Anti-Candida Biofilm Activity of Pterostilbene or Crude Extract from Non-Fermented Grape Pomace Entrapped in Biopolymeric Nanoparticles






Abstract

1. Introduction
2. Results
2.1. Physicochemical Characterization of PLGA NPs Loaded With PTB and Pomace Extracts
2.2. Chemical Characterization of Pomace Extracts
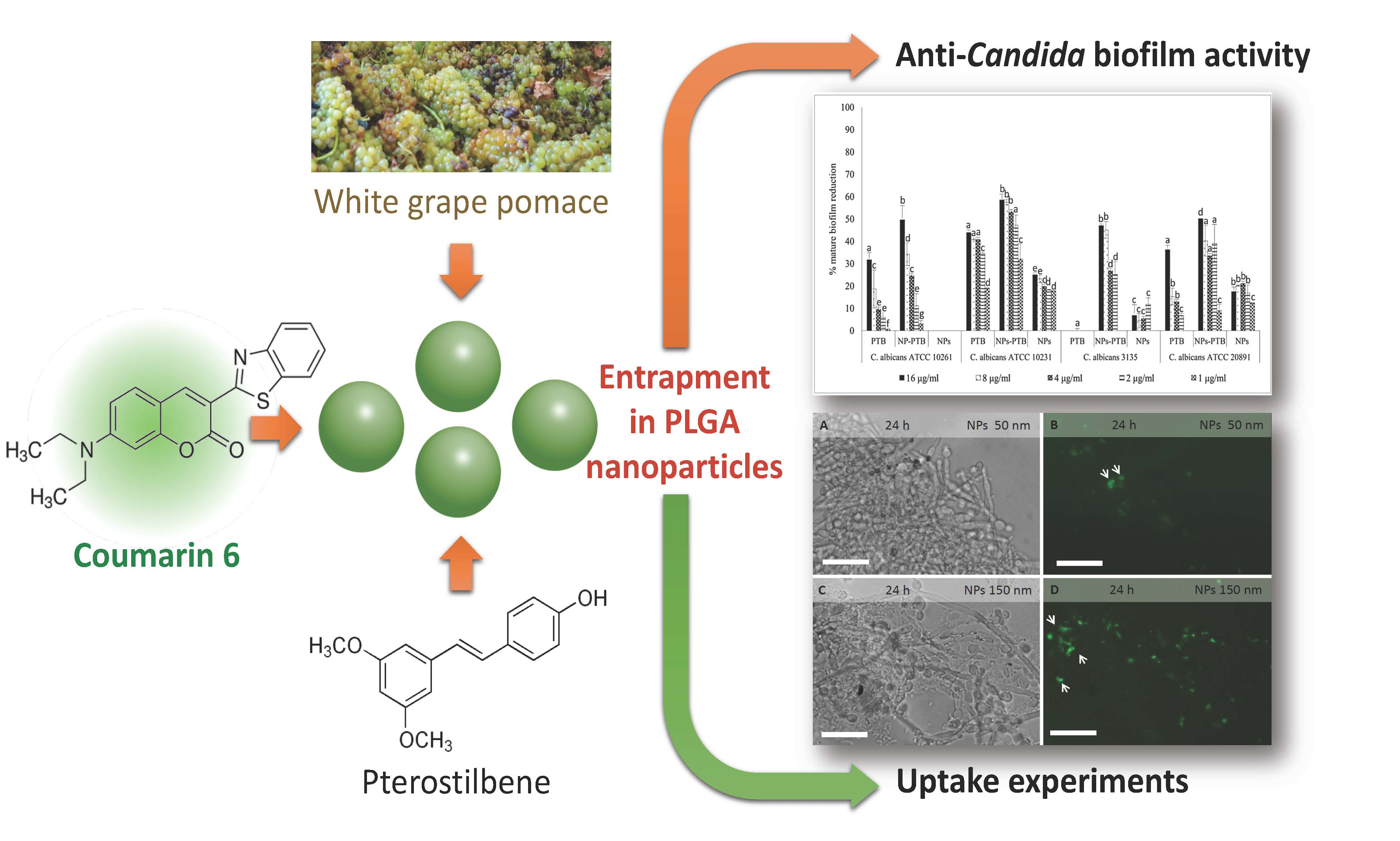
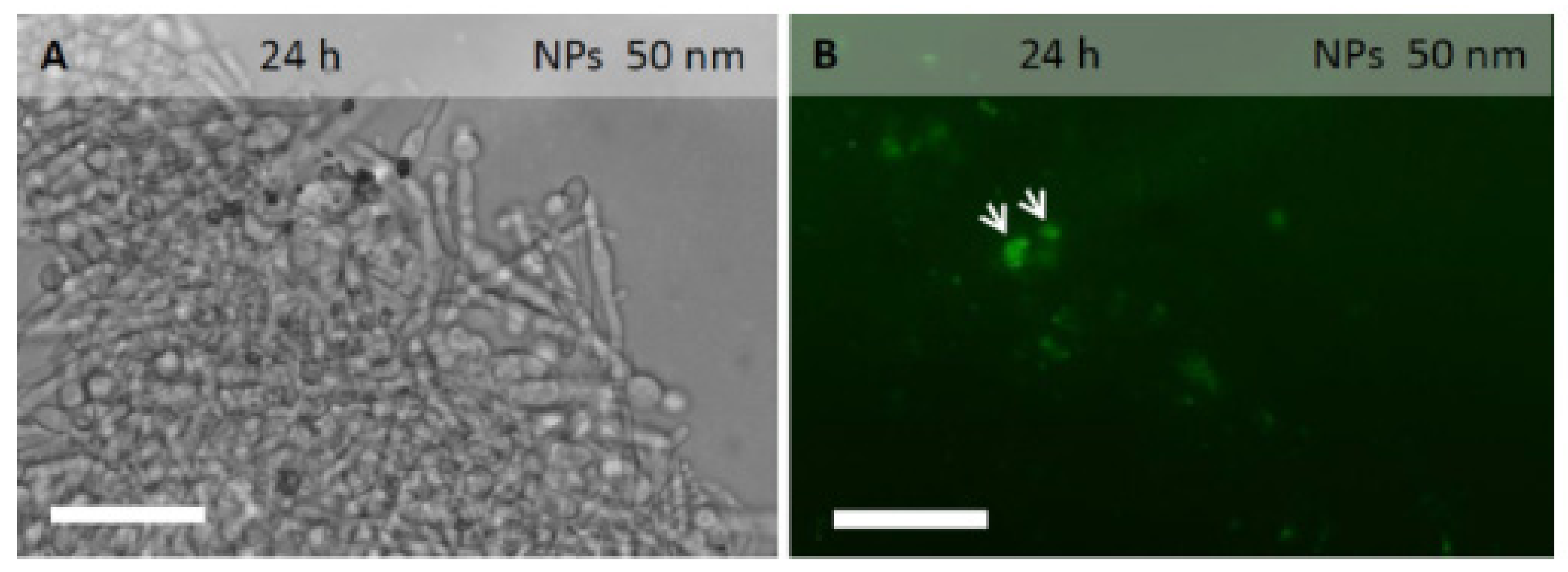
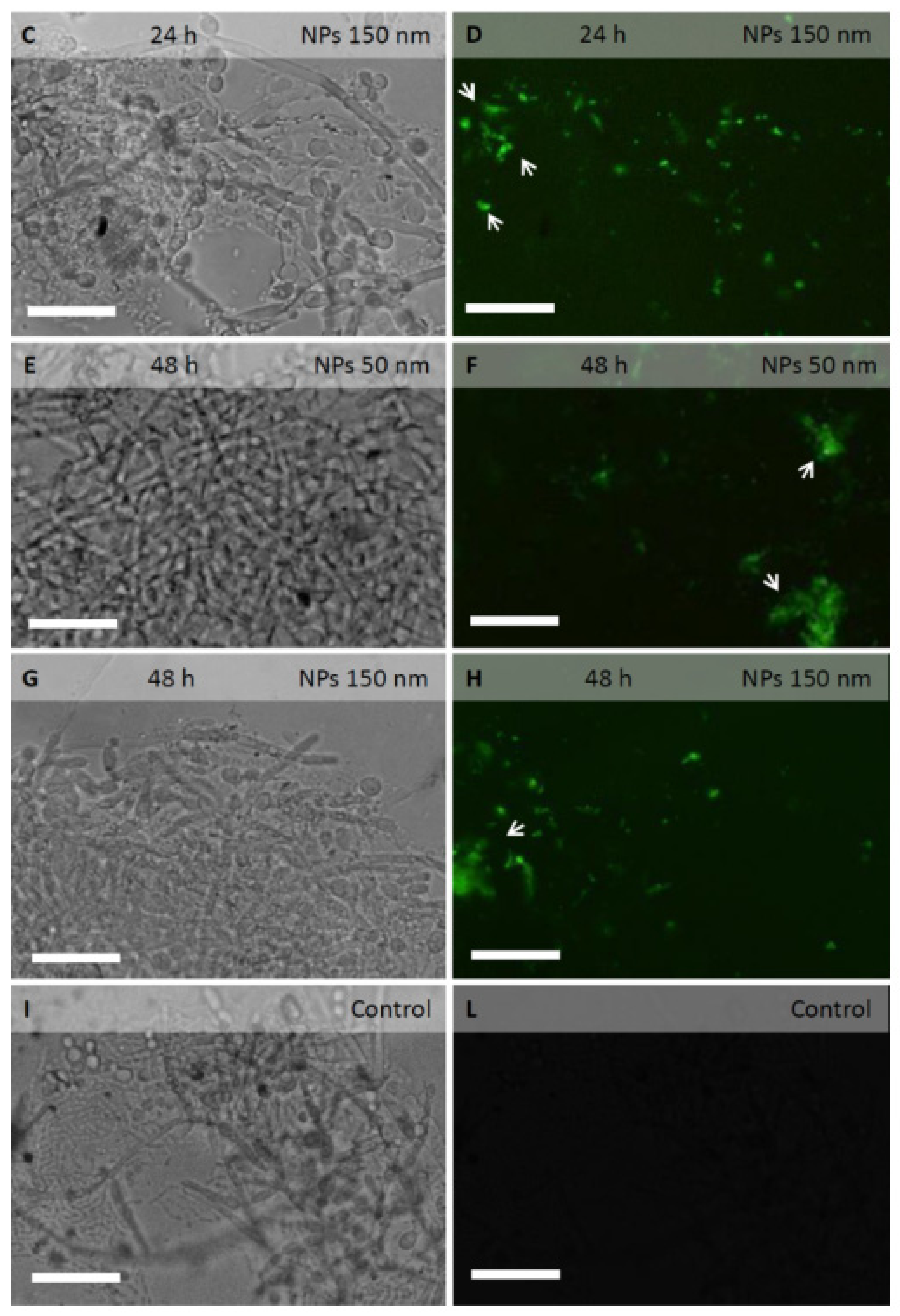
2.3. PLGA NPs Localization in C. albicans Planktonic Cells and Biofilm
2.4. Antifungal Activity of PTB and Pomace Extract-Loaded PLGA NPs on C. albicans Planktonic Cells
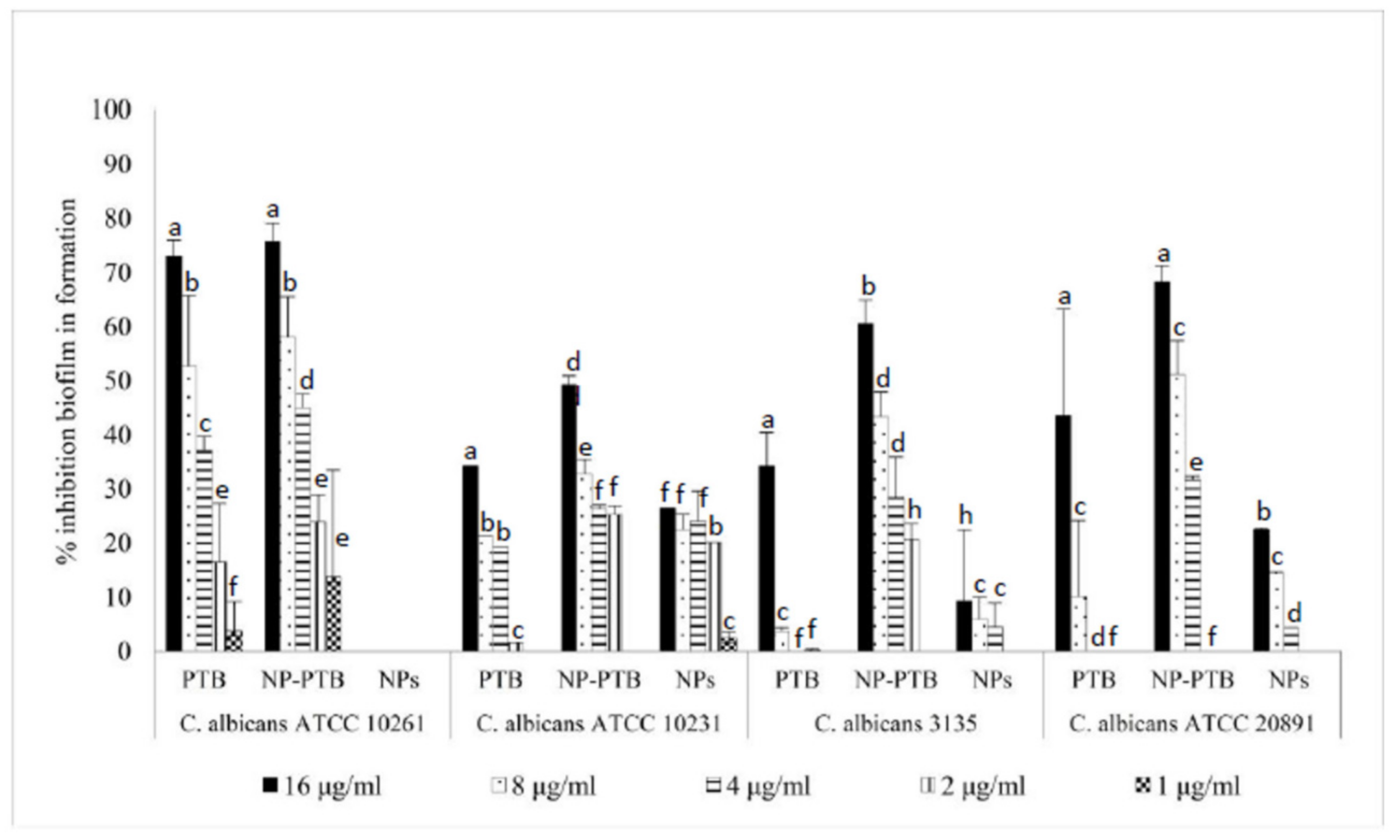
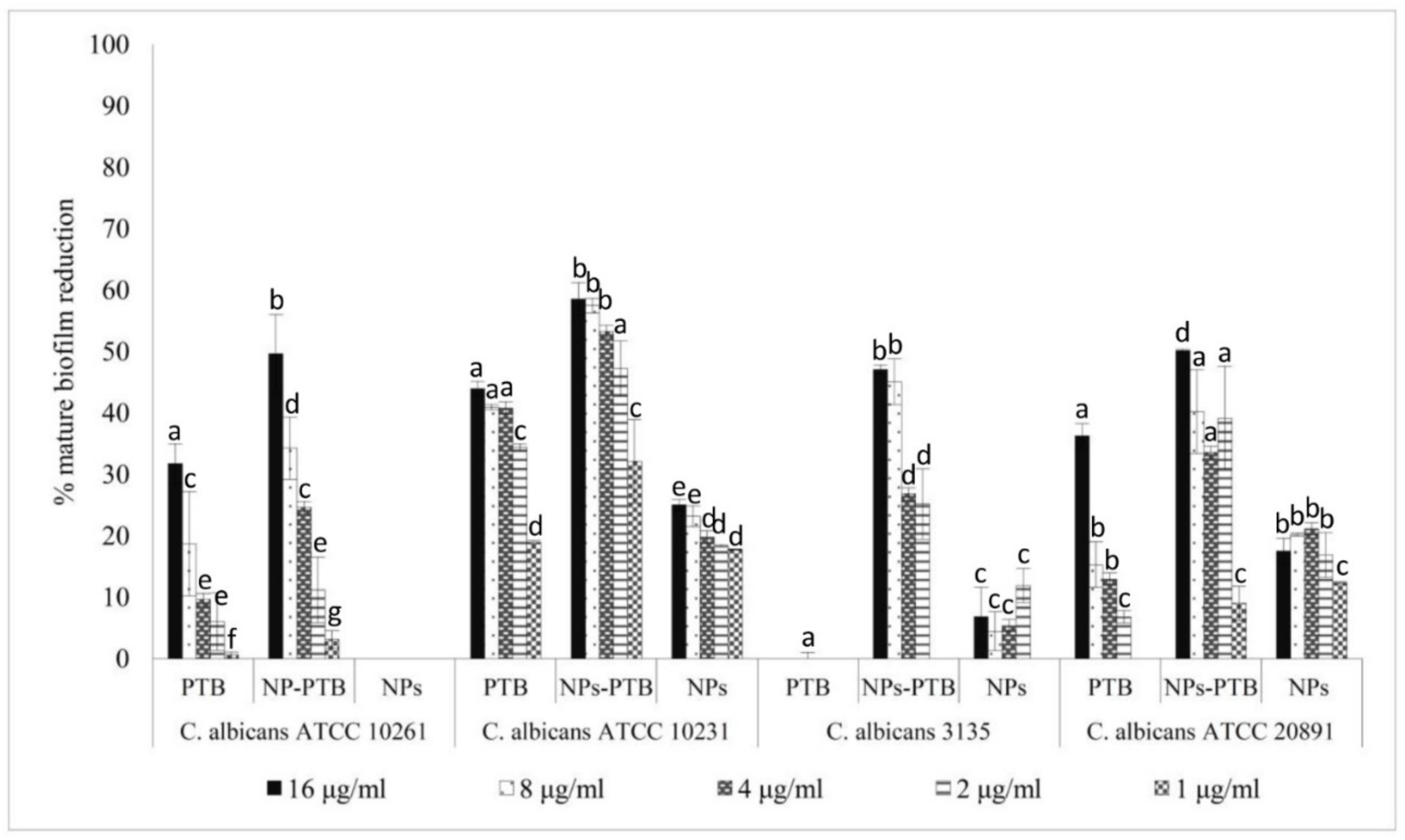
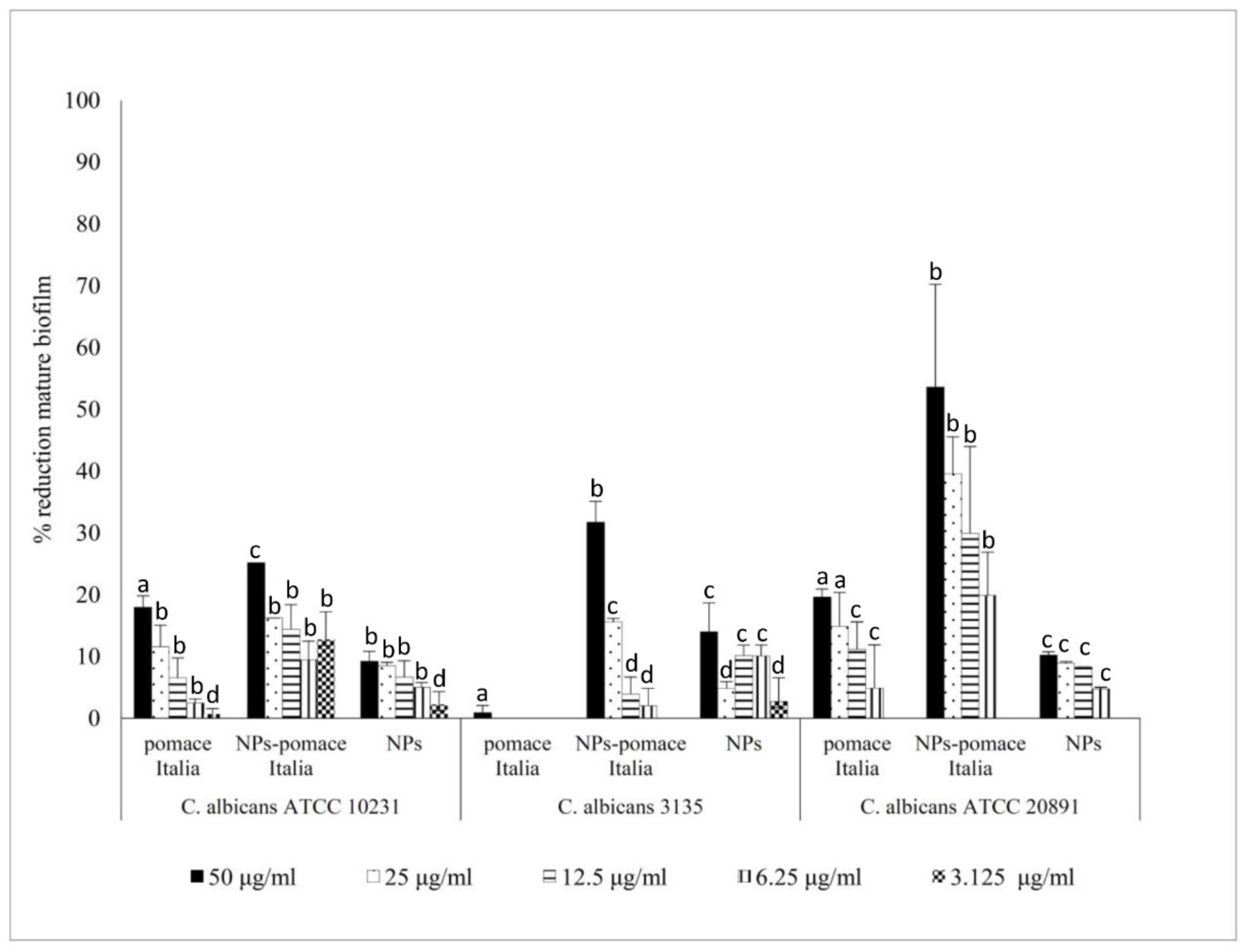
2.5. Antifungal Activity of PTB and Pomace Extract-Loaded PLGA NPs on C. albicans Biofilm
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Pomace Extract Preparation
5.2. NP Preparation and Characterization
5.3. Chemical Characterization of Non-Fermented Grape Pomace
5.4. Determination of Entrapment Efficiency of PTB and Pomace Extract
5.5. Antifungal Susceptibility Testing
5.6. In Vitro Biofilm Formation Assay
5.7. Epifluorescence Microscopy Analysis
5.8. Statistical analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Calderone, R.A.; Fonzi, W.A. Virulence factors of Candida albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Uppuluri, P.; Lopez Ribot, J.L. Candida albicans Biofilms. In Candida albicans: Cellular and Molecular Biology; Prasad, R., Ed.; Springer: Cham, Switzerland, 2017; pp. 63–75. [Google Scholar]
- Nett, J.E. Future directions for anti-biofilm therapeutics targeting Candida. Expert Rev. Anti Infect. Ther. 2014, 12, 375–382. [Google Scholar] [CrossRef]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet. Infect. Dis. 2017, 17, e383–e392. [Google Scholar] [CrossRef]
- Guo, N.; Liu, J.; Wu, X.; Bi, X.; Meng, R.; Wang, X.; Xiang, H.; Deng, X.; Yu, L. Antifungal activity of thymol against clinical isolates of fluconazole-sensitive and-resistant Candida albicans. J. Med. Microbiol. 2009, 58, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- De Vita, D.; Friggeri, L.; D’Auria, F.D.; Pandolfi, F.; Piccoli, F.; Panella, S.; Palamara, A.T.; Simonetti, G.; Scipione, L.; Di Santo, R.; Costi, R.; Tortorella, S. Activity of caffeic acid derivatives against Candida albicans biofilm. Bioorg. Med. Chem. Lett. 2014, 24, 1502–1505. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Lal, P.; Pruthi, V. Prevention of Candida albicans biofilm by plantoils. Mycopathologia 2008, 165, 13–19. [Google Scholar] [CrossRef]
- Evensen, N.A.; Braun, P.C. The effects of tea polyphenols on Candida albicans: Inhibition of biofilm formation and proteasome inactivation. Can. J. Microbiol. 2009, 55, 1033–1039. [Google Scholar] [CrossRef]
- Jung, H.J.; Seu, Y.B.; Lee, D.G. Candicidal action of resveratrol isolated from grapes on human pathogenic yeast C. albicans. J. Microbiol. Biotechnol. 2007, 17, 1324–1329. [Google Scholar] [PubMed]
- Kolouchová, I.; Maťátková, O.; Paldrychová, M.; Kodeš, Z.; Kvasničková, E.; Sigler, K.; Čejková, A.; Šmidrkal, J.; Demnerová, K.; Masák, J. Resveratrol, pterostilbene, and baicalein: Plant-derived anti-biofilm agents. Folia Microbial. 2018, 63, 261–272. [Google Scholar] [CrossRef]
- Simonetti, G.; Santamaria, A.R.; D’Auria, F.D.; Mulinacci, N.; Innocenti, M.; Cecchini, F.; Pericolini, E.; Gabrielli, E.; Panella, S.; Antonacci, D.; et al. Evaluation of anti-Candida activity of Vitis vinifera L. seed extracts obtained from wine and table cultivars. BioMed Res. Int. 2014. Available online: http://dx.doi.org/10.1155/2014/127021 (accessed on 29 May 2019). [CrossRef]
- Pasqua, G.; Simonetti, G.; D’Auria, F.D.; Santamaria, A.R.; Antonacci, D. Estratti Ottenuti da Semi e/o Vinacce di Vitis vinifera e Relative Impieghi Come Agenti Antifungini. Patent application number: 0001407378, 2014. [Google Scholar]
- Pasqua, G.; Simonetti, G. Antimicrobial and antiviral activities of grape seed extracts. In Grape Seeds: Nutrient Content, Antioxidant Properties and Health Benefits; Rodriguez, J.M.L., Ruiz, D.F., Eds.; Nova Science Publishers: New York, NY, USA, 2016; pp. 222–233. [Google Scholar]
- Simonetti, G.; D’Auria, F.D.; Mulinacci, N.; Innocenti, M.; Antonacci, D.; Angiolella, L.; Santamaria, A.R.; Valletta, A.; Donati, L.; Pasqua, G. Anti-dermatophyte and anti-malassezia activity of extracts rich in polymeric flavan-3-ols obtained from Vitis vinifera seeds. Phytother. Res. 2017, 31, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, G.; D’Auria, F.D.; Mulinacci, N.; Milella, R.A.; Antonacci, D.; Innocenti, M.; Pasqua, G. Phenolic content and in vitro antifungal activity of unripe grape extracts from agro-industrial wastes. Nat. Prod. Res. 2017, 33, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.A.; Carson, C.F.; Riley, T.V.; Nielsen, J.B. A review of the toxicity of Melaleuca alternifolia (tea tree) oil. Food. Chem. Toxicol. 2006, 44, 616–625. [Google Scholar] [CrossRef]
- Peixoto, C.M.; Dias, M.I.; Alves, M.J.; Calhelha, R.C.; Barros, L.; Pinho, S.P.; Ferreira, I.C. Grape pomace as a source of phenolic compounds and diverse bioactive properties. Food Chem. 2018, 253, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Li, D.D.; Zhao, L.X.; Mylonakis, E.; Hu, G.H.; Zou, Y.; Huang, T.K.; Yan, L.; Jiang, Y.Y. In vitro and in vivo activities of pterostilbene against Candida albicans biofilms. Antimicrob. Agents Chemother. 2014, 58, 2344–2355. [Google Scholar] [CrossRef]
- Chronopoulou, L.; Di Domenico, E.G.; Ascenzioni, F.; Palocci, C. Positively charged biopolymeric nanoparticles for the inhibition of Pseudomonas aeruginosa biofilms. J. Nanopart. Res. 2016, 18, 308–317. [Google Scholar] [CrossRef]
- Mishra, G.; Mittal, N.; Sharma, A. Multifunctional Mesoporous Carbon Capsules and their Robust Coatings for Encapsulation of Actives: Antimicrobial and Anti-bioadhesion Functions. ACS Appl. Mater. Interfaces. 2017, 9, 19371–19379. [Google Scholar] [CrossRef] [PubMed]
- Soliman, G.M. Nanoparticles as safe and effective delivery systems of antifungal agents: Achievements and challenges. Int. J. Pharm. 2017, 523, 15–32. [Google Scholar] [CrossRef]
- Valletta, A.; Chronopoulou, L.; Palocci, C.; Baldan, B.; Donati, L.; Pasqua, G. Poly(lactic-co-glycolic) acid nanoparticles uptake by Vitis vinifera and grapevine-pathogenic fungi. J. Nanopart. Res. 2014, 16, 2744–2758. [Google Scholar] [CrossRef]
- Palocci, C.; Valletta, A.; Chronopoulou, L.; Donati, L.; Bramosanti, M.; Brasili, E.; Baldan, B.; Pasqua, G. Endocytic pathways involved in PLGA nanoparticle uptake by grapevine cells and role of cell wall and membrane in size selection. Plant Cell. Rep. 2017, 36, 1917–1928. [Google Scholar] [CrossRef]
- Bramosanti, M.; Chronopoulou, L.; Grillo, F.; Valletta, A.; Palocci, C. Microfluidic assisted nanoprecipitation of antiviral-loaded polymeric nanoparticles. Colloids Surf. A Physicochem. Eng. Asp. 2017, 532, 369–376. [Google Scholar] [CrossRef]
- Yang, M.; Du, K.; Hou, Y.; Xie, S.; Dong, Y.; Li, D.; Du, Y. Synergistic Antifungal Effect of Amphotericin B-Loaded Poly (Lactic-Co-Glycolic Acid) Nanoparticles and Ultrasound against Candida albicans Biofilms. Antimicrob. Agents Chemother. 2019, 63, e02022-18. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Saville, S.P.; Thomas, D.P.; Lopez-Ribot, J.L. Candida biofilms: An update. Eukaryotic Cell. 2005, 4, 633–638. [Google Scholar] [CrossRef]
- Lohse, M.B.; Gulati, M.; Johnson, A.D.; Nobile, C.J. Development and regulation of single-and multi-species Candida albicans biofilms. Nat. Rev. Microbiol. 2018, 16, 19. [Google Scholar] [CrossRef]
- Dominguez, E.G.; Andes, D.R. Candida Biofilm Tolerance: Comparison of Planktonic and Biofilm Resistance Mechanisms. In Candida albicans: Cell. Mol. Biol.; Prasad, R., Ed.; Springer: Cham, Switzerland, 2017; pp. 77–92. [Google Scholar]
- Singh, R.; Kumari, A.; Kaur, K.; Sethi, P.; Chakrabarti, A. Relevance of antifungal penetration in biofilm-associated resistance of Candida albicans and non-albicans Candida species. J. Med. Microb. 2018, 67, 922–926. [Google Scholar] [CrossRef]
- Agrawal, S.; Bhatt, M.; Rai, S.K.; Bhatt, A.; Dangwal, P.; Agrawal, P.K. Silver nanoparticles and its potential applications: A review. J. Pharmacogn. Phytochem. 2018, 7, 930–937. [Google Scholar]
- Rai, M.; Yadav, A.; Gade, A. Silver nanoparticles as a new generation of antimicrobials. Biotechnol. Adv. 2009, 27, 76–83. [Google Scholar] [CrossRef]
- Kumari, A.; Yadav, S.K.; Yadav, S.C. Biodegradable polymeric nanoparticles based drug delivery systems. Colloids Surf. B Biointerfaces. 2010, 75, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; Le Breton, A.; Préat, V. PLGA-based nanoparticles: An overview of biomedical applications. J. Control. Release. 2012, 161, 505–522. [Google Scholar] [CrossRef]
- Yano, A.; Kikuchi, S.; Takahashi, T.; Kohama, K.; Yoshida, Y. Inhibitory effects of the phenolic fraction from the pomace of Vitis coignetiae on biofilm formation by Streptococcus mutans. Arch. Oral Biol. 2012, 57, 711–719. [Google Scholar] [CrossRef]
- Kammerer, D.; Claus, A.; Carle, R.; Schieber, A. Polyphenol screening of pomace from red and white grape varieties (Vitis vinifera L.) by HPLC-DAD-MS/MS. J. Agric. Food Chem. 2004, 52, 4360–4367. [Google Scholar] [CrossRef] [PubMed]
- Chronopoulou, L.; Sparago, C.; Palocci, C. A modular microfluidic platform for thesynthesis of biopolymeric nanoparticles entrapping organic actives. J. Nanopart. Res. 2014, 16, 2703–2713. [Google Scholar] [CrossRef]
- Provencher, S. Contin. A general purpose constrained regularization program for inverting noisy linear algebraic and integral equations. Comput. Phys. Commun. 1982, 27, 229–242. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V.L. Total phenol analysis: Automation and comparison withmanual methods. Am. J. En. Vitic. 1977, 28, 49–55. [Google Scholar]
- CLSI Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; approved standard-third edition; CLSI document M27-A3; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008.
- CLSI Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts; 4th informational supplement. CLSI document M27-S4; Clinical and Laboratory Standards Institute: Wayne, PA, USA.
- Pierce, C.G.; Uppuluri, P.; Tristan, A.R.; Wormley, F.L., Jr.; Mowat, E.; Ramage, G.; Lopez Ribot, J.L. A simple and reproducible 96-well plate-based method for the formation of fungal biofilms and its application to antifungal susceptibility testing. Nat. Protoc. 2008, 3, 1494–1500. [Google Scholar] [CrossRef] [PubMed]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PLGA NPs | NPs Size (nm) ± SD | PI ± SD |
|---|---|---|
| Empty PLGA NPs | 50 ± 12 | 0.3 ± 0.03 |
| Coumarin 6-loaded PLGA NPs | 50 ± 14 | 0.4 ± 0.04 |
| PTB-loaded PLGA NPs | 50 ± 10 | 0.2 ± 0.02 |
| Pomace crude extract-loaded PLGA NPs | 50 ± 13 | 0.3 ± 0.03 |
| Coumarin 6-loaded PLGA NPs | 150 ± 14 | 0.2 ± 0.02 |
| m/z | Retention Time (min) | Metabolite | Fragments |
|---|---|---|---|
| 178.9 | 2.1–3.3 | Caffeic acid | 58.7/70.6/89/85.1 |
| 289.1 | 30.5–30.9 | Catechin/Epicatechin | 109/124.8/203.1/151.1/245.2/123/161.1205.1/137/221.2/289/179/108.9/124.8/203.1/122.8/150.9/204.7/245.3/289 |
| 168.8 | 11–12.3 22.43 | Gallic acid | 125/168.6/79.1/80/96.8/107.1/125/169.1/78.8/ 97.2/106.9 |
| 577.1 | 28.6–29.8/32–33.2/33.4–33.8/34.7–38.4/42.8–43 | Procyanidin | 407.3/289.1/425.2/125/450.8/577 |
| 191 | 2.5–5.6 | Quinic acid | 110.8/72.7/85.1/58.8/98.9/117/111/86.8/85 |
| 195 | 1.3–2.5 5.6 | Unknown | 74.7/98.8/86.9/85.1/58.7/70.7/89/128.9/194.8 |
| 224.9 | 2.1–2.8/15.7 | Unknown | 89.1/58.7/70.6/112.8/100.9/85.1/118.8 |
| 161.1 | 2.2–2.5/19.5–20.1 | Unknown | 70.6/85/72.6/58.8/94.9/100.8/56.7/83.1/113.1 |
| 253.1 | 2.2–4.2 | Unknown | 88.8/72.7/70.6/87.2/113/119.2/100.7 |
| 439.1 | 2.4 | Unknown | 97.1/78.9/161.1/178.6 |
| 331 | 21.8–22.6 | Unknown | 169.1/125/331/168.1/124.9/313.1 |
| 293.1 | 269–27.1/27.7–27.8 | Unknown | 130.9/89/118.6/293.1/100.9/112.8/85.1 |
| 579.2 | 29-29.7/32.2–33.1/33.4–33.8/34.8-35.1 | Unknown | 289.1/245.2/579.2/ 426.8/409.4/453.2/288.8 |
| 441.1 | 42.2–42.5 | Unknown | 169/389.1/125/271.2/441.2 |
| 477 | 43.7–44 | Unknown | 301.2/476.8/113.1/175 |
| 463 | 43-43.1/43.3–43.6 | Unknown | 463.2/301.2/178.9/343.1/150.6 463.3/300.2/300.8/178.9/343 |
| 493.3 | 49.7/50.1–50.2/50.3-50.6 | Unknown | 315.2/447.4/161.1/131/493.3/447/160.7/314.9/447.4/315.1/130.6/161.2/118.9/179/191.2/447.3/149.2/179.1/131/88.9/118.9/161/191.2/315/251.3/221.1/47.4/149/493.3/179.1/101.2/160.9/89/118.8/130.6/190.7/251.1/221.1 |
| 507.2 | 50.3–50.9 | Unknown | 149.1/292.9/88.9/167.2/233.1/190.6/124.6/100.5/221.1/507.2 |
| MIC 50 (µg/mL) | |||||
|---|---|---|---|---|---|
| C. albicans | PTB | PTB + NPs | Pomace Extract | Pomace + NPs | NPs |
| 10,231 | >16 | >16 | 12.5 ± 0.0 | 50 ± 0 | >50 |
| 3135 | >16 | >16 | 6.25 ± 0.0 | 50 ± 0 | >50 |
| 20,891 | >16 | >16 | 1.4 ± 0.4 | 12.5 ± 0.0 | >50 |
| 10,261 | >16 | >16 | 12.5 ± 0.0 | 50 ± 0 | >50 |
| GM | >16 | >16 | 6.06 | 35.35 | >50 |
| Median | >16 | >16 | 9.45 | 50 | >50 |
| MIC 90 (µg/mL) | |||||
| C.albicans | PTB | PTB + NPs | Pomace Extract | Pomace + NPs | NPs |
| 10,231 | >16 | >16 | 25 ± 0 | 50 ± 0 | >50 |
| 3135 | >16 | >16 | 25 ± 0 | 50 ± 0 | >50 |
| 20,891 | >16 | >16 | 7.49 ± 3.05 | 50 ± 0 | >50 |
| 10,261 | >16 | >16 | 21.87 ± 6.25 | 50 ± 0 | >50 |
| GM | >16 | >16 | 17.76 | 50 | >50 |
| Median | >16 | >16 | 25 | 50 | >50 |
| MIC 100 (µg/mL) | |||||
| C.albicans | PTB | PTB + NPs | Pomace extract | Pomace + NPs | NPs |
| 10,231 | >16 | >16 | >50 | >50 | >50 |
| 3135 | >16 | >16 | >50 | >50 | >50 |
| 20,891 | >16 | >16 | >50 | >50 | >50 |
| 10,261 | >16 | >16 | >50 | >50 | >50 |
| GM | >16 | >16 | >50 | >50 | >50 |
| Median | >16 | >16 | >50 | >50 | >50 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simonetti, G.; Palocci, C.; Valletta, A.; Kolesova, O.; Chronopoulou, L.; Donati, L.; Di Nitto, A.; Brasili, E.; Tomai, P.; Gentili, A.; et al. Anti-Candida Biofilm Activity of Pterostilbene or Crude Extract from Non-Fermented Grape Pomace Entrapped in Biopolymeric Nanoparticles. Molecules 2019, 24, 2070. https://doi.org/10.3390/molecules24112070
Simonetti G, Palocci C, Valletta A, Kolesova O, Chronopoulou L, Donati L, Di Nitto A, Brasili E, Tomai P, Gentili A, et al. Anti-Candida Biofilm Activity of Pterostilbene or Crude Extract from Non-Fermented Grape Pomace Entrapped in Biopolymeric Nanoparticles. Molecules. 2019; 24(11):2070. https://doi.org/10.3390/molecules24112070
Chicago/Turabian StyleSimonetti, Giovanna, Cleofe Palocci, Alessio Valletta, Olga Kolesova, Laura Chronopoulou, Livia Donati, Antonio Di Nitto, Elisa Brasili, Pierpaolo Tomai, Alessandra Gentili, and et al. 2019. "Anti-Candida Biofilm Activity of Pterostilbene or Crude Extract from Non-Fermented Grape Pomace Entrapped in Biopolymeric Nanoparticles" Molecules 24, no. 11: 2070. https://doi.org/10.3390/molecules24112070
APA StyleSimonetti, G., Palocci, C., Valletta, A., Kolesova, O., Chronopoulou, L., Donati, L., Di Nitto, A., Brasili, E., Tomai, P., Gentili, A., & Pasqua, G. (2019). Anti-Candida Biofilm Activity of Pterostilbene or Crude Extract from Non-Fermented Grape Pomace Entrapped in Biopolymeric Nanoparticles. Molecules, 24(11), 2070. https://doi.org/10.3390/molecules24112070