1. Introduction
Cordycepin (3′-deoxyadenosine), a cytotoxic nucleoside analogue found in
Cordyceps militaris, has attracted considerable attention due to its diverse therapeutic potentials associated with anticancer, antitumor, anti-inflammatory, antimicrobial and antiviral effects [
1,
2,
3]. Cordycepin serves as an important marker for the quality control of
C. militaris.
The biosynthetic mechanism of cordycepin in purine biosynthesis has attracted widespread interest in the past seven decades since the discovery of cordycepin in 1950 [
4]. Notably, based on a comparative genetic analysis of two distantly homogenous and cordycepin-producing fungi (
C. militaris and
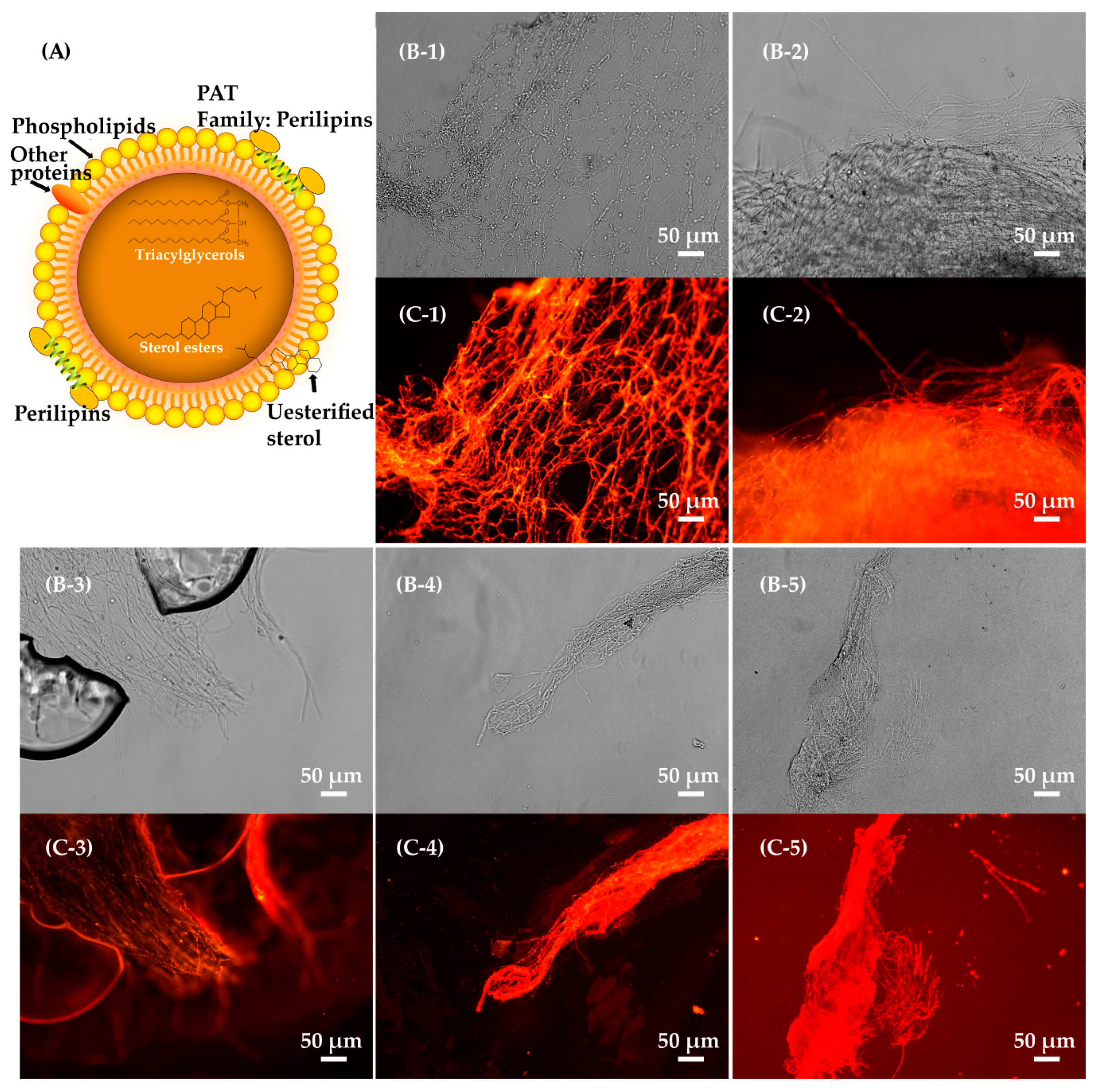
Aspergillus nidulans), the Cns1 and Cns2 proteins, responsible for cordycepin biosynthesis, were determined to be colocalized on a type of organelle in
Cordyceps militaris using GFP. Then, the Nile red method was employed to demonstrate that these organelles were lipid droplets in the mycelium of
C. militaris [
5]. To date, none of experiments have not been performed to investigate the effects of lipid droplets on Cns1 and Cns2 proteins in the mycelium of cordycepin-producing fungi. Previous work demonstrated that lipid droplets were involved in protein biosynthesis and degradation in eukaryotes [
6], suggesting that the effects of lipid droplets on the Cns1 and Cns2 proteins in the mycelium of cordycepin-producing fungi were still unclear. For example, whether lipid droplets can modify Cns1 and Cns2 proteins in the mycelium of cordycepin-producing fungi requires further investigated. In other words, the lipid droplets in cells may have potential effects (positive or negative) on cordycepin accumulation in the mycelium of cordycepin-producing fungi.
Lipid droplets, spherical cellular organelles consisting of a neutral lipid core enclosed by a phospholipid monolayer membrane covered with many multifunctional binding proteins, are involved in multiple intracellular processes, such as protein storage and degradation and nucleic acid processing [
6,
7]. The neutral lipid content in cells is associated with lipid droplet content. The compositions of neutral lipids in lipid droplets markedly vary and are associated with cell type and culture conditions. Major types of neutral lipids in lipid droplets are different from those in fungi and mammals, such as triacylglycerols and steryl esters in fungi (budding yeast) [
8], and triacylglycerol, cholesterol ester and ether lipids in in vivo mammalian cell lines [
9]. However, to date, the quantitative relationship between the neutral lipid content and cordycepin accumulation in the mycelia of cordycepin-producing fungi remains unclear.
The relative fluorescence intensity of Nile red-stained smooth muscle cells was first used as a reliable indicator to quantify neutral lipid content [
10], and the use of this proxy has been demonstrated in different microorganisms [
11,
12,
13]. The strain, the solvent in which Nile red is dissolved and the measurement conditions affect the combination of Nile red and neutral lipid, causing variable relative fluorescence intensities [
14]. Therefore, the measurement conditions for determining fluorescence intensity in a specific microorganism play a vital role in investigating the relationship between neutral lipid content and cordycepin accumulation in cordycepin-producing fungi.
In this study, fifteen mutant strains producing different levels of cordycepin were obtained from two C. militaris starting strains after LiCl+60Coγ compound mutagenesis. Lyophilized mycelium powder (LMP) was used as the original material to optimize the Nile red method to investigate the quantitative relationship between the cordycepin concentration in both liquid broth and LMP and the corresponding relative fluorescence intensity (RFI) from Nile red-stained LMP in cordycepin-producing fungi.
3. Discussion
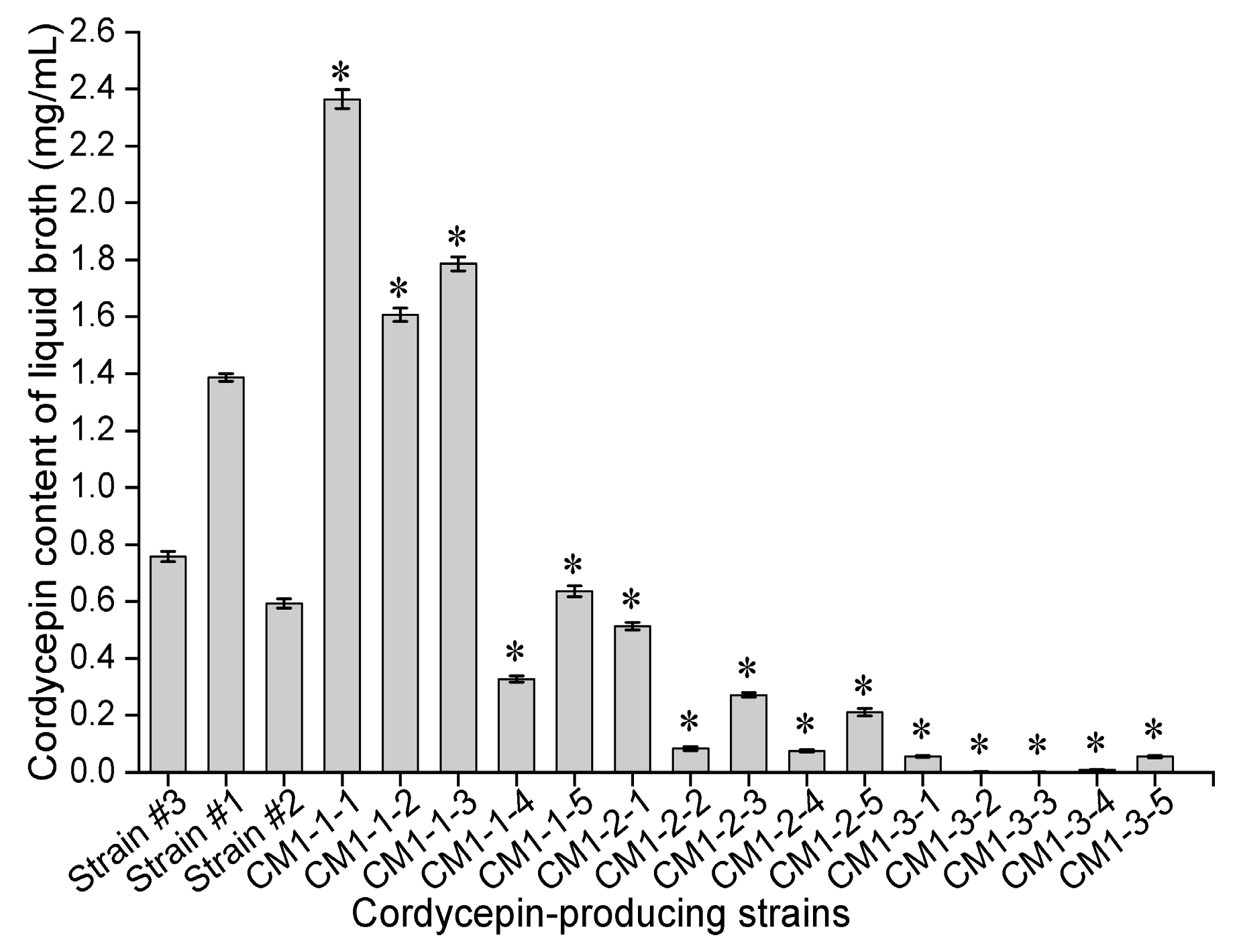
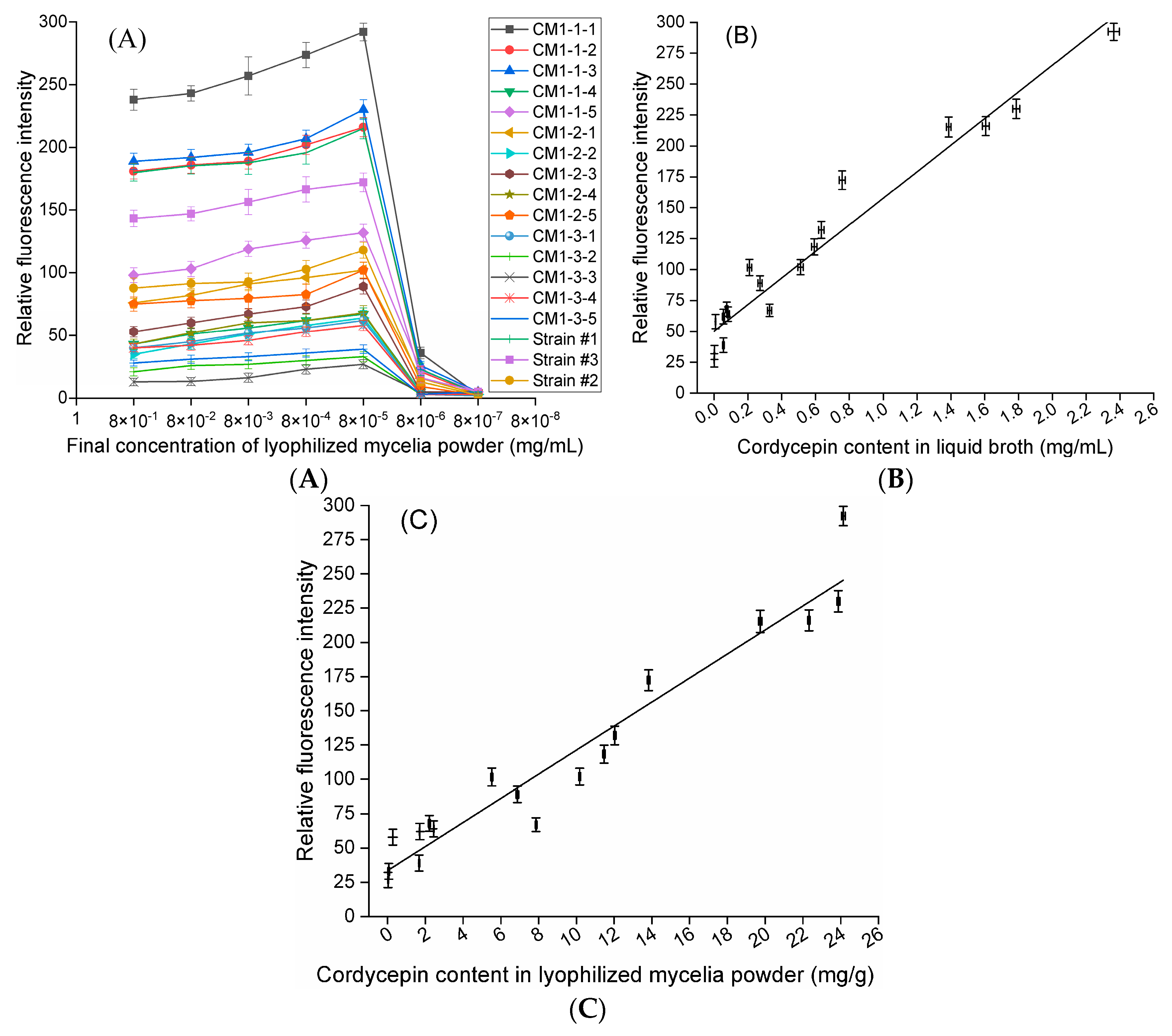
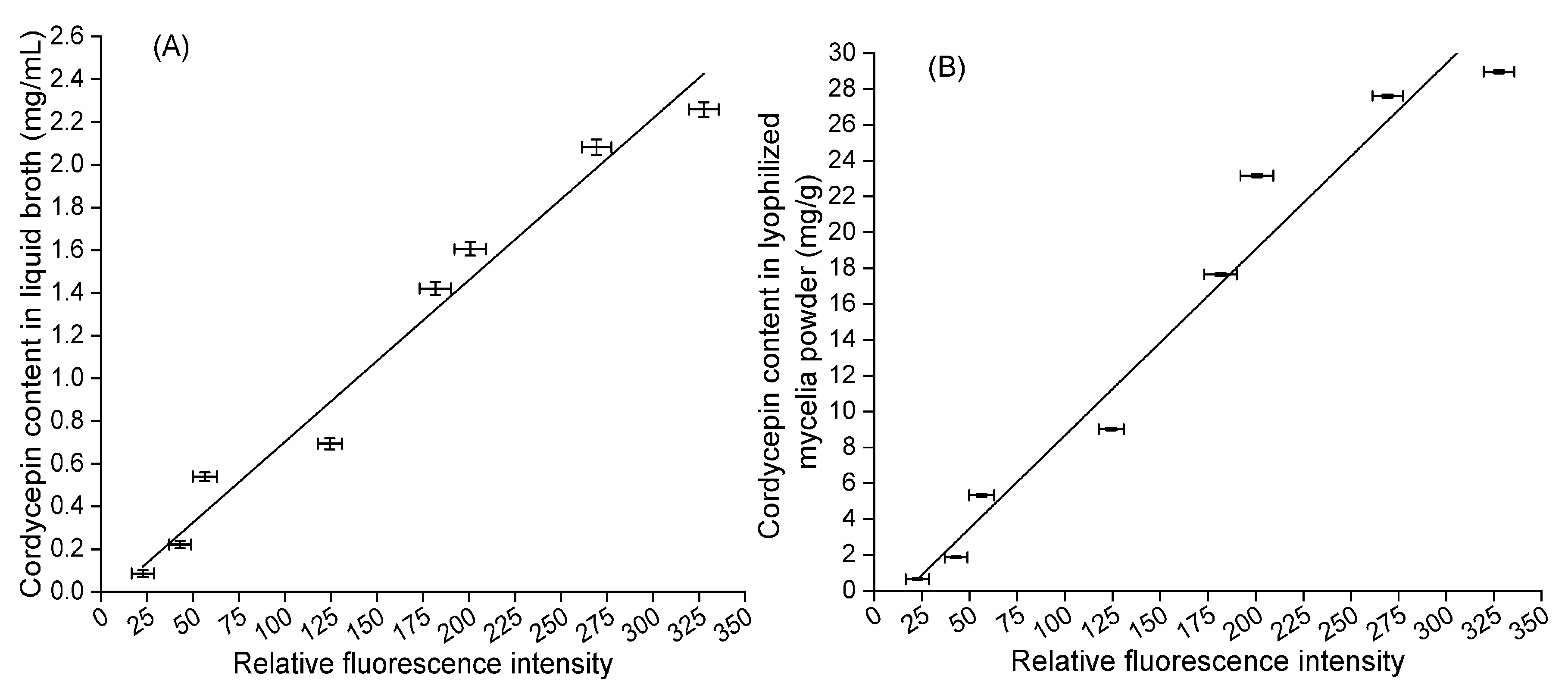
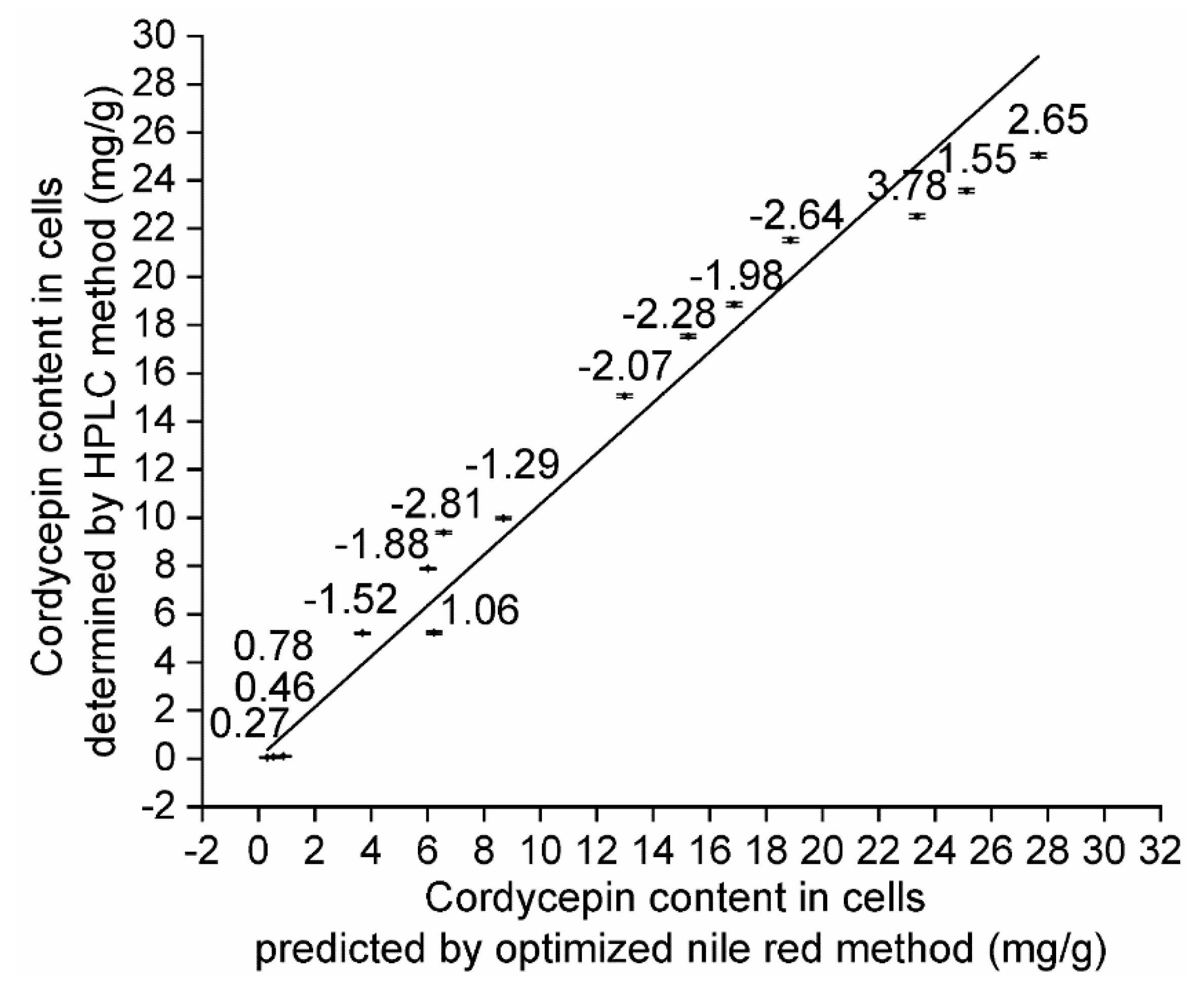
In our study, an optimized methylcellulose-based medium, a type of semisolid medium, was first used for isolating mutant spores of fungi. In brief, a fungal spore suspension was piped into optimized semisolid methylcellulose medium, and the medium thoroughly shaken to homogeneously disperse these spores. Then, the relative positions of these spores in optimized semisolid methylcellulose medium were fixed during static cultivation, and these spores formed single colonies and were separately transferred into downstream medium in 96-deep-well plates using a pipette with a 1000 μL tip. Notably, compared to traditional methods for isolating single fungal spores, such as the spread plate method, this novel method is simple, convenient and efficient for several reasons: (a) optimized semisolid methylcellulose medium is liquid at 0~70 °C, which is suitable for the growth of fungi; (b) pipette operation is simple, convenient and efficient; and (c) single spores can form single colonies inside the medium and on the surface of the medium. In addition, LMP of fungi was first used as the original material to measure neutral lipid content in lipid droplets using the Nile red method, and the quantitative relationships between the cordycepin concentration in liquid broth, cordycepin content in LMP, and corresponding RFI from Nile red-stained LMP were investigated. Eighteen stable mutants (CM1-1-1~CM1-3-5), individually producing from 0.001 (CM1-3-3) to 2.363 (CM1-1-1) mg/mL cordycepin, were finally obtained after LiCl+60Coγ compound mutagenesis. The RFI values from the Nile red-stained LMP of these cordycepin-producing strains were linearly correlated with the corresponding cordycepin concentration in liquid broth (R2 = 0.9332) when the LMP final concentration was 8 × 10−5 mg/mL. The mutant CM1-1-1, with the highest RFI from Nile red-stained LMP among the eighteen strains, was selected for optimizing the Nile red method, and the optimal method for RFI determination can be described as follows: 120 μL of LMP suspension, which was obtained by mixing LMP suspension prepared in PBS (1.11 × 10−4 mg/mL) with 10% DMSO (v/v) prior to a reaction in darkness at 30 °C for 10 min, was mixed with 30 µL of Nile red solution at final concentration of 3.2 µg/mL and thoroughly vortexed for 5 min at 30 °C prior to the determination of fluorescence intensities. Using this optimal method, the cordycepin concentration in liquid broth (R2 = 0.9727) and cordycepin content in LMP (R2 = 0.9613) were demonstrated to be linearly correlated with the corresponding RFI from Nile red-stained LMP of eight samples of mutant CM1-1-1 harvested under different culture conditions. In addition, the cordycepin content in LMP measured by the optimized Nile red method was linearly correlated to that determined by an HPLC method using five strains cultured for different times (R2 = 0.9627). In conclusion, in 18 cordycepin-producing fungi, neutral lipids in lipid droplets are required during cordycepin accumulation and considered potential biomarkers of cordycepin biosynthesis.
Cordycepin is a self-toxic component in cordycepin-producing fungi. These fungi can develop the capacity to resist and detoxify this toxic compound. 3′AMP is potentially formed and transferred into lipid droplets, and then cordycepin is biosynthesized by the two enzymes Cns1 and Cns2, which are colocalized on lipid droplets [
5]. Our data showed a significant relationship between the RFI from Nile red-stained LMP and cordycepin accumulation in cordycepin-producing fungi, suggesting a marked correlation between neutral lipids in lipid droplets and cordycepin biosynthesis. Therefore, we speculate that lipid droplets in these cordycepin-producing fungi enclose cordycepin to prevent it from contacting targets within their own cell and transport cordycepin to the vicinity of the cell membrane. Then, cordycepin is pumped into the extracellular matrix by a specific transporter. This mechanism potentially enhances the resistance and detoxification capacities of these fungi toward self-toxic cordycepin. In other words, lipid droplets may improve the cordycepin-producing capacity of these fungi. Thus, at most, 25.93% cordycepin remained in the cells of these strains (cultured at 25 °C for 30 d) used in our experiments. In other words, over 74.07% of cordycepin was pumped into liquid broth. Among these strains, mutant CM1-1-1 retained the least cordycepin in cells, which was less than 12.6%, under different culture conditions. In brief, neutral lipids in lipid droplets in the mycelium of cordycepin-producing fungi may be associated with detoxification of toxic cordycepin, but the effects are limited.
In addition, the multiple functions of lipid droplet-associated proteins may illuminate the positive effect of lipid droplets on cordycepin accumulation. Since lipid droplet-associated proteins vary substantially in different fungi, lipid droplet-resident proteins are involved in different signals, such as those involving signal peptides, posttranslation modification and nucleic acids, which may help these proteins localize lipid droplets [
6]. Notably, previous work demonstrated that
C. militaris mutant G81-3 produced abundant cordycepin in the mycelial stage by liquid surface culture. The over saturation of cordycepin resulted in crystallization when the cordycepin concentration in liquid broth exceeded approximately 7.4 g/L at 25 °C. The maximum capacity of cordycepin production reached 14.3 g/L through a decrease in the beginning substrate level and a water-supplementing strategy [
16]. Then, some work for the strain G81-3 in the mycelia stage when grown in liquid surface culture need further investigation to take better advantage of this excellent strain: (a) Whether cordycepin crystallization occurs in cells; (b) if cordycepin crystallization appears in cells, how do G81-3 cells respond to this issue; and (c) if cordycepin is biosynthesized without crystallization in cells throughout the mycelial stage, in spite of cordycepin being pumped outside cells through cordycepin transporters, can additional mechanisms also decrease the effective concentration of cordycepin in cells? For example, cordycepin might be enclosed in neutral lipids in lipid droplets of cells to downregulate the cordycepin level in the cytoplasm. (d) the detailed mechanisms that G81-3 can produced such large amount of cordycepin in the mycelia stage require further investigation. In addition, abundant cordycepin is produced during the fruiting stage of
C. militaris; during this time, signal transduction is markedly more active than it is in mycelia [
17]. As a result, the neutral lipid content may increase to overcome the unfavourable increase in cordycepin during the fruiting stage of cordycepin-producing fungi. Therefore, more cordycepin may be produced by mutant G81-3 in the fruiting stage than in the mycelial stage if G81-3 can form a fruiting body. Further investigations are required to clarify these issues. Our future work will focus on analysing the genetic expression and dynamics involved in droplet synthesis and cordycepin accumulation in
C. militaris cells.
4. Materials and Methods
4.1. Strains and Medium
C. militaris CGMCC 3.4655 (strain#2), Bacillus subtilis CGMCC 1.1849 and A. nidulans CGMCC 3.15737 (strain#3) were obtained from the China General Microbiological Culture Collection Center (Beijing, China). C. militaris CICC 14014 (strain #1) was obtained from the China Center of Industrial Culture Collection (Beijing, China). C. militaris and A. nidulans were maintained on PDA and YPD slants, respectively. Bacillus subtilis was cultured on nutrient agar (NA) medium. Basic liquid medium (g/L): glucose, 34; peptone, 17; KH2PO4, 1; and MgSO4·7H2O, 1. Optimized semisolid medium (g/L): basic liquid medium; methylcellulose, 23. Medium #A (g/L): glucose, 16; peptone, 7; KH2PO4, 1; and MgSO4·7H2O, 1. Medium #B (g/L): glucose, 6; peptone, 2; KH2PO4, 1; and MgSO4·7H2O, 1.
4.2. Materials
Nile red, DMSO and methylcellulose (viscosity: 4000 MPa·s) were obtained from Shanghai Sangon Biotech Limited Corporation (Shanghai, China). Nile red solution: 0.1 mg per 1 mL of acetone, was diluted with acetone to the appropriate concentration just prior to each assay. Cordycepin was obtained from the National Institutes for Food and Drug Control (Beijing, China). LiCl was obtained from Sinopharm Chemical Reagent Limited Corporation (Beijing, China). Methylene blue solution (0.1%) was obtained from Beijing Solarbio Science and Technology Co., Ltd. (Beijing, China).
4.3. Preparation of Spore Suspension, Lethality Assays and Antimicrobial Tests
Spore suspensions of strain #1 and strain #2, which had markedly different cordycepin-producing capabilities, were individually diluted with sterile PBS (pH 7.2) to 1 × 106 living conidia/mL and examined by methylene blue staining, optical microscopy (CX23, OLYMPUS, Tokyo, Japan) and hemocytometry. MSSs of strain #1 and strain #2 were prepared by mixing the same volume of each suspension.
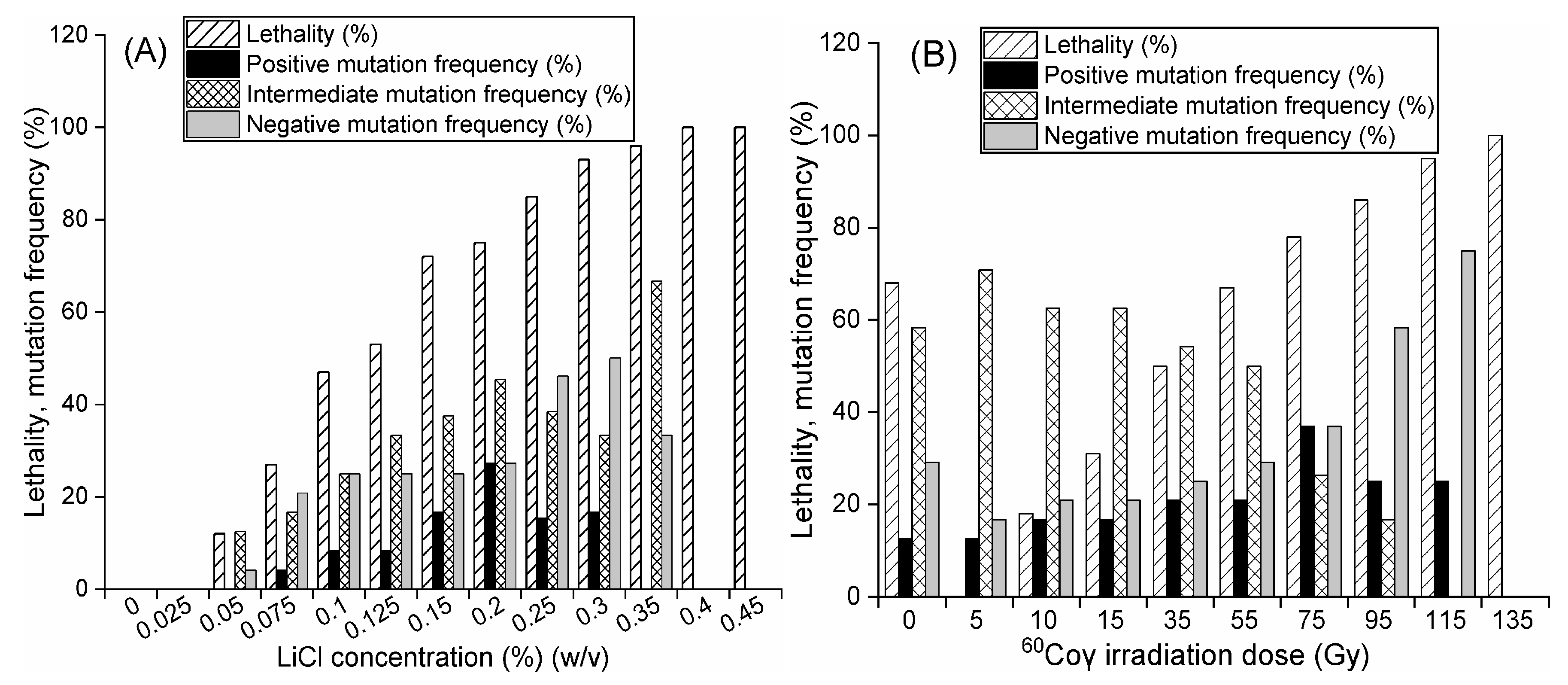
Mutant MSSs were appropriately diluted with sterile PBS, transferred into 20 mL of optimized semisolid medium in a 50 mL Erlenmeyer flask, fully shaken to homogeneously blend the spores, and incubated statically in darkness at 25 °C for 72 h. The relative positions of mutant MSSs in optimized semisolid medium were fixed during static cultivation. Variable isolates were transferred into downstream medium using a pipette with a 1000 µL tip. Lethality was calculated with the equation below by observing different isolates in optimized semisolid medium.
Eight random isolates under each mutation condition were separately transferred into 1.8 mL of basic liquid medium per well in 96-deep-well plates and incubated statically at 25 °C in darkness for 10 d (LiCl mutation) or 30 d (LiCl+
60Coγ compound mutagenesis and screening of stable mutants). Antimicrobial tests were performed with
B. subtilis [
18]. Eight sterile paper disks (7 mm) saturated with liquid broth were placed on each NA plate and incubated in darkness at 30 °C for 12 h. Lethality and mutant frequency were calculated by comparing these groups with the nonirradiated groups:
Using Equations (1)–(4) the mutants which showed an inhibition diameter over 20% larger than that of strain #1 were defined as positive mutants, and the mutants with a 20% or smaller inhibition diameter than strain #2 were considered negative mutants. NB is the number of single colonies before mutagenesis, NA is the number of single colonies after mutagenesis, PMF is the positive mutant frequency, NEC is the number of positive mutant colonies, NVC is the number of viable colonies, NMF is the negative mutant frequency, IMF is the intermediate mutant frequency and NDC is the number of negative mutant colonies.
4.4. Mutation Procedure
According to the modified mutation method using LiCl [
19] that stimulated base substitution and significantly enhanced the mutation efficiency in combined mutagenesis [
19,
20], MSSs were incubated with different concentrations of LiCl (0.025, 0.05, 0.075, 0.1, 0.125, 0.15, 0.2, 0.25, 0.3, 0.35, 0.4 and 0.45%) (
w/
v) prepared in sterile PBS and cultured in darkness at 25 °C for 12 h. Lethality assays and antimicrobial tests were performed, and the optimal concentration of LiCl was selected according to lethality and mutant frequency.
According to the modified mutation method under
60Coγ ray [
21], which mechanistically induced a large number of double-stranded breaks of chromosomal DNA and caused genetic variability [
22], after treatment with LiCl at the optimal concentration for 5 h, MSSs were irradiated under a
60Coγ ray (
60Co irradiator, CN-101, CNNC, Gansu Tianchen Irradiation Technology Co. Ltd., Lanzhou, China) at different doses (5, 10, 15, 35, 55, 75, 95, 115 and 135 Gy) for 15 min. Positive, intermediate and negative mutants under each dose of
60Coγ irradiation were preliminarily screened according to their inhibition diameters being obviously different from those of their initial strains in antimicrobial tests. Throughout six d of cultivation on PDA slants for six generations, 15 stable mutants were individually cultured at 25 °C for 30 d, selected by comparing the inhibition diameters of liquid broth obtained from mutants of the first generation with those of the corresponding mutants of the sixth generation using antimicrobial test method as described in
Section 4.3.
A single colony of each selected strain (stable mutants, strain #1, strain #2 and strain #3), prepared using the method described in
Section 4.3, was individually transferred into 20 mL of basic liquid medium in a 50 mL centrifuge tube and incubated in darkness at 25 °C for 30 d, and liquid broth was filtered through 22 µm filter paper prior to determining the cordycepin concentration in liquid broth using HPLC. HPLC was performed with the following parameters: Agilent 1260 platform (Santa Clara, CA, USA), DiamonsilC18(2) (5 μm × 4.6 mm × 250 mm), 15% methanol (Merck, Germany): 85% water (
v/
v), flow rate: 1 mL/min, 35°C, injection volume: 20 µL, and detection wavelength: 260 nm. The mycelia of these strains were used for determining fluorescence intensities.
4.5. LMP Preparation
The mycelia of the stable mutants, strain #1, strain #2 and strain #3 were filtered with three-layer gauze, washed with pure water three times, frozen in a cryogenic refrigerator (Forma 900 series, Thermo, Waltham, MA, USA) at −80 °C for 12 h, dried in a vacuum freeze dryer (Labconco, Kansas, MO, USA) for 28 h, ground into LMP in a mortar, and then filtered through a 90 mesh sieve. The mortar and sieve were washed with 95% ethanol three times prior to the next LMP being treated. According to a previously described method [
23], water extracts (10 mL) of treated LMP (0.1 g) of the stable mutants, strain #1, strain #2 and strain #3 were prepared to determine the cordycepin concentration in the cells using HPLC. The residual treated LMP was stored at −20 °C before being used in subsequent experiments.
4.6. Determination of Fluorescence Intensity by a Multiscan Spectrum
LMP (10 ± 0.1 mg) was weighed with an electronic balance (Shanghai Yueping Scientific Instrument Co., Ltd., Shanghai, China) and diluted with 10 mL of PBS prior to each assay. Then, 120 µL of LMP suspension was mixed with 30 μL of Nile red solution at a final concentration of 3.2 μg/mL. Treated LMP was agitated on a 96-well plate oscillator vigorously for 5 min before determining the fluorescence intensities on a microplate reader (Infinite M200 pro, Männedorf, Kanton Zürich, Switzerland) with a 490 nm excitation wavelength and a 598 nm emission wavelength. The RFI was calculated after subtracting the self-fluorescence of Nile red and the fluorescence intensity of the LMP suspension at 598 nm.
4.7. Optimizing the Nile red Method
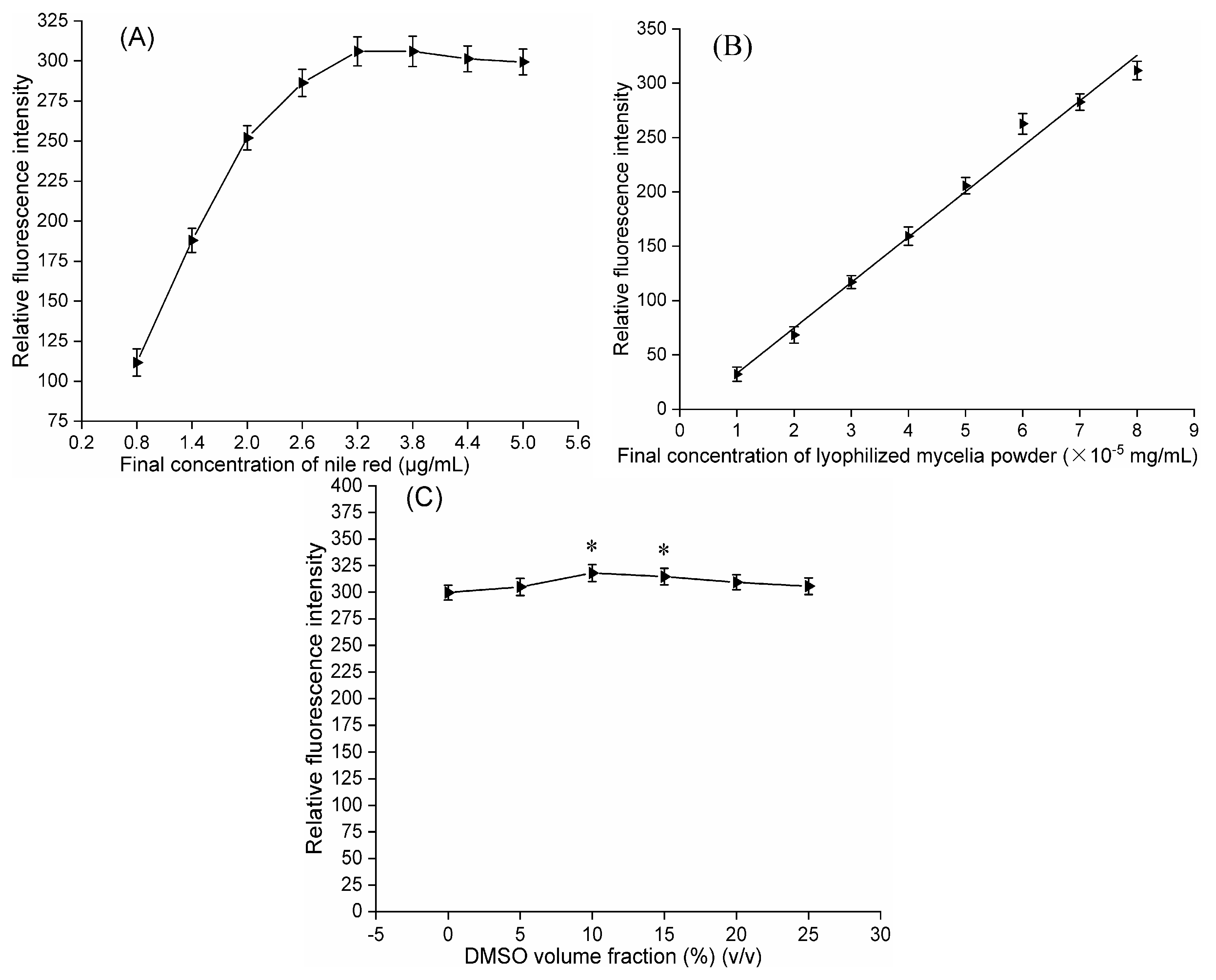
For optimization, 120 µL of LMP suspensions at seven different concentrations (1, 1 × 10
−1, 1 × 10
−2, 1 × 10
−3, 1 × 10
−4, 1 × 10
−5 and 1 × 10
−6 mg/mL) were mixed with 30 µL of Nile red solution at a final concentration of 3.2 µg/mL and vortexed vigorously in darkness at 30 °C for 5 min [
24] before determining the fluorescence intensities. The relationships between the cordycepin concentration in liquid broth, cordycepin concentration in LMP, and the corresponding RFI from Nile red-stained LMP were evaluated at the optimal concentration of each LMP for the selected eighteen strains, with the highest RFI from Nile red-stained LMP determined at seven different concentrations. The strain with the maximal RFI from Nile red-stained LMP at the optimal concentration was selected for the following experiments.
Next, 120 µL aliquots of LMP suspension of the selected strain at the optimal concentration were individually mixed with 30 µL of different Nile red solutions in acetone (4, 7, 10, 13, 16 and 19 µg/mL) at different final concentrations (0.8, 1.4, 2.0, 2.6, 3.2 and 3.8 µg/mL) and vortexed vigorously at 30°C in darkness for 5 min prior to determining the fluorescence intensity.
Then, 120 µL of LMP suspensions of the selected strain at eight different concentrations (1, 0.875, 0.75, 0.625, 0.5, 0.375, 0.25 and 0.125 × 10−4 mg/mL) were individually mixed with 30 µL of Nile red solution at the optimal final concentration and vortexed vigorously at 30 °C in darkness for 5 min before the determination of the fluorescence intensity.
To maintain the optimal final concentration of LMP, LMP suspensions were individually mixed with a volume ratio of DMSO (5%, 10%, 15%, 20% and 25%) (v/v) and reacted in darkness at 30 °C for 10 min. For this purpose, 120 µL of pretreated LMP suspension was mixed with 30 µL of Nile red solution at the optimal final concentration and vortexed vigorously in darkness at 30 °C for 5 min before determining fluorescence intensity.
4.8. Cordycepin Content Quantification by Relative Fluorescence Intensity
A single colony of the selected strain that was prepared using the method described in
Section 4.3 was transferred into 20 mL of basic liquid medium in a 50 mL centrifuge tube and statically cultured under eight different conditions (#1–#8) as follows: conditions #1–#2 consisted of 25°C, 30 d and different media (medium #A and medium #B), while conditions #3–#8 consisted of 25°C, different culture times (5, 10, 15, 20, 25 and 30 d) and basic liquid medium. The relationships between the cordycepin concentration in liquid broth, cordycepin content in LMP and the corresponding RFI from Nile red-stained LMP were investigated.
4.9. Data Analysis
Three replicates of each experiment were performed. The results were expressed as the mean ± standard deviation and analysed through one-way ANOVA using SPSS 22.0 software (International Business Machines Corporation, Armonk, NY, USA). Statistical differences at the 0.05 level using the least significant difference (LSD) method were considered significant.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






