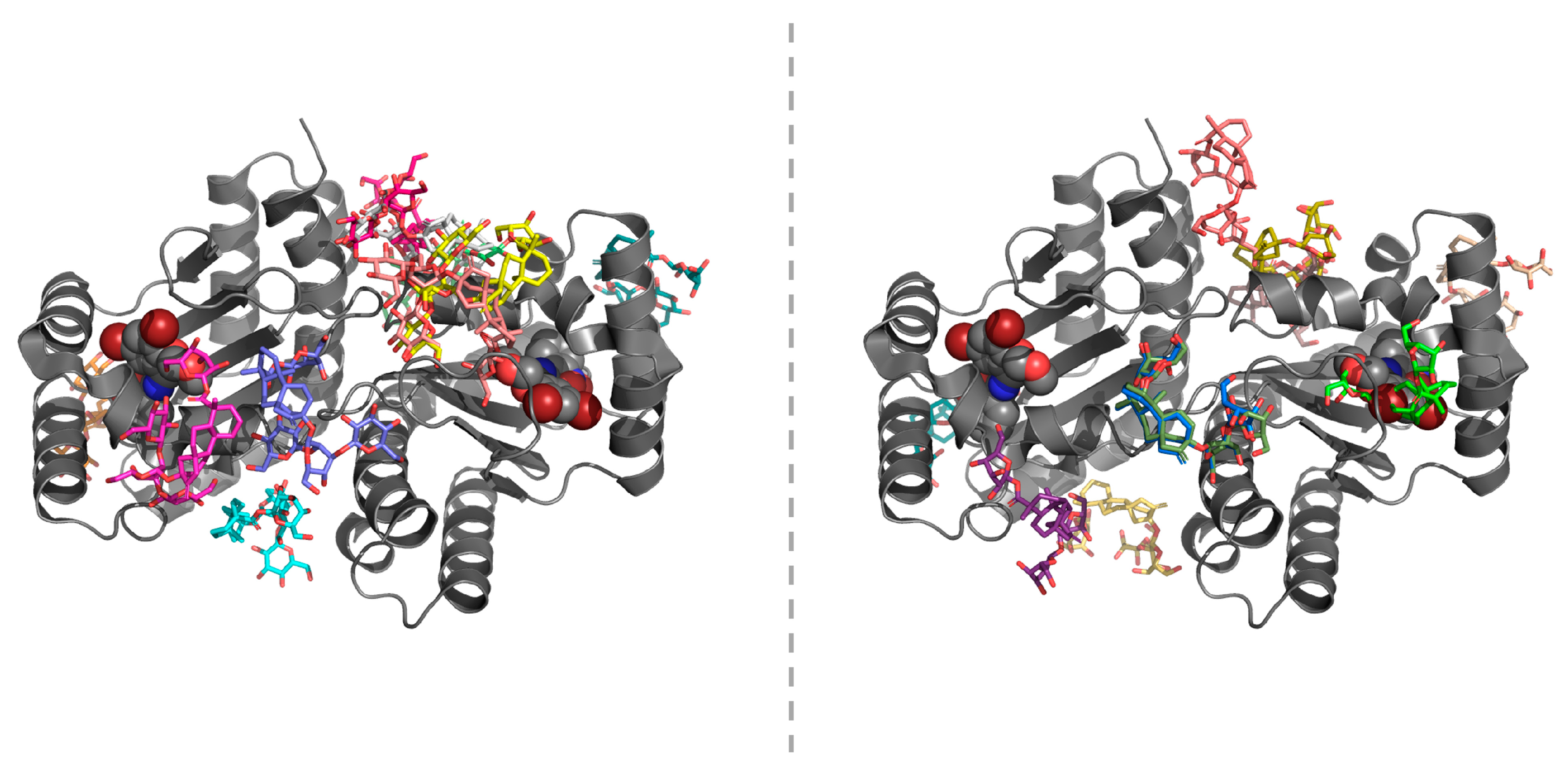
2.4. In-Silico Studies
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Bacterial Strains
4.3. Strain Cultivation
4.4. Effects of Stevia Extracts and Stevia-Derived Compounds on Bacterial QS
4.5. LC-MS Analysis of CSHS
4.6. Molecular Docking
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Samuel, P.; Ayoob, T.K.; Magnuson, A.B.; Wölwer-Rieck, U.; Jeppesen, B.P.; Rogers, J.P.; Rowland, I.; Mathews, R. Stevia Leaf to Stevia Sweetener: Exploring ItsScience, Benefits, and Future Potential. J. Nutr. 2018, 148, 1186S–1205S. [Google Scholar] [CrossRef]
- Brandle, J.E.; Starratt, A.; Gijzen, M. Stevia rebaudiana: Its agricultural, biological, and chemical properties. Can. J. Plant Sci. 1998, 78, 527–536. [Google Scholar] [CrossRef]
- Hutapea, A.M.; Toskulkao, C.; Buddhasukh, D.; Wilairat, P.; Glinsukon, T. Digestion of Stevioside, a Natural Sweetener, by Various Digestive Enzymes. J. Clin. Biochem. Nutr. 1997, 23, 177–186. [Google Scholar] [CrossRef]
- Gardana, C.; Simonetti, P.; Canzi, E.; Zanchi, R.; Pietta, P. Metabolism of Stevioside and Rebaudioside A from Stevia rebaudiana Extracts by Human Microflora. J. Agric. Food Chem. 2003, 51, 6618–6622. [Google Scholar] [CrossRef]
- Nikiforov, A.I.; Rihner, M.O.; Eapen, A.K.; Thomas, J.A. Metabolism and Toxicity Studies Supporting the Safety of Rebaudioside D. Int. J. Toxicol. 2013, 32, 261–273. [Google Scholar] [CrossRef]
- Purkayastha, S.; Markosyan, A.; Prakash, I.; Bhusari, S.; Pugh, G.; Lynch, B.; Roberts, A. Steviol glycosides in purified stevia leaf extract sharing the same metabolic fate. Regul. Toxicol. Pharmacol. 2016, 77, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Chen, S.C.; Chan, P.; Chu, Y.L.; Yang, H.Y.; Cheng, J.T. Mechanism of the hypoglycemic effect of stevioside, a glycoside of Stevia rebaudiana. Planta Med. 2005, 71, 108–113. [Google Scholar] [CrossRef]
- Ilić, V.; Vukmirović, S.; Stilinović, N.; Čapo, I.; Arsenović, M.; Milijašević, B. Insight into anti-diabetic effect of low dose of stevioside. Biomed. Pharmacother. 2017, 90, 216–221. [Google Scholar] [CrossRef]
- Ripken, D.; van der Wielen, N.; Wortelboer, H.M.; Meijerink, J.; Witkamp, R.F.; Hendriks, H.F.J. Steviol Glycoside Rebaudioside A Induces Glucagon-like Peptide-1 and Peptide YY Release in a Porcine ex Vivo Intestinal Model. J. Agric. Food Chem. 2014, 62, 8365–8370. [Google Scholar] [CrossRef]
- van der Wielen, N.; ten Klooster, J.P.; Muckenschnabl, S.; Pieters, R.; Hendriks, H.F.J.; Witkamp, R.F.; Meijerink, J. The Noncaloric Sweetener Rebaudioside A Stimulates Glucagon-Like Peptide 1 Release and Increases Enteroendocrine Cell Numbers in 2-Dimensional Mouse Organoids Derived from Different Locations of the Intestine. J. Nutr. 2016, 146, 2429–2435. [Google Scholar] [CrossRef]
- Jeppesen, P.B.; Gregersen, S.; Poulsen, C.R. Stevioside acts directly on pancreatic beta cells to secrete insulin: Actions independent of cyclic adenosine monophosphate and adenosine triphosphate-sensitive K+-channel activity. Metabolism 2000, 49, 208–214. [Google Scholar] [CrossRef]
- Philippaert, K.; Pironet, A.; Mesuere, M.; Sones, W.; Vermeiren, L.; Kerselaers, S.; Pinto, S.; Segal, A.; Antoine, N.; Gysemans, C.; et al. Steviol glycosides enhance pancreatic beta-cell function and taste sensation by potentiation of TRPM5 channel activity. Nat. Commun. 2017, 8, 14733. [Google Scholar] [CrossRef]
- Becker, S.L.; Chiang, E.; Plantinga, A.; Carey, H.V.; Suen, G.; Swoap, S.J. Effect of stevia on the gut microbiota and glucose tolerance in a murine model of diet-induced obesity. FEMS Microbiol. Ecol. 2020, 96, fiaa079. [Google Scholar] [CrossRef] [PubMed]
- Nettleton, J.E.; Cho, N.A.; Klancic, T.; Nicolucci, A.C.; Shearer, J.; Borgland, S.L.; Johnston, L.A.; Ramay, R.H.; Tuplin, E.N.; Chleilat, F.; et al. Maternal low-dose aspartame and stevia consumption with an obesogenic diet alters metabolism, gut microbiota and mesolimbic reward system in rat dams and their offspring. Gut 2020, 69, 1807–1817. [Google Scholar] [CrossRef] [PubMed]
- Nettleton, J.E.; Klancic, T.; Schick, A.; Choo, A.C.; Shearer, J.; Borgland, S.L.; Chleilat, F.; Mayengbam, S.; Reimer, R.A. Low-Dose Stevia (Rebaudioside A) Consumption Perturbs Gut Microbiota and the Mesolimbic Dopamine Reward System. Nutrients 2019, 11, 1248. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, T.; Dong, S.; Xiong, Y.; Wei, H.; Xu, F. The Effects of Rebaudioside A on Microbial Diversity in Mouse Intestine. Food Sci. Technol. Res. 2014, 20, 459–467. [Google Scholar] [CrossRef]
- Mahalak, K.; Firrman, J.; Tomasula, P.M.; Nunez, A.; Lee, J.-J.; Bittinger, K.; Rinaldi, M.; Liu, L. The impact of steviol glycosides and erythritol on the human and Cebus apella gut microbiome. J. Agric. Food Chem. 2020, 68, 13093–13101. [Google Scholar] [CrossRef]
- Jimenez, A.G.; Sperandio, V. Quorum Sensing and the Gut Microbiome. In Quorum Sensing: Molecular Mechanism and Biotechnological Application; Tommonaro, G., Ed.; Academic Press: Amsterdam, The Netherlands, 2019; pp. 151–169. [Google Scholar]
- Atkinson, S.; Williams, P. Quorum sensing and social networking in the microbial world. J. R. Soc. Interface 2009, 6, 959–978. [Google Scholar] [CrossRef]
- Mukherjee, S.; Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Azimi, S.; Klementiev, A.D.; Whiteley, M.; Diggle, S.P. Bacterial Quorum Sensing During Infection. Annu. Rev. Microbiol. 2020, 74, 201–219. [Google Scholar] [CrossRef]
- Wang, S.; Payne, G.F.; Bentley, W.E. Quorum Sensing Communication: Molecularly Connecting Cells, Their Neighbors, and Even Devices. Annu. Rev. Chem. Biomol. Eng. 2020, 11, 447–468. [Google Scholar] [CrossRef] [PubMed]
- Bassler, L.B.; Greenberg, P.E.; Stevens, M.A. Cross-Species Induction of Luminescence in the Quorum-Sensing Bacterium Vibrio harveyi. J. Bacteriol. 1997, 179, 4043–4045. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Schauder, S.; Potier, N.; Van Dorsselaer, A.; Pelczer, I.; Bassler, B.L.; Hughson, F.M. Structural identification of a bacterial quorum-sensing signal containing boron. Nature 2002, 415, 545–549. [Google Scholar] [CrossRef]
- Xavier, K.B.; Bassler, B.L. Interference with AI-2-mediated bacterial cell–cell communication. Nature 2005, 437, 750–753. [Google Scholar] [CrossRef] [PubMed]
- An, J.H.; Goo, E.; Kim, H.; Seo, Y.-S.; Hwang, I. Bacterial quorum sensing and metabolic slowing in a cooperative population. Proc. Natl. Acad. Sci. USA 2014, 111, 14912–14917. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, X.; Xu, X.; Fu, V.Y. Microbe social skill: The cell-to-cell communication between microorganisms. Sci. Bull. 2017, 62, 516–524. [Google Scholar] [CrossRef]
- Lazazzera, B.A. The intracellular function of extracellular signaling peptides. Peptides 2001, 22, 1519–1527. [Google Scholar] [CrossRef]
- Okada, M.; Sato, I.; Cho, S.J.; Iwata, H.; Nishio, T.; Dubnau, D.; Sakagami, Y. Structure of the Bacillus subtilis quorum-sensing peptide pheromone ComX. Nat. Chem. Biol. 2005, 1, 23–24. [Google Scholar] [CrossRef]
- Thoendel, M.; Kavanaugh, J.S.; Flack, C.E.; Horswill, A.R. Peptide Signaling in the Staphylococci. Chem. Rev. 2011, 111, 117–151. [Google Scholar] [CrossRef]
- Whitehead, N.A.; Barnard, A.M.L.; Slater, H.; Simpson, N.J.L.; Salmond, G.P.C. Quorum-sensing in Gram-negative bacteria. FEMS Microbiol. Rev. 2001, 25, 365–404. [Google Scholar] [CrossRef]
- Dobretsov, S.; Teplitski, M.; Paul, V. Mini review: Quorum sensing in the marine environment and its relationship to biofouling. Biofouling 2009, 25, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.-H.; Wang, L.-H.; Xu, J.-L.; Zhang, H.-B.; Zhang, X.-F.; Zhang, L.-H. Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature 2001, 411, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Sitnikov, D.M.; Schineller, J.B.; Baldwin, T.O. Transcriptional regulation of bioluminesence genes from Vibrio fischeri. Mol. Microbiol. 1995, 17, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed]
- Landman, C.; Besse, A.; Maubert, M.; Brot, L.; Humbert, L.; Cosnes, J.; Beaugerie, L.; Trugnan, G.; Sokol, H.; Rainteau, D.; et al. Quorum Sensing Driven by N-Acyl-Homoserine Lactone in Inflammatory Bowel Diseases Associated Dysbiosis. Gastroenterology 2013, 144, S310. [Google Scholar] [CrossRef]
- Le Balc’h, E.; Landman, C.; Tauziet, E.; Brot, L.; Quevrain, E.; Rainteau, D.; Grill, J.-P.; Thenet, S.; Seksik, P. 3-oxo-C12:2-HSL, a new N-acyl-homoserine lactone identified in gut ecosystem exerts an anti-inflammatory effect and does not modify paracellular permeability. J. Crohns Colitis 2017, 11, S479–S480. [Google Scholar] [CrossRef]
- Landman, C.; Grill, J.-P.; Mallet, J.-M.; Marteau, P.; Humbert, L.; Le Balc’h, E.; Maubert, M.-A.; Perez, K.; Chaara, W.; Brot, L.; et al. Inter-kingdom effect on epithelial cells of the N-Acyl homoserine lactone 3-oxo-C12:2, a major quorum-sensing molecule from gut microbiota. PLoS ONE 2018, 13, e0202587. [Google Scholar] [CrossRef]
- Kim, C.S.; Gatsios, A.; Cuesta, S.; Lam, Y.C.; Wei, Z.; Chen, H.; Russell, R.M.; Shine, E.E.; Wang, R.; Wyche, T.P.; et al. Characterization of Autoinducer-3 Structure and Biosynthesis in E. coli. ACS Cent. Sci. 2020, 6, 197–206. [Google Scholar] [CrossRef]
- Goh, E.B.; Yim, G.; Tsui, W.; McClure, J.; Surette, M.G.; Davies, J. Transcriptional modulation of bacterial gene expression by sub-inhibitory concentrations of antibiotics. Proc. Natl. Acad. Sci. USA 2002, 99, 17025–17030. [Google Scholar] [CrossRef]
- Steckel, A.; Schlosser, G. An Organic Chemist’s Guide to Electrospray Mass Spectrometric Structure Elucidation. Molecules 2019, 24, 611. [Google Scholar] [CrossRef]
- Bukelman, O.; Amara, N.; Mashiach, R.; Krief, P.; Meijler, M.M.; Alfonta, L. Electrochemical Studies of Biofilm Formation and Inhibition. Chem. Commun. 2009, 2836–2838. [Google Scholar] [CrossRef]
- Duan, K.; Surette, M.G. Environmental Regulation of Pseudomonas aeruginosa PAO1 Las and Rhl Quorum-Sensing Systems. J. Bacteriol. 2007, 4827–4836. [Google Scholar] [CrossRef]
- O’Reilly, M.C.; Dong, S.H.; Rossi, F.M.; Karlen, K.M.; Kumar, R.S.; Nair, S.K.; Blackwell, H.E. Structural and Biochemical Studies of Non-native Agonists of the LasR Quorum-Sensing Receptor Reveal an L3 Loop “Out” Conformation for LasR. Cell Chem. Biol. 2018, 25, 1128–1139. [Google Scholar] [CrossRef]
- Bottomley, M.J.; Muraglia, E.; Bazzo, R.; Carfì, A. Molecular insights into quorum sensing in the human pathogen Pseudomonas aeruginosa from the structure of the virulence regulator LasR bound to its autoinducer. J. Biol. Chem. 2007, 282, 13592–13600. [Google Scholar] [CrossRef]
- Bursulaya, B.D.; Totrov, M.; Abagyan, R.; Brooks, C.L., III. Comparative study of several algorithms for flexible ligand docking. J. Comput. Aided Mol. Des. 2003, 17, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Deniņa, I.; Semjonovs, P.; Fomina, A.; Treimane, R.; Linde, R. The influence of stevia glycosides on the growth of Lactobacillus reuteri strains. Lett. Appl. Microbiol. 2013, 58, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Karimi, S.; Jonsson, H.; Lundh, T.; Roos, S. Lactobacillus reuteri strains protect epithelial barrier integrity of IPEC-J2 monolayers from the detrimental effect of enterotoxigenic Escherichia coli. Physiol. Rep. 2018, 6, e13514. [Google Scholar] [CrossRef]
- Wang, Q.-P.; Browman, D.; Herzog, H.; Neely, G.G. Non-nutritive sweeteners possess a bacteriostatic effect and alter gut microbiota in mice. PLoS ONE 2018, 13, e0199080. [Google Scholar] [CrossRef]
- Joint FAO/WHO expert committee on food additives. In Proceedings of the Eighty-Seventh Meeting, Rome, Italy, 4–13 June 2019; Available online: http://www.fao.org/3/ca5270en/ca5270en.pdf (accessed on 21 March 2020).
- Manefield, M.; Rasmussen, T.B.; Henzter, M.; Andersen, J.B.; Steinberg, P.; Kjelleberg, S.; Givskov, M. Halogenated furanones inhibit quorum sensing through accelerated LuxR turnover. Microbiology 2002, 148, 1119–1127. [Google Scholar] [CrossRef]
- Koch, B.; Liljefors, T.; Persson, T.; Nielsen, J.; Kjelleberg, S.; Givskov, M. The LuxR receptor: The sites of interaction with quorum-sensing signals and inhibitors. Microbiology 2005, 151, 3589–3602. [Google Scholar] [CrossRef]
- Moore, J.D.; Rossi, F.M.; Welsh, M.A.; Nyffeler, K.E.; Blackwell, H.E. A Comparative Analysis of Synthetic Quorum Sensing Modulators in Pseudomonas aeruginosa: New Insights into Mechanism, Active Efflux Susceptibility, Phenotypic Response, and Next-Generation Ligand Design. J. Am. Chem. Soc. 2015, 137, 14626–14639. [Google Scholar] [CrossRef] [PubMed]
- Paczkowski, J.E.; Mukherjee, S.; McCready, A.R.; Cong, J.P.; Aquino, C.J.; Kim, H.; Henke, B.R.; Smith, C.D.; Bassler, B.L. Flavonoids Suppress Pseudomonas aeruginosa Virulence through Allosteric Inhibition of Quorum-sensing Receptors. J Biol. Chem. 2017, 292, 4064–4076. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Swem, L.R.; Swem, D.L.; Stauff, D.L.; O’Loughlin, C.T.; Jeffrey, P.D.; Bassler, B.L.; Hughson, F.M. A strategy for antagonizing quorum sensing. Mol. Cell 2011, 42, 199–209. [Google Scholar] [CrossRef]
- Pessi, G.; Haas, D. Transcriptional control of the hydrogen cyanide biosynthetic genes hcnABC by the anaerobic regulator ANR and the quorum-sensing regulators LasR and RhlR in Pseudomonas aeruginosa. J. Bacteriol. 2000, 182, 6940–6949. [Google Scholar] [CrossRef]
- Gilbert, K.B.; Kim, T.H.; Gupta, R.; Greenberg, E.P.; Schuster, M. Global position analysis of the Pseudomonas aeruginosa quorum-sensing transcription factor LasR. Mol. Microbiol. 2009, 73, 1072–1085. [Google Scholar] [CrossRef]
- Ley, R.; Turnbaugh, P.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Koliada, A.; Syzenko, G.; Moseiko, V.; Budovska, L.; Puchkov, K.; Perederiy, V.; Gavalko, Y.; Dorofeyev, A.; Romanenko, M.; Tkach, S.; et al. Association between body mass index and Firmicutes/Bacteroidetes ratio in an adult Ukrainian population. BMC Microbiol. 2017, 17, 120. [Google Scholar] [CrossRef]
- Riva, A.; Borgo, F.; Lassandro, C.; Verduci, E.; Morace, G.; Borghi, E.; Berry, D. Pediatric obesity is associated with an altered gut microbiota and discordant shifts in Firmicutes populations. Environ. Microbiol. 2017, 19, 95–105. [Google Scholar] [CrossRef]
- Lane, E.R.; Zisman, T.; Suskind, D. The microbiota in inflammatory bowel disease: Current and therapeutic insights. J. Inflamm. Res. 2017, 10, 63–73. [Google Scholar] [CrossRef]
- Landman, C.; Clement, M.; Nsiri, H.; Quevrain, E.; Bazin, T.; Brot, L.; Grill, J.-P.; Maubert, M.-A.; Humbert, L.; Sokol, H.; et al. The N- Acyl-Homoserine Lactone 3-oxo-C12, an Inter-Bacterial Signaling Molecule (Involved in Quorum Sensing), Exerts Effects on the Host: Thus, Implicating Quorum Sensing in Inflammatory Bowel Disease. J. Crohns Colitis 2015, 148, S717–S718. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. PubChem Substance and Compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Liu, Y.; Grimm, M.; Dai, W.T.; Hou, M.C.; Xiao, Z.X.; Cao, Y. CB-Dock: A web server for cavity detection-guided protein-ligand blind docking. Acta Pharmacol. Sin. 2020, 41, 138–144. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Closed-Loop Conformation (PDB Entry: 2UV0) | Open-Loop Conformation (PDB Entry: 6D6P) | |||
|---|---|---|---|---|
| Chain E | Chain G | Chain A | Chain B | |
| Steviol | −3.0 kcal mol−1 | −2.5 kcal mol−1 | −9.0 kcal mol−1 | −8.7 kcal mol−1 |
| 3-Oxo-C12-HSL | −8.9 kcal mol−1 | −8.9 kcal mol−1 | − | − |
| Compound 19 | − | − | −11.5 kcal mol−1 | −10.9 kcal mol−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markus, V.; Share, O.; Teralı, K.; Ozer, N.; Marks, R.S.; Kushmaro, A.; Golberg, K. Anti-Quorum Sensing Activity of Stevia Extract, Stevioside, Rebaudioside A and Their Aglycon Steviol. Molecules 2020, 25, 5480. https://doi.org/10.3390/molecules25225480
Markus V, Share O, Teralı K, Ozer N, Marks RS, Kushmaro A, Golberg K. Anti-Quorum Sensing Activity of Stevia Extract, Stevioside, Rebaudioside A and Their Aglycon Steviol. Molecules. 2020; 25(22):5480. https://doi.org/10.3390/molecules25225480
Chicago/Turabian StyleMarkus, Victor, Orr Share, Kerem Teralı, Nazmi Ozer, Robert S. Marks, Ariel Kushmaro, and Karina Golberg. 2020. "Anti-Quorum Sensing Activity of Stevia Extract, Stevioside, Rebaudioside A and Their Aglycon Steviol" Molecules 25, no. 22: 5480. https://doi.org/10.3390/molecules25225480
APA StyleMarkus, V., Share, O., Teralı, K., Ozer, N., Marks, R. S., Kushmaro, A., & Golberg, K. (2020). Anti-Quorum Sensing Activity of Stevia Extract, Stevioside, Rebaudioside A and Their Aglycon Steviol. Molecules, 25(22), 5480. https://doi.org/10.3390/molecules25225480