
Response of Bioactive Metabolite and Biosynthesis Related Genes to Methyl Jasmonate Elicitation in Codonopsis pilosula

Abstract
1. Introduction
2. Results
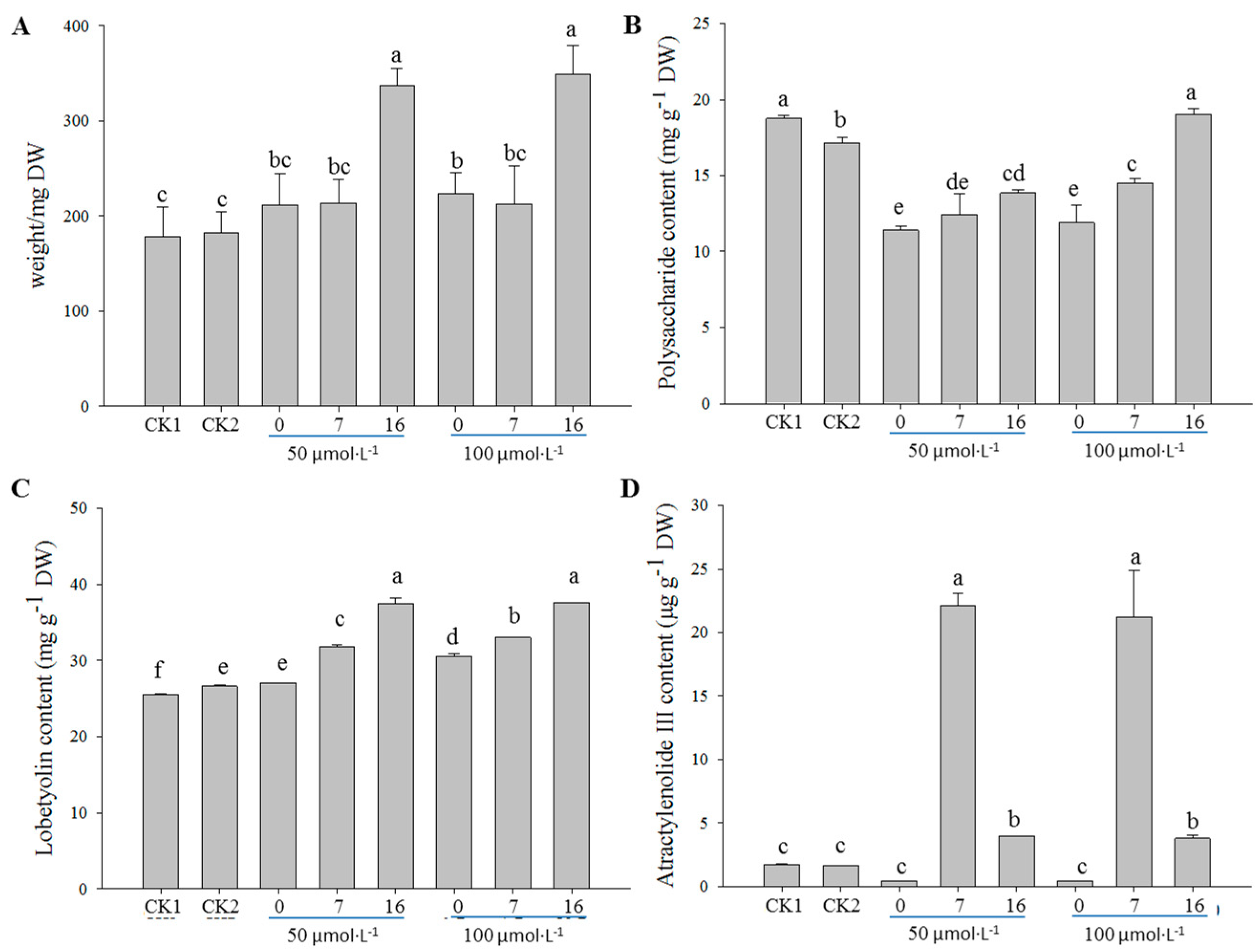
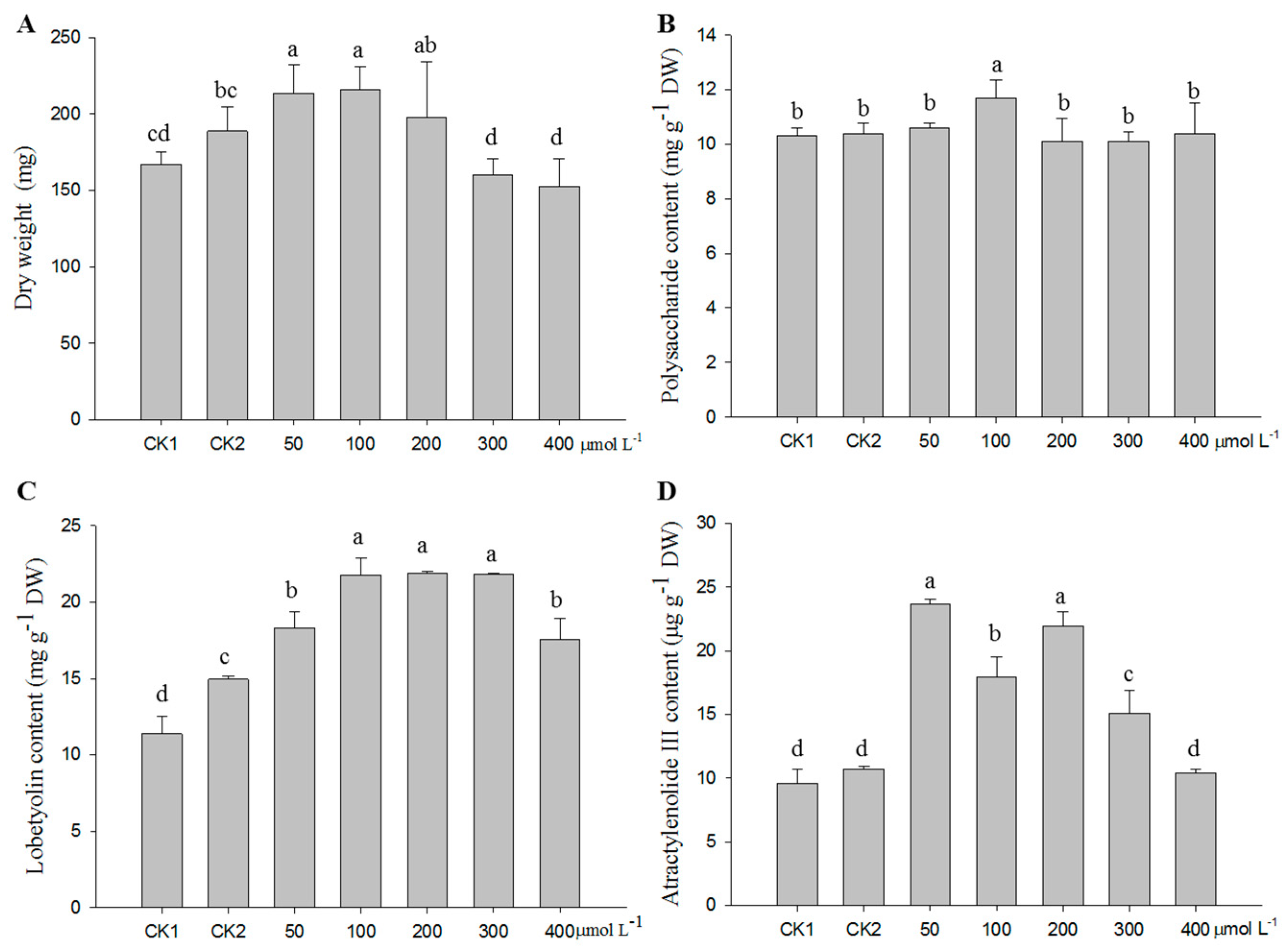
2.1. Optimization of the MeJA Treatment
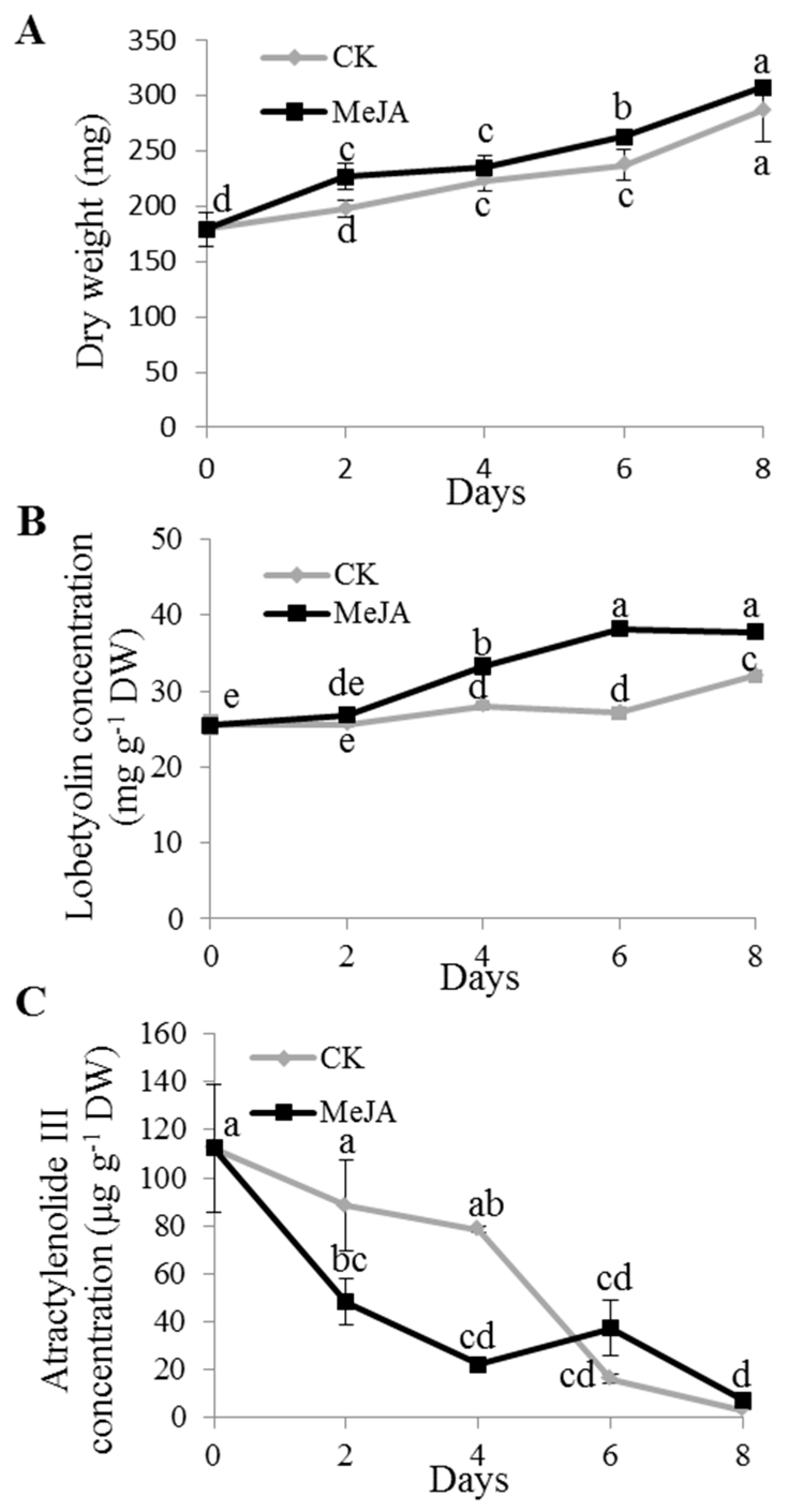
2.2. Dynamics of Lobetyolin and Atractylenolide III Accumulation under MeJA Treatment
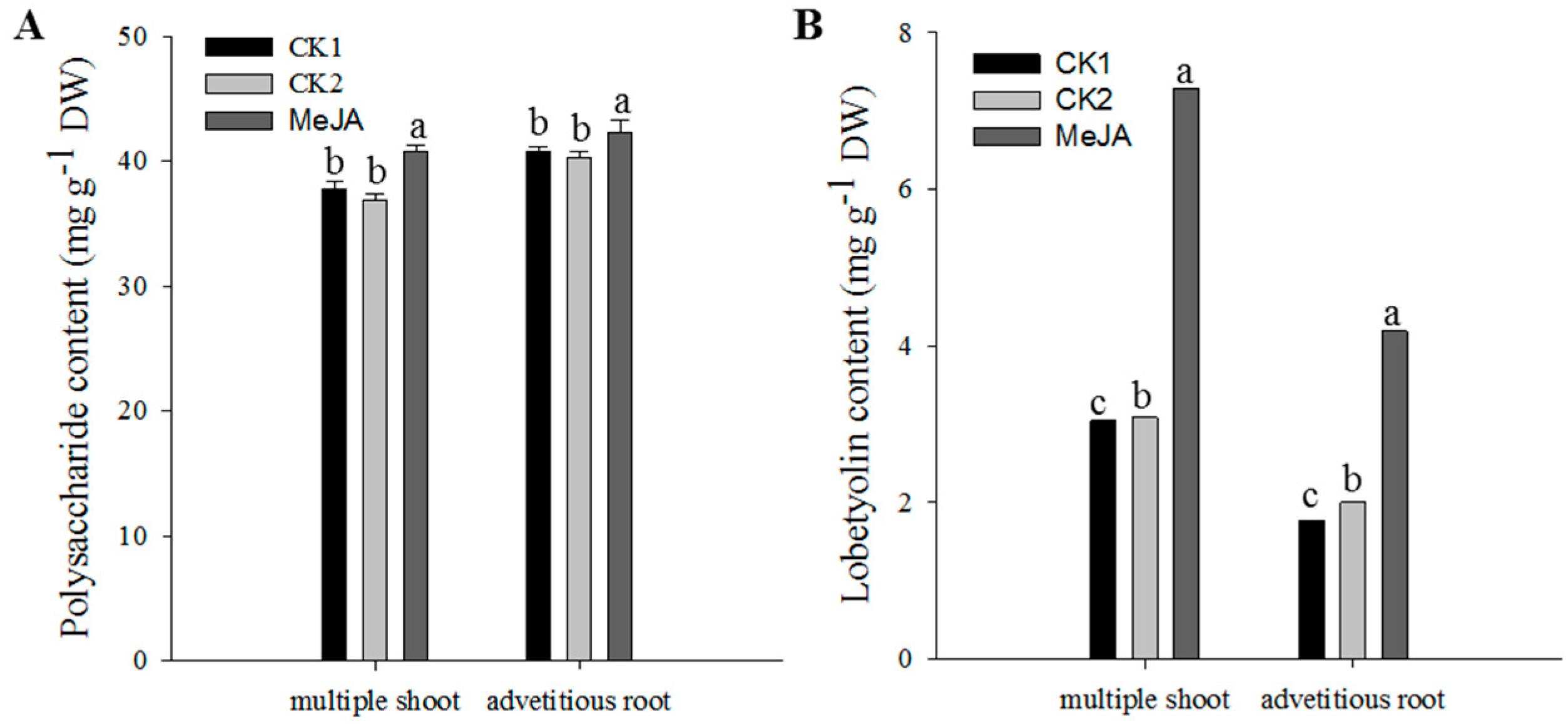
2.3. Effect of MeJA on Metabolite Accumulation in C. pilosula Multiple Shoots and Adventitious Roots
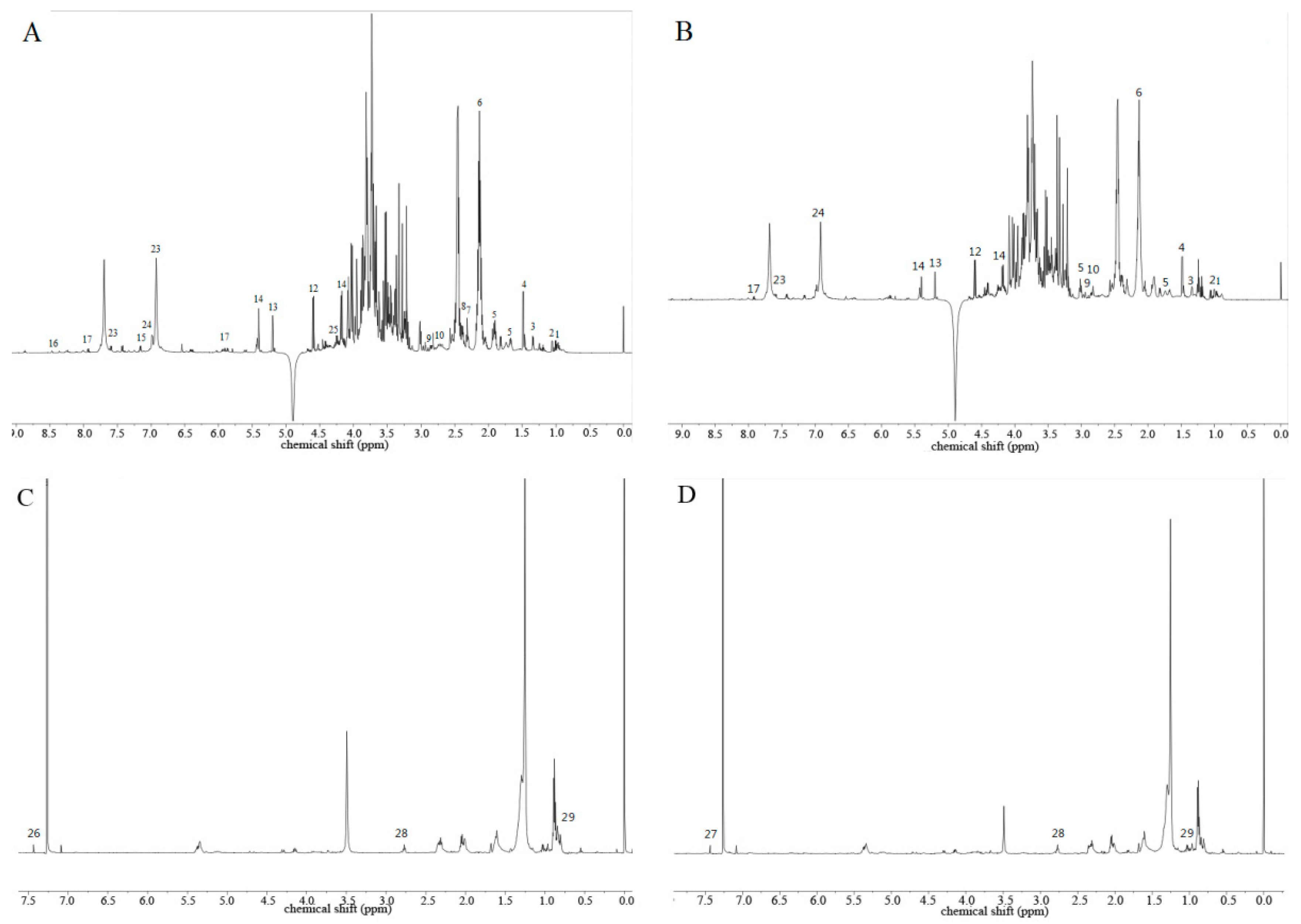
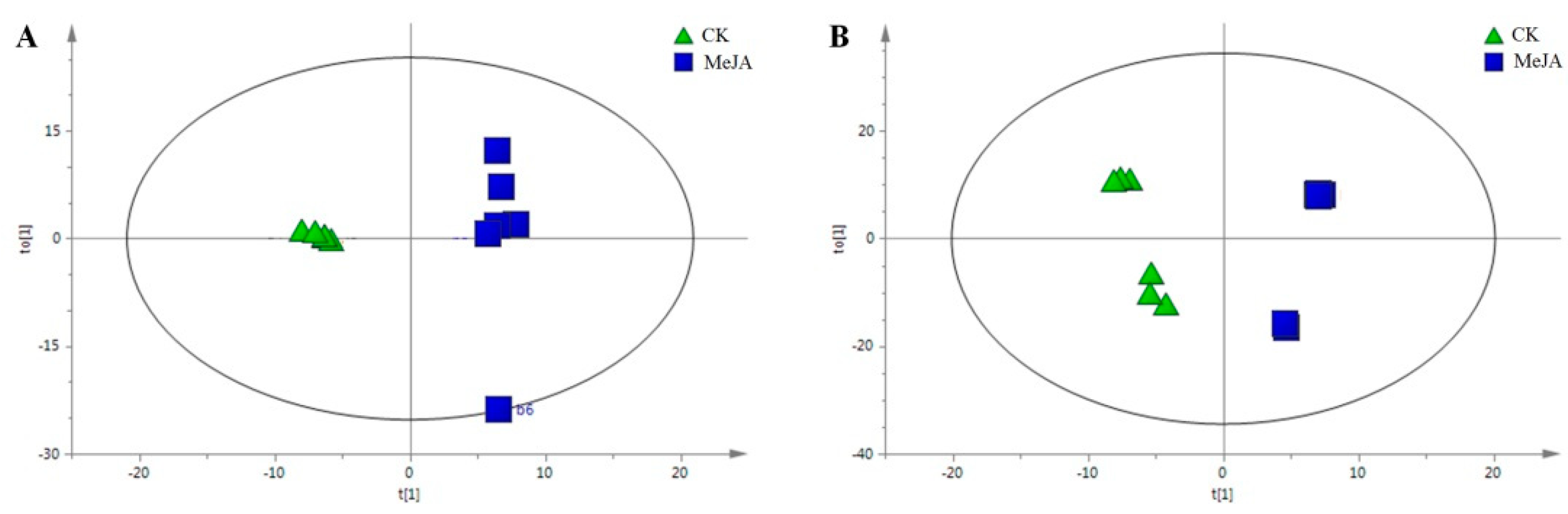
2.4. Metabolomic Profiling of C. pilosula Adventitious Root by 1H-NMR
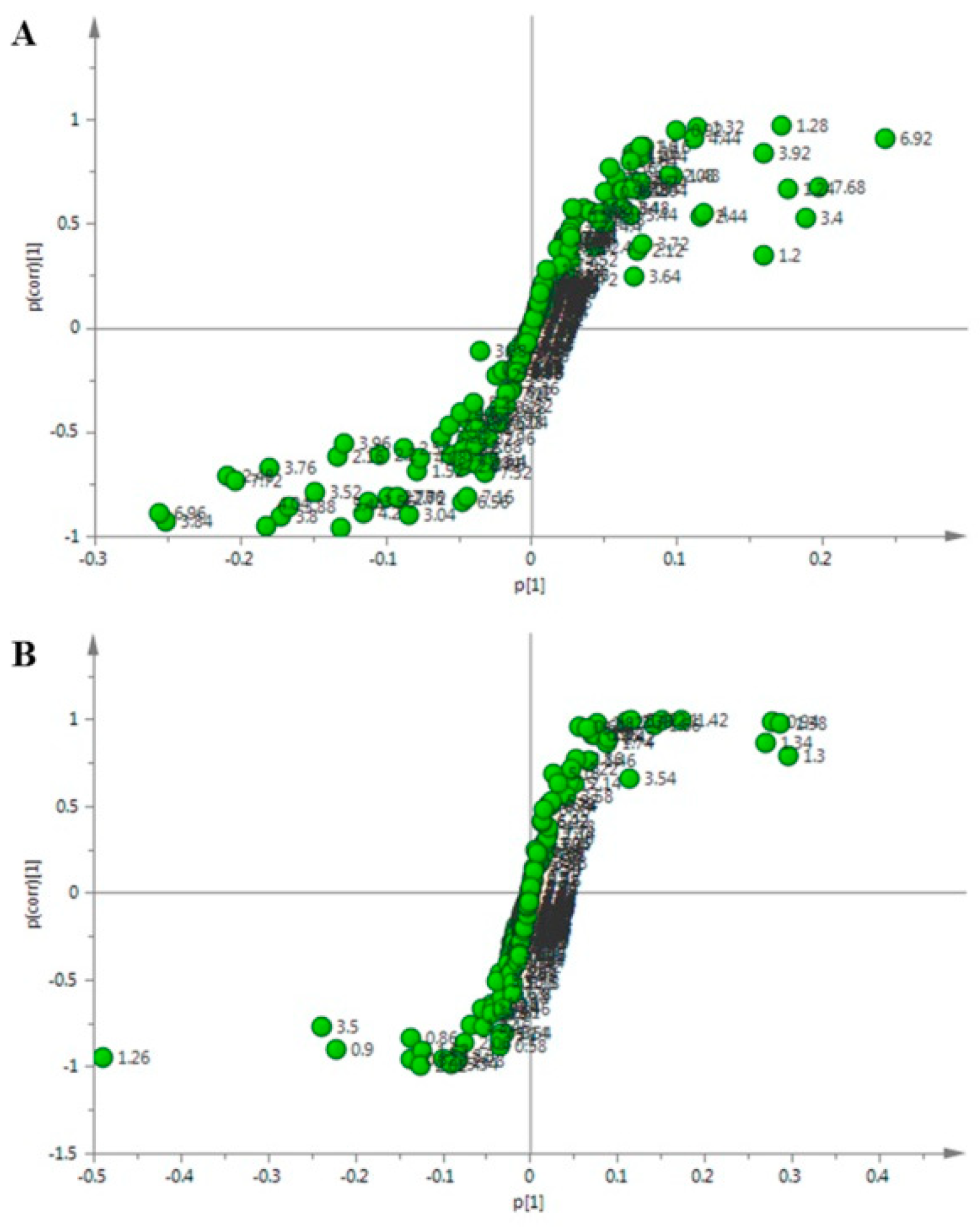
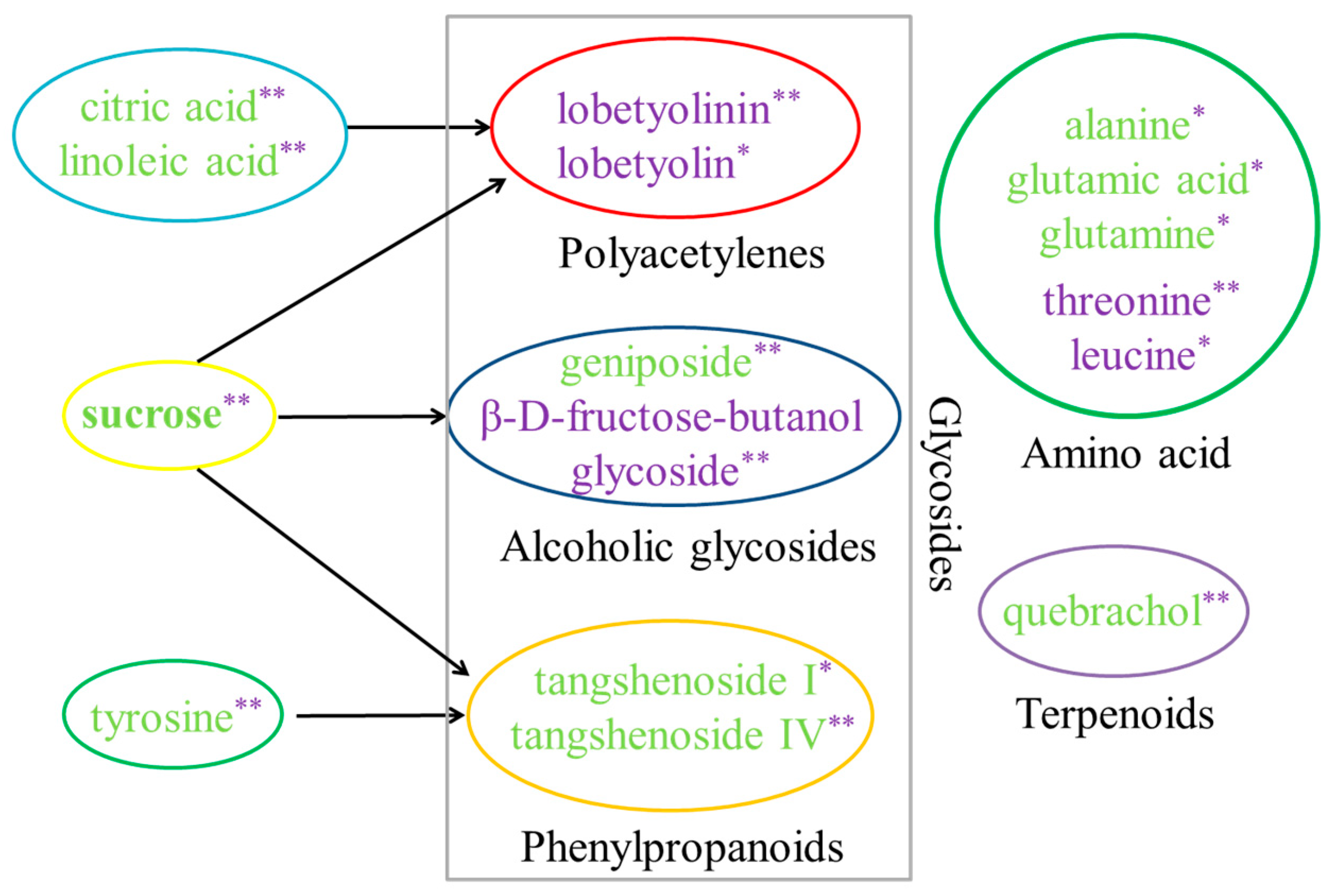
2.5. Identification of the Differential Metabolites in C. pilosula Adventitious Roots upon MeJA Treatment
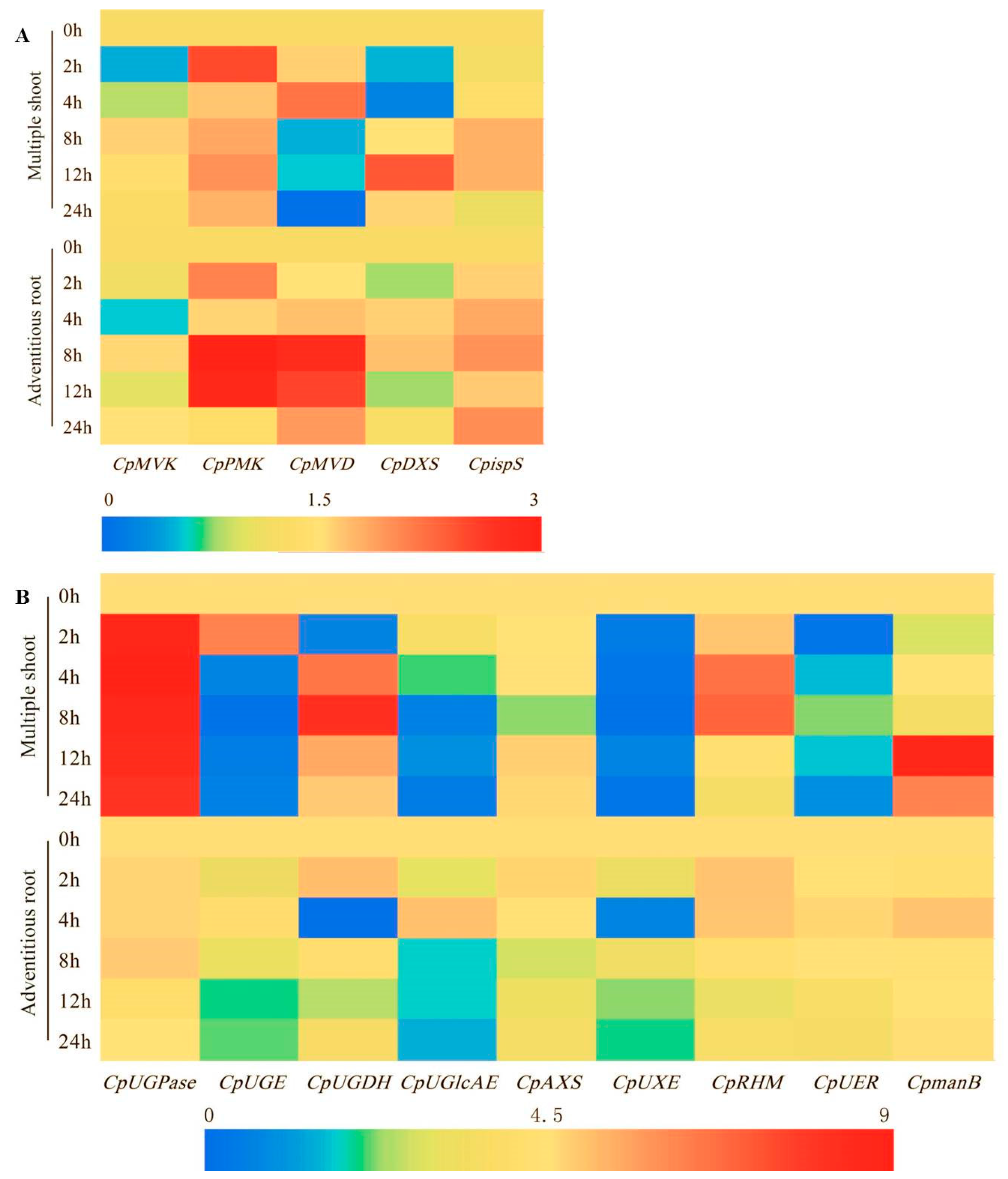
2.6. Effect of MeJA on the Expression of the Genes Involved in the Biosynthesis of the Main Bioactive Components in C. pilosula
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatment
4.2. Quantification of Bioactive Metabolites
4.3. 1H-NMR Analysis
4.4. Gene Expression Analysis
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MeJA | Methyl jasmonate |
| PLS-DA | Partial least square- discriminant analysis |
| OPLS-DA | Orthogonal partial least square-discriminant analysis |
| UGPase | UDP-glucose pyrophosphorylase |
| PMK | Mevalonate-5-phosphate kinase |
References
- Normile, D. The new face of traditional Chinese medicine. Science 2003, 299, 188–190. [Google Scholar] [CrossRef]
- Raskin, I.; Ribnicky, D.; Komarnytsky, S.; Llic, N.; Poulev, A.; Borisjuk, N.; Brinker, A.; Moreno, D.A.; Ripoll, C.; Yakoby, N.; et al. Plants and human health in the twenty-first century. Trends Biotechnol. 2002, 20, 522–531. [Google Scholar] [CrossRef]
- Hussain, M.; Sarfaraj, F.S.; Ansari, S.; Rahman, M.; Akhlaquer, A.I.Z.; Saeed, M. Current approaches toward production of secondary plant metabolites. J. Pharm. Bioallied. Sci. 2012, 4, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.Q.; Fang, X.; Wu, X.M.; Mao, Y.B.; Wang, L.J.; Chen, X.Y. Transcriptional regulation of plant secondary metabolism. J. Integr. Plant Biol. 2012, 54, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.R.; Ravishankar, G.A. Plant cell cultures: chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar]
- Thakur, G.S.; Sharma, R.; Sanodiya, B.S.; Baghel, R.; Thakur, R.; Singh, B.N.; Savita, A.; Dubey, A.; Sikarwar, L.; Jaiswal, P.; et al. In vitro induction of tuber formation for the synthesis of secondary metabolites in Chlorophytum borivilianum Sant. et Fernand. Afr. J. Biotechnol. 2013, 12, 2900–2907. [Google Scholar]
- Kim, H.J.; Park, K.J.; Lim, J.H. Metabolomic analysis of phenolic compounds in buckwheat (Fagopyrum esculentum M.) sprouts treated with methyl jasmonate. J. Agr. Food. Chem. 2011, 59, 5707–5713. [Google Scholar] [CrossRef]
- Singh, A.; Dwivedi, P. Methyl-jasmonate and salicylic acid as potent elicitors for secondary metabolite production in medicinal plants: A review. J. Pharmacogn. Phytochem. 2018, 7, 750–757. [Google Scholar]
- Zhong, J.J. Biochemical engineering of the production of plant-specific secondary metabolites by cell suspension cultures. Adv. Biochem. Eng. Biot. 2001, 72, 1–26. [Google Scholar]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Cheong, J.J.; Choi, Y.D. Methyl jasmonate as a vital substance in plants. Trends Genet. 2003, 19, 409–413. [Google Scholar] [CrossRef]
- Koo, A.J.K.; Howe, G.A. The wound hormone jasmonate. Phytochemistry 2009, 70, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.Y.; Kim, S.A.; Cho, K.S.; Yun, H.K. Expression of genes related to flavonoid and stilbene synthesis as affected by signalling chemicals and Botrytis cinerea in grapevines. Biol. Plant. 2014, 58, 758–767. [Google Scholar] [CrossRef]
- Onrubia, M.; Moyano, E.; Bonfill, M.; Cusido, R.M.; Goossens, A.; Palazón, J. Coronatine, a more powerful elicitor for inducing taxane biosynthesis in Taxus media cell cultures than methyl jasmonate. J. Plant Physiol. 2013, 170, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Ram, M.; Prasad, K.V.; Singh, S.K.; Hada, B.S.; Kumar, S. Influence of salicylic acid and methyl jasmonate elicitation on anthocyanin production in callus cultures of Rosa hybrid L. Plant Cell Tiss. Org. 2013, 113, 459–467. [Google Scholar] [CrossRef]
- Cao, S.; Cai, Y.; Yang, Z.; Joyce, D.C.; Zheng, Y. Effect of MeJA treatment on polyamine, energy status and anthracnose rot of loquat fruit. Food Chem. 2014, 145, 86–89. [Google Scholar] [CrossRef]
- Dučaiová, Z.; Sajko, M.; Mihaličová, S.; Repčák, M. Dynamics of accumulation of coumarin-related compounds in leaves of Matricaria chamomilla, after methyl jasmonate elicitation. J. Plant Growth Regul. 2016, 79, 81–94. [Google Scholar] [CrossRef]
- Zhou, P.F.; Yang, J.Z.; Zhu, J.H.; He, S.J.; Zhang, W.J.; Yu, R.M.; Zi, J.C.; Song, L.Y.; Huang, X.S. Effects of β-cyclodextrin and methyl jasmonate on the production of vindoline, catharanthine, and ajmalicine in Catharanthus roseus cambial meristematic cell cultures. App. Microbiol. Biot. 2015, 99, 7035–7045. [Google Scholar] [CrossRef]
- Saeed, S.; Ali, H.; Khan, T.; Kayani, W.; Khan, M.A. Impacts of methyl jasmonate and phenyl acetic acid on biomass accumulation and antioxidant potential in adventitious roots of Ajuga bracteosa Wall ex Benth.; a high valued endangered medicinal plant. Physiol. Mol. Biol. Pla. 2017, 23, 229–237. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science Press: Beijing, China, 2010; Volume I, pp. 264–265. [Google Scholar]
- Ishida, S.; Okasaka, M.; Ramos, F.; Kashiwada, Y.; Takaishi, Y.; Kodzhimatov, O.K.; Ashurmetov, O. New alkaloid from the aerial parts of Codonopsis clematidea. J. Nat. Med. 2008, 62, 236–238. [Google Scholar] [CrossRef] [PubMed]
- He, J.Y.; Zhu, S.; Goda, Y.; Cai, S.Q.; Komatsu, K. Quality evaluation of medicinally-used Codonopsis species and Codonopsis Radix based on the contents of pyrrolidine alkaloids, phenylpropanoid and polyacetylenes. J. Nat. Med. 2014, 68, 326–339. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.P.; Wang, D.; Cao, L.Y.; Sun, H.F. Transcriptome sequencing of Codonopsis pilosula and identification of candidate genes involved in polysaccharide biosynthesis. PLos ONE 2015, 10, e0117342. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Liu, Y.; Guo, Q.; Xu, C.; Zhu, C.; Shi, J. Sesquiterpene glycosides from the roots of Codonopsis pilosula. Acta. Pharm. Sin. B. 2016, 6, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Wang, Z.T.; Li, L.Y.; Zhong, G.Y. Protective effect of lobetyolin on gastric mucosa of experimental gastric ulcer in rats. J. Emerg. Tradit. Chin. Med. 2008, 17, 963–964. [Google Scholar]
- Endo, K.; Taquchi, T.; Taquchi, F.; Hikino, H.; Yamahara, J.; Fujimura, H. Antiinflammatory prinviples of Atractylodes Rhizomes. Chem. Pharm. Bull. 1979, 27, 2954–2958. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.X. Immunological adjuvant effect of a water-soluble polysaccharide, CPP, from the roots of Codonopsis pilosula on the immune responses to ovalbumin in mice. Chem. Biodivers. 2009, 6, 890–896. [Google Scholar] [CrossRef]
- Li, J.; Wang, T.; Zhu, Z.; Yang, F.; Cao, L.; Gao, J. Structure features and anti-Gastric ulcer effects of inulin-type fructan CPP-αfrom the roots of Codonopsis pilosula Franch.) Nannf. Molecules 2017, 22, 2258. [Google Scholar] [CrossRef]
- Li, A.P.; Li, Z.Y.; Xing, J.; Zhang, L.Z.; Qin, X.M. Chemical characterization of different vinegars by NMR-based metabolomics approach. Food Sci. 2013, 34, 247–253. [Google Scholar]
- Tian, D.; Li, Z.Y.; Fan, S.C.; Jia, J.P.; Qin, X.M. NMR-based analysis of water soluble extracts of different Astragali Radix. Acta Pharm. Sin. 2014, 49, 89–94. [Google Scholar]
- He, Q.; Zhu, E.Y.; Wang, Z.T.; Yu, G.X.; Xu, L.S.; Hu, Z.B. Study on Chemical Constitutes of Codonopsis pilosula. Chin. J. Pharm. 2006, 41, 10–12. [Google Scholar]
- Han, G.R.; He, X.F.; Yang, J.H.; Tian, Z.Y. Study on Chemical Constitutes of Codonopsis pilosula. China J. Chin. Mater. Med. 1990, 15, 41–42. [Google Scholar]
- Yuda, M.; Ohtani, K.; Mizutani, K.; Kasai, R.; Tanaka, O.; Jia, M.R.; Ling, Y.R.; Pu, X.F.; Saruwatari, Y.L. Neolignan glycosides from roots of Codonopsis tangshen. Phytochemistry 1990, 29, 1989–1993. [Google Scholar] [CrossRef]
- Qi, H.Y.; Wang, R.; Liu, Y.; Shi, Y.P. Studies on the chemical constituents of Codonopsis pilosula. J. Chin. Med. Mater. 2011, 34, 546–548. [Google Scholar]
- Ren, Z.B.; Zhang, Z.X.; Zhou, W.L.; Wang, J.H.; Li, X. Study on Chemical Constitutes of the flowers of Trollius Ledebourii. Mod. Chin. Med. 2006, 8, 14–15. [Google Scholar]
- Zhu, E.Y.; He, Q.; Wang, Z.T.; Xu, L.S.; Xu, G.J. Study on Chemical Constitutes of Codonopsis pilosula. J. China Pharm. Univ. 2001, 32, 94–95. [Google Scholar]
- Guillén, M.D.; Ruiz, A. High resolution 1H nuclear magnetic resonance in the study of edible oils and fats. Trends Food Sci. Technol. 2001, 12, 328–338. [Google Scholar] [CrossRef]
- Harborne, J.B.; Turner, B.L. Plant Chemosystematics; Academic Press: London, UK, 1984. [Google Scholar]
- De Geyter, N.; Gholami, A.; Goormachtig, S.; Goossens, A. Transcriptional machineries in jasmonate-elicited plant secondary metabolism. Trends Plant Sci. 2012, 17, 349–359. [Google Scholar] [CrossRef]
- Czerpak, R.; Piotrowska, A.; Szulecka, K. Jasmonic acid affects changes in the growth and some components content in alga Chlorella vulgaris. Acta. Physiol. Plant. 2006, 28, 195–203. [Google Scholar] [CrossRef]
- Wang, R.; Xu, S.; Wang, N.; Xia, B.; Jiang, Y.; Wang, R. Transcriptome analysis of secondary metabolism pathway, transcription factors, and transporters in response to methyl jasmonate in Lycoris aurea. Front. Plant Sci. 2016, 7, 1971. [Google Scholar] [CrossRef]
- Namdeo, A.G. Plant cell elicitation for production of secondary metabolites: A review. Pharmacogn. Rev. 2007, 1, 69–79. [Google Scholar]
- Yang, X.; Zhao, Y.; Ruan, Y.; Yang, Y. Development and application of a capillary electrophoretic method for the composition analysis of a typical heteropolysaccharide from Codonopsis pilosula Nannf. Biol. Pharm. Bull. 2008, 31, 1860–1865. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.J.; Liu, D.; Hu, Z.B. Study on relationship between uridine diphosphate glucose pyrophosphorylase activity and polysaccharide content in hairy root of Astragalus membranaceus. Plant Physiol. J. 2000, 14, 53–55. [Google Scholar]
- Payyavula, R.S.; Tschaplinski, T.J.; Jawdy, S.S.; Sykes, R.W.; Tuskan, G.A.; Kalluri, U.C. Metabolic profiling reveals altered sugar and secondary metabolism in response to UGPase overexpression in Populus. BMC Plant Biol. 2014, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.C.; Tsai, T.H.; Kuo, C.L. Chemical constituents comparison of Codonopsis tangshen, Codonopsis pilosula var. modesta and Codonopsis pilosula. Nat. Prod. Res. 2013, 27, 1812–1815. [Google Scholar] [CrossRef] [PubMed]
- Aguiló-Aguayo, I.; Gangopadhyay, N.; Lyng, J.G.; Brunton, N.P. Impact of pulsed light on colour, carotenoid, polyacetylene and sugar content of carrot slices. Innov. Food Sci. Emerg. 2017, 42, 49–55. [Google Scholar] [CrossRef]
- Wang, N.; Yuan, C. Overview of natural polyacetylene compounds. For. Med. Pharm. 1990, 3, 129–132. [Google Scholar]
- Ali, M.; Abbasi, B.H.; Ali, G.S. Elicitation of antioxidant secondary metabolites with jasmonates and gibberellic acid in cell suspension cultures of Artemisia absinthium L. Plant Cell Tiss. Org. 2015, 120, 1099–1106. [Google Scholar] [CrossRef]
- Garcia, D.E.; Keasling, J.D. Kinetics of phosphomevalonate kinase from Saccharomyces cerevisiae. PLoS ONE 2014, 9, e87112. [Google Scholar] [CrossRef]
- Ye, L.; Liu, S.; Xie, F.; Zhao, L.; Wu, X. Enhanced production of polysaccharides and triterpenoids in Ganoderma lucidum fruit bodies on induction with signal transduction during the fruiting stage. PLoS ONE 2018, 13, e0196287. [Google Scholar] [CrossRef]
- Ren, A.; Qin, L.; Shi, L.; Dong, X.; Mu, D.S.; Li, Y.X.; Zhao, M.W. Methyl jasmonate induces ganoderic acid biosynthesis in the basidiomycetous fungus, Ganoderma lucidum. Bioresource Technol. 2010, 101, 6785–6790. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Li, X.X.; Ji, J.J.; Sun, H.F.; Gao, J.P. Study on induction of multiple shoot and culture of adventitious root in Codonopsis pilosula. J. Chin. Med. Mater. 2017, 10, 2270–2274. [Google Scholar]
- Yang, J.; Su, Q.; Liu, E.; Gao, J.P. Content of atractylenolide III and lobetyolin in Codonopsis Radix from different regions. J. Shanxi Med. Univ. 2010, 41, 698–702. [Google Scholar]
- Zhen, X.; Gao, J.P.; Cao, L.Y. Measurement of polysaccharide in Lu Dangshen. Chin. Arch. Tradit. Chin. Med. 2014, 3, 498–500. [Google Scholar]
- Cao, L.Y.; Li, X.X.; Wang, D.; Sun, H.F.; Gao, J.P. Validation of reliable reference genes for accurate normalization in RT-qPCR analysis of Codonopsis pilosula. Chin. Tradit. Herbal. Drug 2017, 9, 226–235. [Google Scholar] [CrossRef]
Sample Availability: Not available. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Number | Compound | δH (J in Hz) | Reference |
|---|---|---|---|
| 1 | leucine | δ 0.94 (t, 3.3) | [30] |
| 2 | valine | δ 1.06 (d, 7.0), 1.01 (d, 7.0) | [30] |
| 3 | threonine | δ 1.34 (d, 6.5) | [30] |
| 4 | alanine | δ 3.73(m), 1.49 (d, 7.2) | [30] |
| 5 | arginine | δ 1.92–1.89 (m), 1.70–1.65 (m) | [30] |
| 6 | glutamic acid | δ 2.38(m), 2.13 (m) | [30] |
| 7 | acetylglutamate | δ 2.32 (t, 7.2) | [30] |
| 8 | glutamine | δ 2.46 (m), 2.13 (m) | [30] |
| 9 | asparaginic acid | δ 2.84 (dd, 16.4, 6.5) | [31] |
| 10 | citric acid | δ 2.72 (d, 4.6) | [31] |
| 11 | choline | δ 3.22 (s) | [31] |
| 12 | β-glucose | δ 4.60 (d, 7.9) | [31] |
| 13 | α-glucose | δ 5.20 (d, 3.7) | [31] |
| 14 | sucrose | δ 5.41 (d, 3.8), 4.18 (d, 8.7) | [31] |
| 15 | tyrosine | δ 7.16 (d, 8.7) | [30] |
| 16 | formic acid | δ 8.47 (s) | [31] |
| 17 | uridine | δ 7.86 (d, 8.7), 5.89 (dd, 10.6, 4.3), 4.35 (d, 7.7), 4.14 (dd, 8.8, 6.2), 3.80 (dd, 6.3, 2.3) | [32] |
| 18 | tangshenoside I | δ 4.60 (d, 7.9), 3.75 (s), 2.82 (s), 2.46 (d, 7.5), 1.41 (s) | [33] |
| 19 | Tangshenoside IV | δ 6.93 (s), 3.83 (s), 3.80 (s), 3.72 (s), 2.62 (d, 3.2) | [34] |
| 20 | lobetyolinin | δ 4.43 (d, 7.3), 4.37 (d, 6.4), 3.74 (dd, 11.5, 5.2), 3.53 (t, 4.3) | [35] |
| 21 | lobetyolin | δ 4.35 (d, 7.7), 4.24 (d, 2.6), 4.18 (dd, 9.6, 3.8) | [35] |
| 22 | β-d-fructose-butanol glycoside | δ 4.06 (d, 2.5), 1.57–1.51 (m) | [32] |
| 23 | vanillic acid | δ 7.54 (d, 2.4), 6.85 (d, 8.0) | [36] |
| 24 | aurantiamarin I | δ 6.93 (s), 3.81 (s), 1.06 (d, 7.0) | [35] |
| 25 | geniposide | δ 7.51 (s), 5.80 (s),5.17 (d, 3.7), 4.20 (d, 3.3), 3.87 (d, 2.3) | [32] |
| 28 | augelicin | δ 6.37 (d, 7.1) | [37] |
| 29 | psoralen | δ 7.48 (s), 6.37 (d, 7.1) | [37] |
| 30 | linoleic fatty acid | δ 2.77 (t, 6.6), 1.30–1.20 (m) | [38] |
| 31 | β-quebrachol | δ 0.89 (t, 5.9), 0.87 (d, 7.0) | [35] |
| Gene | Protein | Primer (5′–3′) | Product (bp) |
|---|---|---|---|
| CpGAPDH | glyceraldehyde-3-phosphate dehydrogenase | F: TGCTTCGTTCAACATCATTC | 164 |
| R: CATAACTGGCTGCCTTCTCC | |||
| CpMVK | mevalonate kinase | F: GACACAAAAGTTGGGAGGAACAC | 126 |
| R: GGTAGCCAGTTCATTGCTGATAGA | |||
| CpPMK | phosphomevalonate kinase | F: CTGCCGTAGTTGCTGCTTTACTT | 88 |
| R: TTCGTGGCTGTTTCTTGGTG | |||
| CpMVD | Methylovalerate decarboxylase | F: CAAGATGCTGGCGTTCAGG | 92 |
| R: CCTTTGGTTTTCTGCGTTGG | |||
| CpDXS | 1-deoxyxylose-5-phosphate synthase | F: TTGGCATAGCCGAACAACA | 94 |
| R: TGGAGGAAAGACGAGTAAATAGCAC | |||
| CpispS | isoprene synthase | F: ACGAACACTGCATCAAAGAATCTC | 112 |
| R: TGAACCCCAAAACCATCTCC | |||
| CpUGPase | UDP-glucose pyrophosphorylase | F: TTTACCCTTGAGAACGACG | 192 |
| R: TCTGATGGCTATGTGACCC | |||
| CpUGE | UDP-glucose isomerase | F: CGGGGTACATCTGTGCTTG | 161 |
| R: ATGCCATACTTTGCCTTCC | |||
| CpUGDH | UDP-glucose dehydrogenase | F: GATGCTTATGAGGCGACGAA | 193 |
| R: GAGGCTTACCAATGGAGTAGACAAT | |||
| CpUGlcAE | UDP galaotcse 4-epimerase | F: CACGGGATTTTACCTACAT | 95 |
| R: CTTCTTCTTACCGCCTGAT | |||
| CpAXS | UDP-apiose/xylose synthase | F: GATAAAGGCGATGACGATA | 146 |
| R: AAGACGGTTCAAGAAGGTG | |||
| CpUXE | UDP xylose epiisomerase | F: TGTTGGCACAGGAAGAGGT | 97 |
| R: CCGACGAGGAAGGAAATCA | |||
| CpRHM | rhamnose synthase | F: TCGGCATTCGGACTCTAAG | 111 |
| R: TCCTGAAGGTTCTGGCATT | |||
| CpUER | UDP-4-keto-6-deoxyglucose isoreductase | F: CCAATCCCCGTAACTTCAT | 131 |
| R: TATTCCAGTCAGGTTCCTCTTT | |||
| CpmanB | mannase B | F: AACGCCAACTGAGACAAC | 86 |
| R: GCACTCTTACAGCACCGA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, J.-j.; Feng, Q.; Sun, H.-f.; Zhang, X.-j.; Li, X.-x.; Li, J.-k.; Gao, J.-p. Response of Bioactive Metabolite and Biosynthesis Related Genes to Methyl Jasmonate Elicitation in Codonopsis pilosula. Molecules 2019, 24, 533. https://doi.org/10.3390/molecules24030533
Ji J-j, Feng Q, Sun H-f, Zhang X-j, Li X-x, Li J-k, Gao J-p. Response of Bioactive Metabolite and Biosynthesis Related Genes to Methyl Jasmonate Elicitation in Codonopsis pilosula. Molecules. 2019; 24(3):533. https://doi.org/10.3390/molecules24030533
Chicago/Turabian StyleJi, Jiao-jiao, Qi Feng, Hai-feng Sun, Xue-jun Zhang, Xiao-xiao Li, Jian-kuan Li, and Jian-ping Gao. 2019. "Response of Bioactive Metabolite and Biosynthesis Related Genes to Methyl Jasmonate Elicitation in Codonopsis pilosula" Molecules 24, no. 3: 533. https://doi.org/10.3390/molecules24030533
APA StyleJi, J.-j., Feng, Q., Sun, H.-f., Zhang, X.-j., Li, X.-x., Li, J.-k., & Gao, J.-p. (2019). Response of Bioactive Metabolite and Biosynthesis Related Genes to Methyl Jasmonate Elicitation in Codonopsis pilosula. Molecules, 24(3), 533. https://doi.org/10.3390/molecules24030533