Perspective on the Therapeutics of Anti-Snake Venom



Abstract
1. Introduction
2. Methodology
3. Current Information in the Design of New Antivenoms
- Immediate hypersensitivity reaction to the alien immunoglobulins, including anaphylactic and pyrogenic reactions such as chills, rigor, headache, and tachycardia. Delayed antivenom reactions or serum sickness is observed after 8 to 12 days of treatment; these are characterized by cutaneous eruptions, fever, and allergies, among other effects [18];
- Limited efficacy of antivenom therapy to protect the affected organ/s against immediate local tissue damage and low stability;
- Ineffectiveness of the antivenom due to significant geographic variation in the composition of the venom;
- Antigenic reactivity due to the taxonomic diversity of the snakes;
- Improper use of the antivenom due to incorrect medical management, which contributes to a high incidence of adverse reactions, a low toxin neutralizing potency, or both.
- The instability in the antivenom market;
- Little financial incentive for pharmacists and health centers to sell antivenom due to the low profit margins;
- Lack of comprehensive data on how many doses of antivenom are required and where they should be distributed [20].
4. Current Drugs for the Treatment of Snakebites in the United States
- Antivenin® Wyeth (equine), approved to treat envenomation from the American coral (Micrurus fulvius) [27];
- Crofab® Crotalidae polyvalent immune FAB (ovine), approved in 2000 for snakebite envenomation from multiple species, including Crotalus atrox (western diamondback rattlesnake), Crotalus adamanteus (eastern diamondback rattlesnake), Crotalus scutulatus (Mojave rattlesnake), Agkistrodon contortrix (copperhead), and Agkistrodon piscivorus (cottonmouth or water moccasin) [28];
- Anavip® Crotalidae Immune F(ab’)2 (equine), approved in 2015 to treat envenomation from the rattlesnakes Crotalus durissus and Bothrops asper [29].
5. Folkloric Medicine in the Treatment of Snake Envenomation
- The isolation or derivatization of bioactive substances from natural sources;
- The evaluation of safety and efficacy using pharmacological methods;
- Evaluation of safety and efficacy by conventional pharmacological methods (pharmacodynamics, toxicology, and pharmacokinetics);
- Regulatory approval of the therapeutic agent to be used in the market and in post-marketing supervision, as well as pharmacovigilance.
6. Mechanisms of Action of Antivenoms Derived from Herbs
6.1. Protein Precipitation
6.2. Enzyme Inactivation/Inhibition
6.3. Chelation Activity
6.4. Adjuvant Action
6.5. Antioxidant Activity
7. Natural Products in the Development of Antivenom Agents
8. Drawbacks of Herbal Products in the Treatment of Snake Envenomation
9. Challenges Involved in the Discovery of Antivenoms
10. Snake Venom as a Source of Therapeutic Agents
11. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Available online: https://www.who.int/neglected_diseases/EB132_R7_en.pdf (accessed on 5 March 2019).
- Warrell, D.A.; Gutiérrez, J.M.; Calvete, J.J.; Williams, D. New approaches & technologies of venomics to meet the challenge of human envenoming by snakebites in India. Indian J. Med Res. 2013, 138, 38–59. [Google Scholar] [PubMed]
- WHO. Available online: https://www.who.int/snakebites/en/ (accessed on 9 September 2018).
- Fry, B.G. Snakebite: When the Human Touch Becomes a Bad Touch. Toxins 2018, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Ralph, R.; Sharma, S.K.; Faiz, M.A.; Ribeiro, I.; Rijal, S.; Chappuis, F.; Kuch, U. The timing is right to end snakebite deaths in South Asia. BMJ 2019, 364, k5317. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins. Available online: https://www.who.int/bloodproducts/snake_antivenoms/snakeantivenomguide/en/ (accessed on 9 December 2018).
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Primers 2017, 14, 17063. [Google Scholar]
- Kini, R.M.; Doley, R. Structure, function and evolution of three-finger toxins: Mini proteins with multiple targets. Toxicon 2010, 56, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M. Understanding and confronting snakebite envenoming: The harvest of cooperation. Toxicon 2016, 109, 51–62. [Google Scholar] [CrossRef]
- Chippaux, J.P.; Goyffon, M. Venomous and poisonous animals. I. Overview. Med Trop 2006, 66, 215–220. [Google Scholar]
- Giovannini, P.; Howes, M.-J.R. Medicinal plants used to treat snakebite in Central America: Review and assessment of scientific evidence. J. Ethnopharmacol. 2017, 199, 240–256. [Google Scholar] [CrossRef]
- Félix-Silva, J.; Silva-Junior, A.A.; Zucolotto, S.M.; Fernandes-Pedrosa, M.D.F. Medicinal plants for the treatment of local tissue damage induced by Snake Venoms: An overview from traditional use to pharmacological evidence. Evid Based Complement. Alternat. Med. 2017, 5748256. [Google Scholar] [CrossRef]
- Upasani, S.V.; Beldar, V.G.; Tatiya, A.U.; Upasani, M.; Surana, S.J.; Patil, D.S. Ethnomedicinal plants used for snakebite in India: a brief overview. Integr. Med. Res. 2017, 6, 114–130. [Google Scholar] [CrossRef]
- Bermúdez-Méndez, E.; Fuglsang-Madsen, A.; Føns, S.; Lomonte, B.; Gutiérrez, J.M.; Laustsen, A.H. Innovative Immunization Strategies for Antivenom Development. Toxins 2018, 10, 452. [Google Scholar] [CrossRef] [PubMed]
- Landon, J.; Smith, D.S. Merits of Sheep Antisera for Antivenom Manufacture. J. Toxicol. Toxin Rev. 2003, 22, 15–22. [Google Scholar] [CrossRef]
- Aird, S.D. Ophidian envenomation strategies and the role of purines. Toxicon 2002, 40, 335–393. [Google Scholar] [CrossRef]
- Heard, K.; O’Malley, G.F.; Dart, R.C. Antivenom therapy in the Americas. Drugs 1999, 58, 5–15. [Google Scholar] [CrossRef] [PubMed]
- De Silva, H.A.; Ryan, N.M.; de Silva, J. Adverse reactions to snake antivenom, and their prevention and treatment. Br. J. Clin. Pharmacol. 2016, 81, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, C.L.S.; Moreira-Dill, L.S.; Fernandes, R.S.; Costa, T.R.; Hage-Melim, L.I.S.; Marcussi, S.; A Carvalho, B.M.; Da Silva, S.L.; Zuliani, J.P.; Fernandes, C.F.C.; et al. Biodiversity as a source of bioactive compounds against snakebites. Curr. Med. Chem. 2014, 21, 2952–2979. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.-P. Estimating the Global Burden of Snakebite Can Help to Improve Management. PLoS Med. 2008, 5, e221. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M.; Sidhu, S.S.; Laustsen, A.H. Biosynthetic Oligoclonal Antivenom (BOA) for Snakebite and Next-Generation Treatments for Snakebite Victims. Toxins 2018, 10, 534. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.; Engmark, M.; Milbo, C.; Johannesen, J.; Lomonte, B.; Gutiérrez, J.; Lohse, B. From Fangs to Pharmacology: The Future of Snakebite Envenoming Therapy. Curr. Pharm. Des. 2016, 22, 5270–5293. [Google Scholar] [CrossRef]
- Williams, H.E.; Layfield, H.J.; Vallance, T.; Patel, K.; Bicknell, A.B.; Trim, S.A.; Vaiyapuri, S. The Urgent Need to Develop Novel Strategies for the Diagnosis and Treatment of Snakebites. Toxins 2019, 11, 363. [Google Scholar] [CrossRef]
- Dandona, R.; Kumar, G.A.; Kharyal, A.; George, S.; Akbar, M.; Dandona, L. Mortality due to snakebite and other venomous animals in the Indian state of Bihar: Findings from a representative mortality study. PLoS ONE 2018, 13, e0198900. [Google Scholar] [CrossRef] [PubMed]
- Gummin, D.D.; Mowry, J.B.; Spyker, D.A.; Brooks, D.E.; Osterthaler, K.M.; Banner, W. Annual Report of the American Association of Poison Control Centers’ National Poison Data System (NPDS): 35th Annual Report. Clin. Toxicol. 2017, 56, 1213–1415. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Gutiérrez, J.M.; Knudsen, C.; Johansen, K.H.; Méndez, E.B.; Cerni, F.A.; Jürgensen, J.A.; Ledsgaard, L.; Esteban, A.M.; Øhlenschlæger, M.; et al. Pros and cons of different therapeutic antibody formats for recombinant antivenom development. Toxicon 2018, 146, 151–175. [Google Scholar] [CrossRef] [PubMed]
- Antivenin® [Prescribing Information]. Wyeth Lab. August 2001. Available online: https://www.fda.gov/downloads/BiologicsBloodVaccines/UCM277363.pdf (accessed on 9 December 2018).
- CroFab® [Prescribing Information]. BTG International Inc. May 2017. Available online: 17 https://www.fda.gov/downloads/BloodBloodProducts/ucm117573.pdf (accessed on 9 December 2018).
- Anavip® [Prescribing Information]. Rare Disease Therapeutics, Inc. May 2015. Available online: ttps://www.fda.gov/downloads/BiologicsBloodVaccines/BloodBloodProducts/ApprovedProducts/LicensedProductsBLAs/FractionatedPlasmaProducts/UCM446175.pdf (accessed on 9 December 2018).
- Scheske, L.; Ruitenberg, J.; Bissumbhar, B. Needs and availability of snake antivenoms: relevance an application of international guidelines. Int. J. Health Policy Manag. 2015, 4, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.I.; Ryan, N.M.; Page, C.B.; Buckley, N.A.; Brown, S.G.; O’Leary, M.A.; Isbister, G.K. The Australian Snakebite Project, 2005-2015 (ASP-20). Med. J. Aust. 2017, 207, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.Y.; Zhou, S.F.; Gao, S.H.; Yu, Z.L.; Zhang, S.F.; Tang, M.K.; Sun, J.N.; Ma, D.L.; Han, Y.F.; Fong, W.F.; et al. New Perspectives on How to Discover Drugs from Herbal Medicines: CAM’s Outstanding Contribution to Modern Therapeutics. Evidence-Based Complement. Alternat. Med. 2013, 2013, 627375. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, H.; Matos, F.J.A. Plantas Medicinais no Brasil: Nativas e Exóticas; Instituto Plantarum de Estudos da Flora: São Paulo, Brasil, 2000. [Google Scholar]
- Calderon, L.D.A.; Zuliani, J.P.; Da Silva, L.H.P.; Ii, I.; Silva-Jardim, I.; Silva, A.D.A.E.; Iii, I.; Ciancaglini, P.; Stábeli, R.G. Amazonian biodiversity: a view of drug development for leishmaniasis and malaria. J. Braz. Chem. Soc. 2009, 20, 1011–1023. [Google Scholar] [CrossRef]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef]
- Chin, Y.-W.; Balunas, M.J.; Chai, H.B.; Kinghorn, A.D. Drug discovery from natural sources. AAPS J. 2006, 8, E239–E253. [Google Scholar] [CrossRef]
- Schmidt, T.; Khalid, S.; Romanha, A.; Alves, T.; Biavatti, M.; Brun, R.; Da Costa, F.; De Castro, S.; Ferreira, V.; De Lacerda, M.; et al. The Potential of Secondary Metabolites from Plants as Drugs or Leads Against Protozoan Neglected Diseases - Part II. Curr. Med. Chem. 2012, 19, 2176–2228. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Houghton, P.J.; Osibogun, I.M. Flowering plants used against snakebite. J. Ethnopharmacol. 1993, 39, 1–29. [Google Scholar] [CrossRef]
- Gomes, A.; Das, R.; Sarkhel, S.; Mishra, R.; Mukherjee, S.; Bhattacharya, S.; Gomes, A. Herbs and herbal constituents active against snakebite. Indian J. Exp. Biol. 2010, 48, 865–878. [Google Scholar] [PubMed]
- Castro, O.; Gutiérrez, J.M.; Barrios, M.; Castro, I.; Romero, M.; Umaña, E. Neutralization of the hemorrhagic effect induced by Bothrops asper (Serpentes: Viperidae) venom with tropical plant extracts. Revista de Biología Tropical 1999, 47, 605–616. [Google Scholar] [PubMed]
- Melo, P.A.; Nascimento, M.C.D.; Mors, W.B.; Suarez-Kurtz, G. Inhibition of the myotoxic and hemorrhagic activities of crotalid venoms by Eclipta prostrata (Asteraceae) extracts and constituents. Toxicon 1994, 32, 595–603. [Google Scholar] [CrossRef]
- Mors, W.B.; Nascimento, M.C.D.; Parente, J.; Da Silva, M.H.; Melo, P.A.; Suarez-Kurtz, G. Neutralization of lethal and myotoxic activities of South American rattlesnake venom by extracts and constituents of the plant Eclipta prostrata (Asteraceae). Toxicon 1989, 27, 1003–1009. [Google Scholar] [CrossRef]
- Núñez, V.; Castro, V.; Murillo, R.; Ponce-Soto, L.A.; Merfort, I.; Lomonte, B. Inhibitory effects of Piper umbellatum and Piper peltatum extracts towards myotoxic phospholipases A2 from Bothrops snake venoms: Isolation of 4-nerolidylcatechol as active principle. Phytochemistry 2005, 66, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Chilpa, R.; Gómez-Garibay, F.; Quijano, L.; Magos-Guerrero, G.A.; Rios, T. Preliminary results on the protective effect of (-)-edunol, a pterocarpan from Brongniartia podalyrioides (Leguminosae), against Bothrops atrox venom in mice. J. Ethnopharmacol. 1994, 42, 199–203. [Google Scholar] [CrossRef]
- Amui, S.F.; Puga, R.D.; Soares, A.M.; Giuliatti, S. Plant-antivenom: Database of anti-venom medicinal plants. Eletronic, J. Biotechnol 2010, 14, 6–7. [Google Scholar] [CrossRef]
- Chandrashekara, K.T.; Nagaraju, S.; Nandini, S.U.; Kemparaju, K. Neutralization of local and systemic toxicity of Daboia russelli venom by Morus alba plant leaf extract. Phytother. Res. 2009, 23, 1082–1087. [Google Scholar] [CrossRef]
- Fung, S.Y.; Tan, N.H.; Liew, S.H.; Sim, S.M.; Aguiyi, J.C. The protective effects of Mucuna pruriens seed extract against histopathological changes induced by Malayan cobra (Naja sputatrix) venom in rats. Trop. Biomed. 2009, 26, 80–84. [Google Scholar]
- Tan, N.H.; Fung, S.Y.; Sim, S.M.; Marinello, E.; Guerranti, R.; Aguiyi, J.C. The protective effect of Mucuna pruriens seeds against snake venom poisoning. J. Ethnopharmacol. 2009, 123, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Scirè, A.; Tanfani, F.; Bertoli, E.; Furlani, E.; Nadozie, H.-O.N.; Cerutti, H.; Cortelazzo, A.; Bini, L.; Guerranti, R. The belonging of gpMuc, a glycoprotein from Mucuna pruriens seeds, to the Kunitz-type trypsin inhibitor family explains its direct anti-snake venom activity. Phytomedicine 2011, 18, 887–895. [Google Scholar] [CrossRef]
- Hasson, S.S.; Al-Jabri, A.A.; Sallam, T.A.; Al-Balushi, M.S.; Mothana, R.A.A. Antisnake Venom Activity of Hibiscus aethiopicus L. against Echis ocellatus and Naja nigricollis. J. Toxicol. 2010, 2010, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mahadeswaraswamy, Y.H.; Devaraja, S.; Kumar, M.S.; Goutham, Y.N.J.; Kemparaju, K. Inhibition of local effects of Indian Daboia/Vipera russelli venom by the methanolic extract of grape (Vitis vinifera L.) seeds. Indian J. Biochem. Biophys. 2009, 46, 154–160. [Google Scholar] [PubMed]
- Nazato, V.S.; Rubem-Mauro, L.; Vieira, N.A.G.; Rocha-Junior, D.D.S.; Silva, M.G.; Lopes, P.S.; Dal-Belo, C.A.; Cogo, J.C.; Dos Santos, M.G.; Da Cruz-Höfling, M.A.; et al. In Vitro Antiophidian Properties of Dipteryx alata Vogel Bark Extracts. Molecules 2010, 15, 5959–5970. [Google Scholar] [CrossRef] [PubMed]
- Puebla, P.; Oshima-Franco, Y.; Franco, L.M.; Dos Santos, M.G.; Da Silva, R.V.; Rubem-Mauro, L.; Feliciano, A.S. Chemical Constituents of the Bark of Dipteryx alata Vogel, an Active Species against Bothrops jararacussu Venom. Molecules 2010, 15, 8193–8204. [Google Scholar] [CrossRef]
- Magalhães, A.; Dos Santos, G.B.; Verdam, M.C.D.S.; Fraporti, L.; Malheiro, A.; Lima, E.S.; Dos-Santos, M.C. Inhibition of the inflammatory and coagulant action of Bothrops atrox venom by the plant species Marsypianthes chamaedrys. J. Ethnopharmacol. 2011, 134, 82–88. [Google Scholar] [CrossRef][Green Version]
- Assafim, M.; De Coriolano, E.C.; Benedito, S.E.; Fernandes, C.P.; Lobo, J.F.R.; Sanchez, E.F.; Rocha, L.M.; Fuly, A.L. Hypericum brasiliense plant extract neutralizes some biological effects of Bothrops jararaca snake venom. J. Ven. Res. 2011, 2, 11–16. [Google Scholar]
- A Ibrahim, M.; Aliyu, A.B.; Abusufiyanu, A.; Bashir, M.; Sallau, A.B. Inhibition of Naja nigricolis (Reinhardt) venom protease activity by Luffa egyptiaca (Mill) and Nicotiana rustica (Linn) extracts. Indian J. Exp. Biol. 2011, 49, 552–554. [Google Scholar]
- Dhananjaya, B.L.; Zameer, F.; Girish, K.S.; D’Souza, C.J.M. Anti-venom potential of aqueous extract of stem bark of Mangifera indica L. against Daboia russellii (Russell’s viper) venom. Indian J. Biochem. Biophys. 2011, 48, 175–183. [Google Scholar] [PubMed]
- Torres, A.M.; Camargo, F.J.; Ricciardi, G.A.L.; Ricciardi, A.I.A.; Dellacassa, E. Neutralizing effects of Nectandra angustifolia extracts against Bothrops neuwiedi snake venom. Nat. Prod. Commun. 2011, 6, 1393–1396. [Google Scholar] [CrossRef] [PubMed]
- Razi, M.T.; Asad, M.H.; Khan, T.; Zabta Chaudhary, M.; Tayyab Ansari, M.; Arshad, M.A.; Saqib, Q.N.U. Antihaemorrhagic potentials of Fagonia cretica against Naja naja karachiensis (Black Pakistan cobra) venom. Nat. Prod. Res. 2011, 25, 1902–1907. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.L.; Marcussi, S.; Fernandes, R.S.; Pereira, P.S.; Januário, A.H.; França, S.C.; Da Silva, S.L.; Soares, A.M.; Lourenço, M.V. Anti-snake venom activities of extracts and fractions from callus cultures of Sapindus saponaria. Pharmacol. Biol. 2012, 50, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Nishijima, C.M.; Rodrigues, C.M.; Silva, M.A.; Lopes-Ferreira, M.; Vilegas, W.; Hiruma-Lima, C.A. Anti-hemorrhagic Activity of Four Brazilian Vegetable Species Against Bothrops jararaca Venom. Molecules 2009, 14, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Ushanandini, S.; Nagaraju, S.; Nayaka, S.C.; Kumar, K.H.; Kemparaju, K.; Girish, K.S. The anti-ophidian properties of Anacardium occidentale bark extract. Immunopharmacol. Immunotoxicol. 2009, 31, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Ushanandini, S.; Nagaraju, S.; Kumar, K.H.; Vedavathi, M.; Machiah, D.K.; Kemparaju, K.; Vishwanath, B.S.; Gowda, T.V.; Girish, K.S. The anti-snake venom properties of Tamarindus indica (leguminosae) seed extract. Phytotherapy Res. 2006, 20, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Rizzini, C.T.; Mors, W.B.; Pereira, N.A. Plantas brasileiras tidas como ativas contra peçonhas de animais, especialmente veneno de cobra. Rev. Bras. Farmacog 1988, 69, 5. [Google Scholar]
- Mors, W.B. Plants against snake-bites. Memorias do Instituto Oswaldo Cruz 1991, 86 (Suppl. 2), 193. [Google Scholar] [CrossRef]
- Martz, W. Plants with a reputation against snakebite. Toxicon 1992, 30, 1131–1142. [Google Scholar] [CrossRef]
- Otero, R.; Fonnegra, R.; Jiménez, S.L.; Núñez, V.; Evans, N.; Alzate, S.P.; E García, M.; Saldarriaga, M.; Del Valle, G.; Osorio, R.G.; et al. Snakebites and ethnobotany in the northwest region of Colombia: Part I: traditional use of plants. J. Ethnopharmacol. 2000, 71, 493–504. [Google Scholar] [CrossRef]
- Soares, A.; Januario, A.; Lourenço, M.; Pereira, A.; Pereira, P.S. Neutralizing effects of Brazilian plants against snake venoms. Drugs Futur. 2004, 29, 1105. [Google Scholar] [CrossRef]
- Lomonte, B.; León, G.; Angulo, Y.; Rucavado, A.; Núñez, V. Neutralization of Bothrops asper venom by antibodies, natural products and synthetic drugs: Contributions to understanding snakebite envenomings and their treatment. Toxicon 2009, 54, 1012–1028. [Google Scholar] [CrossRef] [PubMed]
- Tusha.; Basak, S.; Sarma, G.C.; Rangan, L. Ethnomedical uses of Zingiberaceous plants of Northeast India. J. Ethnopharmacol. 2010, 132, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Molander, M.; Saslis-Lagoudakis, C.H.; Jäger, A.K.; Rønsted, N. Cross-cultural comparison of medicinal floras used against snakebites. J. Ethnopharmacol. 2012, 139, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Dey, A.; Nath De, N. Phytopharmacology of antiophidian botanicals. Inter. J. Pharmacol. 2012, 8, 18. [Google Scholar] [CrossRef]
- Houghton, P.J.; Skari, K.P. The effect on blood clotting of some west African plants used against snakebite. J. Ethnopharmacol. 1994, 44, 99–108. [Google Scholar] [CrossRef]
- Molander, M.; Nielsen, L.; Sogaard, S.; Staerk, D.; Rønsted, N.; Diallo, D.; Chifundera, K.Z.; van Staden, J.; Jäger, A.K. Hyaluronidase, Phospholipase A2 and protease inhibitory activity of plants used in traditional treatments of snake-induced tissue necrosis in Mali, DR Congo and South Africa. J. Ethnopharmacol. 2014, 157, 171–180. [Google Scholar] [CrossRef]
- Vale, L.H.F.; Mendes, M.M.; Hamaguchi, A.; Rodrigues, V.M.; Homsi-Brandeburgo, M.I.; Soares, A.M. Neutralization of Pharmacological and Toxic Activities of Bothrops Snake Venoms by Schizolobium parahyba (Fabaceae) Aqueous Extract and Its Fractions. Basic Clin. Pharmacol. Toxicol. 2008, 103, 104–107. [Google Scholar] [CrossRef]
- Hung, Y.-C.; Sava, V.; Hong, M.-Y.; Huang, G. Inhibitory effects on phospholipase A2 and antivenin activity of melanin extracted from Thea sinensis Linn. Life Sci. 2004, 74, 2037–2047. [Google Scholar] [CrossRef]
- Alam, M.I.; Gomes, A. Adjuvant effects and antiserum action potentiation by a (herbal) compound 2-hydroxy-4-methoxy benzoic acid isolated from the root extract of the Indian medicinal plant “sarsaparilla” (Hemidesmus indicus R. Br.). Toxicon 1998, 36, 1423–1431. [Google Scholar] [CrossRef]
- Chatterjee, I.; Chakravarty, A.; Gomes, A. Daboia russellii and Naja kaouthia venom neutralization by lupeol acetate isolated from the root extract of Indian sarsaparilla Hemidesmus indicus R.Br. J. Ethnopharmacol. 2006, 106, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Gupta, Y.K.; Peshin, S.S. Snake bite in India: current scenario of an old problem. J. Clin. Toxicol. 2014, 4, 1–9. [Google Scholar] [CrossRef]
- Gómez-Betancur, I.; Pereañez, J.A.; Patiño, A.C.; Benjumea, D. Inhibitory effect of pinostrobin from Renealmia alpinia, on the enzymatic and biological activities of a PLA2. Int. J. Biol. Macromol. 2016, 89, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Santhosh, M.S.; Hemshekhar, M.; Sunitha, K.; Thushara, R.M.; Jnaneshwari, S.; Kemparaju, K.; Girish, K.S. Snake Venom Induced Local Toxicities: Plant Secondary Metabolites as an Auxiliary Therapy. Mini-Reviews Med. Chem. 2013, 13, 106–123. [Google Scholar] [CrossRef]
- Leanpolchareanchai, J.; Pithayanukul, P.; Bavovada, R.; Saparpakorn, P. Molecular Docking Studies and Anti-enzymatic Activities of Thai Mango Seed Kernel Extract Against Snake Venoms. Molecules 2009, 14, 1404–1422. [Google Scholar] [CrossRef] [PubMed]
- Toyama, D.; Marangoni, S.; Diz-Filho, E.; Oliveira, S.; Toyama, M. Effect of umbelliferone (7-hydroxycoumarin, 7-HOC) on the enzymatic, edematogenic and necrotic activities of secretory phospholipase A2 (sPLA2) isolated from Crotalus durissus collilineatus venom. Toxicon 2009, 53, 417–426. [Google Scholar] [CrossRef]
- Mors, W.B.; Nascimento, M.C.D.; Pereira, B.M.R.; Pereira, N.A. Plant natural products active against snake bite — the molecular approach. Phytochemistry 2000, 55, 627–642. [Google Scholar] [CrossRef]
- Singh, N.; Jabeen, T.; Sharma, S.; Perbandt, M.; Betzel, C.; Singh, T.P. Crystal structures of the complexes of a group IIA phospholipase A2 with two natural anti-inflammatory agents, anisic acid, and atropine reveal a similar mode of binding. Proteins 2016, 64, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.M.; Ticli, F.K.; Marcussi, S.; Lourenço, M.V.; Januário, A.H.; Sampaio, S.V.; Giglio, J.R.; Lomonte, B.; Pereira, P.S. Medicinal plants with inhibitory properties against snake venoms. Curr. Med. Chem. 2005, 12, 2625–2641. [Google Scholar] [CrossRef]
- Jiménez-Estrada, M.; Velázquez-Contreras, C.; Garibay-Escobar, A.; Sierras-Canchola, D.; Lapizco-Vázquez, R.; Ortiz-Sandoval, C.; Burgos-Hernández, A.; Robles-Zepeda, R.E. In vitro antioxidant and antiproliferative activities of plants of the ethnopharmacopeia from northwest of Mexico. BMC Complement. Altern. Med. 2013, 13, 12. [Google Scholar] [CrossRef] [PubMed]
- Gupta, Y.K.; Peshin, S.S. Do herbal medicines have potential for managing snake bite envenomation? Toxicol. Int. 2012, 19, 89. [Google Scholar] [CrossRef] [PubMed]
- Molander, M.; Staerk, D.; Nielsen, H.M.; Brandner, J.N.; Diallo, D.; Zacharie, C.K.; van Staden, J.; Jäger, A.K. Investigation of skin permeation, ex vivo inhibition of venom-induced tissue destruction, and wound healing of African plants used against snakebites. J. Etnopharmacol. 2015, 165, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Aboul-Enein, H.Y.; Kruk, I.; Kładna, A.; Lichszteld, K.; Michalska, T. Scavenging effects of phenolic compounds on reactive oxygen species. Biopolymers 2007, 86, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Rohn, S.; Rawel, H.M.; Kröll, J. Inhibitory Effects of Plant Phenols on the Activity of Selected Enzymes. J. Agric. Food Chem. 2002, 50, 3566–3571. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.I.; Alam, M.A.; Alam, O.; Nargotra, A.; Taneja, S.C.; Koul, S. Molecular modeling and snake venom phospholipase A 2 inhibition by phenolic compounds: Structure–activity relationship. Eur. J. Med. Chem. 2016, 114, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Nargotra, A.; Sharma, S.; Alam, M.I.; Ahmed, Z.; Bhagat, A.; Taneja, S.C.; Qazi, G.N.; Koul, S. In silico identification of viper phospholipaseA2 inhibitors: validation by in vitro, in vivo studies. J. Mol. Model. 2011, 17, 3063–3073. [Google Scholar] [CrossRef] [PubMed]
- Pereañez, J.A.; Núñez, V.; Patiño, A.C.; Londono, M.; Quintana, J.C. Inhibitory effects of plants phenolic compounds on enzymatic and cytotoxic activities induced by a snake venom phospholipase A2. Vitae 2011, 18, 295–304. [Google Scholar]
- Das, K. Medicinal plants for snake bite treatment – Future focus. Ethnobot. Leaf 2009, 13, 14. [Google Scholar]
- Vishwanath, B.; Rao, A.A.; Gowda, T. Interaction of phospholipase A2 from Vipera russelli venom with aristolochic acid: A circular dichroism study. Toxicon 1987, 25, 939–946. [Google Scholar] [CrossRef]
- Chandra, V.; Jasti, J.; Kaur, P.; Srinivasan, A.; Betzel, C.; Singh, T.P. Structural Basis of Phospholipase A2 Inhibition for the Synthesis of Prostaglandins by the Plant Alkaloid Aristolochic Acid from a 1.7 Å Crystal Structure. Biochemistry 2002, 41, 10914–10919. [Google Scholar] [CrossRef] [PubMed]
- Girish, K.S.; Kemparaju, K. Inhibition of Naja naja venom hyaluronidase by plant-derived bioactive components and polysaccharides. Biochemistry (Moscow) 2005, 70, 948–952. [Google Scholar] [CrossRef]
- Kemparaju, K.; Girish, K.S. Snake venom hyaluronidase: a therapeutic target. Cell Biochem. Funct. 2006, 24, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J. Dietary flavonoid aglycones and their glycosides: ¿Which show better biological significance? Crit. Rev. Food Sci. Nutr. 2017, 57, 1874–1905. [Google Scholar] [CrossRef] [PubMed]
- Kuppusamy, U.R.; Das, N.P. Inhibitory effects of flavonoids on several venom hyaluronidases. Cell. Mol. Life Sci. 1991, 47, 1196–1200. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Betancur, I.; Benjumea, D.; Patiño, A.; Jimenez, N.; Osorio, E. Inhibition of the toxic effects of Bothrops asper venom by pinostrobin, a flavanone isolated from Renealmia alpinia (Rottb.) MAAS. J. Ethnopharmacol. 2014, 155, 1609–1615. [Google Scholar] [CrossRef] [PubMed]
- Preciado, L.M.; Comer, J.; Núñez, V.; Rey-Súarez, P.; Pereañez, J.A. Inhibition of a Snake Venom Metalloproteinase by the Flavonoid Myricetin. Molecules 2018, 23, 2662. [Google Scholar] [CrossRef] [PubMed]
- Gopi, K.; Anbarasu, K.; Renu, K.; Jayanthi, S.; Vishwanath, B.; Jayaraman, G. Quercetin-3-O-rhamnoside from Euphorbia hirta protects against snake Venom induced toxicity. Biochim. Biophys. Acta (BBA) - Gen. Subj. 2016, 1860, 1528–1540. [Google Scholar] [CrossRef] [PubMed]
- Sachetto, A.T.A.; Rosa, J.G.; Santoro, M.L. Rutin (quercetin-3-rutinoside) modulates the hemostatic disturbances and redox imbalance induced by Bothrops jararaca snake venom in mice. PLoS Neglected Trop. Dis. 2018, 12, e0006774. [Google Scholar] [CrossRef]
- Nakagawa, M.; Nakanishi, K.; Darko, L.L.; Vick, J.A. Structures of cabenegrins A-I and A-II, potent anti-snake venoms. Tetrahedron Lett. 1982, 23, 3855–3858. [Google Scholar] [CrossRef]
- Ximenes, R.M.; Alves, R.S.; Pereira, T.P.; Araújo, R.M.; Silveira, E.R.; Rabello, M.M.; Hernandes, M.Z.; Soares, V.C.G.; Bristot, D.; Pires, C.L.; et al. Harpalycin 2 inhibits the enzymatic and platelet aggregation activities of PrTX-III, a D49 phospholipase A2 from Bothrops pirajai venom. BMC Complement. Altern. Med. 2012, 12, 139. [Google Scholar] [CrossRef] [PubMed]
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Methods 2007, 3, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Domingos, T.F.S.; Vallim, M.A.; Cavalcanti, D.N.; Sanchez, E.F.; Teixeira, V.L.; Fuly, A.L. Effect of Diterpenes Isolated of the Marine Alga Canistrocarpus cervicornis against Some Toxic Effects of the Venom of the Bothrops jararaca Snake. Molecules 2015, 20, 3515–3526. [Google Scholar] [CrossRef] [PubMed]
- Muthusamy, K.; Chinnasamy, S.; Nagarajan, S.; Sivaraman, T.; Chinnasamy, S. Isolation and characterization of bioactive compounds of Clematis gouriana Roxb. ex DC against snake venom phospholipase A2 (PLA2) computational and in vitro insights. J. Biomol. Struct. Dyn. 2017, 35, 1936–1949. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, J.O.; Fernandes, R.S.; Ticli, F.K.; Oliveira, C.Z.; Mazzi, M.V.; Franco, J.J.; Giuliatti, S.; Pereira, P.S.; Soares, A.M.; Sampaio, S.V. Triterpenoid saponins, new metalloprotease snake venom inhibitors isolated from Pentaclethra macroloba. Toxicon 2007, 50, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Preciado, L.M.; Rey-Suárez, P.; Henao, I.C.; Pereañez, J.A. Betulinic, oleanolic and ursolic acids inhibit the enzymatic and biological effects induced by a P-I snake venom metalloproteinase. Chem. Interactions 2018, 279, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Moura, L.D.A.; Sanchez, E.F.; Bianco, É.M.; Pereira, R.C.; Teixeira, V.L.; Fuly, A.L. Antiophidian properties of a dolastane diterpene isolated from the marine brown alga Canistrocarpus cervicornis. Biomed. Prev. Nutrit. 2011, 1, 61–66. [Google Scholar] [CrossRef]
- Dharmappa, K.K.; Kumar, R.V.; Nataraju, A.; Pereira, R.C.; Teixeira, V.L.; Fuly, A.L. Anti-inflammatory activity of oleanolic acid by inhibition of secretory phospholipase A2. Planta Med. 2009, 75, 211–215. [Google Scholar] [CrossRef]
- Batina, M.D.F.C.; Cintra, A.C.O.; Veronese, E.L.G.; Lavrador, M.A.S.; Giglio, J.R.; Pereira, P.S.; Dias, D.A.; França, S.C.; Sampaio, S.V. Inhibition of the Lethal and Myotoxic Activities of Crotalus durissus terrificus Venom by Tabernaemontana catharinensis: Identification of One of the Active Components. Planta Med. 2000, 66, 424–428. [Google Scholar] [CrossRef]
- Menecucci, C.D.S.; Mucellini, K.L.; De Oliveira, M.M.; Higashi, B.; De Almeida, R.T.R.; Porto, C.; Pilau, E.J.; Gonçalves, J.E.; Gonçalves, R.A.C.; De Oliveira, A.J.B. Latex from Tabernaemontana catharinensis (A. DC)—Apocynaceae: An alternative for the sustainable production of biologically active compounds. Ind. Crop. Prod. 2019, 129, 74–84. [Google Scholar] [CrossRef]
- Samy, R.P.; Gopalakrishnakone, P.; Chow, V.T. Therapeutic application of natural inhibitors against snake venom phospholipase A2. Bioinformation 2012, 8, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, P.; Bhattacharyya, D. Characterization of the aqueous extract of the root of Aristolochia indica: Evaluation of its traditional use as an antidote for snake bites. J. Ethnopharmacol. 2013, 145, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, P.; Bera, I.; Chakraborty, S.; Ghoshal, N.; Bhattacharyya, D. Aristolochic acid and its derivatives as inhibitors of snake venom L -amino acid oxidase. Toxicon 2017, 138, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.A.H.; Cardoso, F.F.; Cavalcante, W.G.L.; Soares, A.M.; Dal-Pai, M.; Gallacci, M.; Fontes, M.R.M. Structural Basis for the Inhibition of a Phospholipase A2-Like Toxin by Caffeic and Aristolochic Acids. PLoS ONE 2015, 10, e0133370. [Google Scholar] [CrossRef] [PubMed]
- Ticli, F.K.; Hage, L.I.; Cambraia, R.S.; Pereira, P.S.; Magro, A.J.; Fontes, M.R.; Stabeli, R.G.; Giglio, J.R.; França, S.C.; Soares, A.M.; et al. Rosmarinic acid, a new snake venom phospholipase A2 inhibitor from Cordia verbenacea (Boraginaceae): antiserum action potentiation and molecular interaction. Toxicon 2005, 46, 318–327. [Google Scholar] [CrossRef] [PubMed]
- DosSantos, J.I.; Cardoso, F.F.; Soares, A.M.; dal Pai Silva, M.; Gallacci, M.; Fontes, M.R.M. Structural and functional studies of a bothropic myotoxin complexed to rosmarinic acid: new insights into Lys49-PLA₂ inhibition. PLoS ONE 2011, 6, e28521. [Google Scholar]
- Aung, H.T.; Nikai, T.; Niwa, M.; Takaya, Y. Rosmarinic acid in Argusia argentea inhibits snake venom-induced hemorrhage. J. Nat. Med. 2010, 64, 482–486. [Google Scholar] [CrossRef]
- Aung, H.T.; Furukawa, T.; Nikai, T.; Niwa, M.; Takaya, Y. Contribution of cinnamic acid analogues in rosmarinic acid to inhibition of snake venom induced hemorrhage. Bioorganic Med. Chem. 2011, 19, 2392–2396. [Google Scholar] [CrossRef]
- Gopi, K.; Renu, K.; Vishwanath, B.S.; Jayaraman, G. Protective effect of Euphorbia hirta and its components against snake venom induced lethality. J. Ethnopharmacol. 2015, 165, 180–190. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Doley, R.; Saikia, D. Isolation of a snake venom phospholipase A2 (PLA2) inhibitor (AIPLAI) from leaves of Azadirachta indica (Neem): Mechanism of PLA2 inhibition by AIPLAI in vitro condition. Toxicon 2008, 51, 1548–1553. [Google Scholar] [CrossRef]
- Alam, M.I.; Auddy, B.; Gomes, A. Viper venom neutralization by Indian medicinal plant (Hemidesmus indicus and Pluchea indica) root extracts. Phytother. Res. 1996, 10, 58–61. [Google Scholar] [CrossRef]
- Tomaz, M.A.; Patrão-Neto, F.C.; Melo, P.A.; Gopalakrishnakone, P.; Carlini, C.R.; Ligabue-Braun, R. Plant Compounds with Antiophidic Activities, Their Discovery History, and Current and Proposed Applications. Toxinology 2016, 1–16. [Google Scholar] [CrossRef]
- Strauch, M.A.; Tomaz, M.A.; Monteiro-Machado, M.; Ricardo, H.D.; Cons, B.L.; Fernandes, F.F.; El-Kik, C.Z.; Azevedo, M.S.; Melo, P.A. Antiophidic activity of the extract of the Amazon plant Humirianthera ampla and constituents. J. Ethnopharmacol. 2013, 145, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Li, B.; Huang, D.; He, Q.; Yu, X. Anti- Deinagkistrodon acutus venom properties of ethanolic root extract from Cynanchum paniculatum (Bunge) kitag and its GC-MS analysis. J. Ethnopharmacol. 2018, 225, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Melo, P.A.; Pinheiro, D.A.; Ricardo, H.D.; Fernandes, F.F.; Tomaz, M.A.; El-Kik, C.Z.; Strauch, M.A.; Da Fonseca, T.F.; Sifuentes, D.N.; Calil-Eliás, S.; et al. Ability of a synthetic coumestan to antagonize Bothrops snake venom activities. Toxicon 2010, 55, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Januário, A.H.; Santos, S.L.; Marcussi, S.; Mazzi, M.V.; Pietro, R.C.; Sato, D.N.; Ellena, J.; Sampaio, S.V.; França, S.C.; Soares, A.M. Neo-clerodane diterpenoid, a new metalloprotease snake venom inhibitor from Baccharis trimera (Asteraceae): anti-proteolytic and anti-hemorrhagic properties. Chem. Interactions 2004, 150, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, S.L.; Calgarotto, A.K.; Chaar, J.S.; Marangoni, S. Isolation and characterization of ellagic acid derivatives isolated from Casearia sylvestris SW aqueous extract with anti-PLA2 activity. Toxicon 2008, 52, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Esteves, I.; Lima, L.M.; Santos, L.S.; Rodrigues, M.; da Silva, J.M.S.; Perazzo, F.F.; Carvalho, J.C.T. Casearia sylvestris SW essential oil activity in inflammation in rats induced by Bothrops alternatus venom. Int. J. Pharm. Sci. Rev. Res. 2011, 7, 5. [Google Scholar]
- Ode, O.; Asuzu, I. The anti-snake venom activities of the methanolic extract of the bulb of Crinum jagus (Amaryllidaceae). Toxicon 2006, 48, 331–342. [Google Scholar] [CrossRef]
- Shukla, P.K.; Gautam, L.; Sinha, M.; Kaur, P.; Sharma, S.; Singh, T.P. Structures and binding studies of the complexes of phospholipase A2 with five inhibitors. Biochim. Biophys. Acta (BBA) - Proteins Proteom. 2015, 1854, 269–277. [Google Scholar] [CrossRef]
- Esmeraldino, L.E.; Souza, A.M.; Sampaio, S.V. Evaluation of the effect of aqueous extract of Croton urucurana Bailon (Euphorbiaceae) on the hemorrhagic activity induced by the venom of Bothrops jararaca, using new techniques to quantify hemorrhagic activity in rat skin. Phytomedicine 2005, 12, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Passos, G.F.; Fernandes, E.S.; Da Cunha, F.M.; Ferreira, J.; Pianowski, L.F.; Campos, M.M.; Calixto, J.B. Anti-inflammatory and anti-allergic properties of the essential oil and active compounds from Cordia verbenacea. J. Ethnopharmacol. 2007, 110, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, E.S.; Passos, G.F.; Medeiros, R.; Da Cunha, F.M.; Ferreira, J.; Campos, M.M.; Pianowski, L.F.; Calixto, J.B. Anti-inflammatory effects of compounds alpha-humulene and (−)-trans-caryophyllene isolated from the essential oil of Cordia verbenacea. Eur. J. Pharmacol. 2007, 569, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Azofeifa, K.; Angulo, Y.; Lomonte, B. Ability of fucoidan to prevent muscle necrosis induced by snake venom myotoxins: Comparison of high- and low-molecular weight fractions. Toxicon 2008, 51, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Geyer, B.; Kiso, Y.; Hikino, H.; Rao, G. Coumestans as the Main Active Principles of the Liver Drugs Eclipta alba and Wedelia calendulacea. Planta Med. 1986, 52, 370–374. [Google Scholar] [CrossRef]
- Saxena, A.; Singh, B.; Anand, K. Hepatoprotective effects of Eclipta alba on subcellular levels in rats. J. Ethnopharmacol. 1993, 40, 155–161. [Google Scholar] [CrossRef]
- Diogo, L.C.; Fernandes, R.S.; Marcussi, S.; Menaldo, D.L.; Roberto, P.G.; Matrangulo, P.V.F.; Pereira, P.S.; França, S.C.; Giuliatti, S.; Soares, A.M.; et al. Inhibition of Snake Venoms and Phospholipases A2 by Extracts from Native and Genetically Modified Eclipta alba: Isolation of Active Coumestans. Basic Clin. Pharmacol. Toxicol. 2009, 104, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Pithayanukul, P.; Laovachirasuwan, S.; Bavovada, R.; Pakmanee, N.; Suttisri, R. Anti-venom potential of butanolic extract of Eclipta prostrata against Malayan pit viper venom. J. Ethnopharmacol. 2004, 90, 347–352. [Google Scholar] [CrossRef]
- Alam, M.; Auddy, B.; Gomes, A. Isolation, purification and partial characterization of viper venom inhibiting factor from the root extract of the Indian medicinal plant sarsaparilla (Hemidesmus indicus R.Br.). Toxicon 1994, 32, 1551–1557. [Google Scholar] [CrossRef]
- Borges, M.; Alves, D.; Raslan, D.; Piló-Veloso, D.; Rodrigues, V.; Homsi-Brandeburgo, M.; De Lima, M. Neutralizing properties of Musa paradisiaca L. (Musaceae) juice on phospholipase A2, myotoxic, hemorrhagic and lethal activities of crotalidae venoms. J. Ethnopharmacol. 2005, 98, 21–29. [Google Scholar] [CrossRef]
- Min, H.M.; Aye, M.; Taniguchi, T.; Miura, N.; Monde, K.; Ohzawa, K.; Nikai, T.; Niwa, M.; Takaya, Y. A structure and an absolute configuration of (+)-alternamin, a new coumarin from Murraya alternans having antidote activity against snake venom. Tetrahedron Lett. 2007, 48, 6155–6158. [Google Scholar] [CrossRef]
- Torres, M.C.M.; Pinto, F.D.C.L.; Filho, R.B.; Silveira, E.R.; Pessoa, O.D.L.; Jorge, R.J.B.; Ximenes, R.M.; Monteiro, H.S.A.; Evangelista, J.S.A.M.; Diz-Filho, E.B.S.; et al. Antiophidic Solanidane Steroidal Alkaloids from Solanum campaniforme. J. Nat. Prod. 2011, 74, 2168–2173. [Google Scholar] [CrossRef] [PubMed]
- Machiah, D.K.; Girish, K.; Gowda, T.V. A glycoprotein from a folk medicinal plant, Withania somnifera, inhibits hyaluronidase activity of snake venoms. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 143, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.E.A.S.; Soares, J.M.D.; Silva, H.A.L.; de Souza Wanderley, C.W.; Moura, C.A.; de Oliveira-Junior, R.G.; de Oliveira, A.P.; Rolim, L.A.; Costa, E.V.; da Silva Almeida, J.R.G.; et al. Inhibitory effects of Morus nigra L. (Moraceae) against local paw edema and mechanical hypernociception induced by Bothrops jararacussu snake venom in mice. Biomed Pharmacother. 2019, 111, 1046–4056. [Google Scholar] [CrossRef] [PubMed]
- Toyama, M.H.; Toyama, D.O.; Torres, V.M.; Pontes, G.C.; Farias, W.R.L.; Melo, F.R.; Oliveira, S.C.B.; Fagundes, F.H.R.; Diz Filho, E.B.S.; Cavada, B.S. Effects of low molecular weight sulfated galactan fragments from Botryocladia occidentalis on the pharmacological and enzymatic activity of sPLA2 from Crotalus durissus cascavella. Protein J. 2010, 29, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.; Mong, S.; Clarke, M.A.; Kruse, L.I.; Crooke, S.T. Differential effects of manoalide on secreted and intracellular phospholipases. Biochem. Pharmacol. 1987, 36, 733–740. [Google Scholar] [CrossRef]
- Rocha, S.L.G.; Lomonte, B.; Neves-Ferreira, A.G.C.; Trugilho, M.R.O.; Junqueira-De-Azevedo, I.D.L.M.; Ho, P.L.; Domont, G.B.; Gutiérrez, J.M.; Perales, J.; Neves-Ferreira, A.G.C.; et al. Functional analysis of DM64, an antimyotoxic protein with immunoglobulin-like structure from Didelphis marsupialis serum. Eur. J. Biochem. 2002, 269, 6052–6062. [Google Scholar] [CrossRef]
- Pan, S.Y.; Chen, S.B.; Dong, H.G.; Yu, Z.L.; Dong, J.C.; Long, Z.X.; Fong, W.F.; Han, Y.F.; Ko, K.M. New perspectives on Chinese herbal medicine (Zhong-Yao) research and development. Evid-Based Complement. Alternat. Med. 2011, 403709. [Google Scholar] [CrossRef]
- Poupko, J.M.; Baskin, S.I.; Moore, E. The pharmacological properties of anisodamine. J. Appl. Toxicol. 2007, 27, 116–121. [Google Scholar] [CrossRef]
- Eisenbrand, G.; Hippe, F.; Jakobs, S.; Muehlbeyer, S. Molecular mechanisms of indirubin and its derivatives: novel anticancer molecules with their origin in traditional Chinese phytomedicine. J. Cancer Res. Clin. Oncol. 2004, 130, 627–635. [Google Scholar] [CrossRef]
- Desilets, A.R.; Gickas, J.J.; Dunican, K.C. Role of huperzine A in the treatment of Alzheimer’s disease. Ann. Pharmacother. 2009, 43, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Xiang, D.-R.; Sheng, J.-F. The analyse of effectiveness in HBeAg-positive chronic viral hepatitis B treated by adefovir dipivoxil combined with bicyclol. Zhonghua shi yan he lin chuang bing du xue za zhi = Zhonghua shiyan he linchuang bingduxue zazhi = Chin. J. Exp. Clin. Virol. 2009, 23, 299–301. [Google Scholar]
- Laustsen, A.H.; Dorrestijn, N. Integrating Engineering, Manufacturing, and Regulatory Considerations in the Development of Novel Antivenoms. Toxins 2018, 10, 309. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.J.; Gutiérrez, J.-M.; Calvete, J.J.; Wüster, W.; Ratanabanangkoon, K.; Paiva, O.; Brown, N.I.; Casewell, N.R.; Harrison, R.A.; Rowley, P.D.; et al. Ending the drought: New strategies for improving the flow of affordable, effective antivenoms in Asia and Africa. J. Proteom. 2011, 74, 1735–1767. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Lomonte, B.; León, G.; Alape-Girón, A.; Flores-Díaz, M.; Sanz, L.; Angulo, Y.; Calvete, J.J. Snake venomics and antivenomics: Proteomic tools in the design and control of antivenoms for the treatment of snakebite envenoming. J. Proteom. 2009, 72, 165–182. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Fasoli, E.; Sanz, L.; Boschetti, E.; Righetti, P.G. Exploring the Venom Proteome of the Western Diamondback Rattlesnake, Crotalus atrox, via Snake Venomics and Combinatorial Peptide Ligand Library Approaches. J. Proteome Res. 2009, 8, 3055–3067. [Google Scholar] [CrossRef] [PubMed]
- Fasoli, E.; Sanz, L.; Wagstaff, S.; Harrison, R.A.; Righetti, P.G.; Calvete, J.J. Exploring the venom proteome of the African puff adder, Bitis arietans, using a combinatorial peptide ligand library approach at different pHs. J. Proteom. 2010, 73, 932–942. [Google Scholar] [CrossRef] [PubMed]
- Righetti, P.G.; Boschetti, E.; Zanella, A.; Fasoli, E.; Citterio, A. Plucking, pillaging and plundering proteomes with combinatorial peptide ligand libraries. J. Chromatogr. A 2010, 1217, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Arnold, C. Synthetic Biology Tackles Antivenom. Nature 2016, 532, 292. [Google Scholar] [CrossRef][Green Version]
- O’Brien, J.; Lee, S.H.; Onogi, S.; Shea, K.J. Engineering the protein porona of a synthetic polymer nanoparticle for broad-Spectrum Sequestration and Neutralization of Venomous Biomacromolecules. J. Am. Chem. Soc. 2016, 138, 16604–16607. [Google Scholar] [CrossRef]
- Waheed, H.; Moin, S.F.; Choudhary, M.I. Snake Venom: From Deadly Toxins to Life-saving Therapeutics. Curr. Med. Chem. 2017, 24, 1874–1891. [Google Scholar] [CrossRef] [PubMed]
- Hashemzadeh, M.; Furukawa, M.; Goldsberry, S.; Movahed, M.R. Chemical structures and mode of action of intravenous glycoprotein IIb/IIIa receptor blockers: A review. Exp. Clin. Cardiol. 2008, 13, 192–197. [Google Scholar] [PubMed]
- Ondetti, M.A.; Williams, N.J.; Sabo, E.; Pluscec, J.; Weaver, E.R.; Kocy, O. Angiotensin-converting enzyme inhibitors from the venom of Bothrops jararaca. Isolation, elucidation of structure, and synthesis. Biochemistry 1971, 10, 4033–4039. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.A.; Wickremasinghe, N.C.; Shi, S.; Hartgerink, J.D. Nanofibrous snake venom hemostat. ACS Biomater. Sci. Eng. 2015, 1, 1300–1305. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.S.; Cheung, R.C.F.; Xia, L.; Wong, J.H.; Ng, T.B. Snake venom toxins: toxicity and medicinal applications. Appl. Microbiol. Biotechnol. 2016, 100, 6165–6181. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Plant | Description of the Study | Reference |
|---|---|---|---|
| India | Morus alba (Moraceae) | Extracts from Morus alba (Moraceae) are active against Daboia russelli venom, inhibiting the caseinolytic, hyaluronolytic, edematogenic, hemorrhagic, and procoagulant activities. | [47] |
| Nigeria | Mucuna pruriens (Fabaceae) | Seed extract of Mucuna pruriens (Fabaceae) used in Nigerian communities offer significant protection to cardiac muscle tissue and blood vessels, and even protects against the lethality produced by venoms from Naja kaouthia, Naja nivea, and Calloselasma rhodostoma. This protection can be explained from the presence of a Kunitz-type trypsin inhibitor. | [48,49,50] |
| Yemen | Hibiscus aethiopicus (Malvaceae) | Aqueous crude extracts from Hibiscus aethiopicus (Malvaceae) possess significant anti-hemorrhagic and cytoprotective activities against Echis ocellatus and Naja n. nigricollis venoms | [51] |
| India | Vitis vinifera (Vitaceae) | Administration of the methanolic extracts from Vitis vinifera (Vitaceae) resulted in the reduction of local symptoms produced by D. russelli venom due to the inhibition of the proteolytic and hyaluronidase activities reducing edema, myonecrosis, and hemorrhaging. | [52] |
| Brazil | Dipteryx alata (Fabaceae) | Extracts and fractions from Dipteryx alata (Fabaceae) partially neutralized Bothrops jararacussu and Crotalus durissus terrificus venom activities. Hydroalcoholic bark extract from D. alata is active against B. jararacussu venom. | [53,54] |
| Brazil | Marsypianthes chamaedrys (Lamiaceae) | Infusions and crushed leaves from Marsypianthes chamaedrys (Lamiaceae) showed a similar activity produced by antivenom serum against clotting and inflammatory effects of the Bothrops atrox venom | [55] |
| Brazil | Hypericum brasiliense (Guttiferae) | Hypericum brasiliense (Guttiferae) reduces the lethality produced by Bothrops jararaca by inhibiting the edematous and proteolytic activities of the venom. | [56] |
| India | Luffa egyptiaca (Cucurbitaceae) and Nicotiana rustica (Solanaceae) | Ethyl acetate fractions of Luffa egyptiaca (Cucurbitaceae) and Nicotiana rustica (Solanaceae) extracts completely inhibited the protease activity of Naja nigricollis venom. | [57] |
| India | Mangifera indica (Anacardiaceae) | Aqueous extracts from the stem bark of Mangifera indica (Anacardiaceae) inhibited the protease, hyaluronidase, hemorrhagic, fibrinogenolytic, hemolytic, procoagulant, edema, ATPase, and alkaline phosphatase activities produced by D. russelli venom | [58] |
| Argentina | Nectandra angustifolia (Lauraceae) | Ethanolic extracts and essential oils from Nectandra angustifolia (Lauraceae) leaves inhibited the hemolytic and coagulant effects produced by Bothrops neuwiedi venom | [59] |
| Pakistan | Fagonia cretica (Zygophyllaceae) | Methanolic extract from leaves and twigs of Fagonia cretica (Zygophyllaceae) is capable of inhibiting hemorrhage induced by Naja naja karachiensis venom | [60] |
| Brazil | Sapindus saponaria (Sapindaceae) | Fractions of the hydro alcoholic extracts from the callus of Sapindus saponaria (Sapindaceae) partially inhibited the lethality, phospholipase, clotting, edema, hemorrhagic, and myotoxic activities produced by Bothrops jararacussu, Bothrops moojeni, Bothrops alternates, and Crotalus durissus terrificus venoms along with isolated myotoxins and phospholipase A2 (PLA2) | [61] |
| Brazil | Mouriri pusa (Melastomataceae), Byrsonima crassa (Malpighiaceae), and Davilla elliptica (Dilleniaceae) | Methanolic extracts from Mouriri pusa (Melastomataceae), Byrsonima crassa (Malpighiaceae), and Davilla elliptica (Dilleniaceae) blocked local hemorrhages produced by Bothrops jararaca venom. | [62] |
| India | Anacardium occidentale | The efficacy of Anacardium occidentale extract against pharmacological actions induced by Vipera russelli venom was observed from the neutralization of phospholipases, proteases, and hyaluronidases, as well as edema, hemorrhage, lethality, and myonecrosis effects | [63] |
| India | Tamarindus indica (Fabaceae) | Tamarindus indica (Fabaceae) inhibited hyaluronidase, Phospholipase A2 (PLA2), l-amino oxidase (LAAO), and 5′-nucleotidase. Exhibited fibrinogenolytic, edema-inducing, hemorrhagic, indirect hemolytic, coagulant, and myotoxic properties, and protected against venom-toxicity | [64] |
| Brazil | 83 plant species, from 34 families | Inhibition of Phospholipase A2 (PLA2), anti edema, anti lethality, anti clotting, myotoxicity, and antihaemorrhagic activity | [65] |
| Brazil | Mandevilla velutina and Eclipta prostata | Inhibition of creatine kinase release and myotoxic activity | [66] |
| Worldwide | Schumanniophyton magnificum, Aristolochia radix, Diospyros kaki, Alocasia cucullata, Picrasma quassioides, Eclipta prostrata, Curcuma sp., Soja hispida, Diodia scandens, Andrographis paniculata | Inhibition of Phospholipase A2 (PLA2) or other enzymes (Adenosine triphosphatese), and life-prolongation effect post black mamba venom treatment. | [67] |
| Colombia | 77 plant species | Three relevant studies: First study was an inventory with 77 species of plants belonging to 41 families used by Colombian healers along with the methods of preparation, administration, and dosage; second study was a list of 74 ethanolic plant extracts used by folk medicinethat were active against lethal effects produced by Bothrops atrox venom; third study showed 31 extracts with moderate or high neutralizing abilities against the hemorrhagic effect of B. atrox venom | [68] |
| Brazil | Numerous plant species | Review discussing Brazilian plant species displaying neutralizing properties against snake envenomation from an ethnopharmacological perspective | [69] |
| Costa Rica | 40 plant species | 40 plant species belonging to at least 30 families. Neutralization activity of Costa Rican plants towards B. asper venom and toxins | [70] |
| India | 34 plant species | A list of 34 plant species belonging to the Zingiberaceae family traditionally used in Northeast India, where one species presented antivenom activity and five other species have been scientifically validated to be anti-inflammatory | [71] |
| Nepal, China, South Africa, Nicaragua, and Brazil | 310 plant genera from 171 families | Enzyme inhibition activity | [72] |
| India | 69 plant species | 69 plant species belonging to 29 genera and 17 compounds with antiophidian activity or relative properties against venoms from 34 snake species | [73] |
| Nigeria and Ghana | Schumanniophyton magnificum, Strophanthus gratus, Strophanthus hispidus, Mucuna pruriens | Aqueous extracts showed effects on the blood clotting against Echis carinatus envenomation | [74] |
| Mali, DR Congo, South Africa | 94 species of 84 genera | List of plants used traditionally in sub-Saharan Africa. Hyaluronidase, phospholipase A2, and protease inhibitory activity against effects produced by Bitis arietans, and Naja nigricollis venom | [75] |
| Compound | Source | Mechanism of Action | Mode of Administration/Study Level | Reference |
|---|---|---|---|---|
| Aristolochic acids | Aristolochia indica Aristolochia sp. Aristolochia odoratissima Aristolochia fordiana Aristolochia radix | Induction of PLA2 Inhibition of L-amino acid oxidase Anti-lethality | Injected into the mouse foot pad | [96,97,98,99,118,119,120,121] |
| Rosmarinic acid | Cordia verbenacea Argusia argentea | Inhibition of myotoxic activity and PLA2 Anti-hemorrhagic activity | Injected intramuscularly into the right gastrocnemius muscle of mice In vivo study | [122,123,124,125] |
| Quercetin-3-O-α-L-rhamnopyranoside quinic acid, gallic acid, quercetin, kaempferol, luteolin, ellagic acid, chlorogenic acid | Euphorbia hirta | Inhibition of PLA2 | In vitro study | [105,126] |
| Pinostrobin | Renealmia alpinia | Inhibition of myotoxic activity and PLA2 | Intramuscular injection (inhibition of myotoxic activity) and subcutaneous injection (inhibition of edema-inducing activity) | [81] |
| Undisclosed | Azadirachta indica | Inhibition of PLA2 | In vitro study | [127] |
| 2-hydroxy-4-methoxy benzoic acid | Hemidesmus indicus | Neutralization of venom hemorrhagic activity | Injected intradermally into mice | [128] |
| β-sitosterol | Eclipta prostrata, Humirianthera ampla Cynanchum paniculatum | Neutralization of enzymes | In vitro and in vivo studies | [129,130,131] |
| 8-methoxy coumestrol | Medicago sativa | Inhibition of edema, hemorrhage, and cardio toxicity | Injected intravenously into mice | [132] |
| 7α-hydroxy-3,13-clerodadiene-16,15:18,19-diolides | Baccharis trimera | Inhibition of metalloproteases | In vitro and in vivo studies | [133] |
| CAS # 1260387-36-7 CAS # 2242794-02-9 | Clematis gouriana | Inhibition of PLA2 | In vitro and computational studies | [111] |
| Linearol Isolinearol CAS # 88452-92-0 | Canistrocarpus cervicornis | Inhibition of induced hemorrhage, hemolysis, and coagulation | Biological assays | [110,114] |
| Ellagic acid | Casearia silvestris | Anti-myotoxic and anti-edema | In vivo study | [134,135] |
| Resveratrol | Crinum jagus | Neutralization of PLA2, protease, hyaluronidase, L-amino acid oxidase, and 5′-nucleotidase enzyme activities. Anti-myonecrosis and anti-hemorrhagic | Oral and intraperitoneal administration. In vivo study | [136,137] |
| Campesterol, β-sitosterol (its glycoside), stigmasterol, catechin, and gallocatechin | Croton urucurana | Anti-hemorrhagic and anti-lethality | In vivo study | [138] |
| Rosmarinic acid α-Humulene (−)-trans-Caryophyllene α-Pinene and alloaromadendrene | Cordia verbenacea | Anti-inflammatory, anti-myotoxic, anti-edematogenic, and anti-PLA2 activity. Anti-edematogenic and reduction of tumor necrosis factor-α (TNFα) Reduction of platelet activating factor, bradykinin, and anti-edematogenic. Anti-inflammatory and anti-edematogenic | In vivo study | [87,122,139,140] |
| Fucoidan | Fucus vesiculosus | Anti-myotoxic activity, anti-PLA2, and anti-necrosis | In vivo study | [141] |
| Wedelolactone | Eclipta alba | Anti-hepatotoxic, anti-hypertensive, anti-tumor, anti-PLA2, anti-snake venom, and anti-myotoxic-induced PLA2. | In vivo study | [142,143,144] |
| Wedelolactone, sitosterol, and stigmasterol | Eclipta prostrata | Anti-neurotoxic and anti-myotoxic | In vivo study | [145] |
| 2-hydroxy-4-methoxy-benzoic acid and lupeol acetate | Hemidesmus indicus | Anti-defibrinogenatic, anti-edematogenic, anti-PLA2 activity, anti-necrotic, anti-hemorrhagic, anti-coagulant, lipid peroxidase inhibition, superoxide dismutase activity, antiserum action potentiation, anti-lethality, anti-cardiotoxic, and anti-neurotoxic | In vivo study | [78,79,128,146] |
| Pentagalloyl glucopyranose | Mangifera indica | Anti-hemorrhagic, anti-dermonecrotic, and enzymatic activities. Inhibition of protease, hyalunoridase, fibrinogenolytic, procoagulant, anti-edematogenic, anti-ATPase, and alkaline phosphatase | In vivo study | [60,83] |
| Gallic acid | Musa paradisiaca | Anti-PLA2, anti-myotoxic, anti-hemorrhagic, and anti-lethality | In vivo study | [109,147] |
| Alternamin | Murraya alternans | Anti-hemorrhagic | In vivo study | [148] |
| Macrolobins A and B | Pentaclethra macroloba | Anti-proteolytic and anti-hemorrhagic, metalloprotease inhibitors | In vitro and in vivo studies | [112] |
| 4-Nerolidyl-catechol | Piper umbellatum and Piper peltatum | Anti-myotoxic, anti-PLA2, anti-serineprotease, and anti-edematogenic | In vivo study | [42] |
| Solanidane | Solanum campaniforme | Hemorrhagic inhibitor, necrotizing, and myotoxicity effects | In vivo study | [149] |
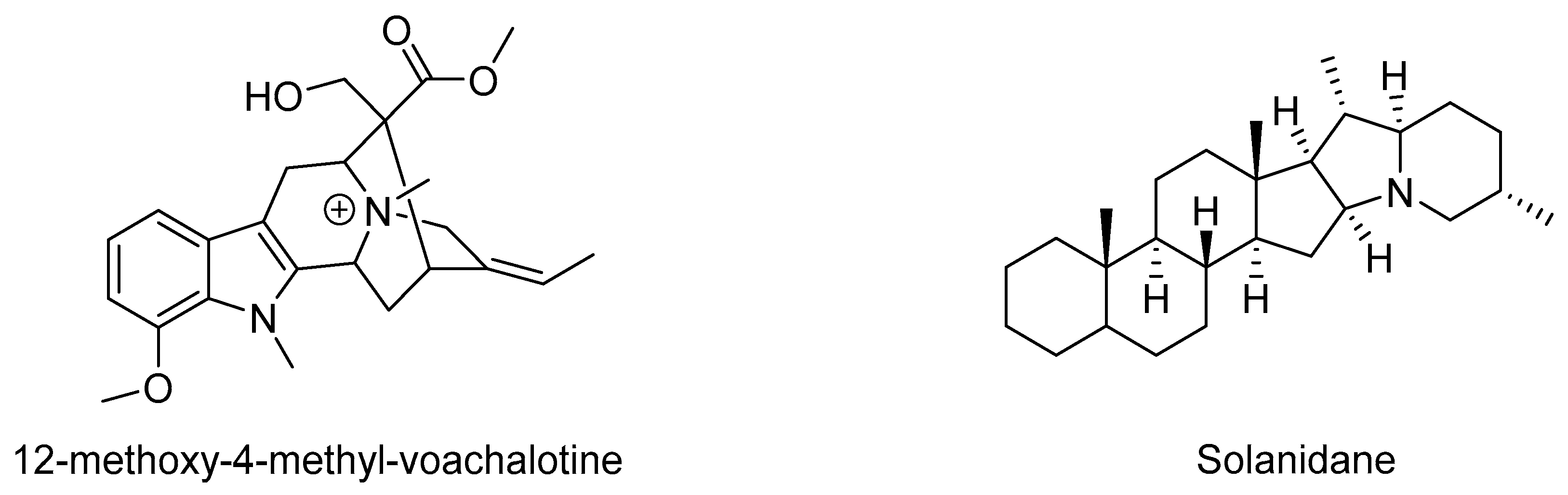
| 12-methoxy-4-methyl-voachalotine | Tabernaemontana catharinensis | Inhibited lethality | In vivo study | [116,117] |
| Whitania Somnifera Glycoprotein (WSG) | Withania somnifera | Anti-edematogenic | In vivo study | [150] |
| β-sitosterol, quercetin-3-O-glucopyranoside, and kaempferol-3-O-glucopyranoside | Morus nigra | Anti-inflammatory and antinociceptive effects | In vivo study | [151] |
| 2-hydroxy-3-methoxy benzaldehyde | Janakia arayalpathra | Anti-PLA2 | In vitro study | [92] |
| Marmin | Aegle marmelos | Anti-snake venom | In vivo study | [85] |
| Cabenegrins AI and A-II | Annona coriaceae | Anti-snake venom | In vitro and in vivo studies | [107] |
| Boc-5 and Boc-10 (sulfated galactans) | Botryocladia occidentalis | Anti-edematogenic, anti-myotoxic, and anti-neurotoxic | In vivo study | [152] |
| Edunol | Brongniartia podalyrioides | Protective effect against Bothrops atrox venom | In vivo study | [45] |
| Dolastane | Canistrocarpus cervicornis | Inhibition of induced hemorrhaging, hemolysis, and coagulation | In vitro and in vivo studies | [116] |
| Manoalide | Luffariella variabilis | Inhibition of extracellular PLA2 activity of cobras | In vitro study | [153] |
| DM64 (acidic glycoprotein) | Didelphis marsupialis | PLA2 inhibitor and prevention of myofiber breakdown caused by myotoxins I (Asp49) and II (Lys49) of B. asper venom | In vitro and in vivo studies | [154] |
| Aristolochic acid (8-methoxy-6-nitrophenanthro(3,4-d)-1,3-dioxole-5-carboxylic acid) Caffeic acid (3-(3,4-dihydroxyphenyl)-2-propenoic acid) | Aristolochia species Vernonia condensate | Inhibition of piratoxin-1 (PrTX-1), a Lys49-PLA2 isolated from Bothrops pirajai venom Inhibition of PrTX-1 and antidote activity against B. jararaca venom | Oral or parenteral administration In vitro and in vivo studies | [85,119] [121] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Betancur, I.; Gogineni, V.; Salazar-Ospina, A.; León, F. Perspective on the Therapeutics of Anti-Snake Venom. Molecules 2019, 24, 3276. https://doi.org/10.3390/molecules24183276
Gómez-Betancur I, Gogineni V, Salazar-Ospina A, León F. Perspective on the Therapeutics of Anti-Snake Venom. Molecules. 2019; 24(18):3276. https://doi.org/10.3390/molecules24183276
Chicago/Turabian StyleGómez-Betancur, Isabel, Vedanjali Gogineni, Andrea Salazar-Ospina, and Francisco León. 2019. "Perspective on the Therapeutics of Anti-Snake Venom" Molecules 24, no. 18: 3276. https://doi.org/10.3390/molecules24183276
APA StyleGómez-Betancur, I., Gogineni, V., Salazar-Ospina, A., & León, F. (2019). Perspective on the Therapeutics of Anti-Snake Venom. Molecules, 24(18), 3276. https://doi.org/10.3390/molecules24183276